HSF1Base: A Comprehensive Database of HSF1 (Heat Shock Factor 1) Target Genes


Abstract
1. Introduction
2. Results
2.1. The HSF1base Database
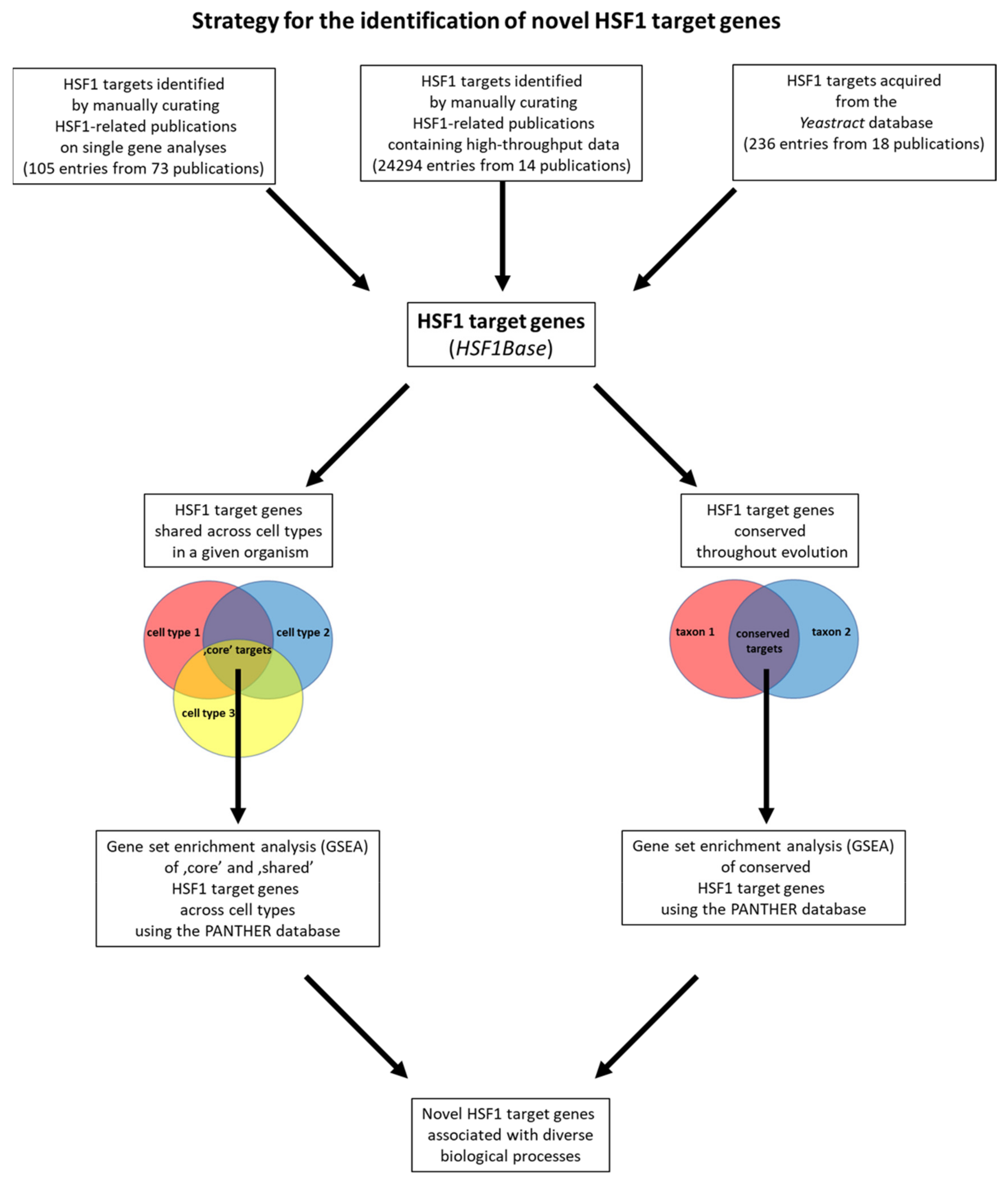
2.2. Strategy for Testing the Applicability of the HSF1Base
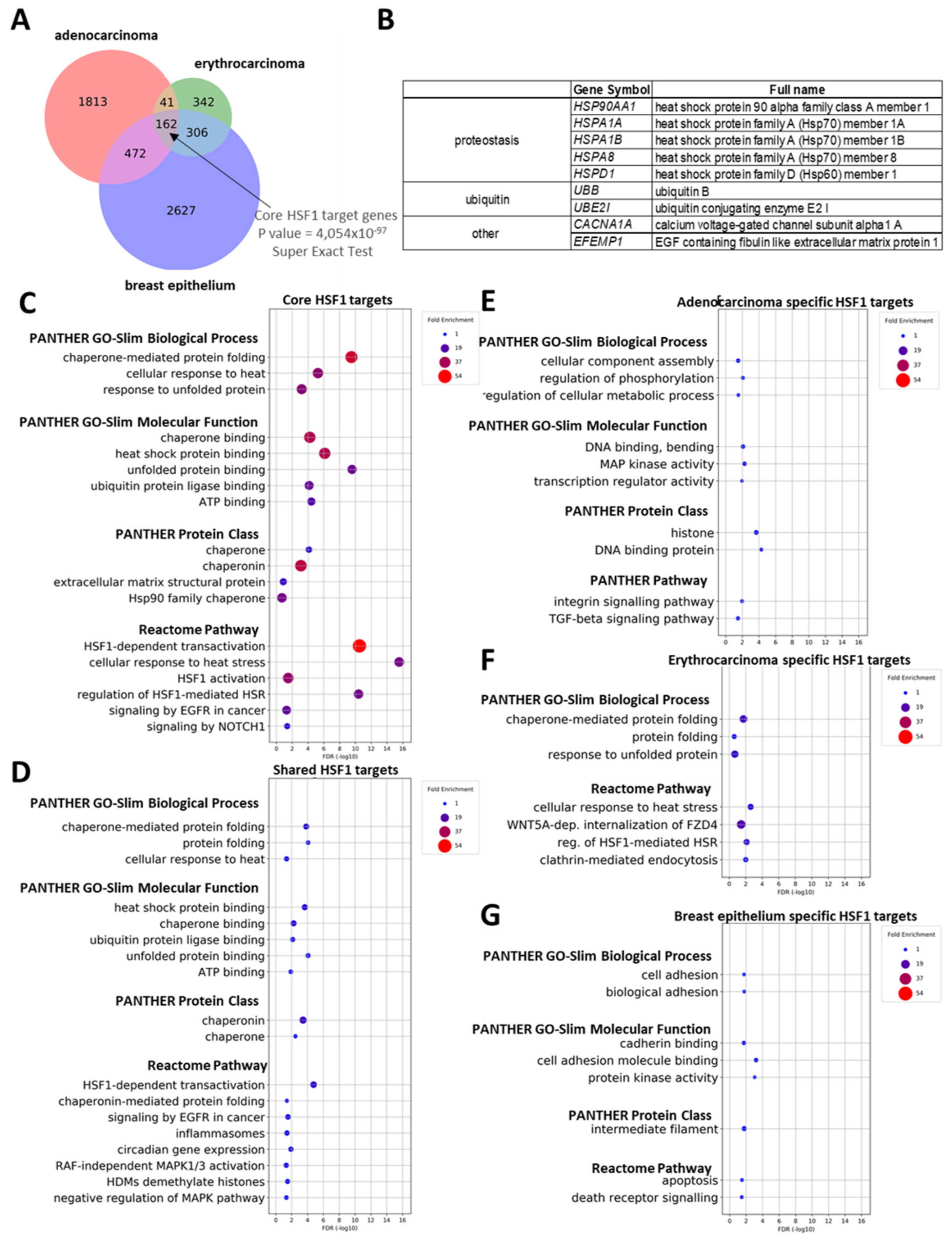
2.3. General and Tissue-Specific HSF1 Target Genes in Human
2.4. Cell Type-Specific and General Ageing-Related HSF1 Target Genes in Human
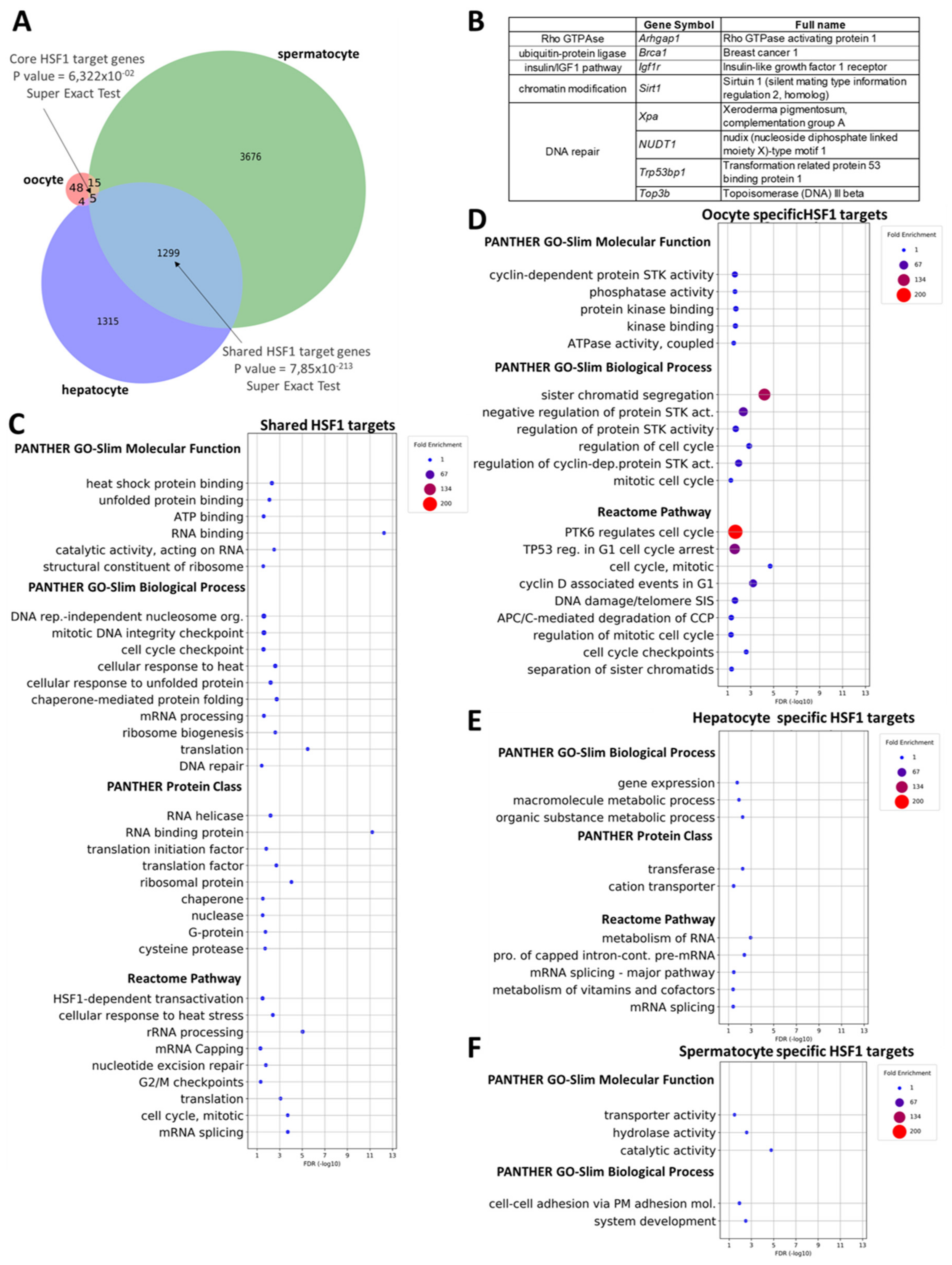
2.5. ‘Shared’ and Cell Type-Specific HSF1 Target Genes in Mice
2.6. Identifying HSF1 Target Genes Related to Ageing in Mouse
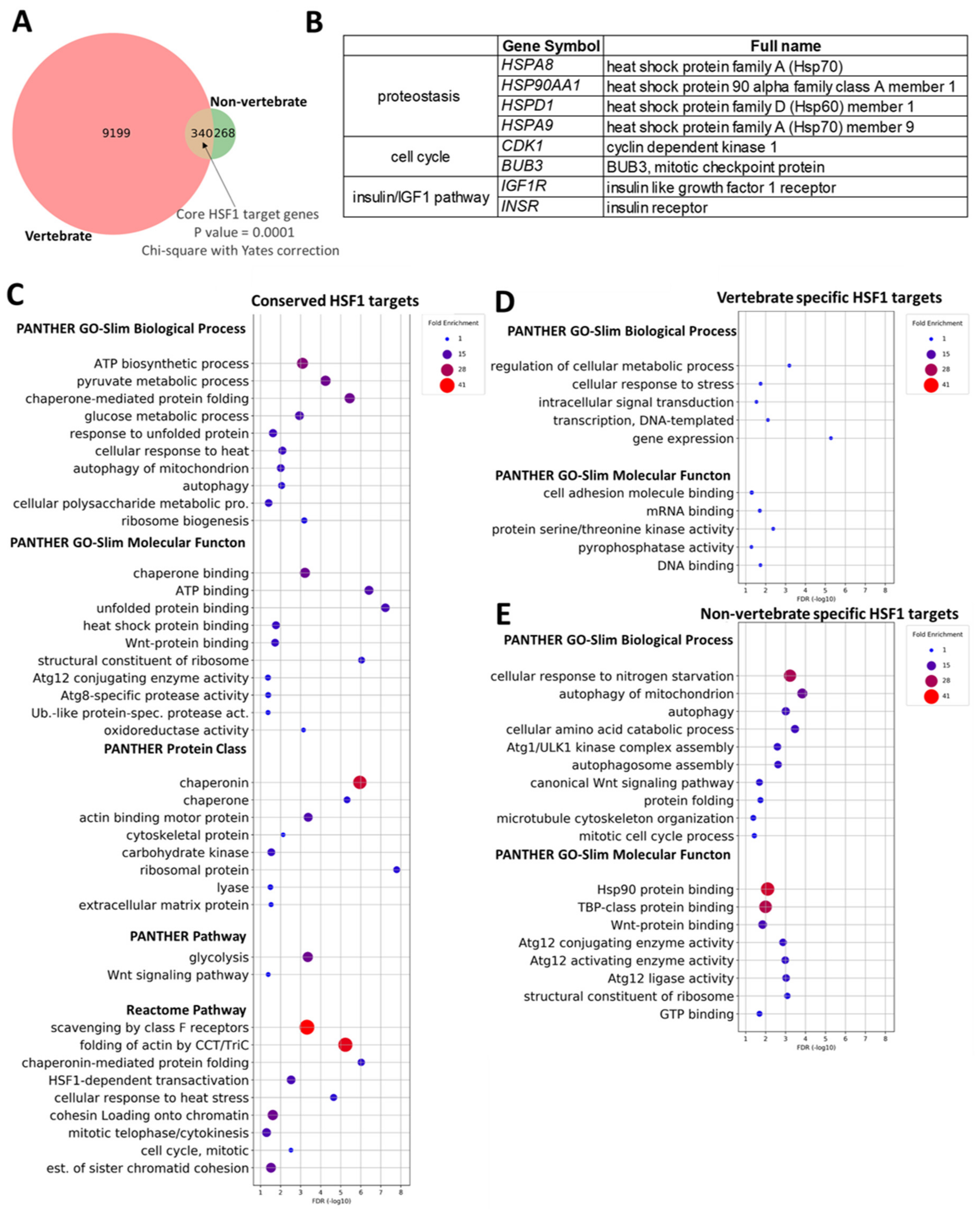
2.7. Novel Conserved HSF1 Target Genes
2.8. Ageing-Related HSF1 Target Genes Conserved Throughout Evolution
3. Discussion
4. Materials and Methods
4.1. Origin of Datasets Used
4.2. Statistical Overrepresentation Tests
4.3. Orthologue Analysis
4.4. Venn Diagrams and Statistical Analysis of Overlaps
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lindquist, S.; Craig, E.A. The heat-shock proteins. Annu. Rev. Genet. 1988, 22, 631–677. [Google Scholar] [CrossRef] [PubMed]
- Akerfelt, M.; Morimoto, R.I.; Sistonen, L. Heat shock factors: Integrators of cell stress, development and lifespan. Nat. Rev. Mol. Cell Biol. 2010, 11, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Richter, K.; Haslbeck, M.; Buchner, J. The heat shock response: Life on the verge of death. Mol. Cell 2010, 40, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, R.I. Cells in stress: Transcriptional activation of heat shock genes. Science 1993, 259, 1409–1410. [Google Scholar] [CrossRef]
- Gomez-Pastor, R.; Burchfiel, E.T.; Thiele, D.J. Regulation of heat shock transcription factors and their roles in physiology and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 4–20. [Google Scholar] [CrossRef]
- Neef, D.W.; Jaeger, A.M.; Thiele, D.J. Heat shock transcription factor 1 as a therapeutic target in neurodegenerative diseases. Nat. Rev. Drug Discov. 2011, 10, 930–944. [Google Scholar] [CrossRef]
- Amin, J.; Ananthan, J.; Voellmy, R. Key features of heat shock regulatory elements. Mol. Cell. Biol. 1988, 8, 3761–3769. [Google Scholar] [CrossRef]
- Wu, C. Heat shock transcription factors: Structure and regulation. Annu. Rev. Cell Dev. Biol. 1995, 11, 441–469. [Google Scholar] [CrossRef]
- Brunquell, J.; Morris, S.; Lu, Y.; Cheng, F.; Westerheide, S.D. The genome-wide role of HSF-1 in the regulation of gene expression in Caenorhabditis elegans. BMC Genom. 2016, 17, 559. [Google Scholar] [CrossRef]
- Hahn, J.-S.; Hu, Z.; Thiele, D.J.; Iyer, V.R. Genome-wide analysis of the biology of stress responses through heat shock transcription factor. Mol. Cell. Biol. 2004, 24, 5249–5256. [Google Scholar] [CrossRef]
- Birch-Machin, I.; Gao, S.; Huen, D.; McGirr, R.; White, R.A.H.; Russell, S. Genomic analysis of heat-shock factor targets in Drosophila. Genome Biol. 2005, 6, R63. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chauve, L.; Phelps, G.; Brielmann, R.M.; Morimoto, R.I. E2F coregulates an essential HSF developmental program that is distinct from the heat-shock response. Genes Dev. 2016, 30, 2062–2075. [Google Scholar] [CrossRef] [PubMed]
- Mahat, D.B.; Salamanca, H.H.; Duarte, F.M.; Danko, C.G.; Lis, J.T. Mammalian Heat Shock Response and Mechanisms Underlying Its Genome-wide Transcriptional Regulation. Mol. Cell 2016, 62, 63–78. [Google Scholar] [CrossRef] [PubMed]
- Trinklein, N.D.; Murray, J.I.; Hartman, S.J.; Botstein, D.; Myers, R.M. The role of heat shock transcription factor 1 in the genome-wide regulation of the mammalian heat shock response. Mol. Biol. Cell 2004, 15, 1254–1261. [Google Scholar] [CrossRef]
- Le Masson, F.; Razak, Z.; Kaigo, M.; Audouard, C.; Charry, C.; Cooke, H.; Westwood, J.T.; Christians, E.S. Identification of heat shock factor 1 molecular and cellular targets during embryonic and adult female meiosis. Mol. Cell. Biol. 2011, 31, 3410–3423. [Google Scholar] [CrossRef]
- Mendillo, M.L.; Santagata, S.; Koeva, M.; Bell, G.W.; Hu, R.; Tamimi, R.M.; Fraenkel, E.; Ince, T.A.; Whitesell, L.; Lindquist, S. HSF1 drives a transcriptional program distinct from heat shock to support highly malignant human cancers. Cell 2012, 150, 549–562. [Google Scholar] [CrossRef]
- Barna, J.; Csermely, P.; Vellai, T. Roles of heat shock factor 1 beyond the heat shock response. Cell. Mol. Life Sci. 2018, 75, 2897–2916. [Google Scholar] [CrossRef]
- Li, J.; Labbadia, J.; Morimoto, R.I. Rethinking HSF1 in stress, development, and organismal health. Trends Cell Biol. 2017, 27, 895–905. [Google Scholar] [CrossRef]
- Santagata, S.; Hu, R.; Lin, N.U.; Mendillo, M.L.; Collins, L.C.; Hankinson, S.E.; Schnitt, S.J.; Whitesell, L.; Tamimi, R.M.; Lindquist, S.; et al. High levels of nuclear heat-shock factor 1 (HSF1) are associated with poor prognosis in breast cancer. Proc. Natl. Acad. Sci. USA 2011, 108, 18378–18383. [Google Scholar] [CrossRef]
- Dai, C.; Whitesell, L.; Rogers, A.B.; Lindquist, S. Heat shock factor 1 is a powerful multifaceted modifier of carcinogenesis. Cell 2007, 130, 1005–1018. [Google Scholar] [CrossRef]
- Chatterjee, S.; Burns, T.F. Targeting heat shock proteins in cancer: A promising therapeutic approach. Int. J. Mol. Sci. 2017, 18, 1978. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.M.R.; Cunha-Oliveira, T.; Sobral, M.C.; Abreu, P.L.; Alpoim, M.C.; Urbano, A.M. Impact of carcinogenic chromium on the cellular response to proteotoxic stress. Int. J. Mol. Sci. 2019, 20, 4901. [Google Scholar] [CrossRef] [PubMed]
- Carnemolla, A.; Labbadia, J.P.; Lazell, H.; Neueder, A.; Moussaoui, S.; Bates, G.P. Contesting the dogma of an age-related heat shock response impairment; implications for cardiac-specific age-related disorders. Hum. Mol. Genet. 2014, 23, 3641–3656. [Google Scholar] [CrossRef] [PubMed]
- Perfetto, L.; Acencio, M.L.; Bradley, G.; Cesareni, G.; Del Toro, N.; Fazekas, D.; Hermjakob, H.; Korcsmaros, T.; Kuiper, M.; Lægreid, A.; et al. CausalTAB: The PSI-MITAB 2.8 updated format for signalling data representation and dissemination. Bioinformatics 2019, 35, 3779–3785. [Google Scholar] [CrossRef]
- Webb, A.E.; Kundaje, A.; Brunet, A. Characterization of the direct targets of FOXO transcription factors throughout evolution. Aging Cell 2016, 15, 673–685. [Google Scholar] [CrossRef]
- Mi, H.; Muruganujan, A.; Thomas, P.D. PANTHER in 2013: Modeling the evolution of gene function, and other gene attributes, in the context of phylogenetic trees. Nucleic Acids Res. 2013, 41, D377–D386. [Google Scholar] [CrossRef]
- Das, S.; Bhattacharyya, N.P. Transcription regulation of HYPK by Heat Shock Factor. PLoS ONE 2014, 9, e85552. [Google Scholar] [CrossRef]
- Song, S.; Kole, S.; Precht, P.; Pazin, M.J.; Bernier, M. Activation of heat shock factor 1 plays a role in pyrrolidine dithiocarbamate-mediated expression of the co-chaperone BAG. Int. J. Biochem. Cell Biol. 2010, 42, 1856–1863. [Google Scholar] [CrossRef]
- Sawai, M.; Ishikawa, Y.; Ota, A.; Sakurai, H. The proto-oncogene JUN is a target of the heat shock transcription factor HSF1. FEBS J. 2013, 280, 6672–6680. [Google Scholar] [CrossRef]
- Lecomte, S.; Desmots, F.; Le Masson, F.; Le Goff, P.; Michel, D.; Christians, E.S.; Le Dréan, Y. Roles of heat shock factor 1 and 2 in response to proteasome inhibition: Consequence on p53 stability. Oncogene 2010, 29, 4216–4224. [Google Scholar] [CrossRef]
- Benderska, N.; Ivanovska, J.; Rau, T.T.; Schulze-Luehrmann, J.; Mohan, S.; Chakilam, S.; Gandesiri, M.; Ziesché, E.; Fischer, T.; Söder, S.; et al. DAPK-HSF1 interaction as a positive-feedback mechanism stimulating TNF-induced apoptosis in colorectal cancer cells. J. Cell Sci. 2014, 127, 5273–5287. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, A.T.; Marnett, L.J. HSF1-mediated BAG3 expression attenuates apoptosis in 4-hydroxynonenal-treated colon cancer cells via stabilization of anti-apoptotic Bcl-2 proteins. J. Biol. Chem. 2009, 284, 9176–9183. [Google Scholar] [CrossRef] [PubMed]
- Logan, I.R.; McNeill, H.V.; Cook, S.; Lu, X.; Meek, D.W.; Fuller-Pace, F.V.; Lunec, J.; Robson, C.N. Heat shock factor-1 modulates p53 activity in the transcriptional response to DNA damage. Nucleic Acids Res. 2009, 37, 2962–2973. [Google Scholar] [CrossRef] [PubMed]
- Tacutu, R.; Craig, T.; Budovsky, A.; Wuttke, D.; Lehmann, G.; Taranukha, D.; Costa, J.; Fraifeld, V.E.; de Magalhães, J.P. Human Ageing Genomic Resources: Integrated databases and tools for the biology and genetics of ageing. Nucleic Acids Res. 2013, 41, D1027–D1033. [Google Scholar] [CrossRef] [PubMed]
- Korfanty, J.; Stokowy, T.; Widlak, P.; Gogler-Piglowska, A.; Handschuh, L.; Podkowiński, J.; Vydra, N.; Naumowicz, A.; Toma-Jonik, A.; Widlak, W. Crosstalk between HSF1 and HSF2 during the heat shock response in mouse testes. Int. J. Biochem. Cell Biol. 2014, 57, 76–83. [Google Scholar] [CrossRef]
- Vihervaara, A.; Sistonen, L. HSF1 at a glance. J. Cell Sci. 2014, 127, 261–266. [Google Scholar] [CrossRef]
- Akerfelt, M.; Vihervaara, A.; Laiho, A.; Conter, A.; Christians, E.S.; Sistonen, L.; Henriksson, E. Heat shock transcription factor 1 localizes to sex chromatin during meiotic repression. J. Biol. Chem. 2010, 285, 34469–34476. [Google Scholar] [CrossRef]
- Vihervaara, A.; Sergelius, C.; Vasara, J.; Blom, M.A.H.; Elsing, A.N.; Roos-Mattjus, P.; Sistonen, L. Transcriptional response to stress in the dynamic chromatin environment of cycling and mitotic cells. Proc. Natl. Acad. Sci. USA 2013, 110, E3388–E3397. [Google Scholar] [CrossRef]
- Barna, J.; Princz, A.; Kosztelnik, M.; Hargitai, B.; Takács-Vellai, K.; Vellai, T. Heat shock factor-1 intertwines insulin/IGF-1, TGF-β and cGMP signalling to control development and ageing. BMC Dev. Biol. 2012, 12, 32. [Google Scholar] [CrossRef]
- Inouye, S.; Izu, H.; Takaki, E.; Suzuki, H.; Shirai, M.; Yokota, Y.; Ichikawa, H.; Fujimoto, M.; Nakai, A. Impaired IgG production in mice deficient for heat shock transcription factor. J. Biol. Chem. 2004, 279, 38701–38709. [Google Scholar] [CrossRef]
- Singh, I.S.; He, J.-R.; Calderwood, S.; Hasday, J.D. A high affinity HSF-1 binding site in the 5′-untranslated region of the murine tumor necrosis factor-alpha gene is a transcriptional repressor. J. Biol. Chem. 2002, 277, 4981–4988. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Zhong, R.; Chen, C.; Calderwood, S.K. Heat shock factor 1 contains two functional domains that mediate transcriptional repression of the c-fos and c-fms genes. J. Biol. Chem. 2003, 278, 4687–4698. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; He, H.; Yu, L.; Xia, H.H.-X.; Lin, M.C.M.; Gu, Q.; Li, M.; Zou, B.; An, X.; Jiang, B.; et al. HSF1 down-regulates XAF1 through transcriptional regulation. J. Biol. Chem. 2006, 281, 2451–2459. [Google Scholar] [CrossRef] [PubMed]
- Inouye, S.; Fujimoto, M.; Nakamura, T.; Takaki, E.; Hayashida, N.; Hai, T.; Nakai, A. Heat shock transcription factor 1 opens chromatin structure of interleukin-6 promoter to facilitate binding of an activator or a repressor. J. Biol. Chem. 2007, 282, 33210–33217. [Google Scholar] [CrossRef] [PubMed]
- Tamaru, T.; Hattori, M.; Honda, K.; Benjamin, I.; Ozawa, T.; Takamatsu, K. Synchronization of circadian Per2 rhythms and HSF1-BMAL1: CLOCK interaction in mouse fibroblasts after short-term heat shock pulse. PLoS ONE 2011, 6, e24521. [Google Scholar] [CrossRef] [PubMed]
- Tamaru, T.; Hattori, M.; Ninomiya, Y.; Kawamura, G.; Varès, G.; Honda, K.; Mishra, D.P.; Wang, B.; Benjamin, I.; Sassone-Corsi, P.; et al. ROS stress resets circadian clocks to coordinate pro-survival signals. PLoS ONE 2013, 8, e82006. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, G.; Hattori, M.; Takamatsu, K.; Tsukada, T.; Ninomiya, Y.; Benjamin, I.; Sassone-Corsi, P.; Ozawa, T.; Tamaru, T. Cooperative interaction among BMAL1, HSF1, and p53 protects mammalian cells from UV stress. Commun. Biol. 2018, 1, 204. [Google Scholar] [CrossRef]
- Park, S.-J.; Sohn, H.-Y.; Park, S.I. TRAIL regulates collagen production through HSF1-dependent Hsp47 expression in activated hepatic stellate cells. Cell Signal. 2013, 25, 1635–1643. [Google Scholar] [CrossRef]
- Fritah, S.; Col, E.; Boyault, C.; Govin, J.; Sadoul, K.; Chiocca, S.; Christians, E.; Khochbin, S.; Jolly, C.; Vourc’h, C. Heat-shock factor 1 controls genome-wide acetylation in heat-shocked cells. Mol. Biol. Cell 2009, 20, 4976–4984. [Google Scholar] [CrossRef]
- Jones, M.; Wales, C.T.K.; Gristock, R.A.; Jacobs, A.T. Regulation of extracellular matrix gene expression and biosynthetic pathways in pulmonary fibroblasts by HSF1. FASEB J. 2017, 31, 6311. [Google Scholar]
- Dhanani, K.C.H.; Samson, W.J.; Edkins, A.L. Fibronectin is a stress responsive gene regulated by HSF1 in response to geldanamycin. Sci. Rep. 2017, 7, 17617. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Sun, L.; Qin, X.; Liu, T.; Zhang, S.; Liu, Y.; Li, S.; Guo, K. HSF1 promotes the inhibition of EMT-associated migration by low glucose via directly regulating Snail1 expression in HCC cells. Discov. Med. 2016, 22, 87–96. [Google Scholar] [PubMed]
- Dai, W.; Ye, J.; Zhang, Z.; Yang, L.; Ren, H.; Wu, H.; Chen, J.; Ma, J.; Zhai, E.; Cai, S.; et al. Increased expression of heat shock factor 1 (HSF1) is associated with poor survival in gastric cancer patients. Diagn. Pathol. 2018, 13, 80. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Mruk, D.D.; Cheng, C.Y. Intercellular adhesion molecules (ICAMs) and spermatogenesis. Hum. Reprod. Update 2013, 19, 167–186. [Google Scholar] [CrossRef] [PubMed]
- Young, T.R.; Leamey, C.A. Teneurins: Important regulators of neural circuitry. Int. J. Biochem. Cell Biol. 2009, 41, 990–993. [Google Scholar] [CrossRef] [PubMed]
- Kenzelmann, D.; Chiquet-Ehrismann, R.; Tucker, R.P. Teneurins, a transmembrane protein family involved in cell communication during neuronal development. Cell Mol. Life Sci. 2007, 64, 1452–1456. [Google Scholar] [CrossRef]
- Chand, D.; Colacci, M.; Dixon, K.; Kollara, A.; Brown, T.J.; Lovejoy, D.A. C-terminal region of teneurin-1 co-localizes with the dystroglycan complex in adult mouse testes and regulates testicular size and testosterone production. Histochem. Cell Biol. 2014, 141, 191–211. [Google Scholar] [CrossRef]
- Izu, H.; Inouye, S.; Fujimoto, M.; Shiraishi, K.; Naito, K.; Nakai, A. Heat shock transcription factor 1 is involved in quality-control mechanisms in male germ cells. Biol. Reprod. 2004, 70, 18–24. [Google Scholar] [CrossRef]
- Widłak, W.; Benedyk, K.; Vydra, N.; Głowala, M.; Scieglińska, D.; Małusecka, E.; Nakai, A.; Krawczyk, Z. Expression of a constitutively active mutant of heat shock factor 1 under the control of testis-specific hst70 gene promoter in transgenic mice induces degeneration of seminiferous epithelium. Acta Biochim. Pol. 2003, 50, 535–541. [Google Scholar]
- Yang, L.; Wang, Y.; Zhang, Q.; Lai, Y.; Li, C.; Zhang, Q.; Huang, W.; Duan, Y.; Jiang, Z.; Li, X.; et al. Identification of Hsf1 as a novel androgen receptor-regulated gene in mouse Sertoli cells. Mol. Reprod. Dev. 2014, 81, 514–523. [Google Scholar] [CrossRef]
- Lee, Y.-J.; Kim, E.-H.; Lee, J.S.; Jeoung, D.; Bae, S.; Kwon, S.H.; Lee, Y.-S. HSF1 as a mitotic regulator: Phosphorylation of HSF1 by Plk1 is essential for mitotic progression. Cancer Res. 2008, 68, 7550–7560. [Google Scholar] [CrossRef] [PubMed]
- Desai, S.; Liu, Z.; Yao, J.; Patel, N.; Chen, J.; Wu, Y.; Ahn, E.E.-Y.; Fodstad, O.; Tan, M. Heat shock factor 1 (HSF1) controls chemoresistance and autophagy through transcriptional regulation of autophagy-related protein 7 (ATG7). J. Biol. Chem. 2013, 288, 9165–9176. [Google Scholar] [CrossRef] [PubMed]
- Dokladny, K.; Zuhl, M.N.; Mandell, M.; Bhattacharya, D.; Schneider, S.; Deretic, V.; Moseley, P.L. Regulatory coordination between two major intracellular homeostatic systems: Heat shock response and autophagy. J. Biol. Chem. 2013, 288, 14959–14972. [Google Scholar] [CrossRef] [PubMed]
- Kumsta, C.; Chang, J.T.; Schmalz, J.; Hansen, M. Hormetic heat stress and HSF-1 induce autophagy to improve survival and proteostasis in C. elegans. Nat. Commun. 2017, 8, 14337. [Google Scholar] [CrossRef]
- Watanabe, Y.; Tsujimura, A.; Taguchi, K.; Tanaka, M. HSF1 stress response pathway regulates autophagy receptor SQSTM1/p62-associated proteostasis. Autophagy 2017, 13, 133–148. [Google Scholar] [CrossRef]
- Samarasinghe, B.; Wales, C.T.K.; Taylor, F.R.; Jacobs, A.T. Heat shock factor 1 confers resistance to Hsp90 inhibitors through p62/SQSTM1 expression and promotion of autophagic flux. Biochem. Pharmacol. 2014, 87, 445–455. [Google Scholar] [CrossRef]
- Tye, B.W.; Commins, N.; Springer, M.; Pincus, D.; Churchman, L.S. A risk-reward tradeoff of high ribosome production in proliferating cells. BioRxiv 2018. [Google Scholar] [CrossRef]
- Albert, B.; Kos-Braun, I.C.; Henras, A.K.; Dez, C.; Rueda, M.P.; Zhang, X.; Gadal, O.; Kos, M.; Shore, D. A ribosome assembly stress response regulates transcription to maintain proteome homeostasis. Elife 2019, 8. [Google Scholar] [CrossRef]
- Hsu, A.-L.; Murphy, C.T.; Kenyon, C. Regulation of aging and age-related disease by DAF-16 and heat-shock factor. Science 2003, 300, 1142–1145. [Google Scholar] [CrossRef]
- Morley, J.F.; Morimoto, R.I. Regulation of longevity in Caenorhabditis elegans by heat shock factor and molecular chaperones. Mol. Biol. Cell 2004, 15, 657–664. [Google Scholar] [CrossRef]
- Morrow, G.; Samson, M.; Michaud, S.; Tanguay, R.M. Overexpression of the small mitochondrial Hsp22 extends Drosophila life span and increases resistance to oxidative stress. FASEB J. 2004, 18, 598–599. [Google Scholar] [CrossRef] [PubMed]
- Min, J.-N.; Whaley, R.A.; Sharpless, N.E.; Lockyer, P.; Portbury, A.L.; Patterson, C. CHIP deficiency decreases longevity, with accelerated aging phenotypes accompanied by altered protein quality control. Mol. Cell. Biol. 2008, 28, 4018–4025. [Google Scholar] [CrossRef] [PubMed]
- Swindell, W.R.; Masternak, M.M.; Kopchick, J.J.; Conover, C.A.; Bartke, A.; Miller, R.A. Endocrine regulation of heat shock protein mRNA levels in long-lived dwarf mice. Mech. Ageing Dev. 2009, 130, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Baird, N.A.; Douglas, P.M.; Simic, M.S.; Grant, A.R.; Moresco, J.J.; Wolff, S.C.; Yates, J.R.; Manning, G.; Dillin, A. HSF-1-mediated cytoskeletal integrity determines thermotolerance and life span. Science 2014, 346, 360–363. [Google Scholar] [CrossRef]
- McLaughlin, P.J.; Bakall, B.; Choi, J.; Liu, Z.; Sasaki, T.; Davis, E.C.; Marmorstein, A.D.; Marmorstein, L.Y. Lack of fibulin-3 causes early ageing and herniation, but not macular degeneration in mice. Hum. Mol. Genet. 2007, 16, 3059–3070. [Google Scholar] [CrossRef]
- Stone, E.M.; Lotery, A.J.; Munier, F.L.; Héon, E.; Piguet, B.; Guymer, R.H.; Vandenburgh, K.; Cousin, P.; Nishimura, D.; Swiderski, R.E.; et al. A single EFEMP1 mutation associated with both Malattia Leventinese and Doyne honeycomb retinal dystrophy. Nat. Genet. 1999, 22, 199–202. [Google Scholar] [CrossRef]
- Ackerman, M.J.; Clapham, D.E. Ion channels--basic science and clinical disease. N. Engl. J. Med. 1997, 336, 1575–1586. [Google Scholar] [CrossRef]
- Fletcher, C.F.; Lutz, C.M.; O’Sullivan, T.N.; Shaughnessy, J.D.; Hawkes, R.; Frankel, W.N.; Copeland, N.G.; Jenkins, N.A. Absence epilepsy in tottering mutant mice is associated with calcium channel defects. Cell 1996, 87, 607–617. [Google Scholar] [CrossRef]
- Ophoff, R.A.; Terwindt, G.M.; Vergouwe, M.N.; van Eijk, R.; Oefner, P.J.; Hoffman, S.M.; Lamerdin, J.E.; Mohrenweiser, H.W.; Bulman, D.E.; Ferrari, M.; et al. Familial hemiplegic migraine and episodic ataxia type-2 are caused by mutations in the Ca2+ channel gene CACNL1A4. Cell 1996, 87, 543–552. [Google Scholar] [CrossRef]
- Wang, M.C.; Bohmann, D.; Jasper, H. JNK signaling confers tolerance to oxidative stress and extends lifespan in Drosophila. Dev. Cell 2003, 5, 811–816. [Google Scholar] [CrossRef]
- Oh, S.W.; Mukhopadhyay, A.; Svrzikapa, N.; Jiang, F.; Davis, R.J.; Tissenbaum, H.A. JNK regulates lifespan in Caenorhabditis elegans by modulating nuclear translocation of forkhead transcription factor/DAF-16. Proc. Natl. Acad. Sci. USA 2005, 102, 4494–4499. [Google Scholar] [CrossRef] [PubMed]
- Grassilli, E.; Bellesia, E.; Salomoni, P.; Croce, M.A.; Sikora, E.; Radziszewska, E.; Tesco, G.; Vergelli, M.; Latorraca, S.; Barbieri, D.; et al. c-fos/c-jun expression and AP-1 activation in skin fibroblasts from centenarians. Biochem. Biophys. Res. Commun. 1996, 226, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Hilberg, F.; Aguzzi, A.; Howells, N.; Wagner, E.F. c-jun is essential for normal mouse development and hepatogenesis. Nature 1993, 365, 179–181. [Google Scholar] [CrossRef] [PubMed]
- Mi, H.; Muruganujan, A.; Ebert, D.; Huang, X.; Thomas, P.D. PANTHER version 14: More genomes, a new PANTHER GO-slim and improvements in enrichment analysis tools. Nucleic Acids Res. 2019, 47, D419–D426. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Altenhoff, A.M.; Glover, N.M.; Train, C.-M.; Kaleb, K.; Warwick Vesztrocy, A.; Dylus, D.; de Farias, T.M.; Zile, K.; Stevenson, C.; Long, J.; et al. The OMA orthology database in 2018: Retrieving evolutionary relationships among all domains of life through richer web and programmatic interfaces. Nucleic Acids Res. 2018, 46, D477–D485. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhao, Y.; Zhang, B. Efficient Test and Visualization of Multi-Set Intersections. Sci. Rep. 2015, 5, 16923. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Activated | Inhibited | N/D | All | |
|---|---|---|---|---|
| ALL INTERACTIONS | ||||
| All records | 5258 | 1942 | 17,435 | 24,635 |
| HSF1 targets with evidence for direct interaction | 577 | 344 | 17,435 | 18,356 |
| HSF1 targets with evidence for HSF1 dependent regulation | 4681 | 1598 | 0 | 6279 |
| HSF1 targets with evidence for direct interaction and HSF1 dependent regulation | 1869 | 1023 | 0 | 2892 |
| UNIQUE INTERACTIONS | ||||
| All records | 3869 | 1272 | 10,500 | 15,641 |
| HSF1 targets with evidence for direct interaction | 404 | 207 | 10,499 | 11,110 |
| HSF1 targets with evidence for HSF1 dependent regulation | 3877 | 1325 | 0 | 5202 |
| HSF1 targets with evidence for direct interaction and HSF1 dependent regulation | 774 | 547 | 0 | 1321 |
| ALL INTERACTIONS | ||||
| Homo sapiens | 1685 | 958 | 7955 | 10,598 |
| Mus musculus | 135 | 279 | 9315 | 9729 |
| Rattus norvegicus | 465 | 166 | 0 | 631 |
| Caenorhabditis elegans | 2662 | 492 | 0 | 3154 |
| Drosophila melanogaster | 99 | 38 | 54 | 191 |
| Saccharomyces cerevisiae | 211 | 5 | 111 | 327 |
| other | 0 | 3 | 0 | 3 |
| UNIQUE INTERACTIONS | ||||
| Homo sapiens | 876 | 562 | 4182 | 5620 |
| Mus musculus | 83 | 143 | 6153 | 6379 |
| Rattus norvegicus | 454 | 146 | 0 | 600 |
| Caenorhabditis elegans | 2218 | 375 | 0 | 2593 |
| Drosophila melanogaster | 98 | 38 | 54 | 190 |
| Saccharomyces cerevisiae | 140 | 5 | 111 | 256 |
| other | 0 | 3 | 0 | 3 |
| Human ’Core’ HSF1 Target Genes | ||
|---|---|---|
| Extracellular matrix structural protein | ||
| Official Gene Symbol | Novelty (target not characterized in detail) | Regulatory information |
| creld1 | Yes | activated |
| EFEMP1 | Yes | activated |
| LTBP4 | Yes | activated |
| Signaling by NOTCH1 | ||
| Official Gene Symbol | Novelty (target not characterized in detail) | Regulatory information |
| UBB | No | activated |
| UBC | Yes | activated |
| notch1 | Yes | activated |
| TBL1X | Yes | inhibited |
| DTX2 | Yes | activated |
| Apoptotic pathway | ||
| Official Gene Symbol | Novelty (target not characterized in detail) | Regulatory information |
| HSPA1L | Yes | activated |
| HSPA1A | No | activated |
| hspa1b | No | activated |
| JUN | No | activated |
| HSPA6 | Yes | activated |
| BCL2L1 | Yes | activated |
| FOS | Yes | activated |
| ATF3 | No | activated |
| BAG3 | No | activated |
| HSPA8 | Yes | activated |
| MAP4K1 | Yes | activated |
| NFKB2 | Yes | activated |
| CFLAR | Yes | activated |
| Human ’shared’ HSF1 target genes | ||
| Inflammasomes | ||
| Official Gene Symbol | Novelty (target not characterized in detail) | Regulatory information |
| BCL2L1 | Yes | activated |
| SUGT1 | Yes | activated |
| APP | Yes | activated |
| MEFV | Yes | activated |
| HSP90AB1 | Yes | activated |
| NFKB2 | Yes | activated |
| BMAL1:CLOCK,NPAS2 activates circadian gene expression | ||
| Official Gene Symbol | Novelty (target not characterized in detail) | Regulatory information |
| RXRA | Yes | activated |
| NOCT | Yes | activated |
| NCOA2 | Yes | inhibited |
| BHLHE40 | Yes | activated |
| ARNTL | Yes | inhibited |
| SERPINE1 | No | activated |
| DBP | Yes | activated |
| TBL1X | Yes | inhibited |
| RAF-independent MAPK1/3 activation | ||
| Official Gene Symbol | Novelty (target not characterized in detail) | Regulatory information |
| DUSP2 | Yes | activated |
| DUSP10 | Yes | activated |
| DUSP5 | Yes | activated |
| MAP2K2 | Yes | activated |
| DUSP1 | Yes | activated |
| HDMs demethylate histones | ||
| Official Gene Symbol | Novelty (target not characterized in detail) | Regulatory information |
| PHF8 | Yes | activated |
| ARID5B | Yes | activated |
| KDM4B | Yes | activated |
| JMJD6 | Yes | activated |
| Murine ’Shared’ HSF1 Target Genes | |||
|---|---|---|---|
| Ribosome biogenesis | |||
| Official Gene Symbol | Novelty (target not characterized in detail) | Regulatory information | |
| RPL7L1 | Yes | inhibited | |
| XPO1 | Yes | inhibited | |
| G2/M Checkpoints | |||
| Official Gene Symbol | Novelty (target not characterized in detail) | Regulatory information | |
| TOPBP1 | Yes | inhibited | |
| RAD1 | Yes | inhibited | |
| UBC | No | activated | |
| Oocyte | |||
| Sister chromatid segregation | |||
| Official Gene Symbol | Novelty (target not characterized in detail) | Regulatory information | |
| STAG3 | Yes | inhibited | |
| STAG2 | Yes | activated | |
| Spermatocyte | |||
| Cell–cell adhesion via plasma-membrane adhesion molecules | |||
| Official Gene Symbol | Novelty (target not characterized in detail) | Regulated directly by HSF1 | Regulatory information |
| CRB1 | Yes | N/D | N/D |
| TENM4 | Yes | N/D | N/D |
| CDH19 | Yes | N/D | N/D |
| CCDC141 | Yes | N/D | N/D |
| CDH12 | Yes | N/D | N/D |
| CDH18 | Yes | N/D | N/D |
| SDK1 | Yes | N/D | N/D |
| CNTN5 | Yes | N/D | N/D |
| CDH6 | Yes | N/D | N/D |
| CDH26 | Yes | N/D | N/D |
| NECTIN1 | Yes | N/D | N/D |
| CDH15 | Yes | N/D | N/D |
| CDH13 | Yes | N/D | N/D |
| CNTN6 | Yes | N/D | N/D |
| CDH2 | Yes | N/D | N/D |
| NECTIN3 | Yes | N/D | N/D |
| CDH10 | Yes | N/D | N/D |
| MYPN | Yes | N/D | N/D |
| CDH4 | Yes | N/D | N/D |
| HSF1 Target Genes Conserved between Vertebrate and Non-Vertebrate Species | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Human | Mouse | Rat | Worm | Fly | Yeast | ||||||
| autophagy | |||||||||||
| Off. gene symb. | Reg. info. | Off. gene symb. | Reg. info. | Off. gene symb. | Reg. info. | Off. gene symb. | Reg. info. | Off. gene symb. | Reg. info. | Off. gene symb. | Reg. info. |
| FIS1 | N/D | N/D | N/D | N/D | N/D | fis-1 | N/D | N/D | N/D | N/D | N/D |
| atg9a | N/D | atg9a | N/D | atg9a | N/D | atg-9 | N/D | N/D | N/D | N/D | N/D |
| ATG2A | N/D | ATG2A | N/D | N/D | N/D | atg-2 | N/D | N/D | N/D | N/D | N/D |
| HSPA8 | act. | HSPA8 | act. | N/D | N/D | HSP-1 | act. | N/D | N/D | N/D | N/D |
| atg2b | N/D | atg2b | N/D | N/D | N/D | atg-2 | N/D | N/D | N/D | N/D | N/D |
| RB1CC1 | N/D | N/D | N/D | N/D | N/D | epg-7 | N/D | N/D | N/D | N/D | N/D |
| ribosome biogenesis | |||||||||||
| Off. gene symb. | Reg. info. | Off. gene symb. | Reg. info. | Off. gene symb. | Reg. info. | Off. gene symb. | Reg. info. | Off. gene symb. | Reg. info. | Off. gene symb. | Reg. info. |
| RPL35A | N/D | RPL35A | N/D | N/D | N/D | rpl-33 | N/D | N/D | N/D | N/D | N/D |
| RPS27 | N/D | N/D | N/D | N/D | N/D | rps-27 | N/D | N/D | N/D | N/D | N/D |
| SART1 | N/D | SART1 | N/D | N/D | N/D | F19F10.9 | N/D | N/D | N/D | N/D | N/D |
| RPS7 | N/D | N/D | N/D | N/D | N/D | rps-7 | N/D | N/D | N/D | N/D | N/D |
| RPS21 | N/D | RPS21 | N/D | N/D | N/D | rps-21 | N/D | N/D | N/D | N/D | N/D |
| BTRC | N/D | N/D | N/D | N/D | N/D | lin-23 | N/D | N/D | N/D | N/D | N/D |
| RPS14 | act. | N/D | N/D | N/D | N/D | rps-14 | N/D | N/D | N/D | N/D | N/D |
| RPL26 | N/D | RPL26 | N/D | N/D | N/D | rpl-26 | N/D | N/D | N/D | N/D | N/D |
| HEATR3 | N/D | HEATR3 | N/D | N/D | N/D | N/D | N/D | CG10286 | act. | N/D | N/D |
| MRTO4 | N/D | N/D | N/D | MRTO4 | N/D | N/D | N/D | N/D | N/D | MRT4 | act. |
| RPL26L1 | N/D | RPL26 | N/D | N/D | N/D | rpl-26 | N/D | N/D | N/D | N/D | N/D |
| Glycolysis | |||||||||||
| Off. gene symb. | Reg. info. | Off. gene symb. | Reg. info. | Off. gene symb. | Reg. info. | Off. gene symb. | Reg. info. | Off. gene symb. | Reg. info. | Off. gene symb. | Reg. info. |
| PGAM1 | N/D | PGAM1 | N/D | N/D | N/D | N/D | N/D | N/D | N/D | GPM1 | act. |
| GPI | N/D | Gpi1 | N/D | N/D | N/D | N/D | N/D | Pgi | act. | N/D | N/D |
| PGK1 | N/D | PGK1 | N/D | N/D | N/D | N/D | N/D | N/D | N/D | PGK1 | act. |
| Wnt signalling pathway | |||||||||||
| Off. gene symb. | Reg. info. | Off. gene symb. | Reg. info. | Off. gene symb. | Reg. info. | Off. gene symb. | Reg. info. | Off. gene symb. | Reg. info. | Off. gene symb. | Reg. info. |
| SMAD1 | N/D | SMAD1 | inh. | N/D | N/D | sma-2 | N/D | N/D | N/D | N/D | N/D |
| PPP3CB | N/D | PPP3CB | inh. | N/D | N/D | tax-6 | N/D | N/D | N/D | N/D | N/D |
| FBXW11 | inh. | FBXW11 | N/D | N/D | N/D | lin-23 | N/D | N/D | N/D | N/D | N/D |
| Establishment of Sister Chromatid Cohesion | |||||||||||
| Off. gene symb. | Reg. info. | Off. gene symb. | Reg. info. | Off. gene symb. | Reg. info. | Off. gene symb. | Reg. info. | Off. gene symb. | Reg. info. | Off. gene symb. | Reg. info. |
| SMC3 | N/D | SMC3 | N/D | N/D | N/D | N/D | N/D | N/D | N/D | SMC3 | act. |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kovács, D.; Sigmond, T.; Hotzi, B.; Bohár, B.; Fazekas, D.; Deák, V.; Vellai, T.; Barna, J. HSF1Base: A Comprehensive Database of HSF1 (Heat Shock Factor 1) Target Genes. Int. J. Mol. Sci. 2019, 20, 5815. https://doi.org/10.3390/ijms20225815
Kovács D, Sigmond T, Hotzi B, Bohár B, Fazekas D, Deák V, Vellai T, Barna J. HSF1Base: A Comprehensive Database of HSF1 (Heat Shock Factor 1) Target Genes. International Journal of Molecular Sciences. 2019; 20(22):5815. https://doi.org/10.3390/ijms20225815
Chicago/Turabian StyleKovács, Dániel, Tímea Sigmond, Bernadette Hotzi, Balázs Bohár, Dávid Fazekas, Veronika Deák, Tibor Vellai, and János Barna. 2019. "HSF1Base: A Comprehensive Database of HSF1 (Heat Shock Factor 1) Target Genes" International Journal of Molecular Sciences 20, no. 22: 5815. https://doi.org/10.3390/ijms20225815
APA StyleKovács, D., Sigmond, T., Hotzi, B., Bohár, B., Fazekas, D., Deák, V., Vellai, T., & Barna, J. (2019). HSF1Base: A Comprehensive Database of HSF1 (Heat Shock Factor 1) Target Genes. International Journal of Molecular Sciences, 20(22), 5815. https://doi.org/10.3390/ijms20225815