A Single Synonymous Variant (c.354G>A [p.P118P]) in ADAMTS13 Confers Enhanced Specific Activity

 , , , and
, , , and
Abstract
1. Introduction
2. Results
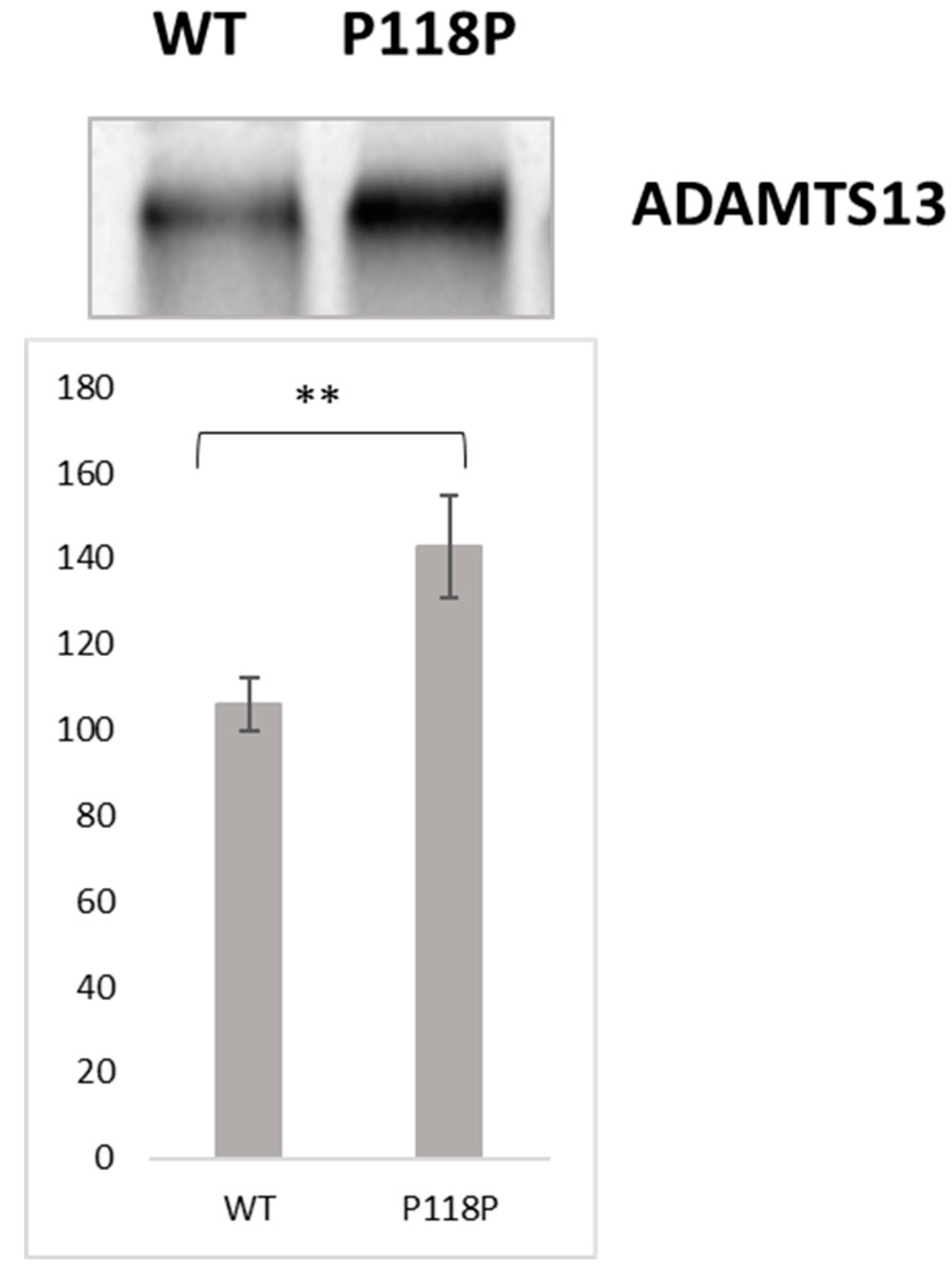
2.1. In Vitro Translation Assay Revealed Higher Protein Yield From c.354G>A [p.P118P] Variant
2.2. In Silico Analysis of c.354G>A Variant Revealed That This Mutation Leads to the Introduction of a More Frequently Used Codon/Codon Pair(s)
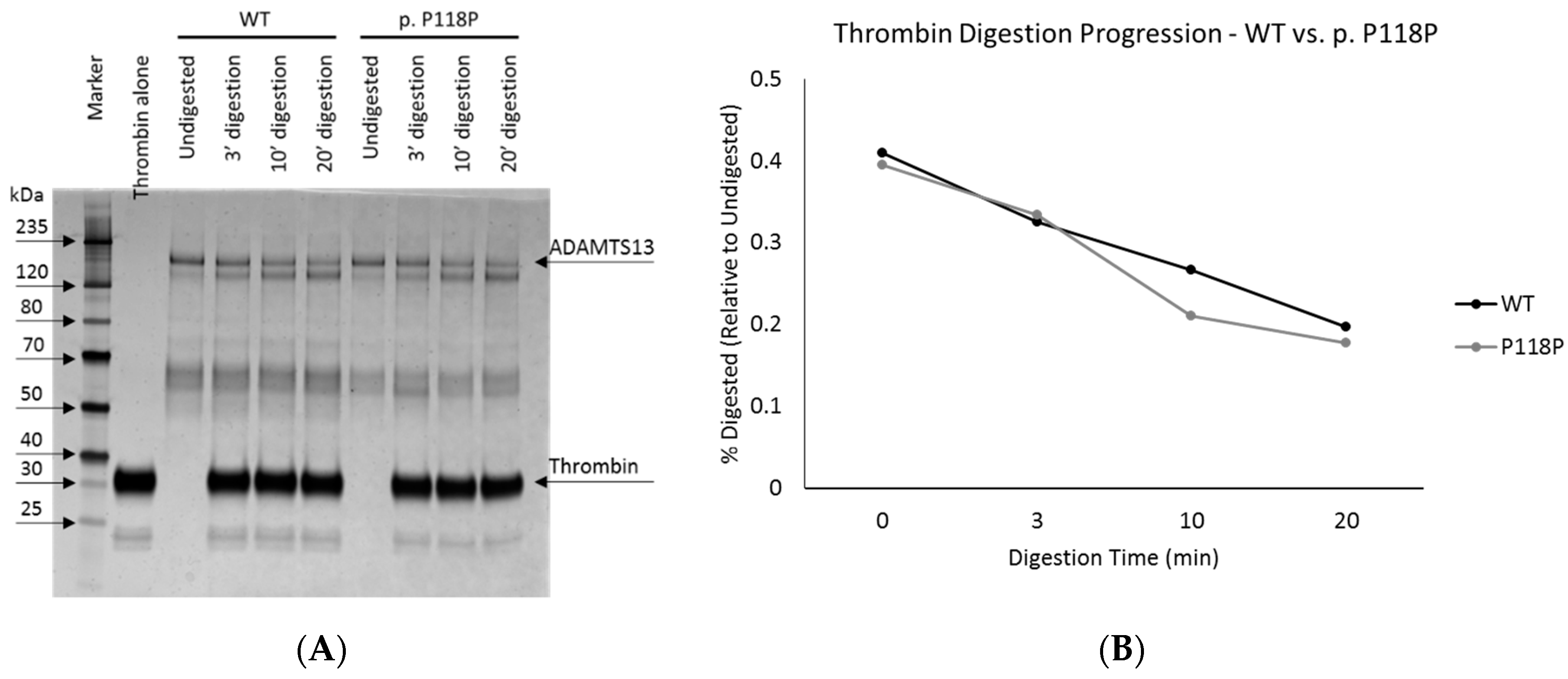
2.3. Limited Proteolysis and Circular Dichroism Analysis of the Wild-Type and p.P118P Variants Did Not Identify Conformational Differences
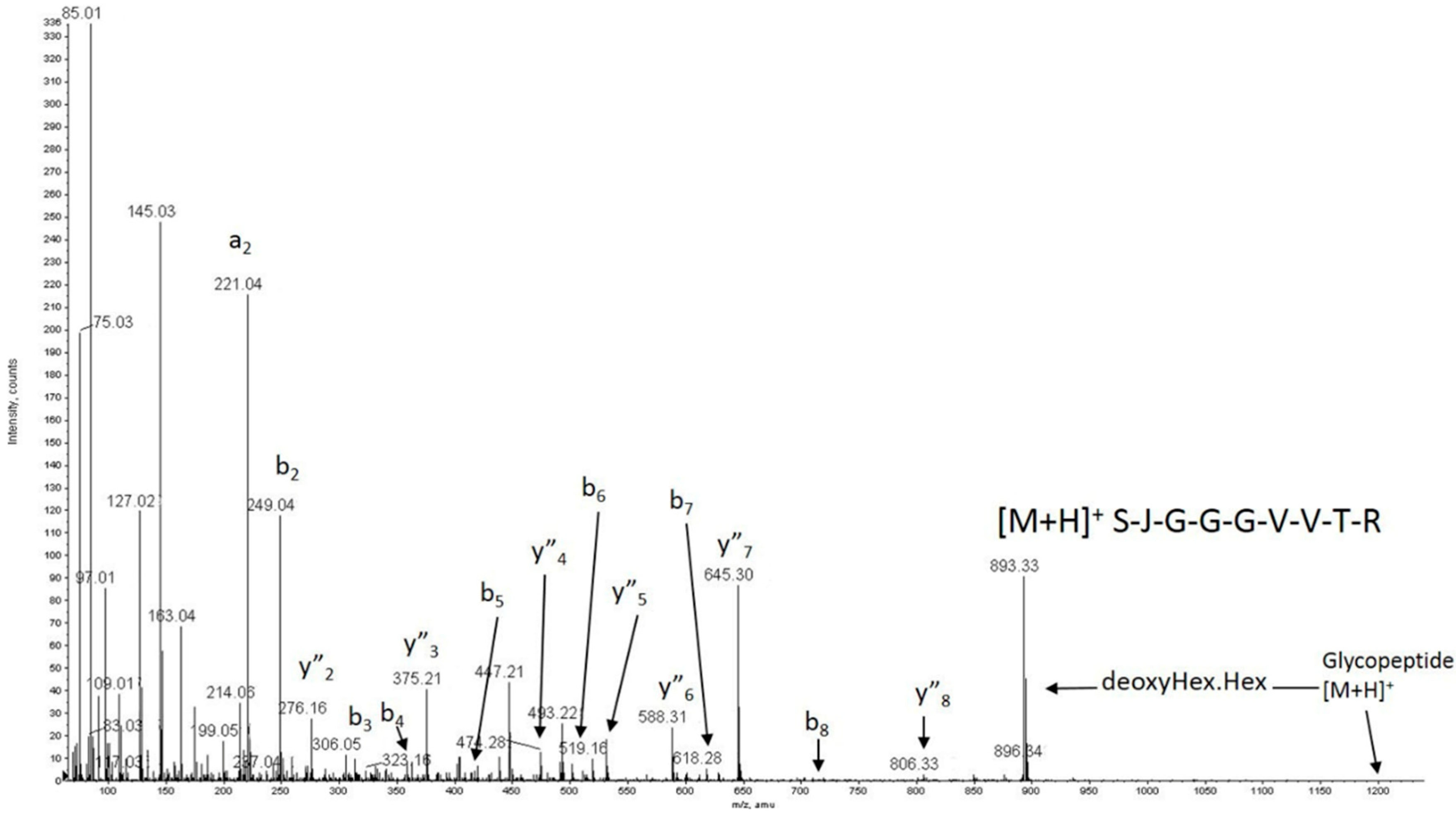
2.4. Post-Translational Modifications Analysis of Wild-Type and p.P118P Variants Showed No Significant Differences, but Revealed Previously Unreported Modifications
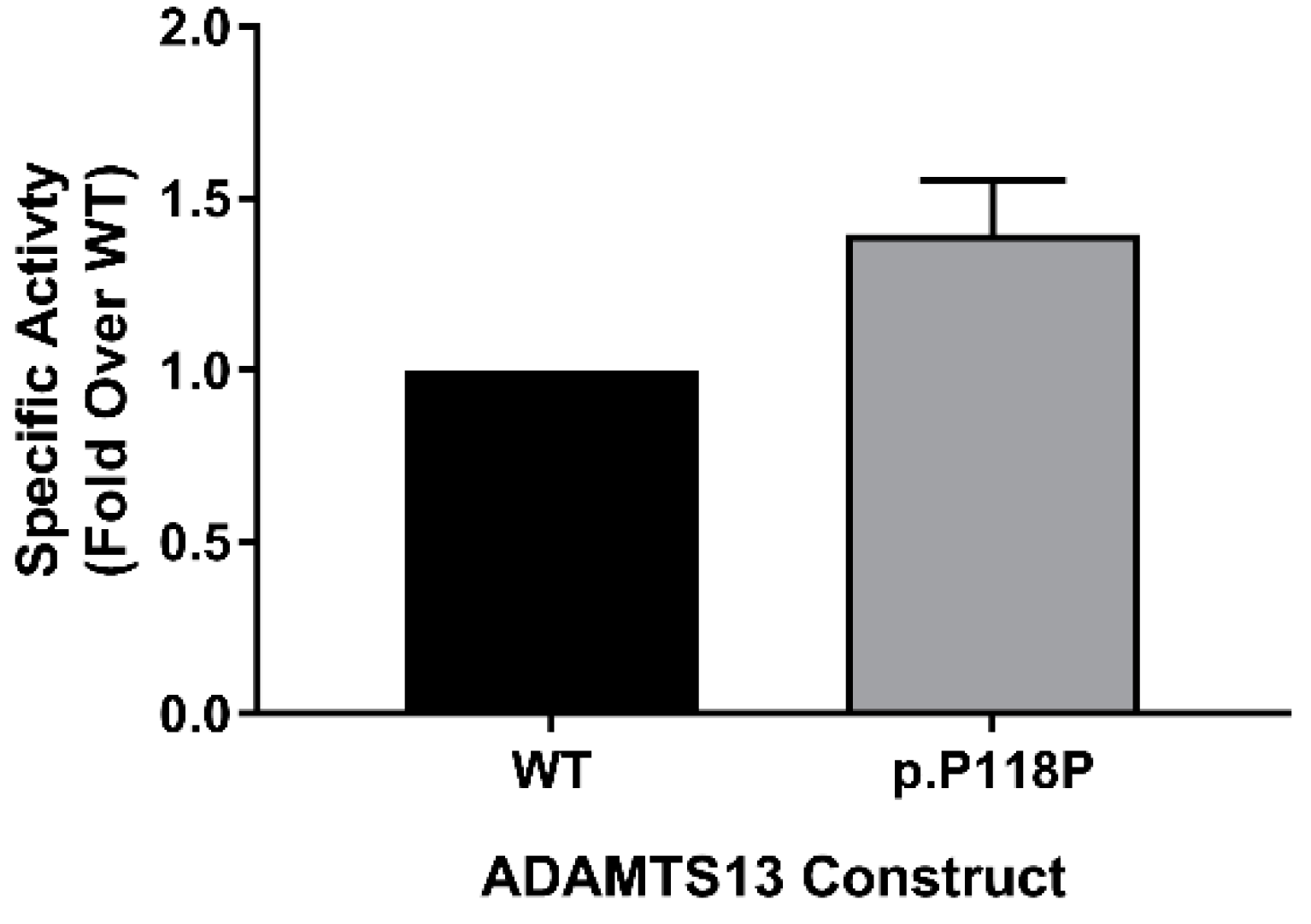
2.5. FRETS-VWF73 Assay Revealed Increased Specific Activity of p.P118P Variant
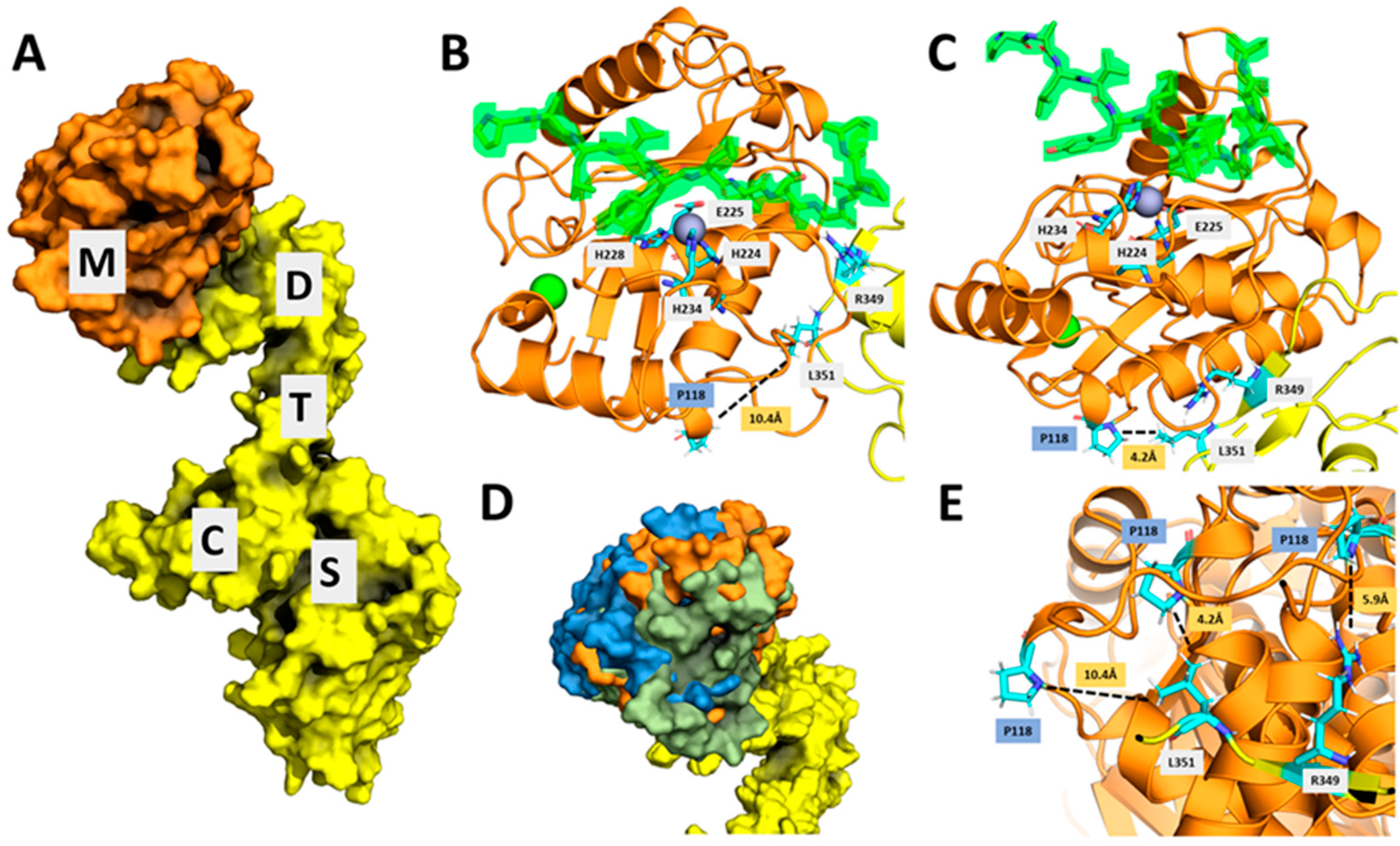
2.6. A Combination of Subtle Conformational Changes Arising from c.354G>A Variant and the Proximity of Proline 118 to an Exosite in Disintegrin Domain May Explain Increased Specific Activity
3. Discussion
4. Materials and Method
4.1. Cell Line Establishment and Maintenance
4.2. ADAMTS13 Purification and Quality Assessment
4.3. In Vitro Translation Assay
4.4. Ribosome Profiling—Library Preparation and Data Processing
4.5. Calculation of Codon and Codon Pair Usage Characteristics
4.6. Limited Proteolysis—Thrombin Digestion
4.7. Circular Dichroism (CD) Analysis
4.8. Post-Translational Modification Analysis by Mass Spectrometry
4.9. Measuring Specific Activity—FRETS-VWF73 Assay
4.10. Modeling the Metalloprotease Domain and Its Docking with the DTCS Domains
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Disclaimer
Abbreviations
| ADAMTS13 | A disintegrin and metalloproteinase with a thrombospondin type 1 motif, member 13 |
| DTCS | Disintegrin (D), thrombospondin type-1 repeat-1 (T), cysteine-rich (C), and spacer (S) domains |
| VWF | von Willebrand factor |
| RSCU | Relative Synonymous Codon Usage |
| PDB | Protein Data Bank |
| RIBO-SEQ | Ribosome Profiling |
| RNA-SEQ | RNA-Sequencing |
| HIVE-CUTS | High-performance Integrated Virtual Environment-Codon Usage Tables |
| CoCoPUTs | Codon and Codon-Pair Usage Tables |
References
- Hunt, R.C.; Simhadri, V.L.; Iandoli, M.; Sauna, Z.E.; Kimchi-Sarfaty, C. Exposing synonymous mutations. Trends Genet. 2014, 30, 308–321. [Google Scholar] [CrossRef] [PubMed]
- Kimchi-Sarfaty, C.; Oh, J.M.; Kim, I.W.; Sauna, Z.E.; Calcagno, A.M.; Ambudkar, S.V.; Gottesman, M.M. A “silent” polymorphism in the mdr1 gene changes substrate specificity. Science 2007, 315, 525–528. [Google Scholar] [CrossRef] [PubMed]
- Komar, A.A. Genetics. Snps, silent but not invisible. Science 2007, 315, 466–467. [Google Scholar] [CrossRef] [PubMed]
- Sauna, Z.E.; Kimchi-Sarfaty, C. Understanding the contribution of synonymous mutations to human disease. Nat. Rev. Genet. 2011, 12, 683–691. [Google Scholar] [CrossRef]
- Supek, F.; Minana, B.; Valcarcel, J.; Gabaldon, T.; Lehner, B. Synonymous mutations frequently act as driver mutations in human cancers. Cell 2014, 156, 1324–1335. [Google Scholar] [CrossRef]
- Plotkin, J.B.; Kudla, G. Synonymous but not the same: The causes and consequences of codon bias. Nat. Rev. Genet. 2011, 12, 32–42. [Google Scholar] [CrossRef]
- Katneni, U.K.; Liss, A.; Holcomb, D.; Katagiri, N.H.; Hunt, R.; Bar, H.; Ismail, A.; Komar, A.A.; Kimchi-Sarfaty, C. Splicing dysregulation contributes to the pathogenicity of several f9 exonic point variants. Mol. Genet. Genom. Med. 2019, 7, e840. [Google Scholar] [CrossRef]
- Kramer, E.B.; Vallabhaneni, H.; Mayer, L.M.; Farabaugh, P.J. A comprehensive analysis of translational missense errors in the yeast saccharomyces cerevisiae. RNA 2010, 16, 1797–1808. [Google Scholar] [CrossRef]
- Kramer, E.B.; Farabaugh, P.J. The frequency of translational misreading errors in e. Coli is largely determined by trna competition. RNA 2007, 13, 87–96. [Google Scholar] [CrossRef]
- Shpaer, E.G. Constraints on codon context in escherichia coli genes. Their possible role in modulating the efficiency of translation. J. Mol. Biol. 1986, 188, 555–564. [Google Scholar] [CrossRef]
- Buhr, F.; Jha, S.; Thommen, M.; Mittelstaet, J.; Kutz, F.; Schwalbe, H.; Rodnina, M.V.; Komar, A.A. Synonymous codons direct cotranslational folding toward different protein conformations. Mol. Cell 2016, 61, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.L. Structure-function and regulation of adamts-13 protease. J. Thromb. Haemost. 2013, 11, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Katneni, U.K.; Ibla, J.C.; Hunt, R.; Schiller, T.; Kimchi-Sarfaty, C. Von willebrand factor/adamts-13 interactions at birth: Implications for thrombosis in the neonatal period. J. Thromb. Haemost. 2019, 17, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Lambers, M.; Goldenberg, N.A.; Kenet, G.; Kirkham, F.J.; Manner, D.; Bernard, T.; Mesters, R.M.; Junker, R.; Stoll, M.; Nowak-Gottl, U. Role of reduced adamts13 in arterial ischemic stroke: A pediatric cohort study. Ann. Neurol 2013, 73, 58–64. [Google Scholar] [CrossRef]
- Miyata, T.; Kokame, K.; Matsumoto, M.; Fujimura, Y. Adamts13 activity and genetic mutations in japan. Hamostaseologie 2013, 33, 131–137. [Google Scholar]
- Mazetto, B.M.; Orsi, F.L.; Barnabe, A.; de Paula, E.V.; Flores-Nascimento, M.C.; Annichino-Bizzacchi, J.M. Increased adamts13 activity in patients with venous thromboembolism. Thromb. Res. 2012, 130, 889–893. [Google Scholar] [CrossRef]
- Katneni, U.K.; Hunt, R.; Hettiarachchi, G.K.; Hamasaki-Katagiri, N.; Kimchi-Sarfaty, C.; Ibla, J.C. Compounding variants rescue the effect of a deleterious adamts13 mutation in a child with severe congenital heart disease. Thromb. Res. 2017, 158, 98–101. [Google Scholar] [CrossRef]
- Edwards, N.C.; Hing, Z.A.; Perry, A.; Blaisdell, A.; Kopelman, D.B.; Fathke, R.; Plum, W.; Newell, J.; Allen, C.E.; Geetha, S.; et al. Characterization of coding synonymous and non-synonymous variants in adamts13 using ex vivo and in silico approaches. PLoS ONE 2012, 7, e38864. [Google Scholar]
- Auton, A.; Abecasis, G.R.; Altshuler, D.M.; Durbin, R.M.; Abecasis, G.R.; Bentley, D.R.; Chakravarti, A.; Clark, A.G.; Donnelly, P.; Eichler, E.E.; et al. A global reference for human genetic variation. Nature 2015, 526, 68–74. [Google Scholar]
- Lek, M.; Karczewski, K.J.; Minikel, E.V.; Samocha, K.E.; Banks, E.; Fennell, T.; O’Donnell-Luria, A.H.; Ware, J.S.; Hill, A.J.; Cummings, B.B.; et al. Analysis of protein-coding genetic variation in 60,706 humans. Nature 2016, 536, 285–291. [Google Scholar] [CrossRef]
- Ingolia, N.T.; Ghaemmaghami, S.; Newman, J.R.S.; Weissman, J.S. Genome-wide analysis in vivo of translation with nucleotide resolution using ribosome profiling. Science (N. Y.) 2009, 324, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Urrutia, A.O.; Hurst, L.D. The signature of selection mediated by expression on human genes. Genome Res. 2003, 13, 2260–2264. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, M.A.; Kurland, C.G.; Pedersen, S. Codon usage determines translation rate in escherichia coli. J. Mol. Biol. 1989, 207, 365–377. [Google Scholar] [CrossRef]
- Tats, A.; Tenson, T.; Remm, M. Preferred and avoided codon pairs in three domains of life. BMC Genom. 2008, 9, 463. [Google Scholar] [CrossRef]
- Sharp, P.M.; Tuohy, T.M.; Mosurski, K.R. Codon usage in yeast: Cluster analysis clearly differentiates highly and lowly expressed genes. Nucleic Acids Res. 1986, 14, 5125–5143. [Google Scholar] [CrossRef]
- Uemura, M.; Tatsumi, K.; Matsumoto, M.; Fujimoto, M.; Matsuyama, T.; Ishikawa, M.; Iwamoto, T.-a.; Mori, T.; Wanaka, A.; Fukui, H.; et al. Localization of adamts13 to the stellate cells of human liver. Blood 2005, 106, 922–924. [Google Scholar]
- Kim, S.J.; Yoon, J.S.; Shishido, H.; Yang, Z.; Rooney, L.A.; Barral, J.M.; Skach, W.R. Translational tuning optimizes nascent protein folding in cells. Science 2015, 348, 444–448. [Google Scholar] [CrossRef]
- Komar, A.A.; Lesnik, T.; Reiss, C. Synonymous codon substitutions affect ribosome traffic and protein folding during in vitro translation. FEBS Lett. 1999, 462, 387–391. [Google Scholar] [CrossRef]
- Crawley, J.T.B.; Lam, J.K.; Rance, J.B.; Mollica, L.R.; O’Donnell, J.S.; Lane, D.A. Proteolytic inactivation of adamts13 by thrombin and plasmin. Blood 2005, 105, 1085–1093. [Google Scholar] [CrossRef]
- Kaufman, R.J. Post-translational modifications required for coagulation factor secretion and function. Thromb. Haemost. 1998, 79, 1068–1079. [Google Scholar] [CrossRef]
- Van Der Wel, H.; Morris, H.R.; Panico, M.; Paxton, T.; North, S.J.; Dell, A.; Thomson, J.M.; West, C.M. A non-golgi alpha 1,2-fucosyltransferase that modifies skp1 in the cytoplasm of dictyostelium. J. Biol. Chem. 2001, 276, 33952–33963. [Google Scholar] [CrossRef] [PubMed]
- Morris, H.R.; Paxton, T.; Dell, A.; Langhorne, J.; Berg, M.; Bordoli, R.S.; Hoyes, J.; Bateman, R.H. High sensitivity collisionally-activated decomposition tandem mass spectrometry on a novel quadrupole/orthogonal-acceleration time-of-flight mass spectrometer. Rapid Commun. Mass Spectrom. 1996, 10, 889–896. [Google Scholar] [CrossRef]
- Teng-umnuay, P.; Morris, H.R.; Dell, A.; Panico, M.; Paxton, T.; West, C.M. The cytoplasmic f-box binding protein skp1 contains a novel pentasaccharide linked to hydroxyproline in dictyostelium. J. Biol. Chem. 1998, 273, 18242–18249. [Google Scholar] [CrossRef] [PubMed]
- Hofsteenge, J.; Huwiler, K.G.; Macek, B.; Hess, D.; Lawler, J.; Mosher, D.F.; Peter-Katalinic, J. C-mannosylation and o-fucosylation of the thrombospondin type 1 module. J. Biol. Chem. 2001, 276, 6485–6498. [Google Scholar] [CrossRef]
- Wang, L.W.; Leonhard-Melief, C.; Haltiwanger, R.S.; Apte, S.S. Post-translational modification of thrombospondin type-1 repeats in adamts-like 1/punctin-1 by c-mannosylation of tryptophan. J. Biol. Chem. 2009, 284, 30004–30015. [Google Scholar] [CrossRef]
- Sorvillo, N.; Kaijen, P.H.; Matsumoto, M.; Fujimura, Y.; van der Zwaan, C.; Verbij, F.C.; Pos, W.; Fijnheer, R.; Voorberg, J.; Meijer, A.B. Identification of n-linked glycosylation and putative o-fucosylation, c-mannosylation sites in plasma derived adamts13. J. Thromb. Haemost. 2014, 12, 670–679. [Google Scholar] [CrossRef]
- Feys, H.B.; Pareyn, I.; Vancraenenbroeck, R.; de Maeyer, M.; Deckmyn, H.; van Geet, C.; Vanhoorelbeke, K. Mutation of the h-bond acceptor s119 in the adamts13 metalloprotease domain reduces secretion and substrate turnover in a patient with congenital thrombotic thrombocytopenic purpura. Blood 2009, 114, 4749–4752. [Google Scholar] [CrossRef]
- Rurali, E.; Noris, M.; Chianca, A.; Donadelli, R.; Banterla, F.; Galbusera, M.; Gherardi, G.; Gastoldi, S.; Parvanova, A.; Iliev, I.; et al. Adamts13 predicts renal and cardiovascular events in type 2 diabetic patients and response to therapy. Diabetes 2013, 62, 3599–3609. [Google Scholar] [CrossRef]
- Akiyama, M.; Nakayama, D.; Takeda, S.; Kokame, K.; Takagi, J.; Miyata, T. Crystal structure and enzymatic activity of an adamts-13 mutant with the east asian-specific p475s polymorphism. J. Thromb. Haemost. 2013, 11, 1399–1406. [Google Scholar] [CrossRef]
- Verbij, F.C.; Stokhuijzen, E.; Kaijen, P.H.P.; van Alphen, F.; Meijer, A.B.; Voorberg, J. Identification of glycans on plasma-derived adamts13. Blood 2016, 128, 51–58. [Google Scholar] [CrossRef]
- Ricketts, L.M.; Dlugosz, M.; Luther, K.B.; Haltiwanger, R.S.; Majerus, E.M. O-fucosylation is required for adamts13 secretion. J. Biol. Chem. 2007, 282, 17014–17023. [Google Scholar] [CrossRef] [PubMed]
- Kokame, K.; Nobe, Y.; Kokubo, Y.; Okayama, A.; Miyata, T. Frets-vwf73, a first fluorogenic substrate for adamts13 assay. Br. J. Haematol. 2005, 129, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Ercig, B.; Wichapong, K.; Reutelingsperger, C.P.M.; Vanhoorelbeke, K.; Voorberg, J.; Nicolaes, G.A.F. Insights into 3d structure of adamts13: A stepping stone towards novel therapeutic treatment of thrombotic thrombocytopenic purpura. Thromb. Haemost. 2018, 118, 28–41. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Liu, W.; Fang, J.; Shi, X.; Wu, J.; Fang, Y.; Lin, J. Afm imaging reveals multiple conformational states of adamts13. J. Biol. Eng. 2019, 13, 9. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Muia, J.; Gupta, G.; Westfield, L.A.; Vanhoorelbeke, K.; Tolia, N.H.; Sadler, J.E. Exploring the “minimal” structure of a functional adamts13 by mutagenesis and small-angle X-ray scattering. Blood 2019, 133, 1909–1918. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, M.; Takeda, S.; Kokame, K.; Takagi, J.; Miyata, T. Crystal structures of the noncatalytic domains of adamts13 reveal multiple discontinuous exosites for von willebrand factor. Proc. Natl. Acad. Sci. USA 2009, 106, 19274–19279. [Google Scholar] [CrossRef]
- Crawley, J.T.; de Groot, R.; Xiang, Y.; Luken, B.M.; Lane, D.A. Unraveling the scissile bond: How adamts13 recognizes and cleaves von willebrand factor. Blood 2011, 118, 3212–3221. [Google Scholar] [CrossRef]
- De Groot, R.; Bardhan, A.; Ramroop, N.; Lane, D.A.; Crawley, J.T.B. Essential role of the disintegrin-like domain in adamts13 function. Blood 2009, 113, 5609–5616. [Google Scholar] [CrossRef]
- Simhadri, V.L.; Hamasaki-Katagiri, N.; Lin, B.C.; Hunt, R.; Jha, S.; Tseng, S.C.; Wu, A.; Bentley, A.A.; Zichel, R.; Lu, Q.; et al. Single synonymous mutation in factor ix alters protein properties and underlies haemophilia b. J. Med. Genet. 2017, 54, 338–345. [Google Scholar] [CrossRef]
- Bartoszewski, R.A.; Jablonsky, M.; Bartoszewska, S.; Stevenson, L.; Dai, Q.; Kappes, J.; Collawn, J.F.; Bebok, Z. A synonymous single nucleotide polymorphism in δf508 cftr alters the secondary structure of the mrna and the expression of the mutant protein. J. Biol. Chem. 2010, 285, 28741–28748. [Google Scholar] [CrossRef]
- Alexaki, A.; Hettiarachchi, G.K.; Athey, J.C.; Katneni, U.K.; Simhadri, V.; Hamasaki-Katagiri, N.; Nanavaty, P.; Lin, B.; Takeda, K.; Freedberg, D.; et al. Effects of codon optimization on coagulation factor ix translation and structure: Implications for protein and gene therapies. Sci. Rep. 2019, 9, 15449. [Google Scholar] [CrossRef] [PubMed]
- Komar, A.A. A pause for thought along the co-translational folding pathway. Trends Biochem. Sci. 2009, 34, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.-H.; Dang, Y.; Zhou, Z.; Wu, C.; Zhao, F.; Sachs, M.S.; Liu, Y. Codon usage influences the local rate of translation elongation to regulate co-translational protein folding. Mol. Cell 2015, 59, 744–754. [Google Scholar] [CrossRef]
- Komar, A.A. Unraveling co-translational protein folding: Concepts and methods. Methods 2018, 137, 71–81. [Google Scholar] [CrossRef]
- Suwanmanee, T.; Hu, G.; Gui, T.; Bartholomae, C.C.; Kutschera, I.; von Kalle, C.; Schmidt, M.; Monahan, P.E.; Kafri, T. Integration-deficient lentiviral vectors expressing codon-optimized r338l human fix restore normal hemostasis in hemophilia b mice. Mol. Ther. 2014, 22, 567–574. [Google Scholar] [CrossRef]
- Dittmar, K.A.; Goodenbour, J.M.; Pan, T. Tissue-specific differences in human transfer rna expression. PLoS Genet. 2006, 2, e221. [Google Scholar] [CrossRef]
- Alexaki, A.; Kames, J.; Holcomb, D.D.; Athey, J.; Santana-Quintero, L.V.; Lam, P.V.N.; Hamasaki-Katagiri, N.; Osipova, E.; Simonyan, V.; Bar, H.; et al. Codon and codon-pair usage tables (cocoputs): Facilitating genetic variation analyses and recombinant gene design. J. Mol. Biol. 2019, 431, 2434–2441. [Google Scholar] [CrossRef]
- Merkin, J.; Russell, C.; Chen, P.; Burge, C.B. Evolutionary dynamics of gene and isoform regulation in mammalian tissues. Science 2012, 338, 1593–1599. [Google Scholar] [CrossRef]
- Hettiarachchi, G.K.; Katneni, U.K.; Hunt, R.C.; Kames, J.M.; Athey, J.C.; Bar, H.; Sauna, Z.E.; McGill, J.R.; Ibla, J.C.; Kimchi-Sarfaty, C. Translational and transcriptional responses in human primary hepatocytes under hypoxia. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 316, G720–G734. [Google Scholar] [CrossRef]
- Sharp, P.M.; Li, W.H. The codon adaptation index-a measure of directional synonymous codon usage bias, and its potential applications. Nucleic Acids Res. 1987, 15, 1281–1295. [Google Scholar] [CrossRef] [PubMed]
- Athey, J.; Alexaki, A.; Osipova, E.; Rostovtsev, A.; Santana-Quintero, L.V.; Katneni, U.; Simonyan, V.; Kimchi-Sarfaty, C. A new and updated resource for codon usage tables. BMC Bioinform. 2017, 18, 391. [Google Scholar] [CrossRef] [PubMed]
- Holcomb, D.D.; Alexaki, A.; Katneni, U.; Kimchi-Sarfaty, C. The kazusa codon usage database, cocoputs, and the value of up-to-date codon usage statistics. Infect. Genet. Evol. 2019, 73, 266–268. [Google Scholar] [CrossRef] [PubMed]
- Consortium, G.T. The genotype-tissue expression (gtex) project. Nat. Genet. 2013, 45, 580–585. [Google Scholar]
- Sauna, Z.E.; Okunji, C.; Hunt, R.C.; Gupta, T.; Allen, C.; Plum, E.; Blaisdell, A.; Grigoryan, V.; Geetha, S.; Fathke, R.; et al. Characterization of conformation-sensitive antibodies to adamts13, the von willebrand cleavage protease. PLoS ONE 2009, 4, e6506. [Google Scholar] [CrossRef] [PubMed]
- Lyskov, S.; Chou, F.-C.; Conchúir, S.Ó.; Der, B.S.; Drew, K.; Kuroda, D.; Xu, J.; Weitzner, B.D.; Renfrew, P.D.; Sripakdeevong, P.; et al. Serverification of molecular modeling applications: The rosetta online server that includes everyone (rosie). PLoS ONE 2013, 8, e63906. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variant | Codon | Relative Synonymous Codon Usage (RSCU) a | ||
|---|---|---|---|---|
| Human Genome | ADAMTS13 cDNA | Human Liver | ||
| WT | CCG | 0.39 | 0.44 | 0.43 |
| c.354G>A | CCA | 1.19 | 0.92 | 1.03 |
| c.354G>C | CCT | 1.21 | 1.19 | 1.15 |
| c.354G>T | CCC | 1.21 | 1.46 | 1.39 |
| Variant | Codon Pairs | Human Genome | Human Liver | ||
|---|---|---|---|---|---|
| Codon Pair Frequency a | Codon Pair Percentile | Codon Pair Frequency | Codon Pair Percentile | ||
| WT | GAC CCG CCG TCC | 179.158 123.832 | 48.315 35.181 | 81.21 53.73 | 28.64 20.73 |
| c.354G>A | GAC CCA CCA TCC | 423.023 311.350 | 84.082 72.192 | 367.07 329.59 | 77.83 74.29 |
| c.354G>C | GAC CCC CCC TCC | 490.152 394.130 | 88.330 81.665 | 664.01 315.10 | 93.12 72.73 |
| c.354G>T | GAC CCT CCT TCC | 635.956 453.802 | 94.189 86.230 | 318.38 217.91 | 73.14 59.16 |
| Un-Mapped Signals Observed in the MS and MS/MS WT Data a | Glycopeptide Assignments from the MS and MS/MS Spectra b | Corresponding Signals Observed in p.P118P c |
|---|---|---|
| 601.22+ | 399SJGGGVVTR407 + deoxyHex.Hex (TSR1) | 601.22+ |
| 765.82+ | 693GPJSVSJGAGLR704 + deoxyHex.Hex (TSR2) | 765.82+ |
| 1380.04+ | 717ELVETVQJQGSQQPPAWPEAJVLEPJPPYWAVGDFGS ASJGGGLR763 + deoxyHex.Hex (TSR3) | 1380.04+ |
| 857.03+ | 889TGAQAAHVWTPVAGSJSVSJGR910+ deoxyHex.Hex (TSR5) | 857.03+ |
| 695.82+ | 959LAAJSVSJGR968 + deoxyHex.Hex (TSR6) | 695.82+ |
| 1017.42+ | 1018VMSLGPJSASJGLGTAR1034 + deoxyHex.Hex (TSR7) | 1017.42+ |
| 859.03+ | 1076WHVGTWMEJSVSJGDGIQR1094 + deoxyHex.Hex (TSR8) | 859.03+ |
| 519.22+ | 387WSSWGPR393 + Hex | 519.22+ |
| 1065.42+ | 1166GLLFSPAPQPR1176 + NeuAc2HexHexNAc (major) | 1065.42+ |
| 832.23+ | 1166GLLFSPAPQPR1176 + NeuAc2Hex2HexNAc2 (minor) | 832.23+ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hunt, R.; Hettiarachchi, G.; Katneni, U.; Hernandez, N.; Holcomb, D.; Kames, J.; Alnifaidy, R.; Lin, B.; Hamasaki-Katagiri, N.; Wesley, A.; et al. A Single Synonymous Variant (c.354G>A [p.P118P]) in ADAMTS13 Confers Enhanced Specific Activity. Int. J. Mol. Sci. 2019, 20, 5734. https://doi.org/10.3390/ijms20225734
Hunt R, Hettiarachchi G, Katneni U, Hernandez N, Holcomb D, Kames J, Alnifaidy R, Lin B, Hamasaki-Katagiri N, Wesley A, et al. A Single Synonymous Variant (c.354G>A [p.P118P]) in ADAMTS13 Confers Enhanced Specific Activity. International Journal of Molecular Sciences. 2019; 20(22):5734. https://doi.org/10.3390/ijms20225734
Chicago/Turabian StyleHunt, Ryan, Gaya Hettiarachchi, Upendra Katneni, Nancy Hernandez, David Holcomb, Jacob Kames, Redab Alnifaidy, Brian Lin, Nobuko Hamasaki-Katagiri, Aaron Wesley, and et al. 2019. "A Single Synonymous Variant (c.354G>A [p.P118P]) in ADAMTS13 Confers Enhanced Specific Activity" International Journal of Molecular Sciences 20, no. 22: 5734. https://doi.org/10.3390/ijms20225734
APA StyleHunt, R., Hettiarachchi, G., Katneni, U., Hernandez, N., Holcomb, D., Kames, J., Alnifaidy, R., Lin, B., Hamasaki-Katagiri, N., Wesley, A., Kafri, T., Morris, C., Bouché, L., Panico, M., Schiller, T., Ibla, J., Bar, H., Ismail, A., Morris, H., ... Kimchi-Sarfaty, C. (2019). A Single Synonymous Variant (c.354G>A [p.P118P]) in ADAMTS13 Confers Enhanced Specific Activity. International Journal of Molecular Sciences, 20(22), 5734. https://doi.org/10.3390/ijms20225734