The Anti-Tumorigenic Activity of Sema3C in the Chick Embryo Chorioallantoic Membrane Model

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
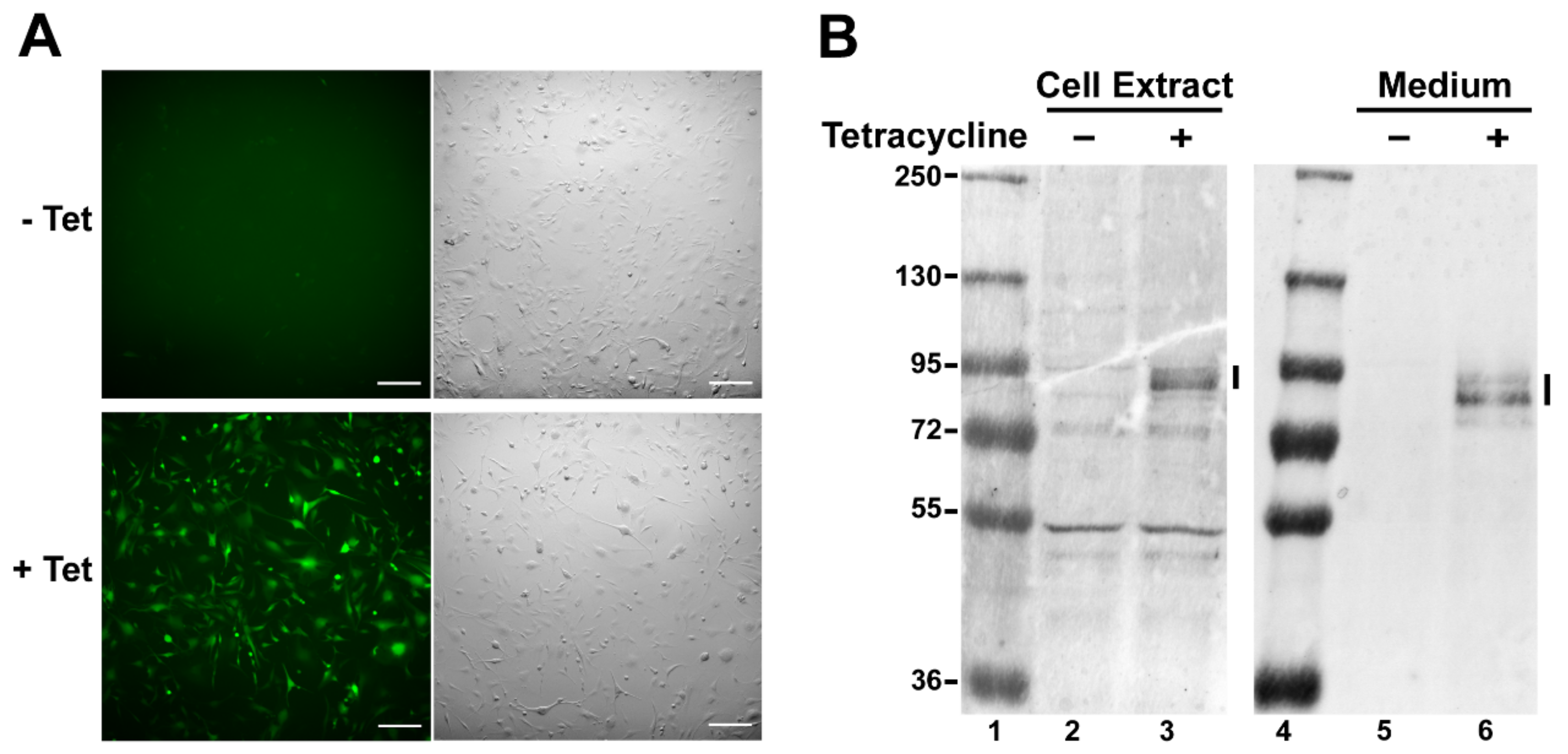
2.1. Construction and Verification of the U87t-Sema3C Cell Line
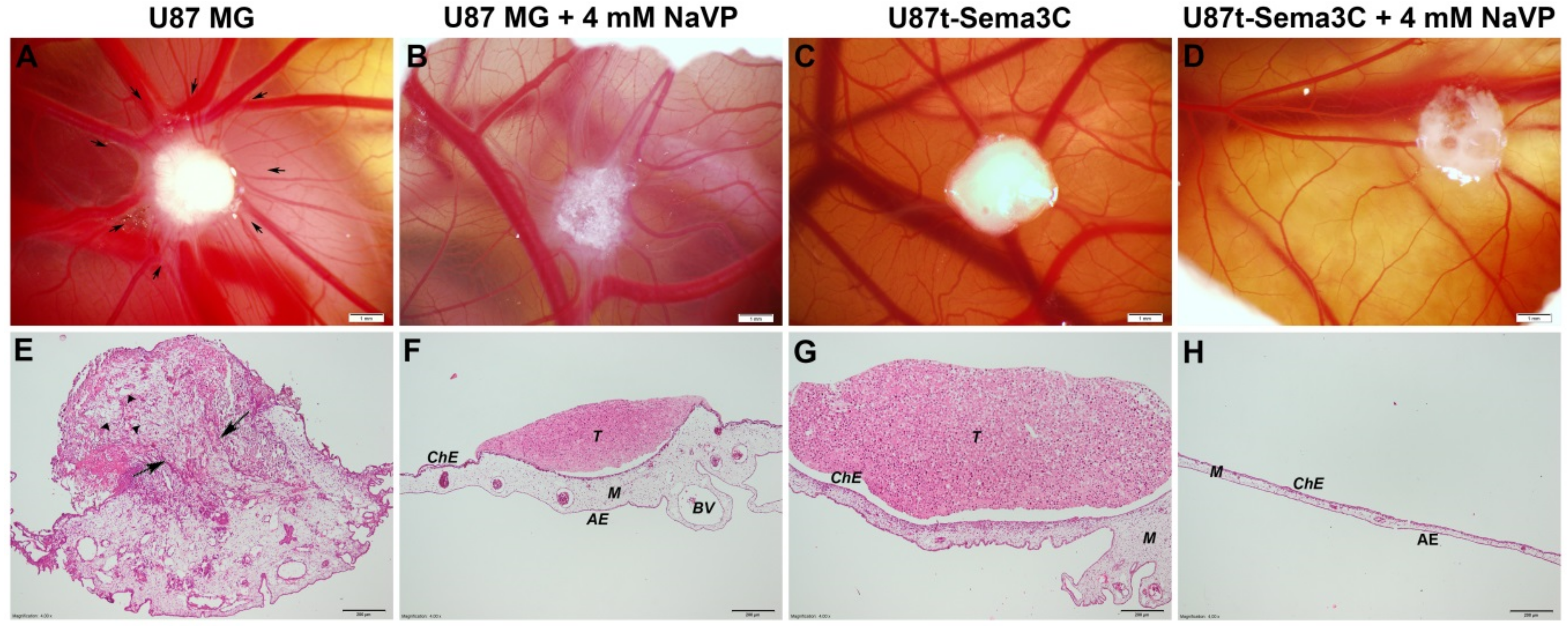
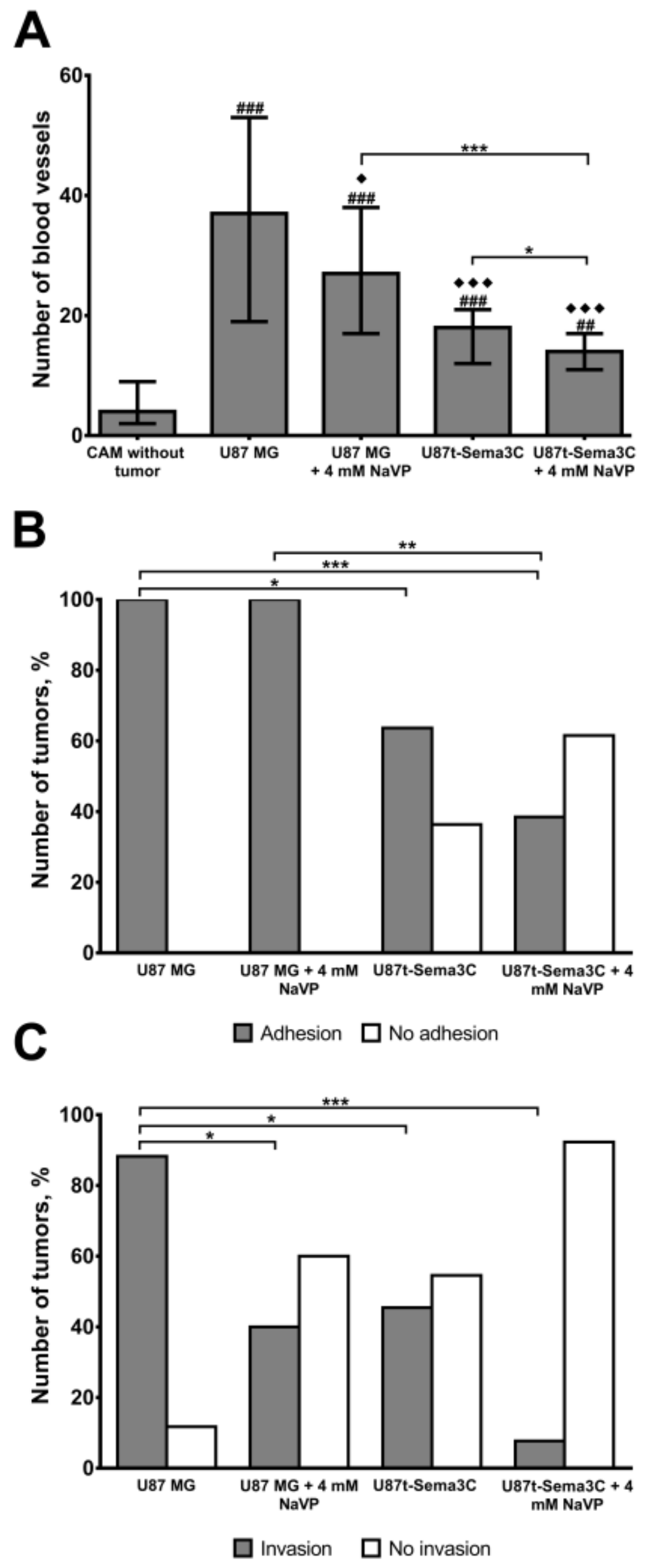
2.2. Sema3C and NaVP Effects on Vascularization and Invasiveness of U87 Cell-Formed Tumors
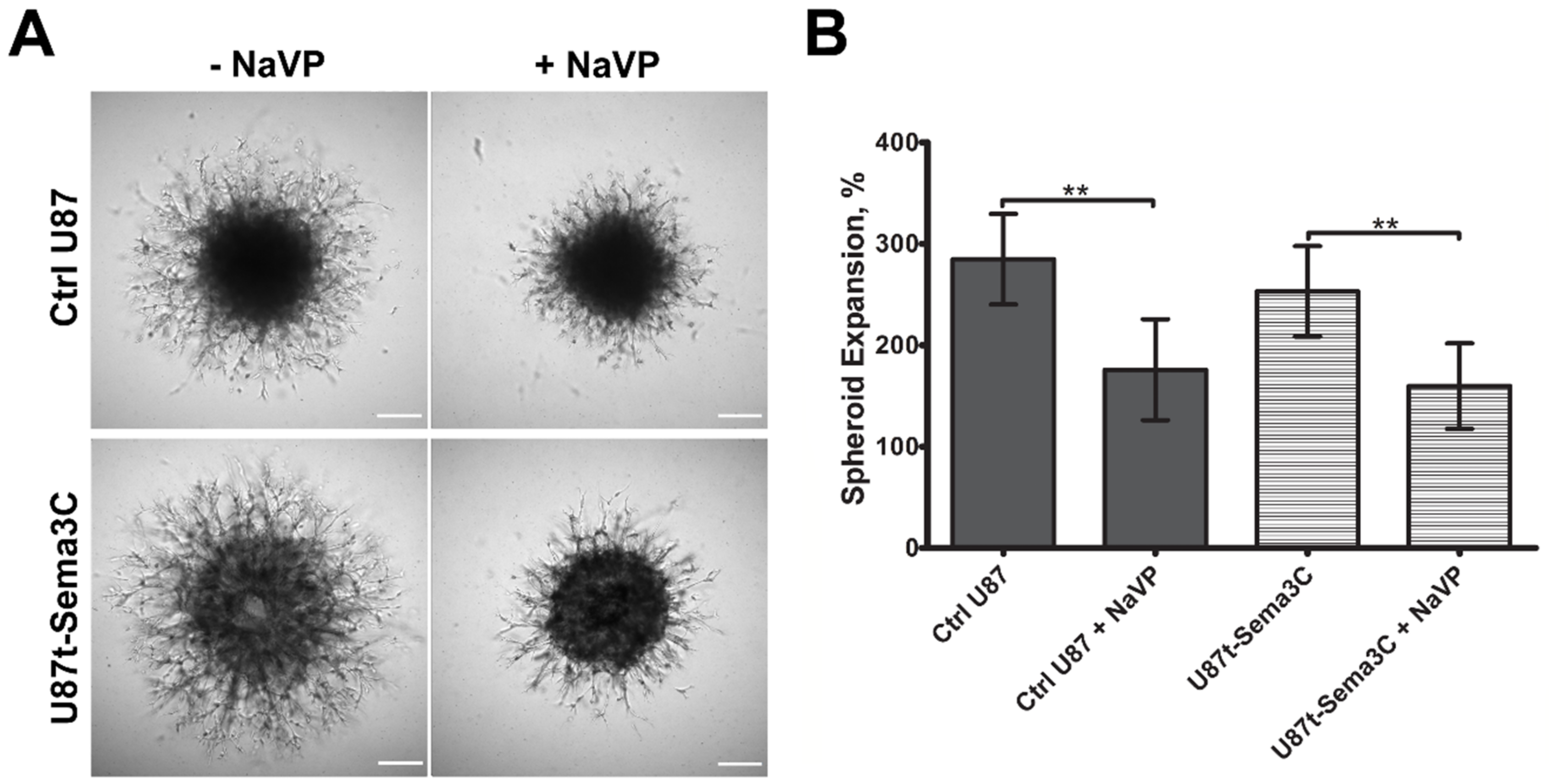
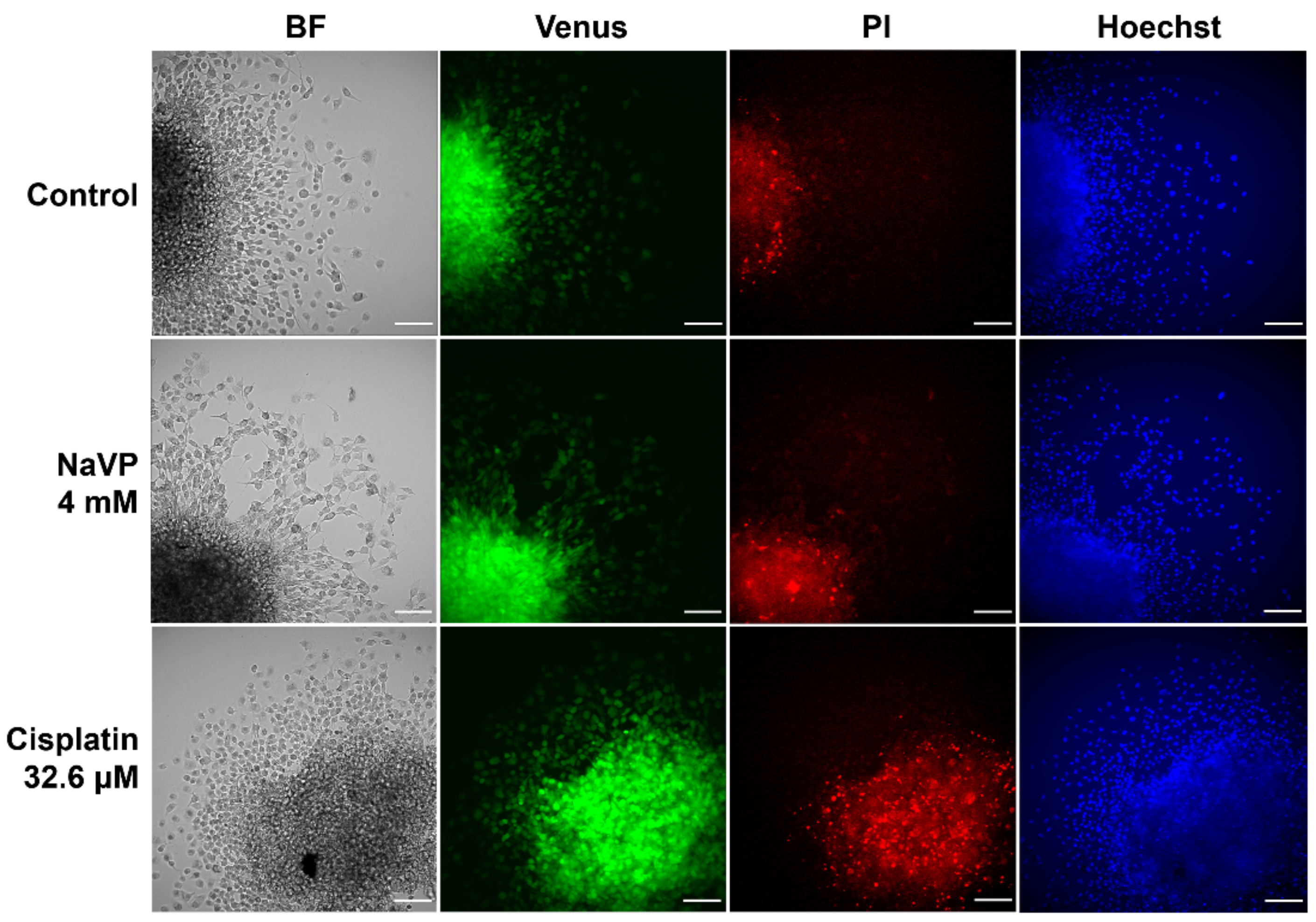
2.3. Effects of Sema3C and NaVP on Glioblastoma U87 Cell Invasion In Vitro
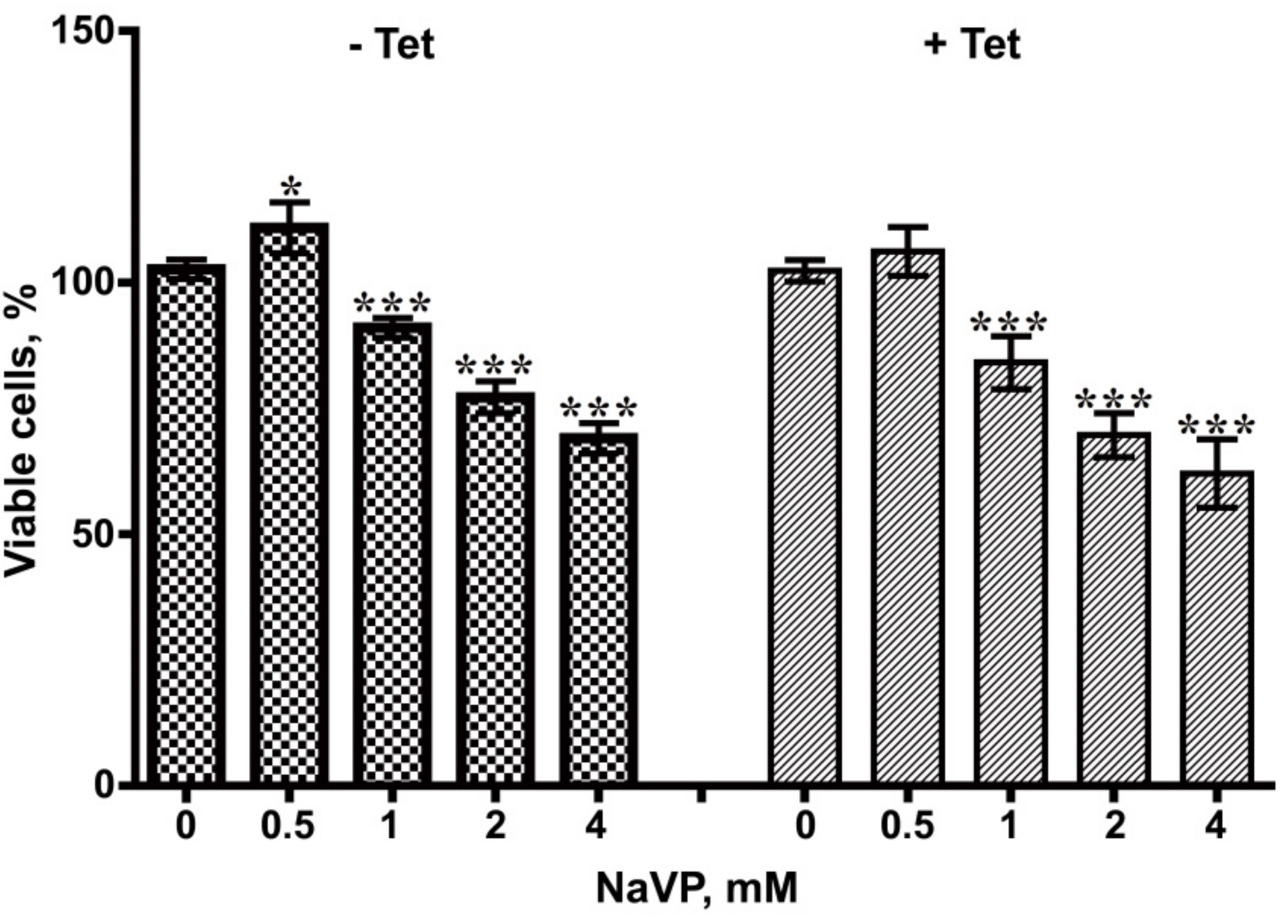
2.4. Effects of NaVP on U87t-Sema3C Cell Viability and Proliferation
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Chemicals
4.2. Generation of Tetracycline-Inducible U87t-Sema3C Cell Line
4.3. Western Blot Analysis
4.4. In Ovo CAM Model and Study Groups
4.5. The Placement of U87 MG and U87t-Sema3C Cells onto the CAM
4.6. Biomicroscopy In Vivo and Histomorphometric Assay
4.7. Assessment of Tumor Adhesion and Invasion into the CAM Mesenchyme
4.8. Cell Viability/Proliferation Assay
4.9. Cell Invasion Assay
4.10. Cell Viability Assay
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CAM | Chicken Embryo Chorioallantoic Membrane |
| NaVP | Sodium Valproate |
| Sema3 | Class 3 Semaphorin |
| GBM | Glioblastoma |
References
- Yazdani, U.; Terman, J.R. The Semaphorins. Genome Biol. 2006, 7, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Alto, L.; Terman, J. Semaphorins and Their Signaling Mechanisms. Methods Mol. Biol. 2017, 1493, 1–25. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Neufeld, G.; Mumblat, Y.; Smolkin, T.; Toledano, S.; Nir-Zvi, I.; Ziv, K.; Kessler, O. The Role of the Semaphorins in Cancer. Cell Adh. Migr. 2016, 10, 652–674. [Google Scholar] [CrossRef] [PubMed]
- Toledano, S.; Nir-Zvi, I.; Engelman, R.; Kessler, O.; Neufeld, G. Class-3 Semaphorins and Their Receptors: Potent Multifunctional Modulators of Tumor Progression. Int. J. Mol. Sci. 2019, 20, 556–576. [Google Scholar] [CrossRef] [PubMed]
- Nasarre, P.; Gemmill, R.M.; Drabkin, H.A. The Emerging Role of Class-3 Semaphorins and Their Neuropilin Receptors in Oncology. Onco. Targets. Ther. 2014, 7, 1663–1687. [Google Scholar] [CrossRef] [PubMed]
- Hui, D.H.F.; Tam, K.J.; Jiao, I.Z.F.; Ong, C.J. Semaphorin 3C as a Therapeutic Target in Prostate and Other Cancers. Int. J. Mol. Sci. 2019, 20, 774. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.; Yu, J.S. Semaphorin 3C and Its Receptors in Cancer and Cancer Stem-Like Cells. Biomedicines 2018, 6, 42. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.-J.; Hu, J.; Uemura, A.; Tetzlaff, F.; Augustin, H.G.; Fischer, A. Semaphorin-3C Signals through Neuropilin-1 and PlexinD1 Receptors to Inhibit Pathological Angiogenesis. EMBO Mol. Med. 2015, 7, 1267–1284. [Google Scholar] [CrossRef] [PubMed]
- Mumblat, Y.; Kessler, O.; Ilan, N.; Neufeld, G. Full-Length Semaphorin-3C Is an Inhibitor of Tumor Lymphangiogenesis and Metastasis. Cancer Res. 2015, 75, 2177–2186. [Google Scholar] [CrossRef] [PubMed]
- Toledano, S.; Lu, H.; Palacio, A.; Ziv, K.; Kessler, O.; Schaal, S.; Neufeld, G.; Barak, Y. A Sema3C Mutant Resistant to Cleavage by Furin (FR-Sema3C) Inhibits Choroidal Neovascularization. PLoS ONE 2016, 11, e0168122. [Google Scholar] [CrossRef] [PubMed]
- Valiulyte, I.; Preitakaite, V.; Tamasauskas, A.; Kazlauskas, A. Importance of the Putative Furin Recognition Site 742RNRR745 for Antiangiogenic Sema3C Activity in Vitro. Brazilian J. Med. Biol. Res. 2018, 51, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Vaitkienė, P.; Skiriutė, D.; Steponaitis, G.; Skauminas, K.; Tamašauskas, A. High Level of Sema3C Is Associated with Glioma Malignancy. Diagn. Pathol. 2015, 10, 58. [Google Scholar] [CrossRef] [PubMed]
- Miyato, H.; Tsuno, N.H.; Kitayama, J. Semaphorin 3C Is Involved in the Progression of Gastric Cancer. Cancer Sci. 2012, 103, 1961–1966. [Google Scholar] [CrossRef] [PubMed]
- Man, J.; Shoemake, J.; Zhou, W.; Fang, X.; Wu, Q.; Rizzo, A.; Prayson, R.; Bao, S.; Rich, J.N.; Yu, J.S. Sema3C Promotes the Survival and Tumorigenicity of Glioma Stem Cells through Rac1 Activation. Cell Rep. 2014, 9, 1812–1826. [Google Scholar] [CrossRef] [PubMed]
- Takano, S.; Yamashita, T.; Ohneda, O. Molecular Therapeutic Targets for Glioma Angiogenesis. J. Oncol. 2010, 2010, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kavaliauskaitė, D.; Stakišaitis, D.; Martinkutė, J.; Šlekienė, L.; Kazlauskas, A.; Balnytė, I.; Lesauskaitė, V.; Valančiūtė, A. The Effect of Sodium Valproate on the Glioblastoma U87 Cell Line Tumor Development on the Chicken Embryo Chorioallantoic Membrane and on EZH2 and P53 Expression. Biomed. Res. Int. 2017, 2017, 6326053. [Google Scholar] [CrossRef] [PubMed]
- Lu, V.M.; Texakalidis, P.; McDonald, K.L.; Mekary, R.A.; Smith, T.R. The Survival Effect of Valproic Acid in Glioblastoma and Its Current Trend: A Systematic Review and Meta-Analysis. Clin. Neurol. Neurosurg. 2018, 174, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Shaloam, D.; Tchounwou, P.B. Cisplatin in Cancer Therapy: Molecular Mechanisms of Action. Eur. J. Pharmacol. 2014, 740, 364–378. [Google Scholar] [CrossRef]
- Mba, G.M.P.; Naunton, M. Valproate: A Simple Chemical with so Much to Offer. J. Clin. Pharm. Ther. 2005, 30, 417–421. [Google Scholar]
- Spina, E.; Perugi, G. Antiepileptic Drugs: Indications Other than Epilepsy. Epileptic. Disord. 2004, 6, 57–75. [Google Scholar] [PubMed]
- Blaheta, R.A.; Michaelis, M.; Jr, J.C.; Herna, P.; Goethe-universita, J.W.; Goethe-universita, J.W. Evolving Anticancer Drug Valproic Acid: Insights into the Mechanism and Clinical Studies. Med. Res. Rev. 2005, 25, 383–397. [Google Scholar] [CrossRef] [PubMed]
- Moradzadeh, M.; Tabarraei, A.; Sadeghnia, H. The Role of Histone Deacetylase (HDAC) as a Biomarker in Cancer. J. Mol. Biomark. Diagn. 2015, 6, 1–4. [Google Scholar] [CrossRef]
- Seto, E.; Yoshida, M. Erasers of Histone Acetylation: The Histone Deacetylase Enzymes. Cold Spring Harb. Perspect. Biol. 2014, 6, a018713. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Ryu, H.; Won, H.; Kwon, S.H. Advances in Epigenetic Glioblastoma Therapy. Oncotarget 2017, 8, 18577–18589. [Google Scholar] [CrossRef] [PubMed]
- Phiel, C.J.; Zhang, F.; Huang, E.Y.; Guenther, M.G.; Lazar, M.A.; Klein, P.S. Histone Deacetylase Is a Direct Target of Valproic Acid, a Potent Anticonvulsant, Mood Stabilizer, and Teratogen*. J. Biol. Chem. 2001, 276, 36734–36741. [Google Scholar] [CrossRef] [PubMed]
- Das, C.; Aguilera, D.; Vasquez, H.; Prasad, P.; Zhang, M.; Wolff, J.; Al, E. Valproic Acid Induces P21 and Topoisomerase-II (α/β) Expression and Synergistically Enhances Etoposide Cytotoxicity in Human Glioblastoma Cell Lines. J. Neurooncol. 2007, 85, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Thotala, D.; Karvas, R.; Engelbach, J.; Garbow, J.; Hallahan, A.; DeWees, T.; Al, E. Valproic Acid Enhances the Efficacy of Radiation Therapy by Protecting Normal Hippocampal Neurons and Sensitizing Malignant Glioblastoma Cells. Oncotarget 2015, 6, 35004–35022. [Google Scholar] [CrossRef] [PubMed]
- Tseng, J.-H.; Chen, C.-Y.; Chen, P.-C.; Hsiao, S.-H.; Fan, C.-C.; Liang, Y.-C.; Al, E. Valproic Acid Inhibits Glioblastoma Multiforme Cell Growth via Paraoxonase 2 Expression. Oncotarget 2017, 8, 14666–14679. [Google Scholar] [CrossRef] [PubMed]
- Osuka, S.; Takano, S.; Watanabe, S.; Ishikawa, E.; Yamamoto, T.; Matsumura, A. Valproic Acid Inhibits Angiogenesis In Vitro and Glioma Angiogenesis In Vivo in the Brain. Neurol. Med. Chir. 2012, 52, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-Y.; Lai, H.-Y.; Chiu, A.; Chan, S.-H.; Hsiao, L.-P.; Lee, S.-T. The Effects of Antiepileptic Drugs on the Growth of Glioblastoma Cell Lines. J. Neurooncol. 2016, 127, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Garcia, C.G.; Romano, I.; Domith, I.; Silva, D.C.L.E.; Cossenza, M.; Kahn, S.A.; Geraldo, L.H.M.; dos Santos Assunção, F.; Romão, L.F.; Lima, F.R.S.; et al. Combination Therapy with Sulfasalazine and Valproic Acid Promotes Human Glioblastoma Cell Death Through Imbalance of the Intracellular Oxidative Response. Mol. Neurobiol. 2018, 55, 6816–6833. [Google Scholar] [CrossRef] [PubMed]
- Tam, K.J.; Hui, D.H.F.; Lee, W.W.; Dong, M.; Tombe, T.; Jiao, I.Z.F.; Khosravi, S.; Takeuchi, A.; Peacock, J.W.; Ivanova, L.; et al. Semaphorin 3 C Drives Epithelial-to-Mesenchymal Transition, Invasiveness, and Stem-like Characteristics in Prostate Cells. Sci. Rep. 2017, 7, 11501–11513. [Google Scholar] [CrossRef] [PubMed]
- Qi, G.; Lu, G.; Yu, J.; Zhao, Y.; Wang, C.; Zhang, H.; Xia, Q. Up-Regulation of TIF1γ by Valproic Acid Inhibits the Epithelial Mesenchymal Transition in Prostate Carcinoma through TGF-β/Smad Signaling Pathway. Eur. J. Pharmacol. 2019, 860, 172551. [Google Scholar] [CrossRef] [PubMed]
- Karayan-Tapon, L.; Wager, M.; Guilhot, J.; Levillain, P.; Marquant, C.; Clarhaut, J.; Potiron, V.; Roche, J. Semaphorin, Neuropilin and VEGF Expression in Glial Tumours: SEMA3G, a Prognostic Marker? Br. J. Cancer 2008, 99, 1153–1160. [Google Scholar] [CrossRef] [PubMed]
- Peacock, J.W.; Takeuchi, A.; Hayashi, N.; Liu, L.; Tam, K.J.; Al Nakouzi, N.; Khazamipour, N.; Tombe, T.; Dejima, T.; Lee, K.C.; et al. SEMA3C Drives Cancer Growth by Transactivating Multiple Receptor Tyrosine Kinases via Plexin B1. EMBO Mol. Med. 2018, 10, 219–238. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zhao, Z.; Guo, S.; Li, J.; Liu, S.; You, Y.; Ni, B.; Wang, H.; Bie, P. Increased semaphorin 3c expression promotes tumor growth and metastasis in pancreatic ductal adenocarcinoma by activating the ERK1/2 signaling pathway. Cancer Lett. 2017, 397, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carrera, I.; Frise, E.; Verena, K.; Mark, L.; Tobias, P.; Stephan, P.; Curtis, R.; Stephan, S.; Benjamin, S.; et al. Fiji—An Open Platform for Biological Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valiulytė, I.; Curkūnavičiūtė, R.; Ribokaitė, L.; Kazlauskas, A.; Vaitkevičiūtė, M.; Skauminas, K.; Valančiūtė, A. The Anti-Tumorigenic Activity of Sema3C in the Chick Embryo Chorioallantoic Membrane Model. Int. J. Mol. Sci. 2019, 20, 5672. https://doi.org/10.3390/ijms20225672
Valiulytė I, Curkūnavičiūtė R, Ribokaitė L, Kazlauskas A, Vaitkevičiūtė M, Skauminas K, Valančiūtė A. The Anti-Tumorigenic Activity of Sema3C in the Chick Embryo Chorioallantoic Membrane Model. International Journal of Molecular Sciences. 2019; 20(22):5672. https://doi.org/10.3390/ijms20225672
Chicago/Turabian StyleValiulytė, Indrė, Rūta Curkūnavičiūtė, Laura Ribokaitė, Arunas Kazlauskas, Monika Vaitkevičiūtė, Kęstutis Skauminas, and Angelija Valančiūtė. 2019. "The Anti-Tumorigenic Activity of Sema3C in the Chick Embryo Chorioallantoic Membrane Model" International Journal of Molecular Sciences 20, no. 22: 5672. https://doi.org/10.3390/ijms20225672
APA StyleValiulytė, I., Curkūnavičiūtė, R., Ribokaitė, L., Kazlauskas, A., Vaitkevičiūtė, M., Skauminas, K., & Valančiūtė, A. (2019). The Anti-Tumorigenic Activity of Sema3C in the Chick Embryo Chorioallantoic Membrane Model. International Journal of Molecular Sciences, 20(22), 5672. https://doi.org/10.3390/ijms20225672