Tissue-Wide Gene Expression Analysis of Sodium/Phosphate Co-Transporters in Pigs

 , , , , and
, , , , and 
Abstract
1. Introduction
2. Results and Discussion
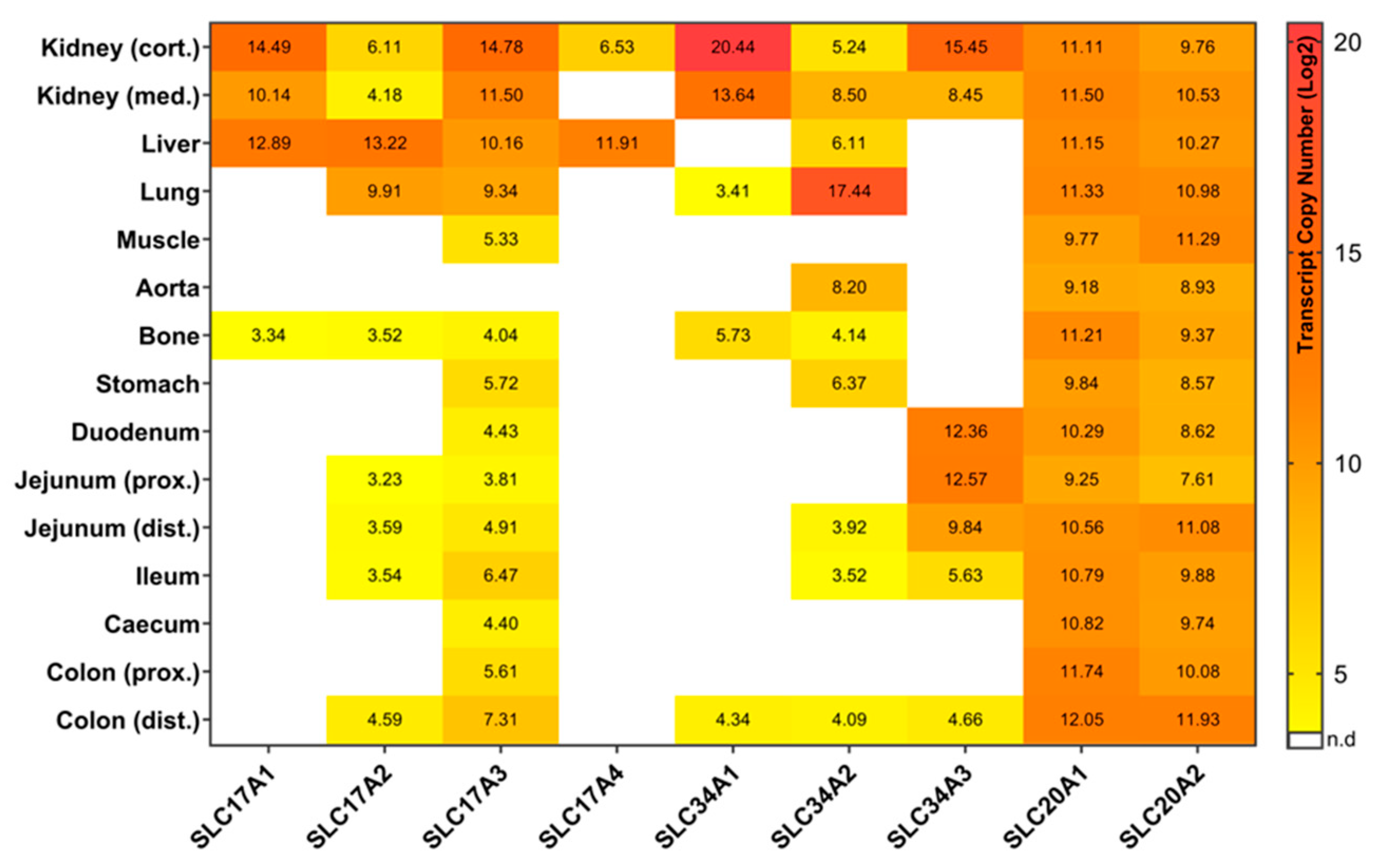
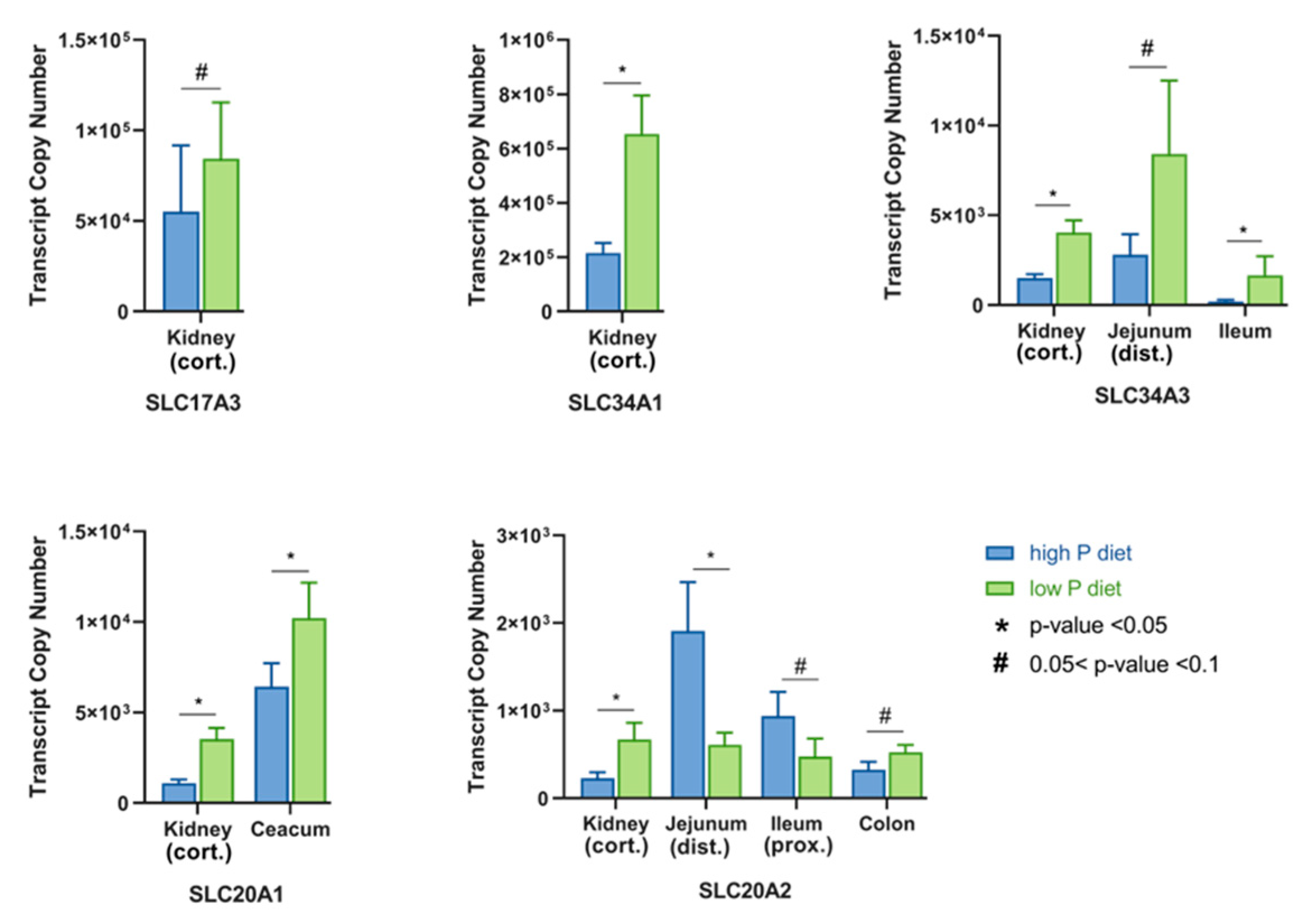
2.1. Type I Sodium/Phosphate Co-Transporters Expression Analysis
2.2. Type II Sodium/Phosphate Co-Transporters Expression Analysis
2.3. Type III Sodium/Phosphate Co-Transporters Expression Analysis
3. Materials and Methods
3.1. Animals
3.2. Tissue Sampling
3.3. RNA Isolation and cDNA Synthesis
3.4. Quantitative Real-Time PCR (qRT-PCR)
3.5. Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| P | Phosphorus |
| qRT-PCR | Quantitative real time polymerase chain reaction |
| SLC | Solute carrier |
References
- Pavlov, E.; Aschar-Sobbi, R.; Campanella, M.; Turner, R.J.; Gomez-Garcia, M.R.; Abramov, A.Y. Inorganic polyphosphate and energy metabolism in mammalian cells. J. Biol. Chem. 2010, 285, 9420–9428. [Google Scholar] [CrossRef]
- Oster, M.; Reyer, H.; Ball, E.; Fornara, D.; McKillen, J.; Sorensen, K.U.; Poulsen, H.D.; Andersson, K.; Ddiba, D.; Rosemarin, A.; et al. Bridging Gaps in the Agricultural Phosphorus Cycle from an Animal Husbandry Perspective- The Case of Pigs and Poultry. Sustainability 2018, 10, 1825. [Google Scholar] [CrossRef]
- Campbell, B.M.; Beare, D.J.; Bennett, E.M.; Hall-Spencer, J.M.; Ingram, J.S.I.; Jaramillo, F.; Ortiz, R.; Ramankutty, N.; Sayer, J.A.; Shindell, D. Agriculture production as a major driver of the Earth system exceeding planetary boundaries. Ecol. Soc. 2017, 22. [Google Scholar] [CrossRef]
- Pokharel, B.B.; Regassa, A.; Nyachoti, C.M.; Kim, W.K. Effect of low levels of dietary available phosphorus on phosphorus utilization, bone mineralization, phosphorus transporter mRNA expression and performance in growing pigs. J. Env. Sci. Health B 2017, 52, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Alexander, L.S.; Qu, A.; Cutler, S.A.; Mahajan, A.; Lonergan, S.M.; Rothschild, M.F.; Weber, T.E.; Kerr, B.J.; Stahl, C.H. Response to dietary phosphorus deficiency is affected by genetic background in growing pigs. J. Anim. Sci. 2008, 86, 2585–2595. [Google Scholar] [CrossRef] [PubMed]
- Reyer, H.; Oster, M.; Wittenburg, D.; Murani, E.; Ponsuksili, S.; Wimmers, K. Genetic Contribution to Variation in Blood Calcium, Phosphorus, and Alkaline Phosphatase Activity in Pigs. Front. Genet. 2019, 10, 590. [Google Scholar] [CrossRef]
- Beck, L.; Karaplis, A.C.; Amizuka, N.; Hewson, A.S.; Ozawa, H.; Tenenhouse, H.S. Targeted inactivation of Npt2 in mice leads to severe renal phosphate wasting, hypercalciuria, and skeletal abnormalities. Proc. Natl. Acad. Sci. USA 1998, 95, 5372–5377. [Google Scholar] [CrossRef]
- Sabbagh, Y.; O'Brien, S.P.; Song, W.P.; Boulanger, J.H.; Stockmann, A.; Arbeeny, C.; Schiavi, S.C. Intestinal Npt2b Plays a Major Role in Phosphate Absorption and Homeostasis. J. Am. Soc. Nephrol. 2009, 20, 2348–2358. [Google Scholar] [CrossRef]
- Iwaki, T.; Sandoval-Cooper, M.J.; Tenenhouse, H.S.; Castellino, F.J. A missense mutation in the sodium phosphate co-transporter Slc34a1 impairs phosphate homeostasis. J. Am. Soc. Nephrol. 2008, 19, 1753–1762. [Google Scholar] [CrossRef]
- Ichikawa, S.; Sorenson, A.H.; Imel, E.A.; Friedman, N.E.; Gertner, J.M.; Econs, M.J. Intronic deletions in the SLC34A3 gene cause hereditary hypophosphatemic rickets with hypercalciuria. J. Clin. Endocrinol. Metab. 2006, 91, 4022–4027. [Google Scholar] [CrossRef]
- Bergwitz, C.; Juppner, H. Regulation of phosphate homeostasis by PTH, vitamin D, and FGF23. Annu. Rev. Med. 2010, 61, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Gattineni, J.; Bates, C.; Twombley, K.; Dwarakanath, V.; Robinson, M.L.; Goetz, R.; Mohammadi, M.; Baum, M. FGF23 decreases renal NaPi-2a and NaPi-2c expression and induces hypophosphatemia in vivo predominantly via FGF receptor 1. Am. J. Physiol. Ren. Physiol. 2009, 297, F282–F291. [Google Scholar] [CrossRef] [PubMed]
- Werner, A.; Moore, M.L.; Mantei, N.; Biber, J.; Semenza, G.; Murer, H. Cloning and Expression of Cdna for a Na/Pi Cotransport System of Kidney Cortex. Proc. Natl. Acad. Sci. USA 1991, 88, 9608–9612. [Google Scholar] [CrossRef] [PubMed]
- Murer, H.; Forster, I.; Biber, J. The sodium phosphate cotransporter family SLC34. Pflug. Arch. 2004, 447, 763–767. [Google Scholar] [CrossRef] [PubMed]
- Collins, J.F.; Bai, L.; Ghishan, F.K. The SLC20 family of proteins: Dual functions as sodium-phosphate cotransporters and viral receptors. Pflug. Arch. 2004, 447, 647–652. [Google Scholar] [CrossRef]
- Yabuuchi, H.; Tamai, I.; Morita, K.; Kouda, T.; Miyamoto, K.; Takeda, E.; Tsuji, A. Hepatic sinusoidal membrane transport of anionic drugs mediated by anion transporter Npt1. J. Pharm. Exp. 1998, 286, 1391–1396. [Google Scholar]
- Chong, S.S.; Kristjansson, K.; Zoghbi, H.Y.; Hughes, M.R. Molecular cloning of the cDNA encoding a human renal sodium phosphate transport protein and its assignment to chromosome 6p21.3-p23. Genomics 1993, 18, 355–359. [Google Scholar] [CrossRef]
- Miyamoto, K.; Tatsumi, S.; Sonoda, T.; Yamamoto, H.; Minami, H.; Taketani, Y.; Takeda, E. Cloning and functional expression of a Na(+)-dependent phosphate co-transporter from human kidney: cDNA cloning and functional expression. Biochem. J. 1995, 305, 81–85. [Google Scholar] [CrossRef]
- Chiba, T.; Matsuo, H.; Kawamura, Y.; Nagamori, S.; Nishiyama, T.; Wei, L.; Nakayama, A.; Nakamura, T.; Sakiyama, M.; Takada, T.; et al. NPT1/SLC17A1 is a renal urate exporter in humans and its common gain-of-function variant decreases the risk of renal underexcretion gout. Arthritis Rheumatol. 2015, 67, 281–287. [Google Scholar] [CrossRef]
- Younus, M.J.; Butterworth, P.J. Sodium-dependent transport of phosphate by rat liver plasma membrane vesicles. Biochim. Biophys. Acta 1993, 1143, 158–162. [Google Scholar] [CrossRef]
- Ghishan, F.K.; Dykes, W. Ontogeny of phosphate transport by rat liver plasma membrane vesicles. J. Dev. Physiol. 1993, 19, 197–201. [Google Scholar] [PubMed]
- Murer, H.; Hernando, N.; Forster, I.; Biber, J. Proximal tubular phosphate reabsorption: Molecular mechanisms. Physiol. Rev. 2000, 80, 1373–1409. [Google Scholar] [CrossRef] [PubMed]
- Busch, A.E.; Schuster, A.; Waldegger, S.; Wagner, C.A.; Zempel, G.; Broer, S.; Biber, J.; Murer, H.; Lang, F. Expression of a renal type I sodium/phosphate transporter (NaPi-1) induces a conductance in Xenopus oocytes permeable for organic and inorganic anions. Proc. Natl. Acad. Sci. USA 1996, 93, 5347–5351. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, S.E.; Gunther, W.; Pearce, S.H.; Thomson, A.; Bianchi, M.L.; Bosio, M.; Craig, I.W.; Fisher, S.E.; Scheinman, S.J.; Wrong, O.; et al. Characterisation of renal chloride channel, CLCN5, mutations in hypercalciuric nephrolithiasis (kidney stones) disorders. Hum. Mol. Genet. 1997, 6, 1233–1239. [Google Scholar] [CrossRef]
- Iharada, M.; Miyaji, T.; Fujimoto, T.; Hiasa, M.; Anzai, N.; Omote, H.; Moriyama, Y. Type 1 Sodium-dependent Phosphate Transporter (SLC17A1 Protein) Is a Cl(-)-dependent Urate Exporter. J. Biol. Chem. 2010, 285, 26107–26113. [Google Scholar] [CrossRef]
- Miyamoto, K.; Haito-Sugino, S.; Kuwahara, S.; Ohi, A.; Nomura, K.; Ito, M.; Kuwahata, M.; Kido, S.; Tatsumi, S.; Kaneko, I.; et al. Sodium-dependent phosphate cotransporters: Lessons from gene knockout and mutation studies. J. Pharm. Sci. 2011, 100, 3719–3730. [Google Scholar] [CrossRef]
- Togawa, N.; Juge, N.; Miyaji, T.; Hiasa, M.; Omote, H.; Moriyama, Y. Wide expression of type I Na+-phosphate cotransporter 3 (NPT3/SLC17A2), a membrane potential-driven organic anion transporter. Am. J. Physiol. Cell Physiol. 2015, 309, C71–C80. [Google Scholar] [CrossRef]
- Sugita, A.; Hayashibara, T.; Yoneda, T. [Role of the type 3 sodium-dependent phosphate transporter in the calcification of growth plate chondrocytes]. Clin. Calcium. 2006, 16, 1723–1729. [Google Scholar]
- Pontén, F.; Jirström, K.; Uhlen, M. The Human Protein Atlas—a tool for pathology. J. Pathol. 2008, 216, 387–393. [Google Scholar] [CrossRef]
- Jutabha, P.; Anzai, N.; Wempe, M.F.; Wakui, S.; Endou, H.; Sakurai, H. Apical voltage-driven urate efflux transporter NPT4 in renal proximal tubule. Nucleosides Nucleotides Nucleic Acids 2011, 30, 1302–1311. [Google Scholar] [CrossRef]
- Jutabha, P.; Anzai, N.; Kitamura, K.; Taniguchi, A.; Kaneko, S.; Yan, K.; Yamada, H.; Shimada, H.; Kimura, T.; Katada, T.; et al. Human sodium phosphate transporter 4 (hNPT4/SLC17A3) as a common renal secretory pathway for drugs and urate. J. Biol. Chem. 2010, 285, 35123–35132. [Google Scholar] [CrossRef] [PubMed]
- Togawa, N.; Miyaji, T.; Izawa, S.; Omote, H.; Moriyama, Y. A Na+-phosphate cotransporter homologue (SLC17A4 protein) is an intestinal organic anion exporter. Am. J. Physiol. Cell Physiol. 2012, 302, C1652–C1660. [Google Scholar] [CrossRef] [PubMed]
- Sreedharan, S.; Shaik, J.H.; Olszewski, P.K.; Levine, A.S.; Schioth, H.B.; Fredriksson, R. Glutamate, aspartate and nucleotide transporters in the SLC17 family form four main phylogenetic clusters: Evolution and tissue expression. BMC Genom. 2010, 11, 17. [Google Scholar] [CrossRef] [PubMed]
- Murer, H.; Hernando, N.; Forster, A.; Biber, J. Molecular aspects in the regulation of renal inorganic phosphate reabsorption: The type IIa sodium/inorganic phosphate co-transporter as the key player. Curr. Opin. Nephrol. Hypertens. 2001, 10, 555–561. [Google Scholar] [CrossRef]
- Ohkido, I.; Segawa, H.; Yanagida, R.; Nakamura, M.; Miyamoto, K. Cloning, gene structure and dietary regulation of the type-IIc Na/Pi cotransporter in the mouse kidney. Pflug. Arch. 2003, 446, 106–115. [Google Scholar] [CrossRef]
- Oster, M.; Just, F.; Busing, K.; Wolf, P.; Polley, C.; Vollmar, B.; Murani, E.; Ponsuksili, S.; Wimmers, K. Toward improved phosphorus efficiency in monogastrics-interplay of serum, minerals, bone, and immune system after divergent dietary phosphorus supply in swine. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 310, R917–R925. [Google Scholar] [CrossRef]
- Xu, H.; Bai, L.; Collins, J.F.; Ghishan, F.K. Molecular cloning, functional characterization, tissue distribution, and chromosomal localization of a human, small intestinal sodium-phosphate (Na+-Pi) transporter (SLC34A2). Genomics 1999, 62, 281–284. [Google Scholar] [CrossRef]
- Hashimoto, M.; Wang, D.Y.; Kamo, T.; Zhu, Y.; Tsujiuchi, T.; Konishi, Y.; Tanaka, M.; Sugimura, H. Isolation and localization of type IIb Na/Pi cotransporter in the developing rat lung. Am. J. Pathol. 2000, 157, 21–27. [Google Scholar] [CrossRef]
- Feild, J.A.; Zhang, L.; Brun, K.A.; Brooks, D.P.; Edwards, R.M. Cloning and functional characterization of a sodium-dependent phosphate transporter expressed in human lung and small intestine. Biochem. Biophys. Res. Commun. 1999, 258, 578–582. [Google Scholar] [CrossRef]
- Walling, M.W. Studies on Mechanisms of 1,25 Dihydroxy-Vitamin-D3-Stimulated Intestinal Phosphate Absorption. J. Dent. Res. 1977, 56, A137. [Google Scholar]
- Fang, R.; Xiang, Z.; Cao, M.; He, J. Different phosphate transport in the duodenum and jejunum of chicken response to dietary phosphate adaptation. Asian-Australas. J. Anim. Sci. 2012, 25, 1457–1465. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Yin, Y.; Wang, Z.; Shen, Y.; Squires, J.E.; Li, J.; Xu, H.; Collins, J.F.; Ghishan, F.K.; Fan, M.Z. Visceral distribution of the type II sodium-dependent phosphate cotransporter (NaPi-II) isomer mRNA and the expression of NaPi-IIc mRNA along the intestinal longitudinal axis in the post-weaned pig. FASEB J. 2006, 20, A1064. [Google Scholar] [CrossRef]
- Bergwitz, C.; Roslin, N.M.; Tieder, M.; Loredo-Osti, J.C.; Bastepe, M.; Abu-Zahra, H.; Frappier, D.; Burkett, K.; Carpenter, T.O.; Anderson, D.; et al. SLC34A3 mutations in patients with hereditary hypophosphatemic rickets with hypercalciuria predict a key role for the sodium-phosphate cotransporter NaPi-IIc in maintaining phosphate homeostasis. Am. J. Hum. Genet. 2006, 78, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Jaureguiberry, G.; Carpenter, T.O.; Forman, S.; Juppner, H.; Bergwitz, C. A novel missense mutation in SLC34A3 that causes hereditary hypophosphatemic rickets with hypercalciuria in humans identifies threonine 137 as an important determinant of sodium-phosphate cotransport in NaPi-IIc. Am. J. Physiol. Ren. Physiol. 2008, 295, F371–F379. [Google Scholar] [CrossRef] [PubMed]
- Segawa, H.; Kaneko, I.; Takahashi, A.; Kuwahata, M.; Ito, M.; Ohkido, I.; Tatsumi, S.; Miyamoto, K. Growth-related renal type II Na/Pi cotransporter. J. Biol. Chem. 2002, 277, 19665–19672. [Google Scholar] [CrossRef]
- Madjdpour, C.; Bacic, D.; Kaissling, B.; Murer, H.; Biber, J. Segment-specific expression of sodium-phosphate cotransporters NaPi-IIa and -IIc and interacting proteins in mouse renal proximal tubules. Pflug. Arch. 2004, 448, 402–410. [Google Scholar] [CrossRef][Green Version]
- Bai, L.; Collins, J.F.; Ghishan, F.K. Cloning and characterization of a type III Na-dependent phosphate cotransporter from mouse intestine. Am. J. Physiol. Cell Physiol. 2000, 279, C1135–C1143. [Google Scholar] [CrossRef]
- Tatsumi, S.; Segawa, H.; Morita, K.; Haga, H.; Kouda, T.; Yamamoto, H.; Inoue, Y.; Nii, T.; Katai, K.; Taketani, Y.; et al. Molecular cloning and hormonal regulation of PiT-1, a sodium-dependent phosphate cotransporter from rat parathyroid glands. Endocrinology 1998, 139, 1692–1699. [Google Scholar] [CrossRef][Green Version]
- Johann, S.V.; Gibbons, J.J.; O'Hara, B. GLVR1, a receptor for gibbon ape leukemia virus, is homologous to a phosphate permease of Neurospora crassa and is expressed at high levels in the brain and thymus. J. Virol. 1992, 66, 1635–1640. [Google Scholar]
- van Zeijl, M.; Johann, S.V.; Closs, E.; Cunningham, J.; Eddy, R.; Shows, T.B.; O'Hara, B. A human amphotropic retrovirus receptor is a second member of the gibbon ape leukemia virus receptor family. Proc. Natl. Acad. Sci. USA 1994, 91, 1168–1172. [Google Scholar] [CrossRef]
- Olah, Z.; Lehel, C.; Anderson, W.B.; Eiden, M.V.; Wilson, C.A. The cellular receptor for gibbon ape leukemia virus is a novel high affinity sodium-dependent phosphate transporter. J. Biol. Chem. 1994, 269, 25426–25431. [Google Scholar] [PubMed]
- Inden, M.; Iriyama, M.; Zennami, M.; Sekine, S.I.; Hara, A.; Yamada, M.; Hozumi, I. The type III transporters (PiT-1 and PiT-2) are the major sodium-dependent phosphate transporters in the mice and human brains. Brain Res. 2016, 1637, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Jensen, N.; Autzen, J.K.; Pedersen, L. Slc20a2 is critical for maintaining a physiologic inorganic phosphate level in cerebrospinal fluid. Neurogenetics 2016, 17, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Byskov, K.; Jensen, N.; Kongsfelt, I.B.; Wielsoe, M.; Pedersen, L.E.; Haldrup, C.; Pedersen, L. Regulation of cell proliferation and cell density by the inorganic phosphate transporter PiT1. Cell Div. 2012, 7, 7. [Google Scholar] [CrossRef]
- Beck, L.; Leroy, C.; Salaun, C.; Margall-Ducos, G.; Desdouets, C.; Friedlander, G. Identification of a novel function of PiT1 critical for cell proliferation and independent of its phosphate transport activity. J. Biol. Chem. 2009, 284, 31363–31374. [Google Scholar] [CrossRef]
- Salaun, C.; Leroy, C.; Rousseau, A.; Boitez, V.; Beck, L.; Friedlander, G. Identification of a novel transport-independent function of PiT1/SLC20A1 in the regulation of TNF-induced apoptosis. J. Biol. Chem. 2010, 285, 34408–34418. [Google Scholar] [CrossRef]
- Couasnay, G.; Bon, N.; Devignes, C.S.; Sourice, S.; Bianchi, A.; Veziers, J.; Weiss, P.; Elefteriou, F.; Provot, S.; Guicheux, J.; et al. PiT1/Slc20a1 is required for endoplasmic reticulum homeostasis, chondrocyte survival and skeletal development. J. Bone Min. Res. 2018, 34, 387–398. [Google Scholar] [CrossRef]
- Bon, N.; Couasnay, G.; Bourgine, A.; Sourice, S.; Beck-Cormier, S.; Guicheux, J.; Beck, L. Phosphate (Pi)-regulated heterodimerization of the high-affinity sodium-dependent Pi transporters PiT1/Slc20a1 and PiT2/Slc20a2 underlies extracellular Pi sensing independently of Pi uptake. J. Biol. Chem. 2018, 293, 2102–2114. [Google Scholar] [CrossRef]
- Gesellschaft für Ernährungsphysiologie. In Empfehlungen zur Energie-und Nährstoffversorgung beim Schwein; DLG Verlag: Frankfurt, Germany, 2006. (In German)




{kind=link}
{kind=link}
{kind=link}
| Gene | Gene Synonyms | Ensembl ID (v. 91) | Description |
|---|---|---|---|
| SLC17A1 | NAPI-1, NPT1 | ENSSSCG00000001107 | Solute carrier family 17 member 1 |
| SLC17A2 | NPT3 | ENSSSCG00000036191 | Solute carrier family 17 member 2 |
| SLC17A3 | NPT4 | ENSSSCG00000037547 | Solute carrier family 17 member 3 |
| SLC17A4 | NPT5 | ENSSSCG00000031944 | Solute carrier family 17 member 4 |
| SLC34A1 | NaPi-2a, NPT2a | ENSSSCG00000037535 | Solute carrier family 34 member 1 |
| SLC34A2 | NaPi-2b, NPT2b | ENSSSCG00000008758 | Solute carrier family 34 member 2 |
| SLC34A3 | NaPi-2c, NPT2c | ENSSSCG00000040105 | Solute carrier family 34 member 3 |
| SLC20A1 | GLVR1, Glvr-1, PiT1 | ENSSSCG00000032288 | Solute carrier family 20 member 1 |
| SLC20A2 | GLVR2, Glvr-2, MLVAR, PiT2, Ram-1 | ENSSSCG00000007027 | Solute carrier family 20 member 2 |
| Tissue | Short | Specification | Trial 1 |
|---|---|---|---|
| Kidney | Kidney cort | Cortex of left kidney | 1, 2 |
| Kidney | Kidney med | Medulla of left kidney | 1, 2 |
| Liver | Liver | Lobulus Spigelii | 1 |
| Stomach | Stomach | Fundus mucosa | 1 |
| Duodenum | Duod | Mucosa, 30–40 cm distal of pylorus | 1, 2 |
| Jejunum (prox.) | Jeju prox | Mucosa, 2 m distal of pylorus | 1, 2 |
| Jejunum (dist.) | Jeju dist | Mucosa, 2 m proximal of the ileocaecal junction | 1, 2 |
| Ileum | Ileum | Mucosa, 20 cm proximal of the ileocaecal junction | 1, 2 |
| Caecum | Caec | Mucosa | 1, 2 |
| Colon (prox.) | Colon prox | Mucosa, 50–60 cm distal of cecolic junction | 1, 2 |
| Colon (dist.) | Colon dist | Mucosa, 50–60 m proximal of rectum | 1, 2 |
| Bone | Bone | Calvarial bone, along the sagittal suture | 1 |
| Muscle | Muscle | Longissimus dorsi, between the 13th and 14th rib | 1 |
| Lung | Lung | Lower tip of the left lung lobe | 1 |
| Aorta | Aorta | Aorta, descending thoracic aorta | 1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wubuli, A.; Reyer, H.; Muráni, E.; Ponsuksili, S.; Wolf, P.; Oster, M.; Wimmers, K. Tissue-Wide Gene Expression Analysis of Sodium/Phosphate Co-Transporters in Pigs. Int. J. Mol. Sci. 2019, 20, 5576. https://doi.org/10.3390/ijms20225576
Wubuli A, Reyer H, Muráni E, Ponsuksili S, Wolf P, Oster M, Wimmers K. Tissue-Wide Gene Expression Analysis of Sodium/Phosphate Co-Transporters in Pigs. International Journal of Molecular Sciences. 2019; 20(22):5576. https://doi.org/10.3390/ijms20225576
Chicago/Turabian StyleWubuli, Aisanjiang, Henry Reyer, Eduard Muráni, Siriluck Ponsuksili, Petra Wolf, Michael Oster, and Klaus Wimmers. 2019. "Tissue-Wide Gene Expression Analysis of Sodium/Phosphate Co-Transporters in Pigs" International Journal of Molecular Sciences 20, no. 22: 5576. https://doi.org/10.3390/ijms20225576
APA StyleWubuli, A., Reyer, H., Muráni, E., Ponsuksili, S., Wolf, P., Oster, M., & Wimmers, K. (2019). Tissue-Wide Gene Expression Analysis of Sodium/Phosphate Co-Transporters in Pigs. International Journal of Molecular Sciences, 20(22), 5576. https://doi.org/10.3390/ijms20225576