Mast Cells and Angiogenesis in Human Plasma Cell Malignancies



Abstract
1. Mast Cells in Human Plasma Cell Malignancies
2. Mast Cells and Tumor Growth
3. Mast Cells and Tumor Angiogenesis
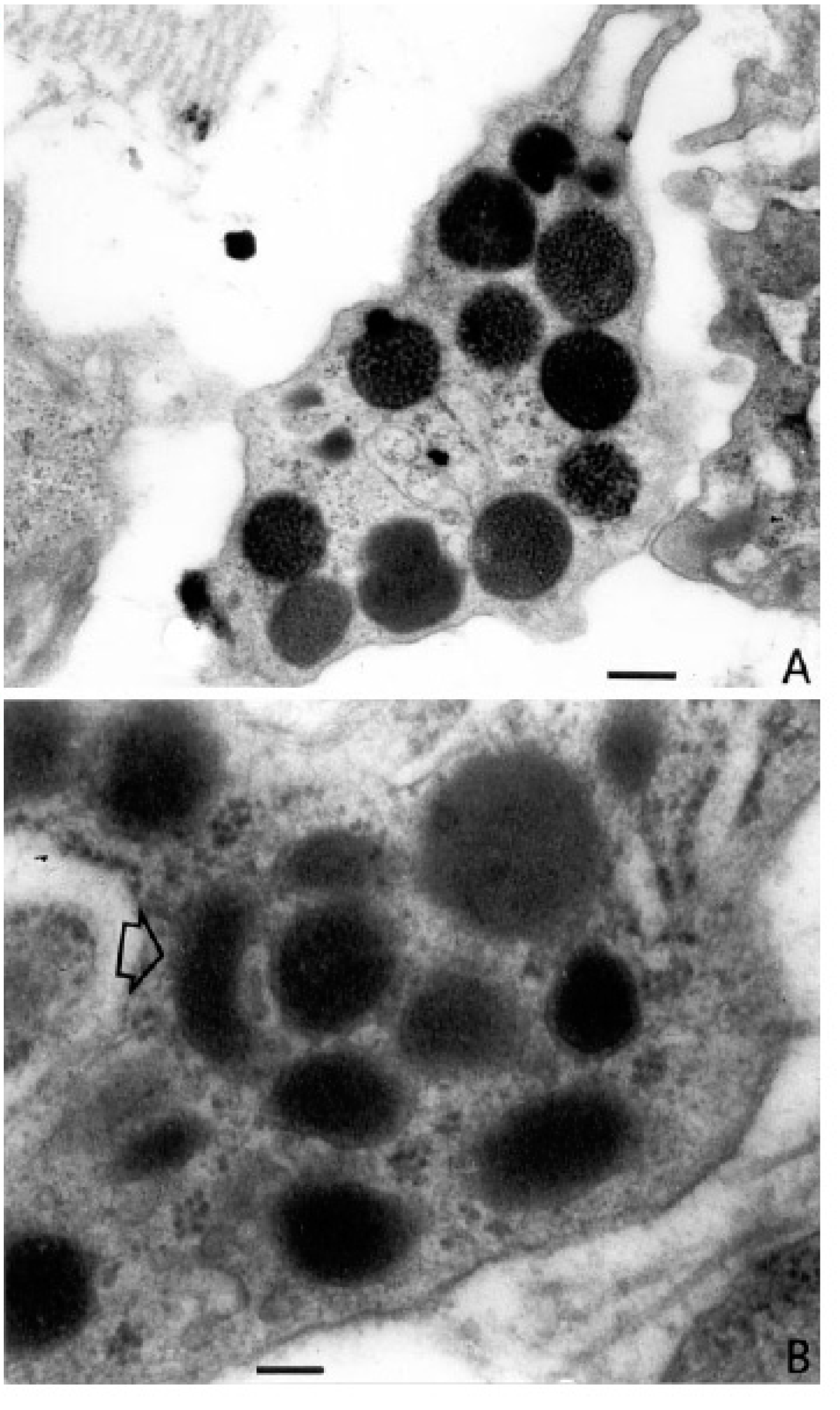
4. The Role of Mast Cells in Angiogenesis in Plasmocytoma
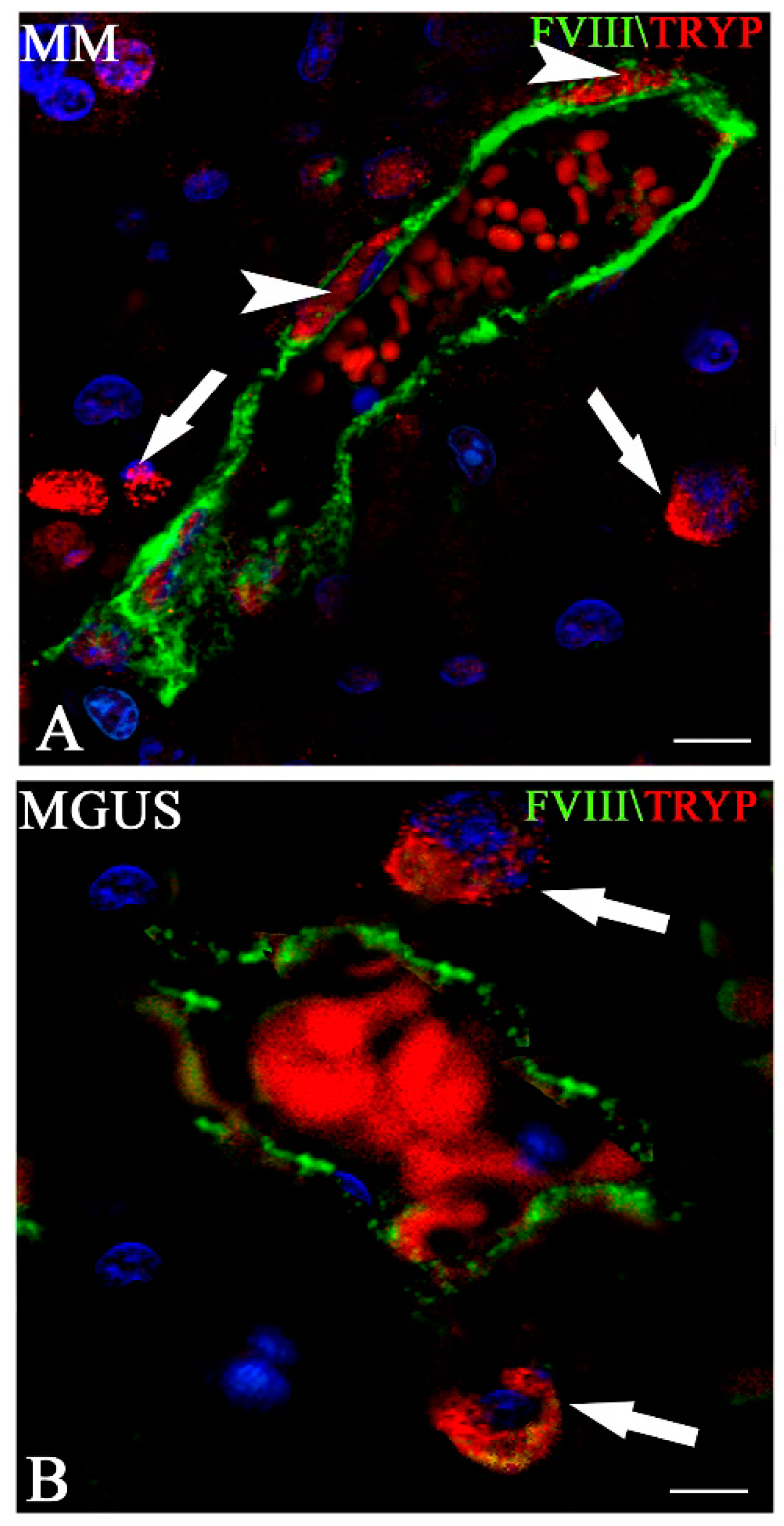
5. The Role of Mast Cells in Angiogenesis in Multiple Myeloma
6. Concluding Remarks and Therapeutic Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Corwin, J.; Lindberg, R.D. Solitary plasmacytoma of bone vs. extramedullary plasmacytoma and their relationship to multiple myeloma. Cancer 1979, 43, 1007–1013. [Google Scholar] [CrossRef]
- Woodruff, R.K.; Whittle, J.M.; Malpas, J.S. Solitary plasmacytoma. I: Extramedullary soft tissue plasmacytoma. Cancer 1979, 43, 2340–2343. [Google Scholar] [CrossRef]
- Bindal, A.K.; Bindal, R.K.; van Loveren, H.; Sawaya, R. Management of intracranial plasmacytoma. J. Neurosurg. 1995, 83, 218–221. [Google Scholar] [CrossRef] [PubMed]
- Klein, B.; Zhang, X.G.; Jourdan, M.; Content, J.; Houssiau, F.; Aarden, L.; Piechaczyk, M.; Bataille, R. Paracrine rather than autocrine regulation of myeloma-cell growth and differentiation by interleukin-6. Blood 1989, 73, 517–526. [Google Scholar] [PubMed]
- Lemoli, R.M.; Fortuna, A.; Grande, A.; Gamberi, B.; Bonsi, L.; Fogli, M.; Amabile, M.; Cavo, M.; Ferrari, S.; Tura, S. Expression and functional role of c-kit ligand (SCF) in human multiple myeloma cells. Br. J. Haematol. 1994, 88, 760–769. [Google Scholar] [CrossRef]
- Ribatti, D.; Crivellato, E. Chapter 4 The Controversial Role of Mast Cells in Tumor Growth. In International Review of Cell and Molecular Biology; Elsevier: Amsterdam, The Netherlands, 2009; pp. 89–131. [Google Scholar]
- Fitzsimons, C.; Molinari, B.; Duran, H.; Palmieri, M.; Davio, C.; Cricco, G.; Bergoc, R.; Rivera, E. Atypical association of H 1 and H 2 histamine receptors with signal transduction pathways during multistage mouse skin carcinogenesis. Inflamm. Res. 1997, 46, 292–298. [Google Scholar] [CrossRef]
- Jeong, H.J.; Oh, H.A.; Nam, S.Y.; Han, N.R.; Kim, Y.S.; Kim, J.H.; Lee, S.J.; Kim, M.H.; Moon, P.D.; Kim, H.M.; et al. The critical role of mast cell-derived hypoxia-inducible factor-1α in human and mice melanoma growth. Int. J. Cancer 2013, 132, 2492–2501. [Google Scholar] [CrossRef]
- Grimbaldeston, M.A.; Nakae, S.; Kalesnikoff, J.; Tsai, M.; Galli, S.J. Mast cell–derived interleukin 10 limits skin pathology in contact dermatitis and chronic irradiation with ultraviolet B. Nat. Immunol. 2007, 8, 1095–1104. [Google Scholar] [CrossRef]
- Ullrich, S.E.; Nghiem, D.X.; Khaskina, P. Suppression of an Established Immune Response by UVA? A Critical Role for Mast Cells. Photochem. Photobiol. 2007, 83, 1095–1100. [Google Scholar] [CrossRef]
- Ribatti, D.; Crivellato, E. Mast cells, angiogenesis, and tumour growth. Biochim. Biophys. Acta 2012, 1822, 2–8. [Google Scholar] [CrossRef]
- Abdel-Majid, R.M.; Marshall, J.S. Prostaglandin E2 Induces Degranulation-Independent Production of Vascular Endothelial Growth Factor by Human Mast Cells. J. Immunol. 2004, 172, 1227–1236. [Google Scholar] [CrossRef]
- Boesiger, J.; Tsai, M.; Maurer, M.; Yamaguchi, M.; Brown, L.F.; Claffey, K.P.; Dvorak, H.F.; Galli, S.J. Mast Cells Can Secrete Vascular Permeability Factor/ Vascular Endothelial Cell Growth Factor and Exhibit Enhanced Release after Immunoglobulin E–dependent Upregulation of Fcε Receptor I Expression. J. Exp. Med. 1998, 188, 1135–1145. [Google Scholar] [CrossRef] [PubMed]
- Grützkau, A.; Krüger-Krasagakes, S.; Baumeister, H.; Schwarz, C.; Kögel, H.; Welker, P.; Lippert, U.; Henz, B.M.; Möller, A. Synthesis, Storage, and Release of Vascular Endothelial Growth Factor/Vascular Permeability Factor (VEGF/VPF) by Human Mast Cells: Implications for the Biological Significance of VEGF206. Mol. Biol. Cell 1998, 9, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Kanbe, N.; Kurosawa, M.; Nagata, H.; Yamashita, T.; Kurimoto, F.; Miyachi, Y. Production of fibrogenic cytokines by cord blood–derived cultured human mast cells. J. Allergy Clin. Immunol. 2000, 106, S85–S90. [Google Scholar] [CrossRef]
- Nilsson, G.; Forsberg-Nilsson, K.; Xiang, Z.; Hallböök, F.; Nilsson, K.; Metcalfe, D.D. Human mast cells express functional TrkA and are a source of nerve growth factor. Eur. J. Immunol. 1997, 27, 2295–2301. [Google Scholar] [CrossRef] [PubMed]
- Qu, Z.; Huang, X.; Ahmadi, P.; Stenberg, P.; Liebler, J.M.; Le, A.-C.; Planck, S.R.; Rosenbaum, J.T. Synthesis of Basic Fibroblast Growth Factor by Murine Mast CellsRegulation by Transforming Growth Factor β, Tumor Necrosis Factor α, and Stem Cell Factor. Int. Arch. Allergy Immunol. 1997, 115, 47–54. [Google Scholar] [CrossRef]
- Qu, Z.; Kayton, R.J.; Ahmadi, P.; Liebler, J.M.; Powers, M.R.; Planck, S.R.; Rosenbaum, J.T. Ultrastructural Immunolocalization of Basic Fibroblast Growth Factor in Mast Cell Secretory Granules: Morphological Evidence for bFGF Release Through Degranulation. J. Histochem. Cytochem. 1998, 46, 1119–1128. [Google Scholar] [CrossRef] [PubMed]
- Qu, Z.; Liebler, J.M.; Powers, M.R.; Galey, T.; Ahmadi, P.; Huang, X.N.; Ansel, J.C.; Butterfield, J.H.; Planck, S.R.; Rosenbaum, J.T. Mast cells are a major source of basic fibroblast growth factor in chronic inflammation and cutaneous hemangioma. Am. J. Pathol. 1995, 147, 564–573. [Google Scholar]
- Walsh, L.J.; Trinchieri, G.; Waldorf, H.A.; Whitaker, D.; Murphy, G.F. Human dermal mast cells contain and release tumor necrosis factor α, which induces endothelial leukocyte adhesion molecule 1. Proc. Natl. Acad. Sci. USA 1991, 88, 4220–4224. [Google Scholar] [CrossRef]
- Moller, A.; Henz, B.M.; Grutzkau, A.; Lippert, U.; Aragane, Y.; Schwarz, T.; Kruger-Krasagakes, S. Comparative cytokine gene expression: Regulation and release by human mast cells. Immunology 1998, 93, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Detmar, M.; Brown, L.F.; Schön, M.P.; Elicker, B.M.; Velasco, P.; Richard, L.; Fukumura, D.; Monsky, W.; Claffey, K.P.; Jain, R.K. Increased Microvascular Density and Enhanced Leukocyte Rolling and Adhesion in the Skin of VEGF Transgenic Mice. J. Investig. Dermatol. 1998, 111, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Detoraki, A.; Staiano, R.I.; Granata, F.; Giannattasio, G.; Prevete, N.; de Paulis, A.; Ribatti, D.; Genovese, A.; Triggiani, M.; Marone, G. Vascular endothelial growth factors synthesized by human lung mast cells exert angiogenic effects. J. Allergy Clin. Immunol. 2009, 123, 1142–1149. [Google Scholar] [CrossRef]
- Gruber, B.L.; Marchese, M.J.; Kiely, J.; Schwartz, L.B.; Schecter, N.M. 520 Human mast cell products degrade connective tissue matrix. J. Allergy Clin. Immunol. 1988, 81, 298. [Google Scholar] [CrossRef]
- Ribatti, D.; Crivellato, E.; Candussio, L.; Nico, B.; Vacca, A.; Roncali, L.; Dammacco, F. Mast cells and their secretory granules are angiogenic in the chick embryo chorioallantoic membrane. Clin. Exp. Allergy 2001, 31, 602–608. [Google Scholar] [CrossRef] [PubMed]
- Norrby, K.; Jakobsson, A.; Sörbo, J. Mast-cell-mediated angiogenesis: A novel experimental model using the rat mesentery. Virchows Archiv B 1986, 52, 195–206. [Google Scholar] [CrossRef]
- Norrby, K.; Jakobsson, A.; Sörbo, J. Mast-cell secretion and angiogenesis, a quantitative study in rats and mice. Virchows Archiv B 1989, 57, 251–256. [Google Scholar] [CrossRef]
- Ribatti, D.; Roncali, L.; Nico, B.; Bertossi, M. Effects of Exogenous Heparin on the Vasculogenesis of the Chorioallantoic Membrane. Cells Tissues Organs 1987, 130, 257–263. [Google Scholar] [CrossRef]
- Sörbo, J.; Norrby, K. Mast-cell histamine expands the microvasculature spatially. Agents Actions 1992, 36, C387–C389. [Google Scholar] [CrossRef]
- Metcalfe, D.D.; Baram, D.; Mekori, Y.A. Mast cells. Physiol. Rev. 1997, 77, 1033–1079. [Google Scholar] [CrossRef]
- Blair, R.J.; Meng, H.; Marchese, M.J.; Ren, S.; Schwartz, L.B.; Tonnesen, M.G.; Gruber, B.L. Human mast cells stimulate vascular tube formation. Tryptase is a novel, potent angiogenic factor. J. Clin. Investig. 1997, 99, 2691–2700. [Google Scholar] [CrossRef]
- De Souza, D.A.; Toso, V.D.; Campos, M.R.D.C.; Lara, V.S.; Oliver, C.; Jamur, M.C. Expression of Mast Cell Proteases Correlates with Mast Cell Maturation and Angiogenesis during Tumor Progression. PLoS ONE 2012, 7, e40790. [Google Scholar] [CrossRef]
- Koskivirta, I.; Rahkonen, O.; Mäyränpää, M.; Pakkanen, S.; Husheem, M.; Sainio, A.; Hakovirta, H.; Laine, J.; Jokinen, E.; Vuorio, E.; et al. Tissue inhibitor of metalloproteinases 4 (TIMP4) is involved in inflammatory processes of human cardiovascular pathology. Histochem. Cell Biol. 2006, 126, 335–342. [Google Scholar] [CrossRef]
- Tanaka, A.; Yamane, Y.; Matsuda, H. Mast Cell MMP-9 Production Enhanced by Bacterial Lipopolysaccharide. J. Vet. Med. Sci. 2001, 63, 811–813. [Google Scholar] [CrossRef] [PubMed]
- Starkey, J.R.; Crowle, P.K.; Taubenberger, S. Mast-cell-deficient W/Wv mice exhibit A decreased rate of tumor angiogenesis. Int. J. Cancer 1988, 42, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Coussens, L.M.; Raymond, W.W.; Bergers, G.; Laig-Webster, M.; Behrendtsen, O.; Werb, Z.; Caughey, G.H.; Hanahan, D. Inflammatory mast cells up-regulate angiogenesis during squamous epithelial carcinogenesis. Genes Dev. 1999, 13, 1382–1397. [Google Scholar] [CrossRef]
- Coussens, L.M.; Tinkle, C.L.; Hanahan, D.; Werb, Z. MMP-9 Supplied by Bone Marrow–Derived Cells Contributes to Skin Carcinogenesis. Cell 2000, 103, 481–490. [Google Scholar] [CrossRef]
- Ribatti, D.; Ennas, M.G.; Vacca, A.; Ferreli, F.; Nico, B.; Orru, S.; Sirigu, P. Tumor vascularity and tryptase-positive mast cells correlate with a poor prognosis in melanoma. Eur. J. Clin. Investig. 2003, 33, 420–425. [Google Scholar] [CrossRef]
- Ribatti, D.; Molica, S.; Vacca, A.; Nico, B.; Crivellato, E.; Roccaro, A.M.; Dammacco, F. Tryptase-positive mast cells correlate positively with bone marrow angiogenesis in B-cell chronic lymphocytic leukemia. Leukemia 2003, 17, 1428–1430. [Google Scholar] [CrossRef]
- Tth, T. Cutaneous malignant melanoma: Correlation between neovascularization and peritumor accumulation of mast cells overexpressing vascular endothelial growth factor. Hum. Pathol. 2000, 31, 955–960. [Google Scholar]
- Glowacki, J.; Mulliken, J.B. Mast cells in hemangiomas and vascular malformations. Pediatrics 1982, 70, 48–51. [Google Scholar]
- Fukushima, N.; Satoh, T.; Sano, M.; Tokunaga, O. Angiogenesis and Mast Cells in Non-Hodgkin’s Lymphoma: A Strong Correlation in Angioimmunoblastic T-Cell Lymphoma. Leuk. Lymphoma 2001, 42, 709–720. [Google Scholar] [CrossRef] [PubMed]
- Marinaccio, C.; Ingravallo, G.; Gaudio, F.; Perrone, T.; Nico, B.; Maoirano, E.; Specchia, G.; Ribatti, D. Microvascular density, CD68 and tryptase expression in human Diffuse Large B-Cell Lymphoma. Leuk. Res. 2014, 38, 1374–1377. [Google Scholar] [CrossRef] [PubMed]
- Rabenhorst, A.; Schlaak, M.; Heukamp, L.C.; Forster, A.; Theurich, S.; von Bergwelt-Baildon, M.; Buttner, R.; Kurschat, P.; Mauch, C.; Roers, A.; et al. Mast cells play a protumorigenic role in primary cutaneous lymphoma. Blood 2012, 120, 2042–2054. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D.; Nico, B.; Vacca, A.; Marzullo, A.; Calvi, N.; Roncali, L.; Dammacco, F. Do mast cells help to induce angiogenesis in B-cell non-Hodgkin’s lymphomas? Br. J. Cancer 1998, 77, 1900–1906. [Google Scholar] [CrossRef]
- Ribatti, D.; Vacca, A.; Nico, B.; Quondamatteo, F.; Ria, R.; Minischetti, M.; Marzullo, A.; Herken, R.; Roncali, L.; Dammacco, F. Bone marrow angiogenesis and mast cell density increase simultaneously with progression of human multiple myeloma. Br. J. Cancer 1999, 79, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D.; Polimeno, G.; Vacca, A.; Marzullo, A.; Crivellato, E.; Nico, B.; Lucarelli, G.; Dammacco, F. Correlation of bone marrow angiogenesis and mast cells with tryptase activity in myelodysplastic syndromes. Leukemia 2002, 16, 1680–1684. [Google Scholar] [CrossRef] [PubMed]
- Molica, S.; Vacca, A.; Crivellato, E.; Cuneo, A.; Ribatti, D. Tryptase-positive mast cells predict clinical outcome of patients with early B-cell chronic lymphocytic leukemia. Eur. J. Haematol. 2003, 71, 137–139. [Google Scholar] [CrossRef]
- Bowrey, P.F.; King, J.; Magarey, C.; Schwartz, P.; Marr, P.; Bolton, E.; Morris, D.L. Histamine, mast cells and tumour cell proliferation in breast cancer: Does preoperative cimetidine administration have an effect? Br. J. Cancer 2000, 82, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Hartveit, F. Mast cells and metachromasia in human breast cancer: Their occurrence, significance and consequence: A preliminary report. J. Pathol. 1981, 134, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Marech, I.; Ammendola, M.; Leporini, C.; Patruno, R.; Luposella, M.; Zizzo, N.; Passantino, G.; Sacco, R.; Farooqi, A.A.; Zuccalà, V.; et al. C-Kit receptor and tryptase expressing mast cells correlate with angiogenesis in breast cancer patients. Oncotarget 2017, 9, 7918. [Google Scholar] [CrossRef]
- Ribatti, D.; Finato, N.; Crivellato, E.; Guidolin, D.; Longo, V.; Mangieri, D.; Nico, B.; Vacca, A.; Beltrami, C.A. Angiogenesis and mast cells in human breast cancer sentinel lymph nodes with and without micrometastases. Histopathology 2007, 51, 837–842. [Google Scholar] [CrossRef]
- Ranieri, G.; Ammendola, M.; Patruno, R.; Celano, G.; Zito, F.A.; Montemurro, S.; Rella, A.; Di Lecce, V.; Gadaleta, C.D.; Battista De Sarro, G.; et al. Tryptase-positive mast cells correlate with angiogenesis in early breast cancer patients. Int. J. Oncol. 2009, 35, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Ammendola, M.; Sacco, R.; Sammarco, G.; Donato, G.; Zuccalà, V.; Romano, R.; Luposella, M.; Patruno, R.; Vallicelli, C.; Verdecchia, G.M.; et al. Mast Cells Positive to Tryptase and c-Kit Receptor Expressing Cells Correlates with Angiogenesis in Gastric Cancer Patients Surgically Treated. Gastroenterol. Res. Pract. 2013, 2013, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Kondo, K.; Muramatsu, M.; Okamoto, Y.; Jin, D.; Takai, S.; Tanigawa, N.; Miyazaki, M. Expression of chymase-positive cells in gastric cancer and its correlation with the angiogenesis. J. Surg. Oncol. 2005, 93, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D.; Guidolin, D.; Marzullo, A.; Nico, B.; Annese, T.; Benagiano, V.; Crivellato, E. Mast cells and angiogenesis in gastric carcinoma. Int. J. Exp. Pathol. 2010, 91, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Yano, H.; Kinuta, M.; Tateishi, H.; Nakano, Y.; Matsui, S.; Monden, T.; Okamura, J.; Sakai, M.; Okamoto, S. Mast cell infiltration around gastric cancer cells correlates with tumor angiogenesis and metastasis. Gastric Cancer 1999, 2, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Ammendola, M.; Sacco, R.; Sammarco, G.; Donato, G.; Montemurro, S.; Ruggieri, E.; Patruno, R.; Marech, I.; Cariello, M.; Vacca, A.; et al. Correlation between Serum Tryptase, Mast Cells Positive to Tryptase and Microvascular Density in Colo-Rectal Cancer Patients: Possible Biological-Clinical Significance. PLoS ONE 2014, 9, e99512. [Google Scholar] [CrossRef] [PubMed]
- Lachter, J.; Stein, M.; Lichtig, C.; Eidelman, S.; Munichor, M. Mast cells in colorectal neoplasias and premalignant disorders. Dis. Colon Rectum 1995, 38, 290–293. [Google Scholar] [CrossRef] [PubMed]
- Ammendola, M.; Sacco, R.; Sammarco, G.; Donato, G.; Zuccalà, V.; Luposella, M.; Patruno, R.; Marech, I.; Montemurro, S.; Zizzo, N.; et al. Mast Cells Density Positive to Tryptase Correlates with Angiogenesis in Pancreatic Ductal Adenocarcinoma Patients Having Undergone Surgery. Gastroenterol. Res. Pract. 2014, 2014, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Longo, V.; Tamma, R.; Brunetti, O.; Pisconti, S.; Argentiero, A.; Silvestris, N.; Ribatti, D. Mast cells and angiogenesis in pancreatic ductal adenocarcinoma. Clin. Exp. Med. 2018, 18, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Benítez–Bribiesca, L.; Wong, A.; Utrera, D.; Castellanos, E. The Role of Mast Cell Tryptase in Neoangiogenesis of Premalignant and Malignant Lesions of the Uterine Cervix. J. Histochem. Cytochem. 2001, 49, 1061–1062. [Google Scholar] [CrossRef] [PubMed]
- Graham, R.M.; Graham, J.B. Cytological prognosis in cancer of the uterine cervix treated radiologically. Cancer 1955, 8, 59–70. [Google Scholar] [CrossRef]
- Ribatti, D.; Finato, N.; Crivellato, E.; Marzullo, A.; Mangieri, D.; Nico, B.; Vacca, A.; Beltrami, C.A. Neovascularization and mast cells with tryptase activity increase simultaneously with pathologic progression in human endometrial cancer. Am. J. Obstet. Gynecol. 2005, 193, 1961–1965. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, A.M.; Mihm, M.C.; Osage, J.E.; Dvorak, H.F. Melanoma. An Ultrastructural Study of the Host Inflammatory and Vascular Responses. J. Investig. Dermatol. 1980, 75, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Reed, J.A.; McNutt, N.S.; Bogdany, J.K.; Albino, A.P. Expression of the mast cell growth factor interleukin-3 in melanocytic lesions correlates with an increased number of mast cells in the perilesional stroma: Implications for melanoma progression. J. Cutan. Pathol. 2006, 23, 495–505. [Google Scholar] [CrossRef]
- Ullah, E.; Nagi, A.H.; Lail, R.A. Angiogenesis and mast cell density in invasive pulmonary adenocarcinoma. J. Cancer Res. Ther. 2012, 8, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Melillo, R.M.; Guarino, V.; Avilla, E.; Galdiero, M.R.; Liotti, F.; Prevete, N.; Rossi, F.W.; Basolo, F.; Ugolini, C.; de Paulis, A.; et al. Mast cells have a protumorigenic role in human thyroid cancer. Oncogene 2010, 29, 6203–6215. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S. Prognostic value of angiogenesis in solitary bone plasmacytoma. Blood 2002, 101, 1715–1717. [Google Scholar] [CrossRef]
- Naganuma, H.; Sakatsume, S.; Sugita, M.; Satoh, E.; Asahara, T.; Nukui, H. Solitary Plasmacytoma of the Skull: Immunohistochemical Study of Angiogenic Factors and Syndecan-1-Two Case Reports. Neurologia Medico-Chirurgica 2004, 44, 195–200. [Google Scholar] [CrossRef]
- Nakayama, T.; Yao, L.; Tosato, G. Mast cell–derived angiopoietin-1 plays a critical role in the growth of plasma cell tumors. J. Clin. Investig. 2004, 114, 1317–1325. [Google Scholar] [CrossRef]
- Swelam, W.M.; Al Tamimi, D.M. Biological impact of vascular endothelial growth factor on vessel density and survival in multiple myeloma and plasmacytoma. Pathol. Res. Pract. 2010, 206, 753–759. [Google Scholar] [CrossRef]
- Vacca, A.; Ribatti, D.; Roncali, L.; Ranieri, G.; Serio, G.; Silvestris, F.; Dammacco, F. Bone marrow angiogenesis and progression in multiple myeloma. Br. J. Haematol. 1994, 87, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Vacca, A.; Ribatti, D.; Presta, M.; Minischetti, M.; Iurlaro, M.; Ria, R.; Albini, A.; Bussolino, F.; Dammacco, F. Bone marrow neovascularization, plasma cell angiogenic potential, and matrix metalloproteinase-2 secretion parallel progression of human multiple myeloma. Blood 1999, 93, 3064–3073. [Google Scholar] [PubMed]
- Ribatti, D.; Nico, B.; Vacca, A. Importance of the bone marrow microenvironment in inducing the angiogenic response in multiple myeloma. Oncogene 2006, 25, 4257–4266. [Google Scholar] [CrossRef] [PubMed]
- Vacca, A.; Ribatti, D. Bone marrow angiogenesis in multiple myeloma. Leukemia 2005, 20, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Nico, B.; Mangieri, D.; Crivellato, E.; Vacca, A.; Ribatti, D. Mast Cells Contribute to Vasculogenic Mimicry in Multiple Myeloma. Stem Cells Dev. 2008, 17, 19–22. [Google Scholar] [CrossRef]
- Mileshkin, L.; Honemann, D.; Gambell, P.; Trivett, M.; Hayakawa, Y.; Smyth, M.; Beshay, V.; Ritchie, D.; Simmons, P.; Milner, A.D.; et al. Patients with multiple myeloma treated with thalidomide: Evaluation of clinical parameters, cytokines, angiogenic markers, mast cells and marrow CD57+ cytotoxic T cells as predictors of outcome. Haematologica 2007, 92, 1075–1082. [Google Scholar] [CrossRef]
- Pappa, C.A.; Tsirakis, G.; Devetzoglou, M.; Zafeiri, M.; Vyzoukaki, R.; Androvitsanea, A.; Xekalou, A.; Sfiridaki, K.; Alexandrakis, M.G. Bone marrow mast cell density correlates with serum levels of VEGF and CXC chemokines ENA-78 and GRO-α in multiple myeloma. Tumor Biol. 2014, 35, 5647–5651. [Google Scholar] [CrossRef]
- Pappa, C.A.; Tsirakis, G.; Roussou, P.; Xekalou, A.; Goulidaki, N.; Konsolas, I.; Alexandrakis, M.G.; Stathopoulos, E.N. Positive correlation between bone marrow mast cell density and ISS prognostic index in patients with multiple myeloma. Leuk. Res. 2013, 37, 1628–1631. [Google Scholar] [CrossRef]
- Pappa, C.A.; Tsirakis, G.; Stavroulaki, E.; Kokonozaki, M.; Xekalou, A.; Konsolas, I.; Alexandrakis, M.G. Mast Cells Influence the Proliferation Rate of Myeloma Plasma Cells. Cancer Investig. 2015, 33, 137–141. [Google Scholar] [CrossRef]
- Devetzoglou, M.; Vyzoukaki, R.; Kokonozaki, M.; Xekalou, A.; Pappa, C.A.; Papadopoulou, A.; Alegakis, A.; Androulakis, N.; Alexandrakis, M.G. High density of tryptase-positive mast cells in patients with multiple myeloma: Correlation with parameters of disease activity. Tumor Biol. 2015, 36, 8491–8497. [Google Scholar] [CrossRef] [PubMed]
- Vyzoukaki, R.; Tsirakis, G.; Pappa, C.A.; Androulakis, N.; Kokonozaki, M.; Tzardi, M.; Alexandrakis, M.G. Correlation of Mast Cell Density with Angiogenic Cytokines in Patients with Active Multiple Myeloma. Clin. Ther. 2016, 38, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Vyzoukaki, R.; Tsirakis, G.; Pappa, C.A.; Devetzoglou, M.; Tzardi, M.; Alexandrakis, M.G. The Impact of Mast Cell Density on the Progression of Bone Disease in Multiple Myeloma Patients. Int. Arch. Allergy Immunol. 2015, 168, 263–268. [Google Scholar] [CrossRef]
- Attar-Schneider, O.; Drucker, L.; Zismanov, V.; Tartakover-Matalon, S.; Rashid, G.; Lishner, M. Bevacizumab attenuates major signaling cascades and eIF4E translation initiation factor in multiple myeloma cells. Lab. Investig. 2012, 92, 178–190. [Google Scholar] [CrossRef] [PubMed]
- White, D.; Kassim, A.; Bhaskar, B.; Yi, J.; Wamstad, K.; Paton, V.E. Results from AMBER, a randomized phase 2 study of bevacizumab and bortezomib versus bortezomib in relapsed or refractory multiple myeloma. Cancer 2013, 119, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Somlo, G.; Lashkari, A.; Bellamy, W.; Zimmerman, T.M.; Tuscano, J.M.; O’Donnell, M.R.; Mohrbacher, A.F.; Forman, S.J.; Frankel, P.; Chen, H.X.; et al. Phase II randomized trial of bevacizumab versus bevacizumab and thalidomide forelapsed/refractory multiple myeloma: A California Cancer Consortium trial. Br. J. Hematol. 2011, 154, 533–535. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tumor | References |
|---|---|
| Haemangioma, haemangioblastoma | [41] |
| Lymphomas | [42,43,44,45] |
| Multiple myeloma | [46] |
| Bagg AlbinoMyelodysplastic syndrome | [47] |
| B-cell chronic lymphocytic leukemia | [38,48] |
| Breast cancer | [49,50,51,52,53] |
| Gastric cancer | [54,55,56,57] |
| Colorectal cancer | [58,59] |
| Pancreatic cancer | [60,61] |
| Uterine cervix cancer | [62,63,64] |
| Melanoma | [39,65,66] |
| Pulmonary adenocarcinoma | [67] |
| Thyroid cancer | [68] |
| Drug | Main Target in Mast Cell |
|---|---|
| Imatinib mesylate (Gleevec, ST1571) | c-kit |
| Sorafenib | c-kit |
| Sunitinib | c-kit |
| Pazopanib (GW786034) | c-kit |
| Axitinib | c-kit |
| Dasatinib | c-kit |
| Enzastaurin | PKC-β |
| Alemtuzumab (Campath) | CD52 |
| CpG activator (Promune) | TLR9 |
| MDX 060 | CD30 (ligand for CD301) * |
| Tanespimycin (17AAG) | Heat shock protein 90 β |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ribatti, D.; Tamma, R.; Vacca, A. Mast Cells and Angiogenesis in Human Plasma Cell Malignancies. Int. J. Mol. Sci. 2019, 20, 481. https://doi.org/10.3390/ijms20030481
Ribatti D, Tamma R, Vacca A. Mast Cells and Angiogenesis in Human Plasma Cell Malignancies. International Journal of Molecular Sciences. 2019; 20(3):481. https://doi.org/10.3390/ijms20030481
Chicago/Turabian StyleRibatti, Domenico, Roberto Tamma, and Angelo Vacca. 2019. "Mast Cells and Angiogenesis in Human Plasma Cell Malignancies" International Journal of Molecular Sciences 20, no. 3: 481. https://doi.org/10.3390/ijms20030481
APA StyleRibatti, D., Tamma, R., & Vacca, A. (2019). Mast Cells and Angiogenesis in Human Plasma Cell Malignancies. International Journal of Molecular Sciences, 20(3), 481. https://doi.org/10.3390/ijms20030481