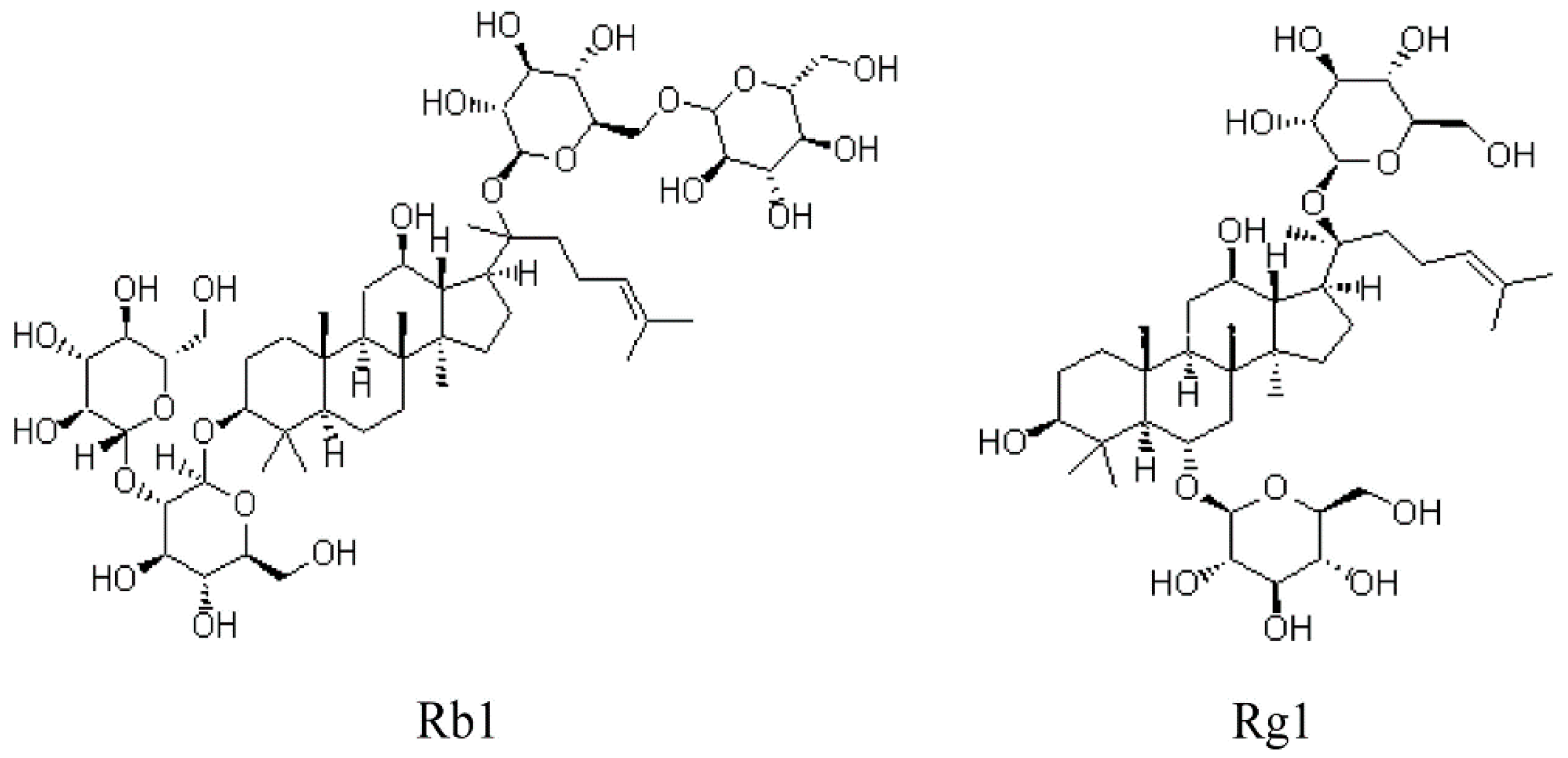
Ginsenosides Rb1 and Rg1 Protect Primary Cultured Astrocytes against Oxygen-Glucose Deprivation/Reoxygenation-Induced Injury via Improving Mitochondrial Function


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
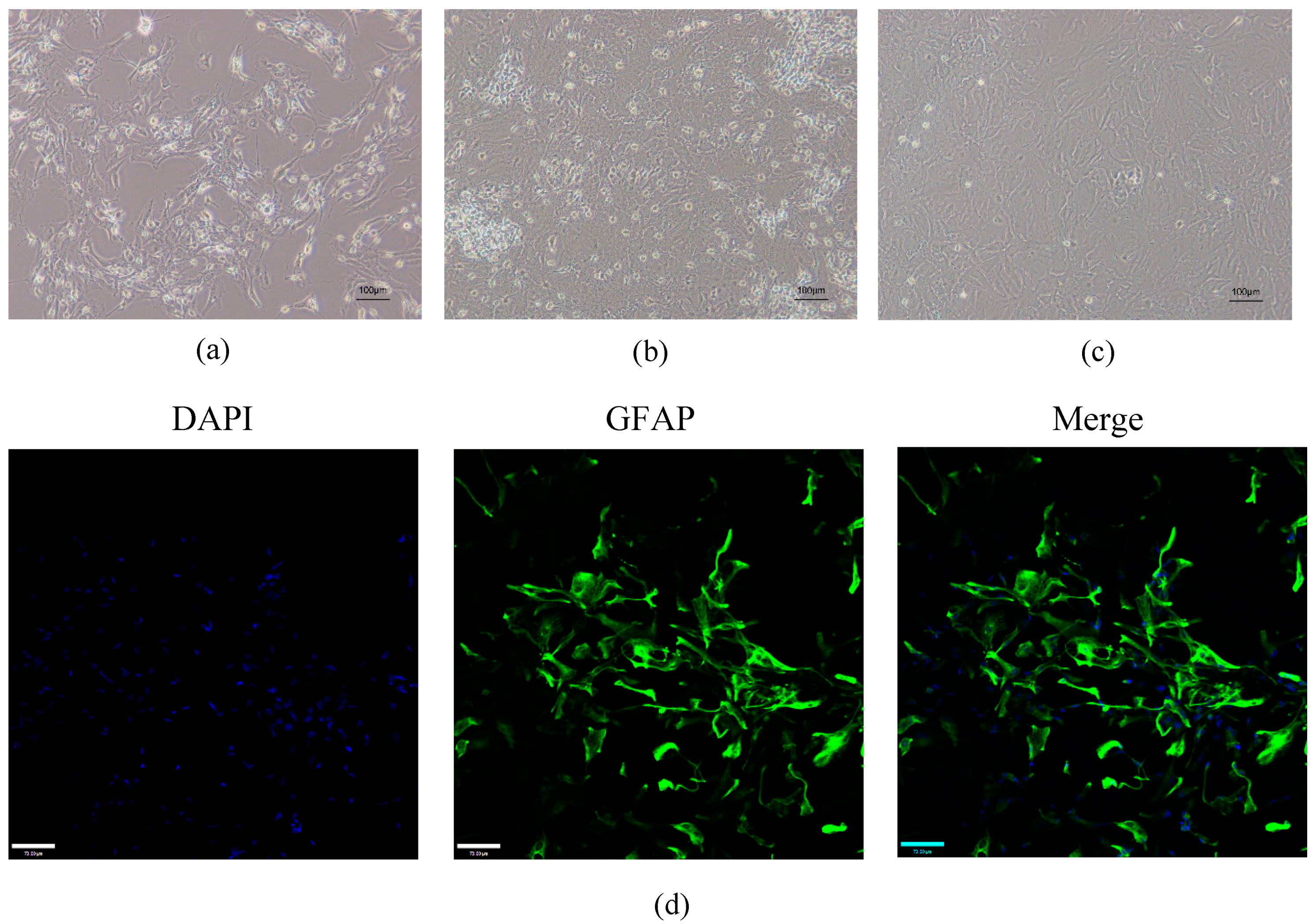
2.1. Validation of Mouse Astrocytes
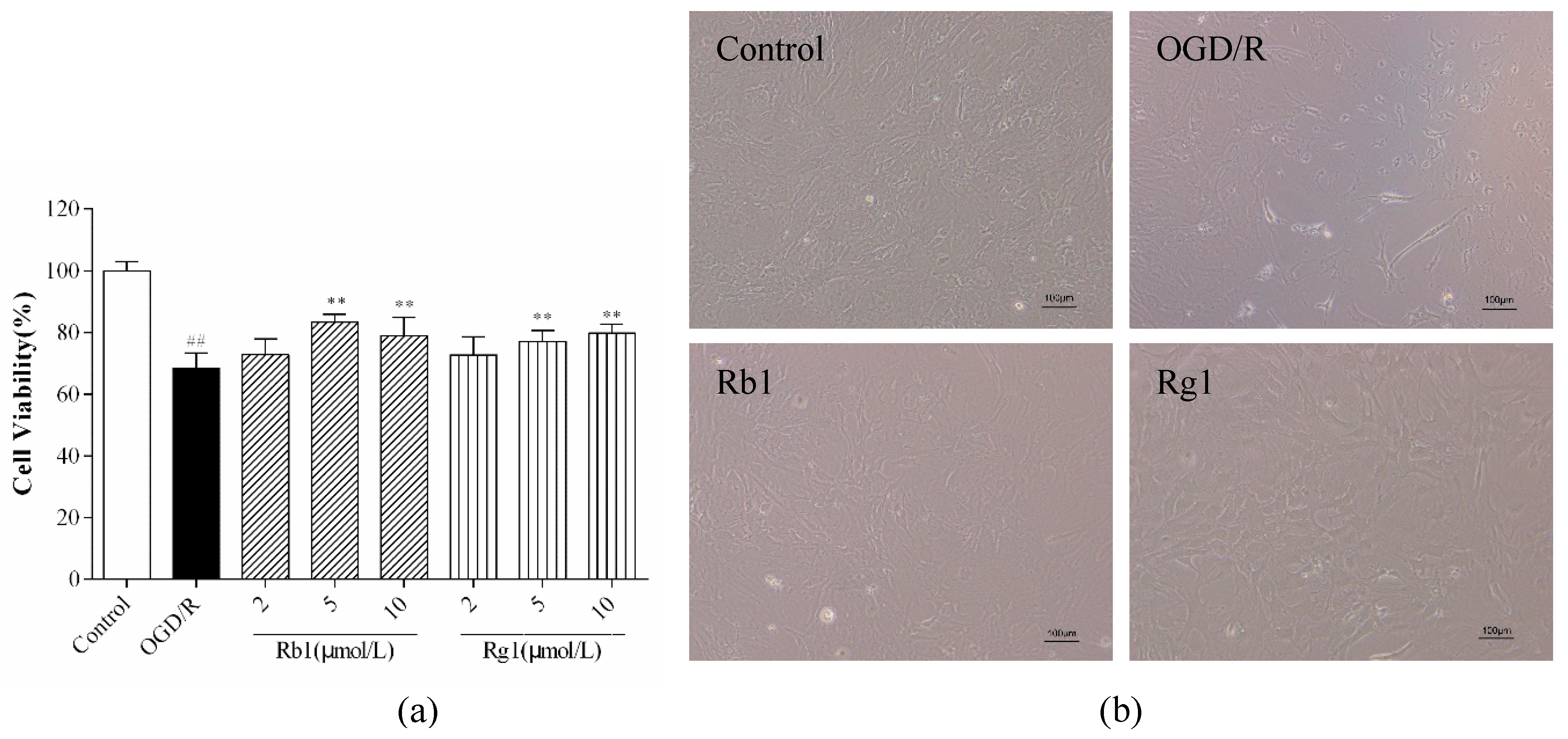
2.2. Rb1 and Rg1 Attenuated OGD/R-Induced Injury in Cultured Astrocyte
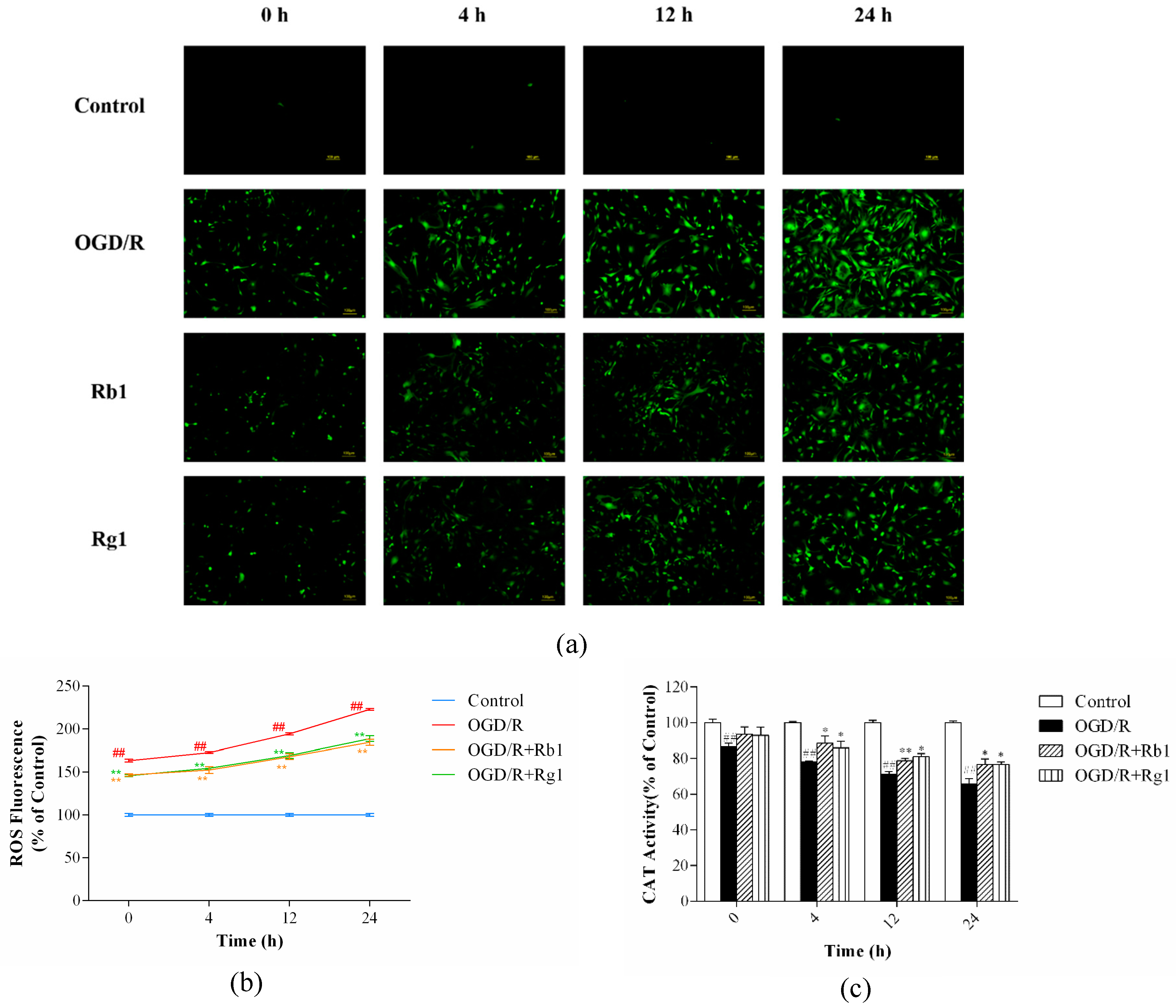
2.3. Rb1 and Rg1 Suppressed ROS Production and Increased CAT Activity in OGD/R-Treated Astrocytes
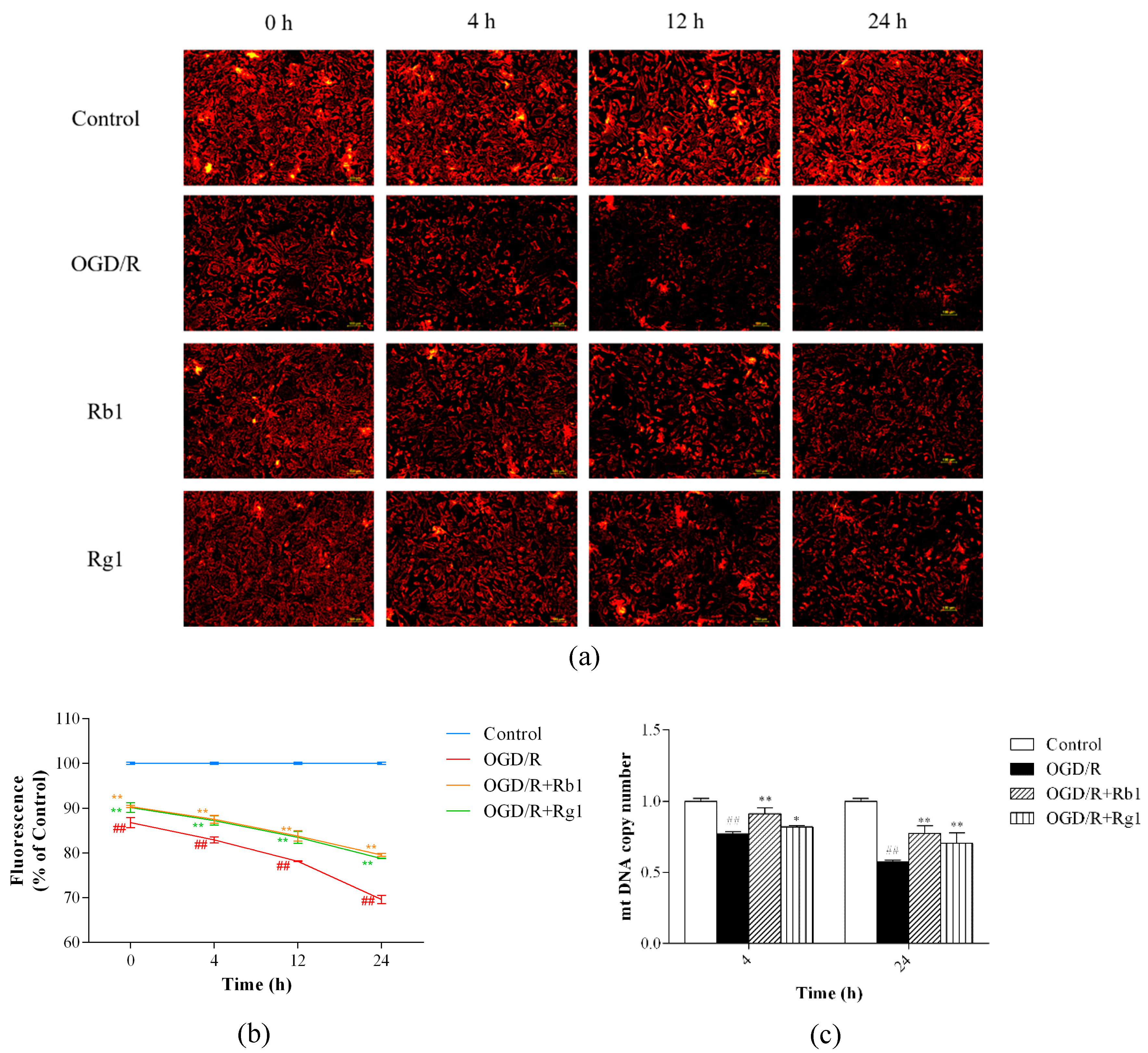
2.4. Rb1 and Rg1 Inhibited MMP Depolarization and Increased mtDNA Content in OGD/R-Treated Astrocytes
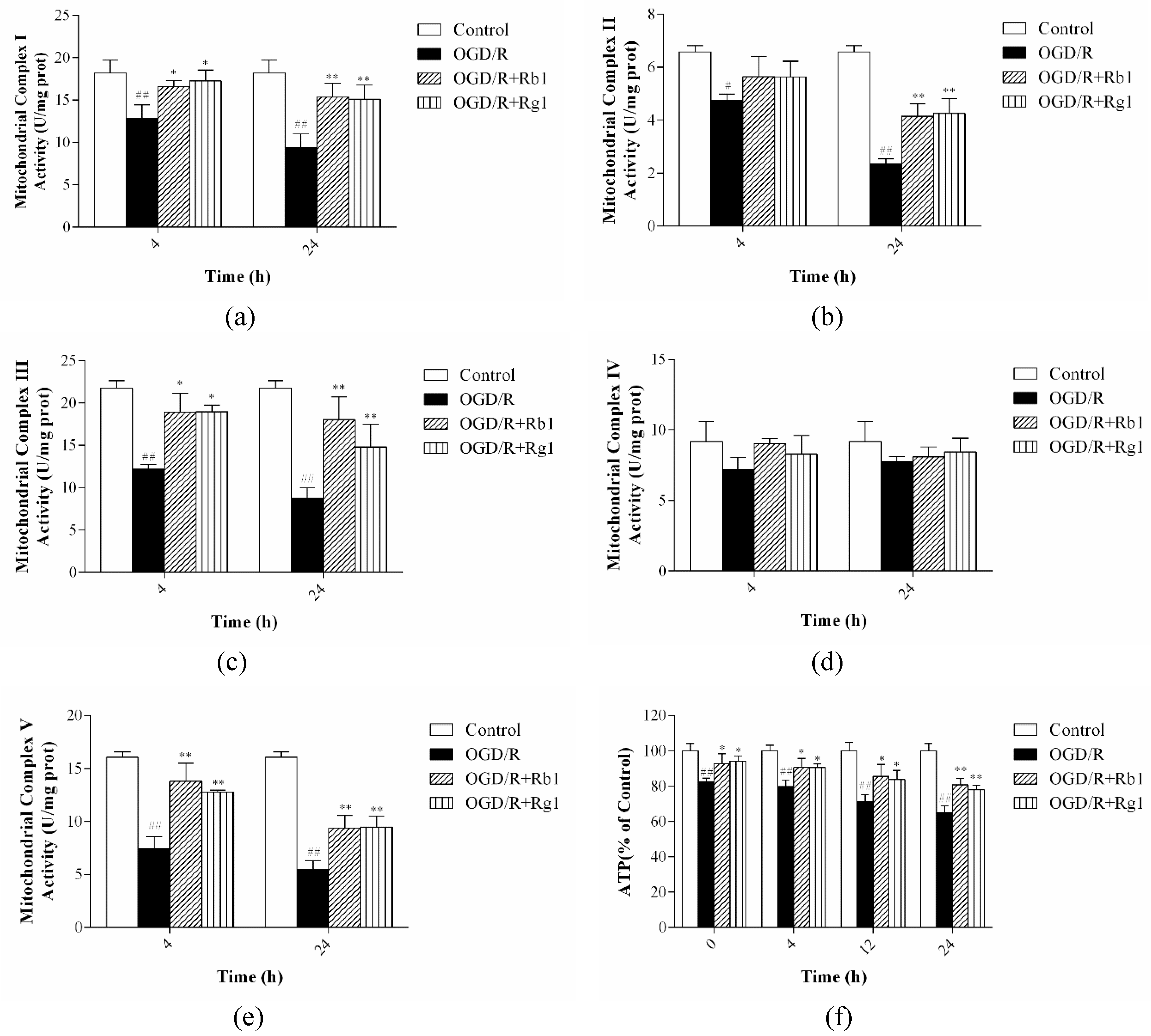
2.5. Rb1 and Rg1 Increased the Activities of Mitochondrial Respiratory Chain Complexes I–V and ATP Levels
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Cell Culture
4.3. Identification of Astrocytes
4.4. OGD/R Procedure and Drug Treatment
4.5. CCK-8 Assay
4.6. Morphology Observation of Astrocytes
4.7. Measurement of Intracellular ROS Levels
4.8. Catalase Activity
4.9. Detection of the Mitochondrial Membrane Potential
4.10. Measurement of Intracellular ATP Levels
4.11. Real-Time Quantitative Polymerase Chain Reaction (PCR) Analysis
4.12. Isolation of Mitochondria from Astrocytes
4.13. Assay of the Activities of Mitochondrial Respiratory Chain Complexes I–V in Astrocytes
4.14. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Rb1 | Ginsenoside Rb1 |
| Rg1 | Ginsenoside Rg1 |
| OGD/R | Oxygen–glucose deprivation/reoxygenation |
| ROS | Reactive oxygen species (ROS) |
| CAT | Catalase |
| NADPH | Nicotinamide adenine dinucleotide phosphate |
| MMP | Mitochondrial membrane potential |
| MPTP | Mitochondrial permeability transition pore |
| mtDNA | Mitochondrial DNA |
| MRC | Mitochondrial respiratory chain |
| ATP | Adenosine triphosphate |
| OXPHOS | Oxidative phosphorylation |
| ETC | Electron transport chain |
| DCFA-DA | 2′,7′-dichlorodihydrofluorescein diacetate |
References
- Annunziato, L.; Boscia, F.; Pignataro, G. Ionic transporter activity in astrocytes, microglia and oligodendrocytes during brain ischemia. J. Cerebr. Blood F Met. 2013, 33, 969–982. [Google Scholar] [CrossRef]
- GuillamónVivancos, T.; GómezPinedo, U.; MatíasGuiu, J. Astrocytes in neurodegenerative diseases (I): Function and molecular description. Neurologia 2015, 30, 119–129. [Google Scholar]
- Robb, S.J.; Robb-Gaspers, L.D.; Scaduto, R.C.; Thomas, A.P.; Connor, J.R. Influence of calcium and iron on cell death and mitochondrial function in oxidatively stressed astrocytes. J. Neurosci. Res. 2015, 55, 674–686. [Google Scholar] [CrossRef]
- Voloboueva, L.A.; Suh, S.W.; Swanson, R.A.; Rona, G.G. Inhibition of mitochondrial function in astrocytes: Implications for neuroprotection. J. Neurochem. 2007, 102, 1383–1394. [Google Scholar] [CrossRef] [PubMed]
- Nah, S.Y.; Mccleskey, E.W. Ginseng root extract inhibits calcium channels in rat sensory neurons through a similar path, but different receptor, as μ-type opioids. J. Ethonpharmacol. 1994, 42, 45–51. [Google Scholar] [CrossRef]
- Nah, S.-Y.; Kim, D.-H.; Rhim, H. Ginsenosides: Are any of them candidates for drugs acting on the central nervous system? CNS Drug Rev. 2007, 13, 381–404. [Google Scholar] [CrossRef]
- Cheng, Y.; Shen, L.H.; Zhang, J.T. Anti-amnestic and anti-aging effects of ginsenoside Rg1 and Rb1 and its mechanism of action. Acta Pharm. 2005, 26, 143–149. [Google Scholar] [CrossRef]
- Zheng, X.; Liang, Y.; Kang, A.; Ma, S.J.; Xing, L.; Zhou, Y.Y.; Dai, C.; Xie, H.; Xie, L.; Wang, G.J.; et al. Peripheral immunomodulation with ginsenoside Rg1 ameliorates neuroinflammation-induced behavioral deficits in rats. Neuroscience 2014, 256, 210–222. [Google Scholar] [CrossRef]
- Li, T.; Shu, Y.J.; Cheng, J.Y.; Liang, R.C.; Dian, S.N.; Lv, X.X.; Yang, M.Q.; Huang, S.L.; Chen, G.; Yang, F. Pharmacokinetics and efficiency of brain targeting of ginsenosides Rg1 and Rb1 given as Nao-Qing microemulsion. Drug Dev. Ind. Pharm. 2015, 41, 224–231. [Google Scholar] [CrossRef]
- Li, Y.; Suo, L.; Liu, Y.; Li, H.; Xue, W. Protective effects of ginsenoside Rg1 against oxygen-glucose-deprivation-induced apoptosis in neural stem cells. J. Neuro. Sci. 2017, 373, 107–112. [Google Scholar] [CrossRef]
- Zeng, X.S.; Zhou, X.S.; Luo, F.C.; Jia, J.J.; Qi, L.; Yang, Z.X.; Zhang, W.; Bai, J. Comparative analysis of the neuroprotective effects of ginsenosides Rg1 and Rb1 extracted from Panax notoginseng against cerebral ischemia. J. Physiol. Pharm. 2014, 92, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.P.; Ding, H.; Lu, J.D.; Tang, Y.H.; Deng, B.X.; Deng, C.Q. Effects of the main active components combinations of Astragalus and Panax notoginseng on energy metabolism in brain tissues after cerebral ischemia-reperfusion in mice. Pharm. Mag. 2015, 11, 732–739. [Google Scholar]
- Xie, W.; Zhou, P.; Sun, Y.; Meng, X.; Dai, Z.; Sun, G.; Sun, X. Protective Effects and Target Network Analysis of Ginsenoside Rg1 in Cerebral Ischemia and Reperfusion Injury: A Comprehensive Overview of Experimental Studies. Cells 2018, 7, 270. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.Z.; Tian, Y.; Li, J. GCN2 suppression attenuates cerebral ischemia in mice by reducing apoptosis and endoplasmic reticulum (ER) stress through the blockage of FoxO3a-regulated ROS production. Biochem. Bioph. Res. Co. 2019, 256, 210–222. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Ling, C.L.; Pang, L.; Wang, Q.; Liu, J.X.; Wang, B.S.; Liang, J.M.; Guo, Y.Z.; Qin, J.; Wang, J.X. Direct Macromolecular Drug Delivery to Cerebral Ischemia Area using Neutrophil-Mediated Nanoparticles. Theranostics 2017, 7, 3260–3275. [Google Scholar] [CrossRef]
- Turrens, J.F. Superoxide Production by the Mitochondrial Respiratory Chain. Biosci. Rep. 1997, 17, 3–8. [Google Scholar] [CrossRef]
- Nissanka, N.; Moraes, C.T. Mitochondrial DNA damage and reactive oxygen species in neurodegenerative disease. FEBS Lett. 2018, 592, 728–742. [Google Scholar] [CrossRef]
- Zhu, Z.; Wang, X. Significance of Mitochondria DNA Mutations in Diseases. Adv. Exp. Med. Biol. 2017, 1038, 219–230. [Google Scholar]
- Malchiodi-Albedi, F.; Domenici, M.R.; Paradisi, S.; Bernardo, A.; Ajmone-Cat, M.A.; Minghetti, L. Astrocytes contribute to neuronal impairment in βA toxicity increasing apoptosis in rat hippocampal neurons. Glia 2001, 34, 68–72. [Google Scholar] [CrossRef]
- Zhao, X.; Ahram, A.; Berman, R.F.; Muizelaar, J.P.; Lyeth, B.G. Early loss of astrocytes after experimental traumatic brain injury. Glia 2003, 44, 140–152. [Google Scholar] [CrossRef]
- Heales, S.J.; Lam, A.A.; Duncan, A.J.; Land, J.M. Neurodegeneration or neuroprotection: The pivotal role of astrocytes. Neurochem. Res. 2004, 29, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Xia, R.; Ji, C.; Zhang, L. Neuroprotective Effects of Pycnogenol Against Oxygen-Glucose Deprivation/Reoxygenation-Induced Injury in Primary Rat Astrocytes via NF-κB and ERK1/2 MAPK Pathways. Cell Physiol. Biochem. 2017, 42, 987–998. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Wang, B.; Liu, Y.; Zhang, J.; Huang, Y.; Cao, P.; Shen, Y.; Lyu, J. Carnosine modulates glutamine synthetase expression in senescent astrocytes exposed to oxygen-glucose deprivation/recovery. Brain Res. Bull. 2017, 130, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Liu, J.; Dong, R.; Zhu, J.; Tao, C.; Zheng, R.; Zhu, S. 14,15-epoxyeicosatrienoic acid promotes production of brain derived neurotrophic factor from astrocytes and exerts neuroprotective effects during ischaemic injury. Neuropath. Appl. Neuro. 2016, 42, 607–620. [Google Scholar] [CrossRef] [PubMed]
- Yingchun, Z.; Qiqiang, T.; Guopin, W.; Han, G. Tanshinone IIA Protects Hippocampal Neuronal Cells from Reactive Oxygen Species through Changes in Autophagy and Activation of Phosphatidylinositol 3-Kinase, Protein Kinas B, and Mechanistic Target of Rapamycin Pathways. Curr. Neurovasc. Res. 2017, 14, 132–140. [Google Scholar]
- Wei, S.; Tong, J.; Xue, Q.; Liu, Y.; Xu, X. Effect of puerarin on transcriptome of astrocyte during oxygen-glucose deprivation/reoxygenation injury. Mol. Cell Biochem. 2016, 425, 1–11. [Google Scholar] [CrossRef]
- Dan, D.J.; Alvarez, L.A.; Zhang, X.; Soldati, T. Reactive oxygen species and mitochondria: A nexus of cellular homeostasis. Redox Biol. 2015, 6, 472–485. [Google Scholar] [CrossRef]
- Bhat, A.H.; Dar, K.B.; Anees, S.; Zargar, M.A.; Masood, A.; Sofi, M.A.; Ganie, S.A. Oxidative stress, mitochondrial dysfunction and neurodegenerative diseases; a mechanistic insight. Biomed. Pharm. 2015, 74, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.M.; Huang, D.Y.; Huang, Y.P.; Hsu, S.H.; Kang, L.Y.; Shen, C.M.; Lin, W.W. Methylglyoxal induces cell death through endoplasmic reticulum stress-associated ROS production and mitochondrial dysfunction. J. Cell Mol. Med. 2016, 20, 1749–1760. [Google Scholar] [CrossRef]
- Moskowitz, M.A.; Lo, E.H.; Iadecola, C. The Science of Stroke: Mechanisms in Search of Treatments. Neuron 2010, 67, 181–198. [Google Scholar] [CrossRef]
- Solaini, G.; Sgarbi, G.; Baracca, A. Oxidative phosphorylation in cancer cells. Biochim. Biophys. Acta 2011, 1807, 534–542. [Google Scholar] [CrossRef]
- Martin, J.; St-Pierre, M.V.; Dufour, J.F. Hit proteins, mitochondria and cancer. Biochim. Biophys. Acta 2011, 1807, 626–632. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.W.; Simone, C.; Armen, A.; Chen, J.Z. Simultaneous Quantification of Mitochondrial DNA Damage and Copy Number in Circulating Blood: A Sensitive Approach to Systemic Oxidative Stress. Biomed. Res. Int. 2013, 2013, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Leon, J.; Sakumi, K.; Castillo, E.; Sheng, Z.; Oka, S.; Nakabeppu, Y. 8-Oxoguanine accumulation in mitochondrial DNA causes mitochondrial dysfunction and impairs neuritogenesis in cultured adult mouse cortical neurons under oxidative conditions. Sci. Rep. 2016, 6, 1–15. [Google Scholar]
- Ming, Y.Z.; Dong, L.Z.; Chen, Z.; Zhen, C. Mild Acidosis Protects Neurons during Oxygen-Glucose Deprivation by Reducing Loss of Mitochondrial Respiration. ACS Chem. Neurosci. 2019, 10, 2489–2497. [Google Scholar]
- Yu, H.; Guan, Q.; Guo, L.; Zhang, H.; Pang, X.; Cheng, Y.; Zhang, X.; Sun, Y. Gypenosides alleviate myocardial ischemia-reperfusion injury via attenuation of oxidative stress and preservation of mitochondrial function in rat heart. Cell Stress Chaperones 2016, 21, 429–437. [Google Scholar] [CrossRef]
- Sha, D.; Wang, L.; Zhang, J.; Qian, L.; Li, Q.; Li, J.; Qian, J.; Gu, S.; Han, L.; Xu, P.; et al. Cocaine- and amphetamine-regulated transcript peptide increases mitochondrial respiratory chain complex II activity and protects against oxygen–glucose deprivation in neurons. Brain Res. 2014, 1582, 107–113. [Google Scholar] [CrossRef]
- Kogot-Levin, A.; Saada, A. Ceramide and the mitochondrial respiratory chain. Biochimie 2014, 100, 88–94. [Google Scholar] [CrossRef]
- Shi, Y.; Sun, C.; Zheng, B.; Li, Y.; Wang, Y. Simultaneous determination of nine ginsenosides in functional foods by high performance liquid chromatography with diode array detector detection. Food Chem. 2010, 123, 1322–1327. [Google Scholar] [CrossRef]
- Chang, H.L.; Kim, J.H. A review on the medicinal potentials of ginseng and ginsenosides on cardiovascular diseases. J. Ginseng Res. 2014, 38, 161–166. [Google Scholar]
- Yu, A.C.; Lau, L.T. Expression of interleukin-1 alpha, tumor necrosis factor alpha and interleukin-6 genes in astrocytes under ischemic injury. Neurochem. Int. 2000, 36, 369–377. [Google Scholar] [CrossRef]
- Rose, J.; Brian, C.; Woods, J.; Pappa, A.; Panayiotidis, M.I.; Powers, R.; Franco, R. Mitochondrial dysfunction in glial cells: Implications for neuronal homeostasis and survival. Toxicology 2017, 391, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Boengler, K.; Hilfiker-Kleiner, D.; Heusch, G.; Schulz, R. Inhibition of permeability transition pore opening by mitochondrial STAT3 and its role in myocardial ischemia/reperfusion. Basic Res. Cardiol. 2010, 105, 771–785. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Sheng, M.; Wang, J.; Teng, T.; Sun, Y.; Yang, Q.; Xu, Z. Zinc improves mitochondrial respiratory function and prevents mitochondrial ROS generation at reperfusion by phosphorylating STAT3 at Ser727. J. Mol. Cell Cardiol. 2018, 118, 169–182. [Google Scholar] [CrossRef]
- National Research Council. Guide for the Care and Use of Laboratory Animals, 8th ed.; NIH Publication: Washington, DC, USA, 2011; pp. 963–965.
- McCarthy, K.D. Preparation of separate astroglial and oligodendroglial cell cultures from rat cerebral tissue. J. Cell Biol. 1980, 85, 890–902. [Google Scholar] [CrossRef]
- Yan, M.; Pei, Z.; Jinqiang, Z.; Chen, Y.; Lin, L.; Han, Z.; Meng, Z.; Xiumei, G.; Xiang, F. Naoxintong Protects Primary Neurons from Oxygen-Glucose Deprivation/Reoxygenation Induced Injury through PI3K-Akt Signaling Pathway. Evid-Based Compl. Alt. 2016, 2016, 1–12. [Google Scholar] [CrossRef]
- Sakamuru, S.; Attene-Ramos, M.S.; Xia, M. Mitochondrial Membrane Potential Assay. Methods Mol. Biol. 2016, 1473, 17–22. [Google Scholar]
- Ylikallio, E.; Page, J.L.; Xu, X. Ribonucleotide reductase is not limiting for mitochondrial DNA copy number in mice. Nucleic Acids Res. 2010, 38, 8208–8218. [Google Scholar] [CrossRef]
- Chunlan, F.; Yuan, Q.; Minke, T. Notoginsenoside R1 attenuates high glucose-induced endothelial damage in rat retinal capillary endothelial cells by modulating the intracellular redox state. Drug Des. Dev. 2017, 11, 3343–3354. [Google Scholar]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, M.; Ma, Q.; Fan, C.; Chen, X.; Zhang, H.; Tang, M. Ginsenosides Rb1 and Rg1 Protect Primary Cultured Astrocytes against Oxygen-Glucose Deprivation/Reoxygenation-Induced Injury via Improving Mitochondrial Function. Int. J. Mol. Sci. 2019, 20, 6086. https://doi.org/10.3390/ijms20236086
Xu M, Ma Q, Fan C, Chen X, Zhang H, Tang M. Ginsenosides Rb1 and Rg1 Protect Primary Cultured Astrocytes against Oxygen-Glucose Deprivation/Reoxygenation-Induced Injury via Improving Mitochondrial Function. International Journal of Molecular Sciences. 2019; 20(23):6086. https://doi.org/10.3390/ijms20236086
Chicago/Turabian StyleXu, Meng, Qing Ma, Chunlan Fan, Xue Chen, Huiming Zhang, and Minke Tang. 2019. "Ginsenosides Rb1 and Rg1 Protect Primary Cultured Astrocytes against Oxygen-Glucose Deprivation/Reoxygenation-Induced Injury via Improving Mitochondrial Function" International Journal of Molecular Sciences 20, no. 23: 6086. https://doi.org/10.3390/ijms20236086
APA StyleXu, M., Ma, Q., Fan, C., Chen, X., Zhang, H., & Tang, M. (2019). Ginsenosides Rb1 and Rg1 Protect Primary Cultured Astrocytes against Oxygen-Glucose Deprivation/Reoxygenation-Induced Injury via Improving Mitochondrial Function. International Journal of Molecular Sciences, 20(23), 6086. https://doi.org/10.3390/ijms20236086