Analysis of Pollen Allergens in Lily by Transcriptome and Proteome Data

 ,
,
Abstract
1. Introduction
2. Results
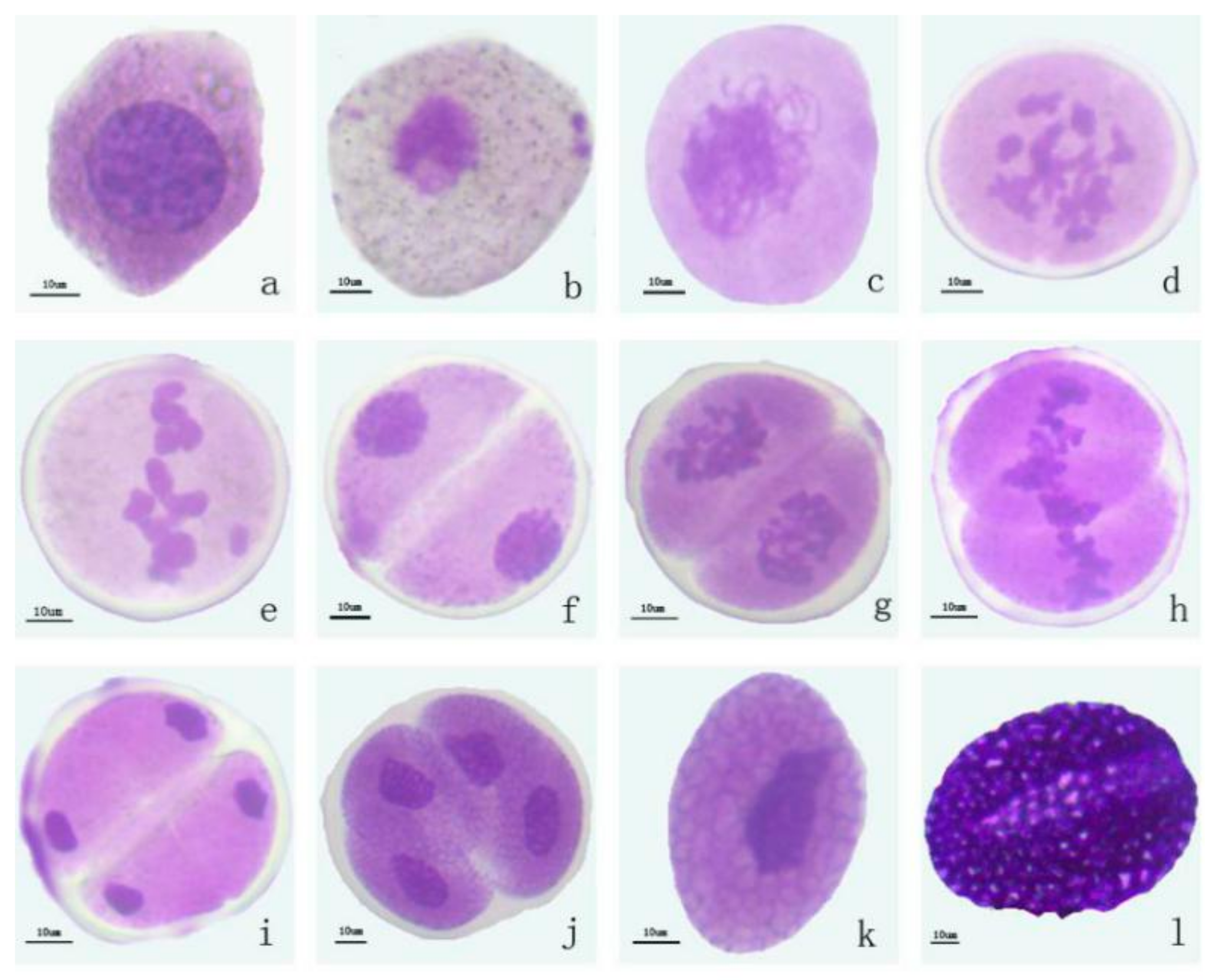
2.1. Morphological and Cytological Characterization of Lily
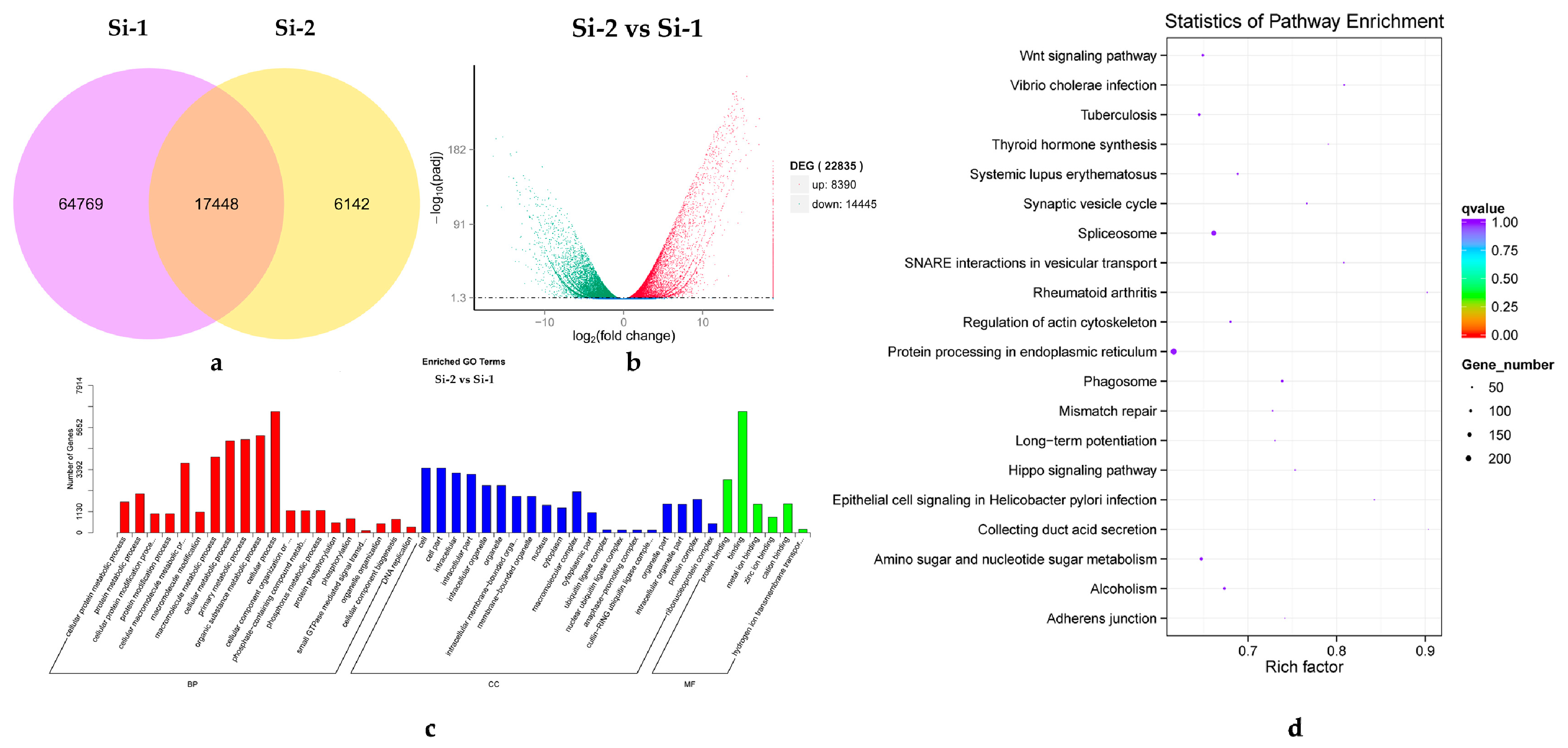
2.2. Transcriptome Sequencing and Transcript Assembly
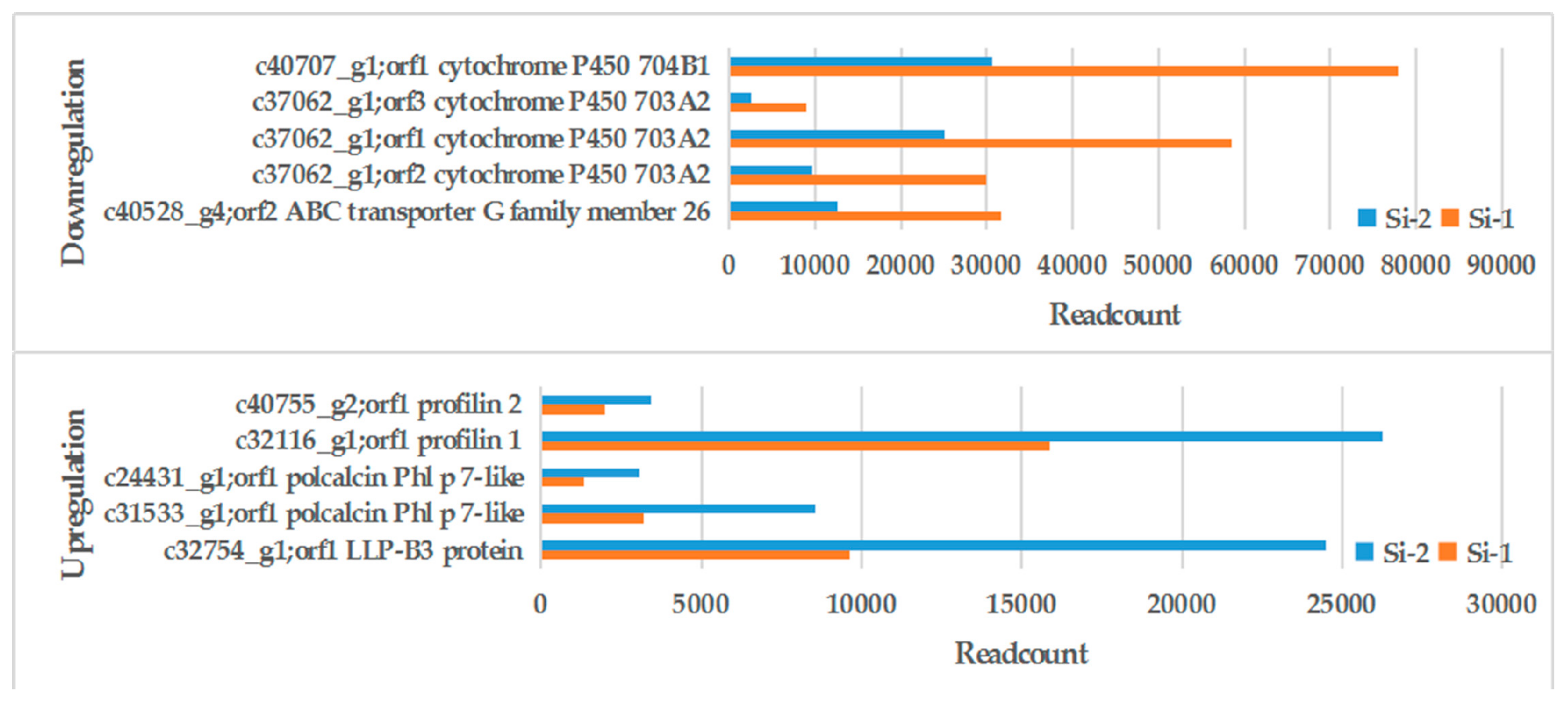
2.3. Gene Function Annotation and Analysis of Differentially Expressed Genes (DEGs)
2.4. Protein Function Annotation and DEPs Analysis
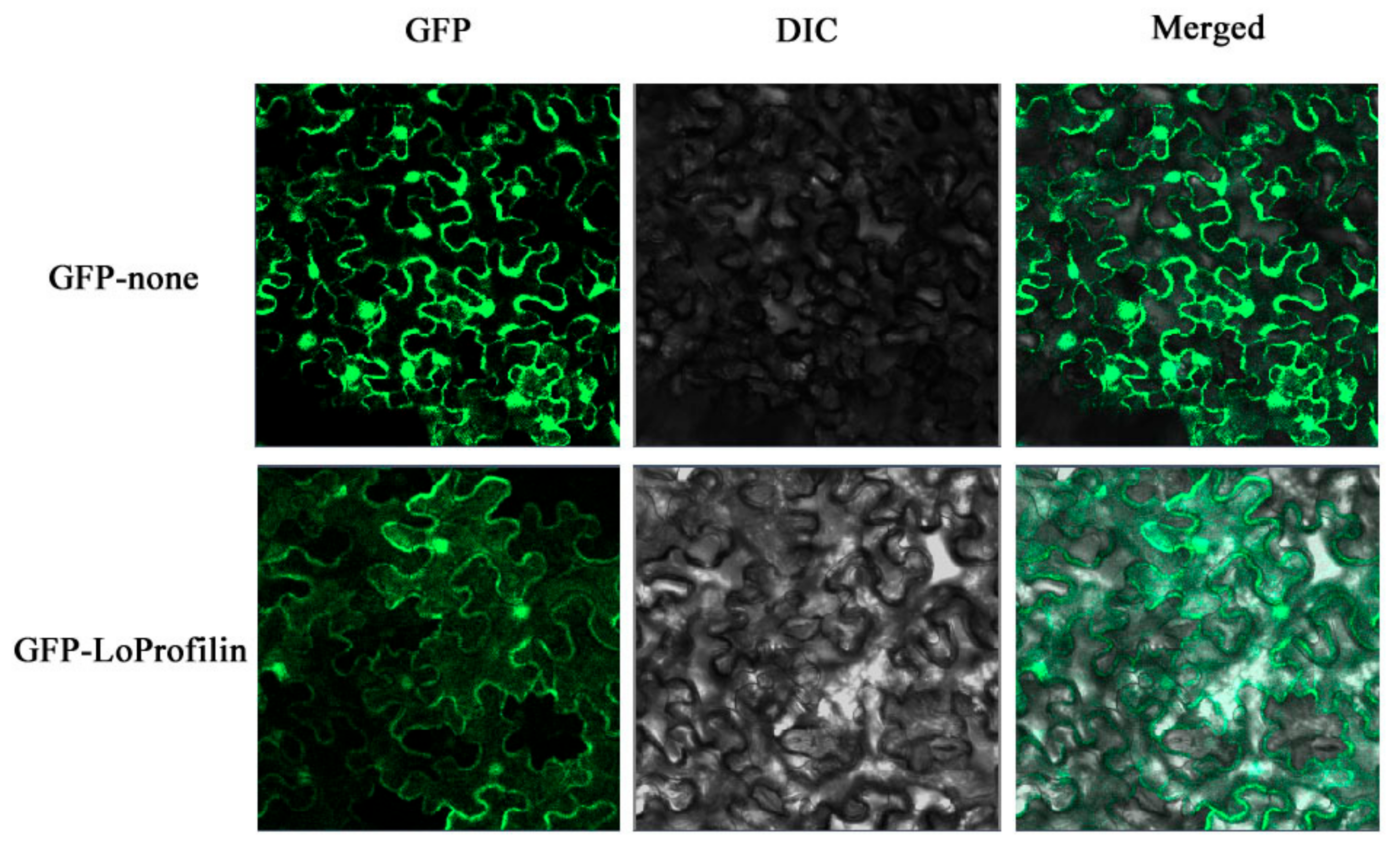
2.5. Gene Cloning and Protein Subcellular Localization
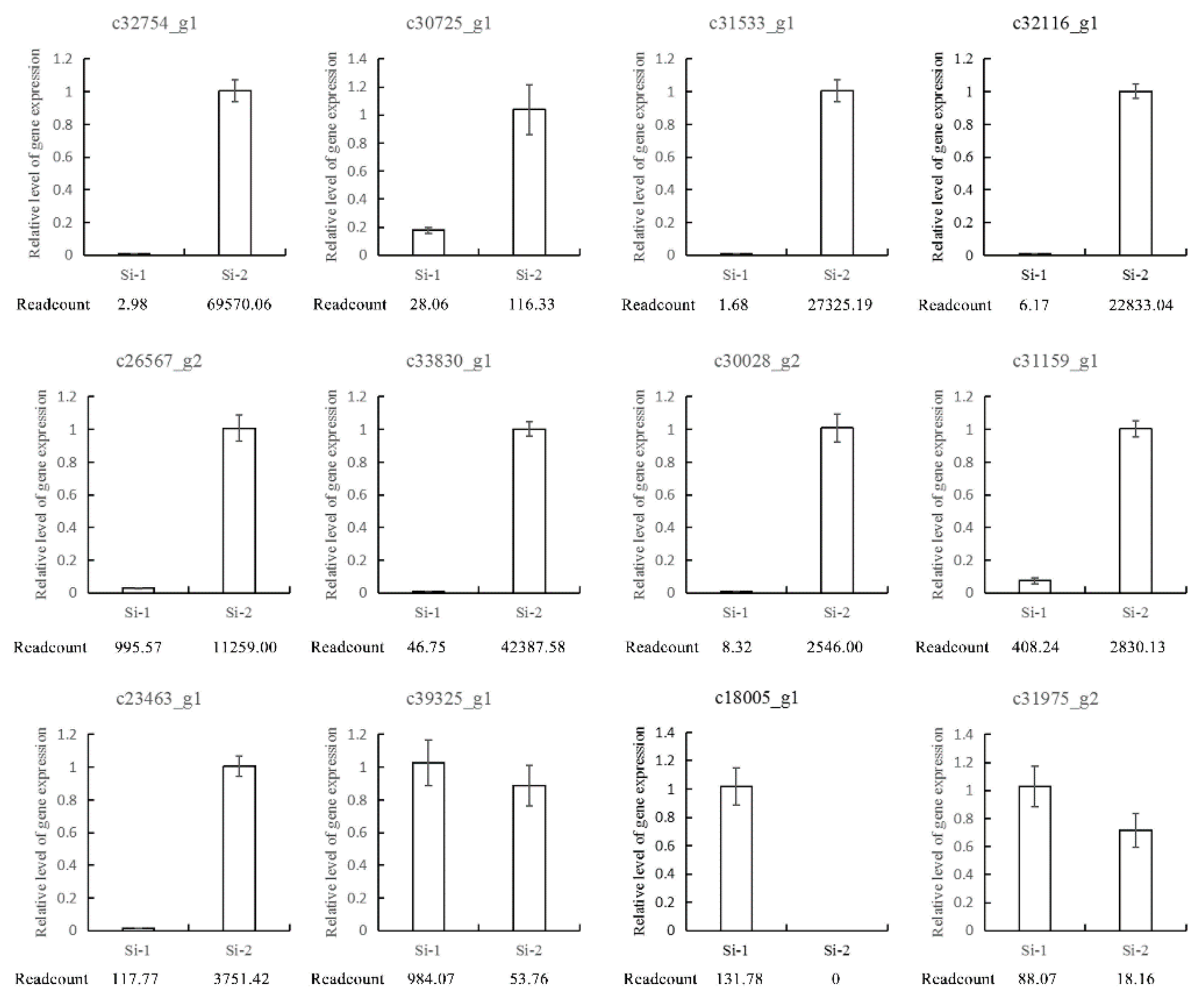
2.6. Expression Analysis of Allergic Related Genes at Different Developmental Stages
3. Discussion
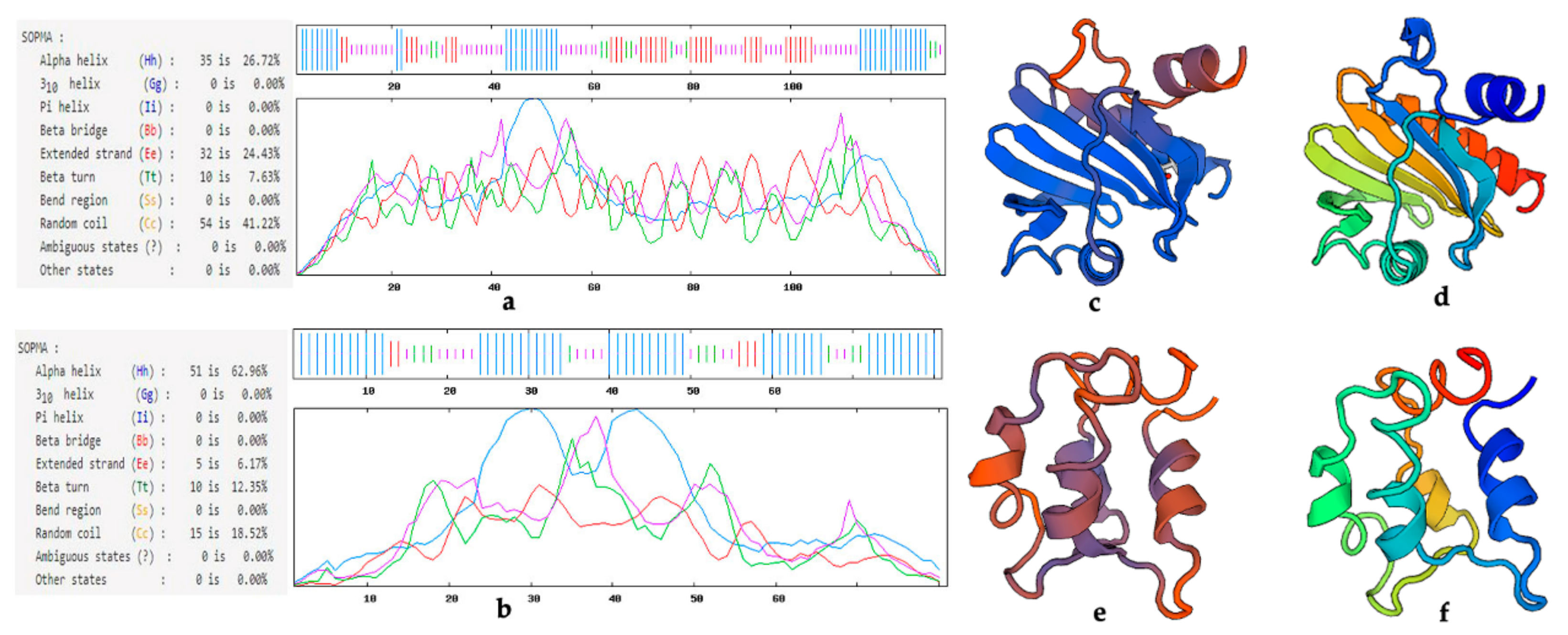
3.1. Allergenic Protein Sequence Structure Analysis
3.2. Profilin Proteins in Pollen Wall
4. Materials and Methods
4.1. Experimental Materials
4.2. Histological Observation and Sample Collection
4.3. RNA Extraction and Quality Determination
4.4. Library Preparation and Transcriptome Sequencing
4.5. Transcriptome Assembly and Functional Annotation
4.6. Gene Differential Expression Analysis
4.7. Quantitative Real-time Polymerase Chain Reaction (qRT-PCR) Analysis
4.8. Total Protein Extraction
4.9. Functional Annotation and Database Analysis
4.10. Isolation of LoProfilin and LoPolcalcin from Lily
4.11. Protein Structure Analysis
4.12. Subcellular Localization of LoProfilin Protein
4.13. Reverse Transcriptase PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| DEG | Differentially expressed gene |
| COG | Cluster of Orthologous Groups |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| GO | Gene Ontology |
| TMT | Tandem mass tag |
| PPI | Protein-protein interaction |
References
- Huang, S.; McDowell, J.M.; Weise, M.J.; Meagher, R.B. The Arabidopsis profilin gene family. Evidence for an ancient split between constitutive and pollen-specific profilin genes. Plant Physiol. 1996, 111, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.M.; Ma, S.L.; Li, W.Q.; Wang, Q.; Cao, H.Y.; Gu, J.H.; Lu, Y.M. Genetic variability and diversity of the main resources of lily assessed via phenotypic characters, pollen morphology, and ISSR markers. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef]
- Gong, B.; Yi, J.; Wu, J.; Sui, J.; Khan, M.A.; Wu, Z.; Zhong, X.; Seng, S.; He, J.; Yi, M. LlHSFA1, a novel heat stress transcription factor in lily (Lilium longiflorum), can interact with LlHSFA2 and enhance the thermotolerance of transgenic Arabidopsis thaliana. Plant Cell Rep. 2014, 33, 1519–1533. [Google Scholar] [CrossRef] [PubMed]
- Sui, J.; He, J.; Wu, J.; Gong, B.; Cao, X.; Seng, S.; Wu, Z.; Wu, C.; Liu, C.; Yi, M. Characterization and Functional Analysis of Transcription Factor LoMYB80 Related to Anther Development in Lily (Lilium Oriental Hybrids). J. Plant Growth Regul. 2015, 34, 545–557. [Google Scholar] [CrossRef]
- Albertine, J.M.; Manning, W.J.; DaCosta, M.; Stinson, K.A.; Muilenberg, M.L.; Rogers, C.A. Projected carbon dioxide to increase grass pollen and allergen exposure despite higher ozone levels. PLoS ONE 2014, 9, e111712. [Google Scholar] [CrossRef]
- Kasperska-Zajac, A.; Brzoza, Z.; Rogala, B. Seasonal Changes in Platelet Activity in Pollen-Induced Seasonal Allergic Rhinitis and Asthma. J. Asthma 2009, 45, 485–487. [Google Scholar] [CrossRef]
- Riediker, M.; Koller, T.; Monn, C. Differences in size selective aerosol sampling for pollen allergen detection using high-volume cascade impactors. Clin. Exp. Allergy 2000, 30, 867–873. [Google Scholar] [CrossRef]
- Zhang, D.; Wilson, Z.A. Stamen specification and anther development in rice. Chin. Sci. Bull. 2009, 54, 2342–2353. [Google Scholar] [CrossRef]
- Altuner, E.M.; Ceter, T.; Alpas, H. High hydrostatic pressure processing: A method having high success potential in pollen protein extraction. High Press. Res. 2012, 32, 291–298. [Google Scholar] [CrossRef]
- Barderas, R.; Villalba, M.; Pascual, C.Y.; Batanero, E.; Rodríguez, R. Profilin (Che a 2) and polcalcin (Che a 3) are relevant allergens of Chenopodium album pollen: Isolation, amino acid sequences, and immunologic properties. J. Allergy Clin. Immunol. 2004, 113, 1192–1198. [Google Scholar] [CrossRef]
- Mas, S.; Garrido-Arandia, M.; Batanero, E.; Purohit, A.; Pauli, G.; Rodríguez, R.; Barderas, R.; Villalba, M. Characterization of profilin and polcalcin panallergens from ash pollen. J. Investig. Allergol. Clin. Immunol. 2014, 24, 257–266. [Google Scholar] [PubMed]
- Hemmer, W.; Focke, M.; Wantke, F.; Götz, M.; Jarisch, R.; Jäger, S.; Götz, M. Ash (Fraxinus excelsior)-pollen allergy in central Europe: Specific role of pollen panallergens and the major allergen of ash pollen, Fra e 1. Allergy 2000, 55, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Wopfner, N.; Gadermaier, G.; Egger, M.; Asero, R.; Ebner, C.; Jahn-Schmid, B.; Ferreira, F. The Spectrum of Allergens in Ragweed and Mugwort Pollen. Int. Arch. Allergy Immunol. 2005, 138, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Valenta, R.; Duchene, M.; Pettenburger, K.; Sillaber, C.; Valent, P.; Bettelheim, P.; Breitenbach, M.; Rumpold, H.; Kraft, D.; Scheiner, O. Identification of profilin as a novel pollen allergen; IgE autoreactivity in sensitized individuals. Science 1991, 253, 557–560. [Google Scholar] [CrossRef]
- Jeong, Y.M.; Mun, J.H.; Lee, I.; Woo, J.C.; Hong, C.B.; Kim, S.G. Distinct Roles of the First Introns on the Expression of Arabidopsis Profilin Gene Family Members. Plant Physiol. 2006, 140, 196–209. [Google Scholar] [CrossRef]
- Niederberger, V.; Hayek, B.; Vrtala, S.; Laffer, S.; Twardosz, A.; Vangelista, L.; Sperr, W.R.; Valent, P.; Rumpold, H.; Kraft, D.; et al. Calcium-dependent immunoglobulin E recognition of the apo- and calcium-bound form of a cross-reactive two EF-hand timothy grass pollen allergen, Phl p 7. FASEB J. 1999, 13, 843–856. [Google Scholar] [CrossRef]
- Grote, M.; Westritschnig, K.; Valenta, R. Immunogold Electron Microscopic Localization of the 2 EF-Hand Calcium-Binding Pollen Allergen Phl p 7 and its Homologues in Pollens of Grasses, Weeds and Trees. Int. Arch. Allergy Immunol. 2008, 146, 113–121. [Google Scholar] [CrossRef]
- Park, J.W.; Ko, S.H.; Kim, C.W.; Jeoung, B.J.; Hong, C.S. Identification and characterization of the major allergen of the Humulus japonicus pollen. Clin. Exp. Allergy 1999, 29, 1080–1086. [Google Scholar] [CrossRef]
- Fu, W.; Gao, Z.; Gao, L.; Jin, J.; Liu, M.; Sun, Y.; Wu, S.; Wu, L.; Ma, H.; Dong, Y.; et al. Identification of a 62-kDa major allergen from Artemisia pollen as a putative galactose oxidase. Allergy 2018, 73, 1041–1052. [Google Scholar] [CrossRef]
- Floch, V.B.L.; Le Mignon, M.; Bouley, J.; Groeme, R.; Jain, K.; Baron-Bodo, V.; Nony, E.; Mascarell, L.; Moingeon, P. Identification of Novel Short Ragweed Pollen Allergens Using Combined Transcriptomic and Immunoproteomic Approaches. PLoS ONE 2015, 10, e0136258. [Google Scholar]
- Radauer, C.; Breiteneder, H. Evolutionary biology of plant food allergens. J. Allergy Clin. Immunol. 2007, 120, 518–525. [Google Scholar] [CrossRef]
- Radauer, C.; Willerroider, M.; Fuchs, H.; Thalhamer, J.; Ferreira, F.; Scheiner, O.; Breiteneder, H.; Hoffmann-Sommergruber, K.; Hoffmann-Sommergruber, K. Cross-reactive and species-specific immunoglobulin E epitopes of plant profilins: An experimental and structure-based analysis. Clin. Exp. Allergy 2006, 36, 920–929. [Google Scholar] [CrossRef]
- Chan, T.F.; Ji, K.M.; Yim, A.K.Y.; Liu, X.Y.; Zhou, J.W.; Li, R.Q.; Yang, K.Y.; Li, J.; Li, M.; Law, P.T.W.; et al. The draft genome, transcriptome, and microbiome of Dermatophagoides farinae reveal a broad spectrum of dust mite allergens. J. Allergy Clin. Immunol. 2015, 135, 539–548. [Google Scholar] [CrossRef]
- Yim, S.; Khare, D.; Kang, J.; Hwang, J.U.; Liang, W.; Martinoia, E.; Zhang, D.; Kang, B.; Lee, Y. Postmeiotic development of pollen surface layers requires two Arabidopsis ABCG-type transporters. Plant Cell Rep. 2016, 35, 1863–1873. [Google Scholar] [CrossRef]
- Aya, K.; Ueguchi-Tanaka, M.; Kondo, M.; Hamada, K.; Yano, K.; Nishimura, M.; Matsuoka, M. Gibberellin Modulates Anther Development in Rice via the Transcriptional Regulation of GAMYB. Plant Cell 2009, 21, 1453–1472. [Google Scholar] [CrossRef]
- Xiong, S.X.; Lu, J.Y.; Lou, Y.; Teng, X.D.; Gu, J.N.; Zhang, C.; Shi, Q.S.; Yang, Z.N.; Zhu, J. The transcription factors MS188 and AMS form a complex to activate the expression of CYP703A2 for sporopollenin biosynthesis in Arabidopsis thaliana. Plant J. 2016, 88, 936–946. [Google Scholar] [CrossRef]
- Dobritsa, A.A.; Shrestha, J.; Morant, M.; Pinot, F.; Matsuno, M.; Swanson, R.; Møller, B.L.; Preuss, D. CYP704B1 Is a Long-Chain Fatty Acid v- Hydroxylase Essential for Sporopollenin Synthesis in Pollen of Arabidopsis. Plant Physiol. 2009, 151, 574–589. [Google Scholar] [CrossRef]
- Blackmore, S.; Wortley, A.H.; Skvarla, J.J.; Rowley, J.R. Pollen wall development in flowering plants. New Phytol. 2007, 174, 483–498. [Google Scholar] [CrossRef]
- Scott, R.J. Stamen Structure and Function. Plant Cell Online 2004, 16 (Suppl. 1), S46–S60. [Google Scholar] [CrossRef]
- Meuter-Gerhards, A.; Riegert, S.; Wiermann, R. Studies on Sporopollenin Biosynthesis in Cucurbita maxima (OUCH.)- II. The Involvement of Aliphatic Metabolism. J. Plant Physiol. 1999, 154, 431–436. [Google Scholar] [CrossRef]
- Zinkl, G.M.; Zwiebel, B.I.; Grier, D.G.; Preuss, D. Pollen-stigma adhesion in Arabidopsis: A species-specific interaction mediated by lipophilic molecules in the pollen exine. Development 1999, 126, 5431–5440. [Google Scholar]
- Yang, R.B.; Liu, L. Research Progress of Pollen Allergy. Chin. Agric. Sci. Bull. 2015, 31, 163–167. (In Chinese) [Google Scholar]
- Luoto, S.; Lambert, W.; Blomqvist, A.; Emanuelsson, C. The identification of allergen proteins in sugar beet (Beta vulgaris) pollen causing occupational allergy in greenhouses. Clin. Mol. Allergy 2008, 6, 7. [Google Scholar] [CrossRef]
- Karakesisoglou, I.; Schleicher, M.; Gibbon, B.C.; Staiger, C.J. Plant profilins rescue the aberrant phenotype of profilin-deficient Dictyostelium cells. Cell Motil. Cytoskelet. 1996, 34, 36–47. [Google Scholar] [CrossRef]
- Martínez, A.; Asturias, J.A.; Monteseirín, J.; Moreno, V.; García-Cubillana, A.; Hernández, M.; De La Calle, A.; Sánchez-Hernández, C.; Pérez-Formoso, J.L.; Conde, J. The allergenic relevance of profilin (Ole e 2) from Olea europaea pollen. Allergy 2002, 57, 17–23. [Google Scholar] [CrossRef]
- Offermann, L.R.; Schlachter, C.R.; Perdue, M.L.; Majorek, K.A.; He, J.Z.; Booth, W.T.; Garrett, J.; Kowal, K.; Chruszcz, M. Structural, Functional, and Immunological Characterization of Profilin Panallergens Amb a 8, Art v 4, and Bet v 2. J. Biol. Chem. 2016, 291, 15447–15459. [Google Scholar] [CrossRef]
- Liu, Q.; Guo, Z. Molecular cloning and characterization of a profilin gene BnPFN from Brassica nigra that expressing in a pollen-specific manner. Mol. Biol. Rep. 2009, 36, 135–139. [Google Scholar] [CrossRef]
- Verdino, P.; Westritschnig, K.; Valenta, R.; Keller, W. The cross-reactive calcium-binding pollen allergen, Phl p 7, reveals a novel dimer assembly. EMBO J. 2002, 21, 5007–5016. [Google Scholar] [CrossRef]
- Christensen, H.E.; Ramachandran, S.; Tan, C.T.; Surana, U.; Dong, C.H.; Chua, N.H. Arabidopsis profilins are functionally similar to yeast profilins: Identification of a vascular bundle-specific profilin and a pollen-specific profilin. Plant J. 1996, 10, 269–279. [Google Scholar] [CrossRef]
- Staiger, C.J.; Goodbody, K.C.; Hussey, P.J.; Valenta, R.; Drøbak, B.K.; Lloyd, C.W. The profilin multigene family of maize: Differential expression of three isoforms. Plant J. 1993, 4, 631–641. [Google Scholar] [CrossRef]
- Mittermann, I.; Swoboda, I.; Pierson, E.; Eller, N.; Kraft, D.; Valenta, R.; Heberle-Bors, E. Molecular cloning and characterization of profilin from tobacco (Nicotiana tabacum): Increased profilin expression during pollen maturation. Plant Mol. Boil. 1995, 27, 137–146. [Google Scholar] [CrossRef]
- Valenta, R.; Ferreira, F.; Grote, M.; Swoboda, I.; Vrtala, S.; Duchêne, M.; Deviller, P.; Meagher, R.B.; McKinney, E.; Heberle-Bors, E. Identification of profilin as an actin-binding protein in higher plants. J. Boil. Chem. 1993, 268, 22777. [Google Scholar]
- Hussey, P.J.; Ketelaar, T.; Deeks, M.J. Control of the Actin Cytoskeleton in Plant Cell Growth. Annu. Rev. Plant Boil. 2006, 57, 109–125. [Google Scholar] [CrossRef]
- Huang, H. Pharmaceutical Cell Biology; China Medical Science and Technology Press: Beijing, China, 2006; pp. 311–312. Available online: http://ebook.njau.edu.cn:8082/Jpath_sky/DsrPath.do?code=E583827EE89C183ECF8E4FB211059A01&ssnumber=11600942&netuser=1&jpgreadmulu=1&displaystyle=0&channel=0&ipside=0&pages=17 (accessed on 22 November 2019).
- Kandasamy, M.K.; McKinney, E.C.; Meagher, R.B. Plant profilin isovariants are distinctly regulated in vegetative and reproductive tissues. Cell Motil. Cytoskelet. 2002, 52, 22–32. [Google Scholar] [CrossRef]
- Rothkegel, M.; Mayboroda, O.; Rohde, M.; Wucherpfennig, C.; Valenta, R.; Jockusch, B.M. Plant and animal profilins are functionally equivalent and stabilize microfilaments in living animal cells. J. Cell Sci. 1996, 109, 83–90. [Google Scholar]
- von Witsch, M.; Baluška, F.; Staiger, C.J.; Volkmann, D. Profilin is associated with the plasma membrane in microspores and pollen. Eur. J. Cell Biol. 1998, 77, 303–312. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Dekkers, B.J.; Willems, L.; Bassel, G.W.; van Bolderen-Veldkamp, R.P.; Ligterink, W.; Hilhorst, H.W.; Bentsink, L. Identification of Reference Genes for RT-qPCR Expression Analysis in Arabidopsis and Tomato Seeds. Plant Cell Physiol. 2012, 53, 28–37. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pollen Development Stage | Size of Bud (mm) | Size of Anther (mm) | Coler of Anther | Color of Pollen |
|---|---|---|---|---|
| Sporulation stage | <36 | <17 | Light green | Transparent |
| Mother cell stage | 36–43 | 17–24 | Light yellow | Light yellow |
| Meiosis stage | 43–53 | 24–29 | Light yellow | Light yellow |
| Tetrad stage | 54–56 | 29–32 | Light yellow | Light yellow |
| Early mononuclear stage | 56–61 | 32–35 | Yellow | Yellow/Dark yellow |
| Mature pollen stage | >61 | >35 | Orange | Orange |
| Sample Name | Raw Reads | Clean Reads | Clean Bases | Q30 1 | Clean Reads Ratio | GC 2 | Total Number of Unigenes | Total Length of Unigenes (bp) |
|---|---|---|---|---|---|---|---|---|
| Si-1 | 65,663,717 | 62,398,416 | 9.36 | 92.56 | 95.03 | 50.81 | 62,531,354 | |
| Si-2 | 67,241,165 | 64,561,039 | 9.68 | 92.85 | 96.01 | 51.72 |
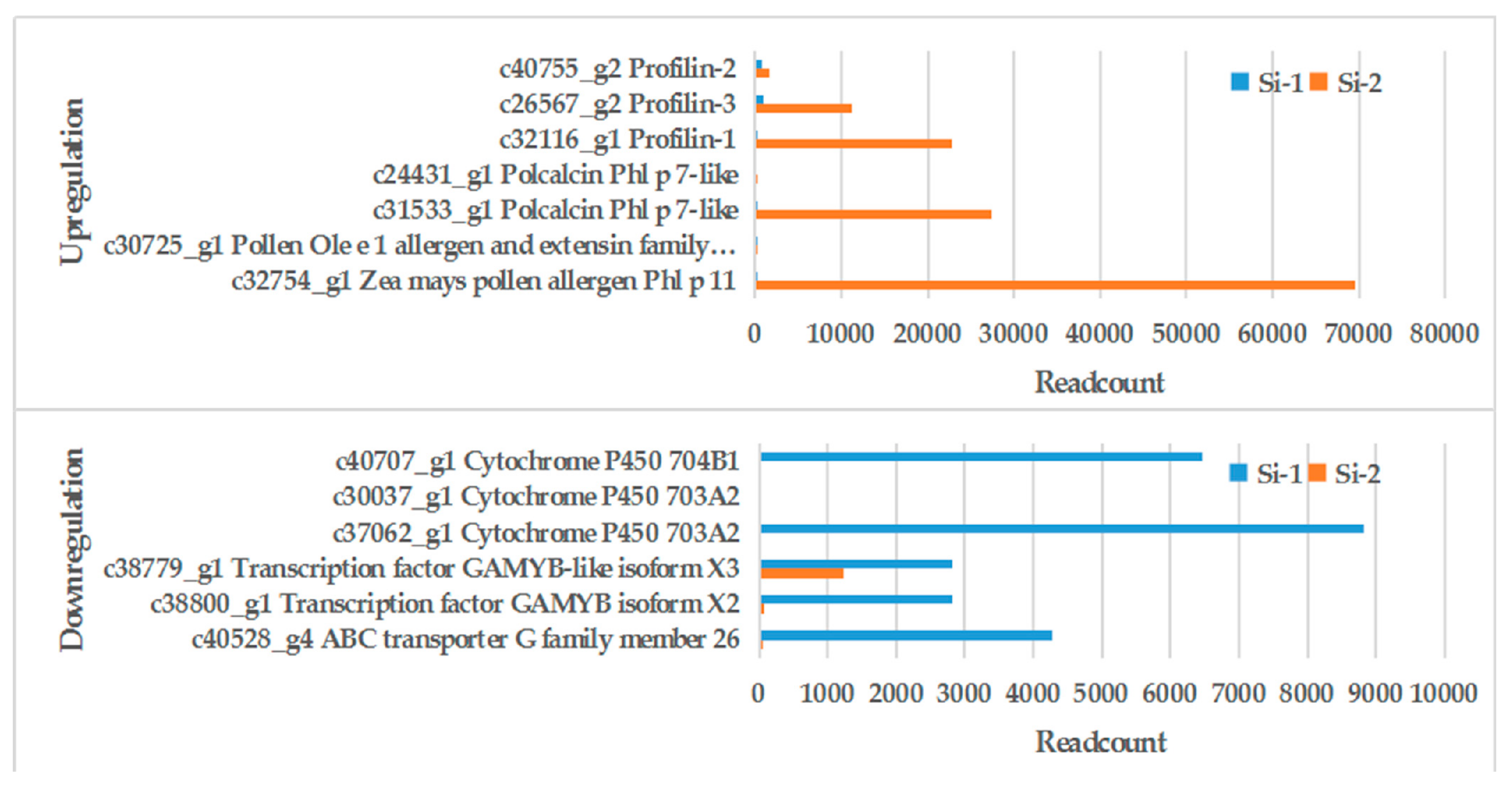
| Gene ID | Si-1 Readcount 1 | Si-2 Readcount 2 | Log2FC 3 (Si-2 vs Si-1) | Up/Down | Annotation |
|---|---|---|---|---|---|
| c40528_g4 | 4272.08 | 18.92 | −7.82 | Down | ABC transporter G family member 26 |
| c38800_g1 | 2816.89 | 64.91 | −5.44 | Down | transcription factor GAMYB isoform X2 |
| c38779_g1 | 2810.16 | 1233.06 | −1.19 | Down | transcription factor GAMYB-like isoform X3 |
| c37062_g1 | 8832.13 | 0 | -Inf | Down | cytochrome P450 703A2 |
| c30037_g1 | 9.48 | 0 | -Inf | Down | cytochrome P450 703A2 |
| c40707_g1 | 6459.93 | 0 | -Inf | Down | cytochrome P450 704B1 |
| c32754_g1 | 2.98 | 69,570.06 | 14.51 | Up | Zea mays pollen allergen Phl p 11 |
| c30725_g1 | 28.06 | 116.33 | 2.05 | Up | pollen Ole e 1 allergen and extensin family protein |
| c31533_g1 | 1.68 | 27,325.19 | 13.99 | Up | polcalcin Phl p 7-like |
| c24431_g1 | 0 | 39.70 | Inf | Up | polcalcin Phl p 7-like |
| c32116_g1 | 6.17 | 22,833.04 | 11.85 | Up | Profilin-1 |
| c26567_g2 | 995.57 | 11,259.00 | 3.50 | Up | Profilin-3 |
| c40755_g2 | 870.39 | 1572.53 | 0.85 | Up | Profilin-2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, J.; Wu, Z.; Wang, X.; Zhang, Y.; Teng, N. Analysis of Pollen Allergens in Lily by Transcriptome and Proteome Data. Int. J. Mol. Sci. 2019, 20, 5892. https://doi.org/10.3390/ijms20235892
Feng J, Wu Z, Wang X, Zhang Y, Teng N. Analysis of Pollen Allergens in Lily by Transcriptome and Proteome Data. International Journal of Molecular Sciences. 2019; 20(23):5892. https://doi.org/10.3390/ijms20235892
Chicago/Turabian StyleFeng, Jingxian, Ze Wu, Xueqian Wang, Yaming Zhang, and Nianjun Teng. 2019. "Analysis of Pollen Allergens in Lily by Transcriptome and Proteome Data" International Journal of Molecular Sciences 20, no. 23: 5892. https://doi.org/10.3390/ijms20235892
APA StyleFeng, J., Wu, Z., Wang, X., Zhang, Y., & Teng, N. (2019). Analysis of Pollen Allergens in Lily by Transcriptome and Proteome Data. International Journal of Molecular Sciences, 20(23), 5892. https://doi.org/10.3390/ijms20235892