Functional Dissection of the Chickpea (Cicer arietinum L.) Stay-Green Phenotype Associated with Molecular Variation at an Ortholog of Mendel’s I Gene for Cotyledon Color: Implications for Crop Production and Carotenoid Biofortification

 , , ,
, , ,  ,
,  ,
,  ,
,
Abstract
1. Introduction
2. Results
2.1. Delayed Degreening Phenotypes in Green Cotyledon Chickpea and Underlying Allelic Variation
2.1.1. Delayed Degreening Phenotypes in Green Cotyledon Chickpea
2.1.2. Identification of Chickpea Ortholog of the Staygreen (SGR) Protein
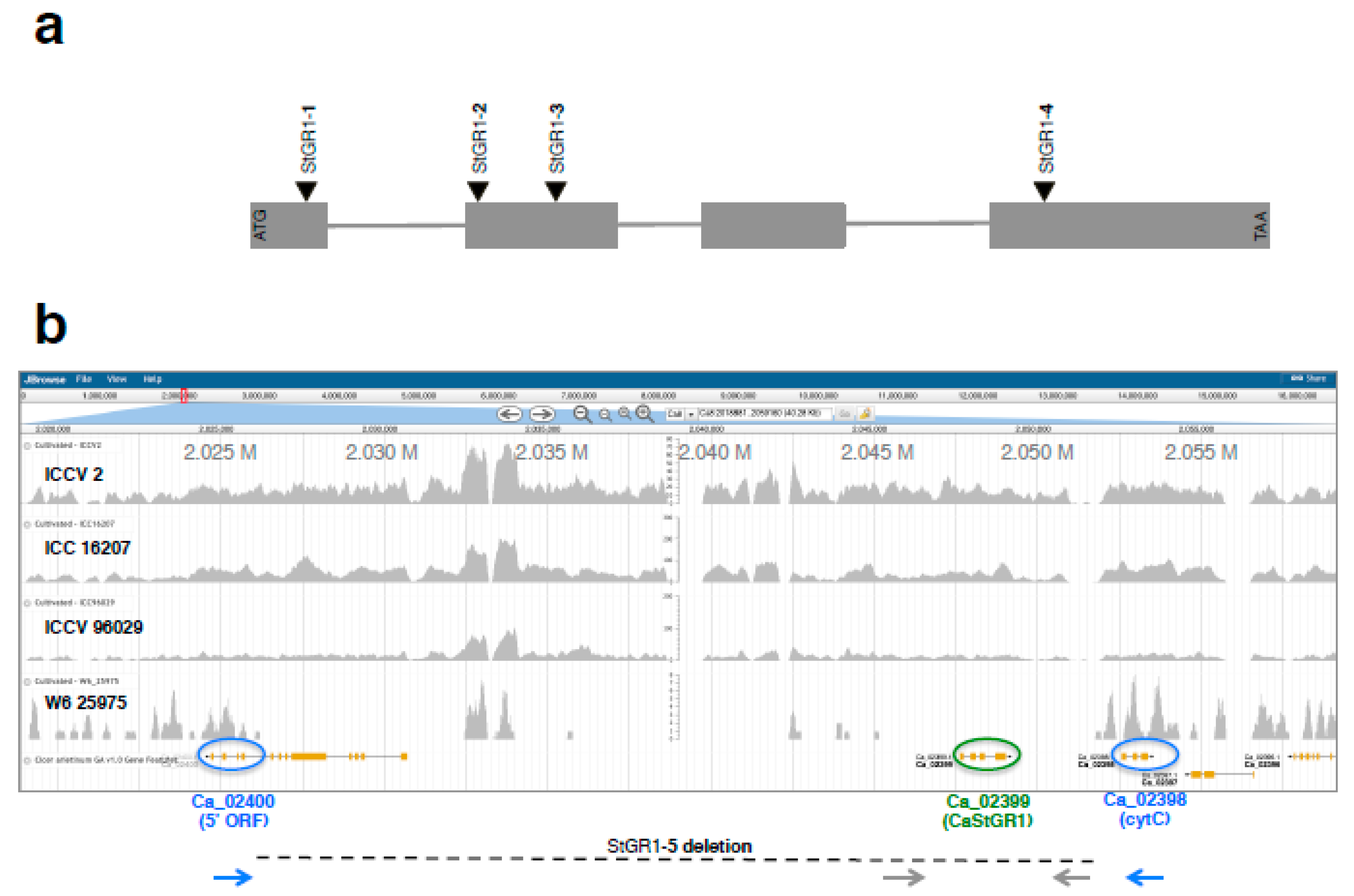
2.1.3. Association of CaStGR1 Sequence Variants with Green Cotyledon Chickpea Germplasm
2.1.4. Whole Genome Skim Sequencing Delimits the Extent of the Deletion in Allele CaStGR1-5
2.1.5. Genetic Cosegregation of Staygreen Sequence Variants with the Green-Cotyledon Trait
2.2. Characterization of Physiological Functions of Green Cotyledon Chickpea
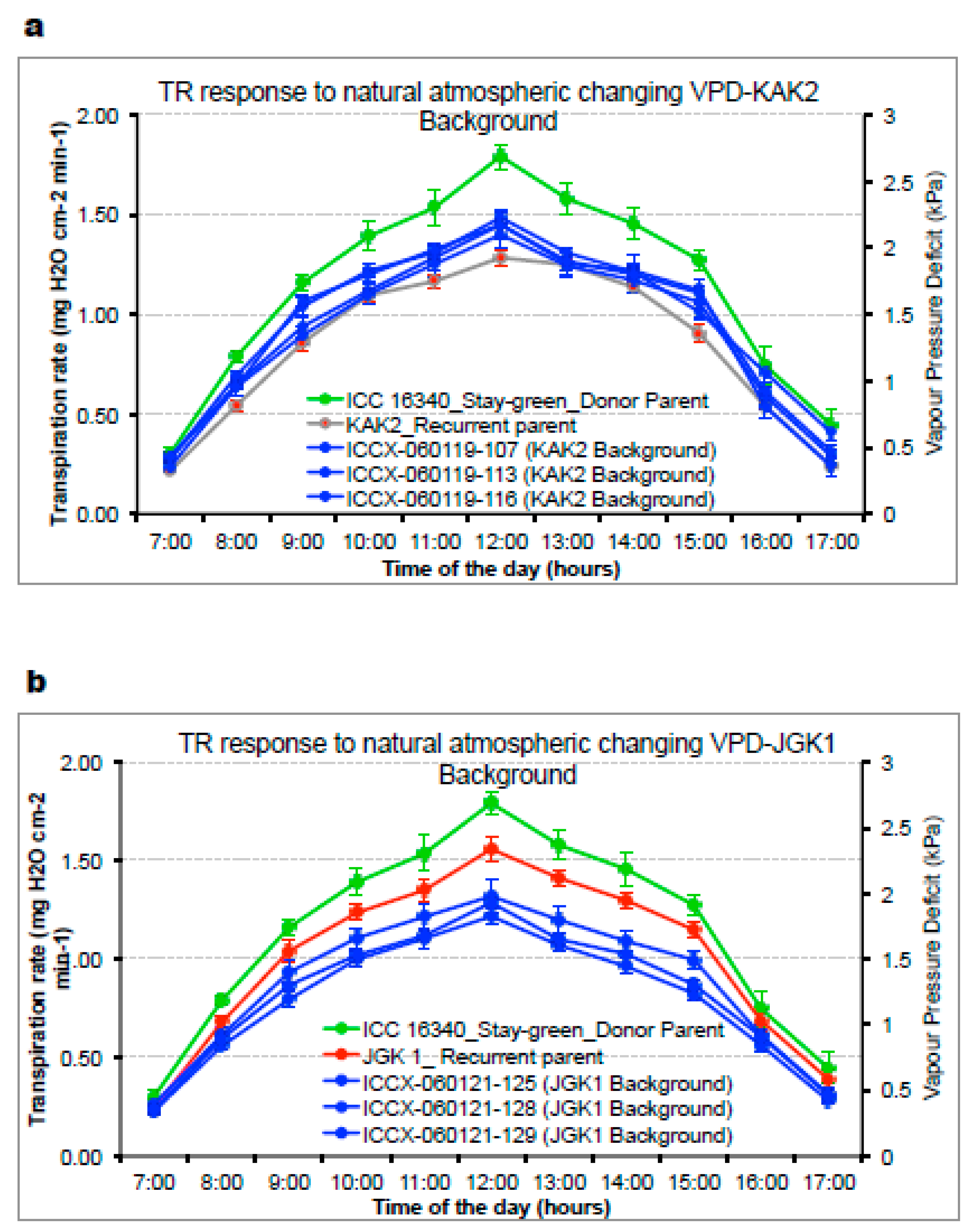
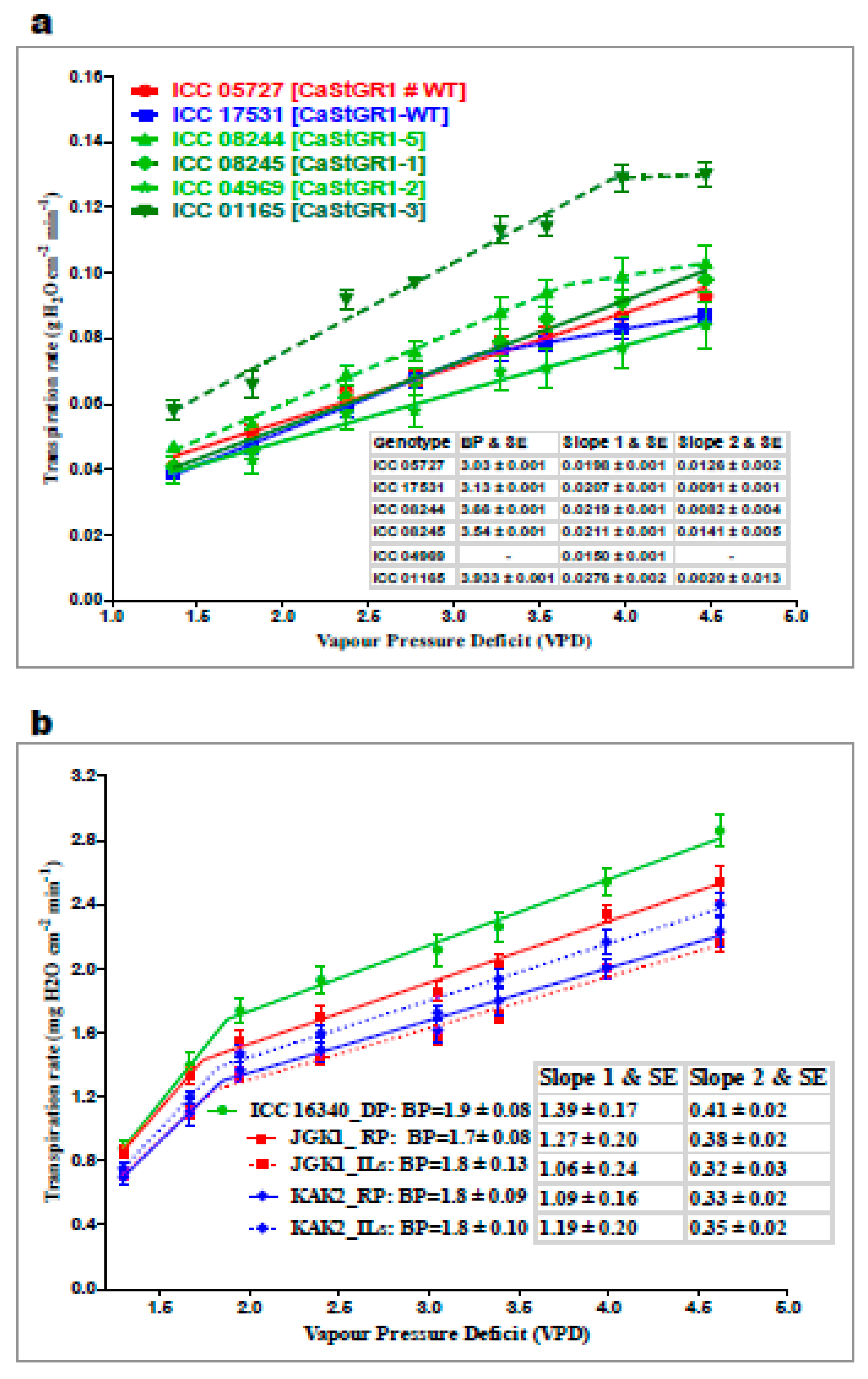
2.2.1. Plant Responsiveness to Soil and Atmospheric Drought (Experiment 1 and 2)
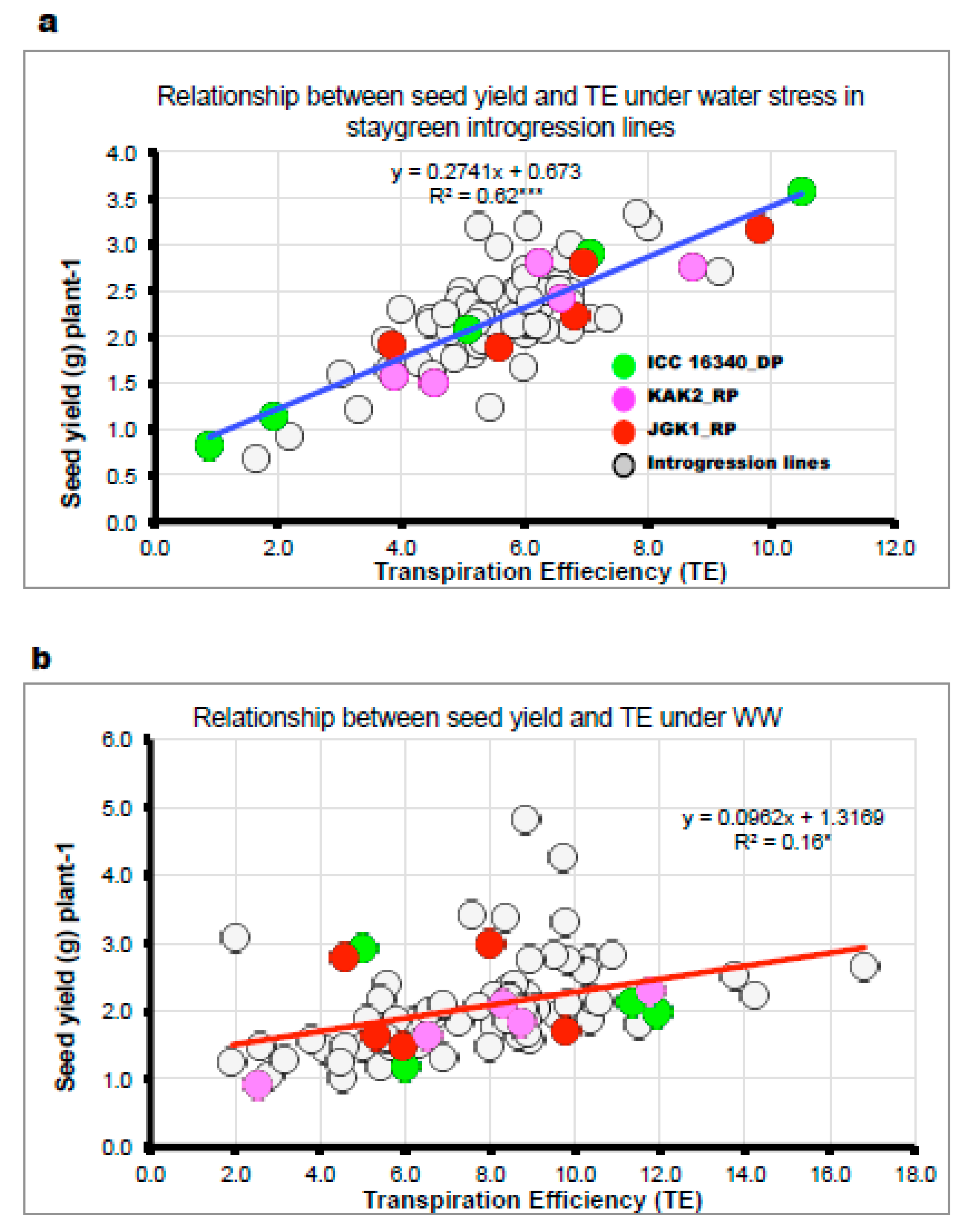
2.2.2. Variation in Plant Growth and Water-Use Related Traits in Lysimetric Facility (Experiment 3a, b)
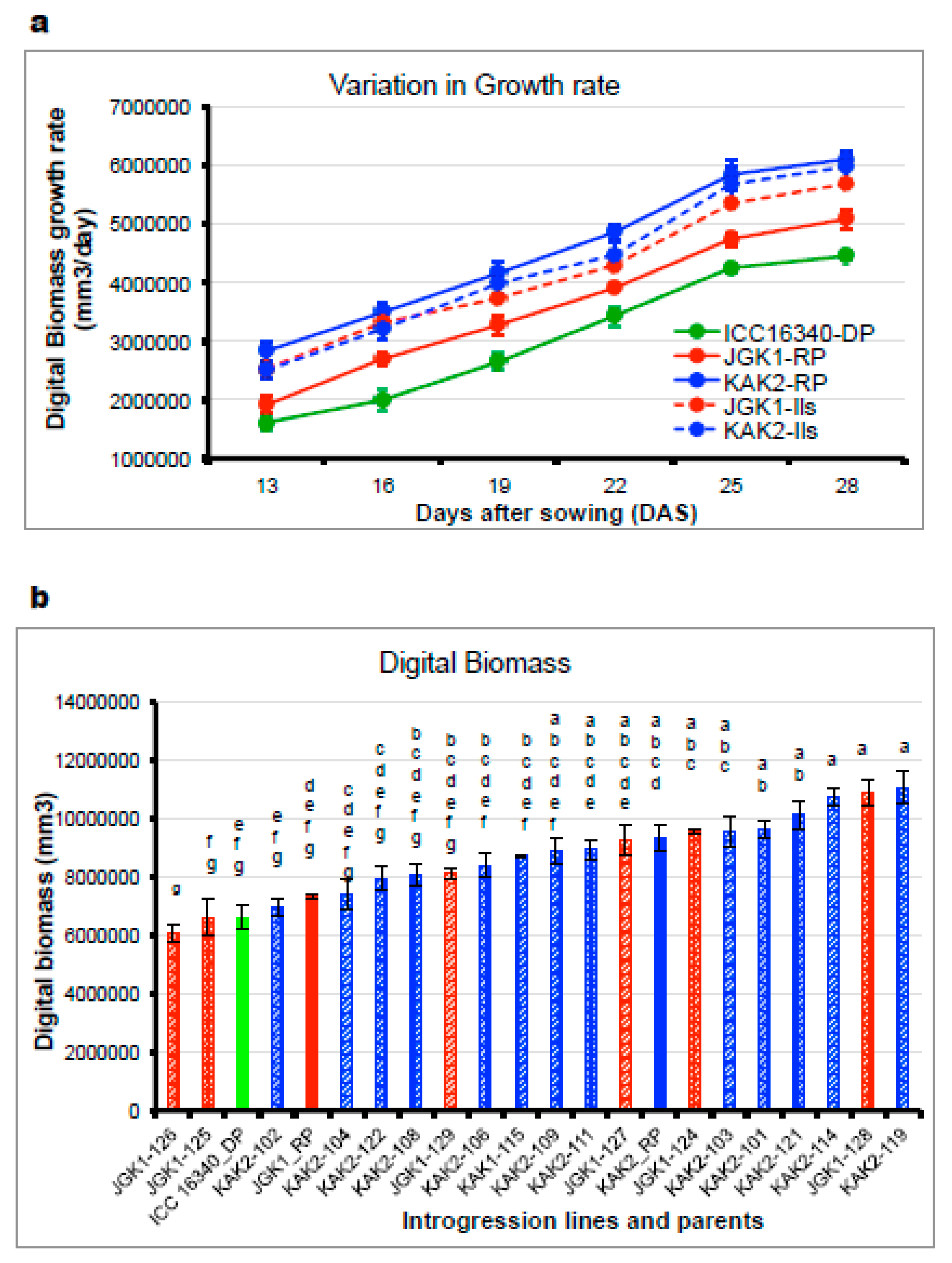
2.2.3. Evaluation of Canopy Growth Related Traits (Experiment 4)
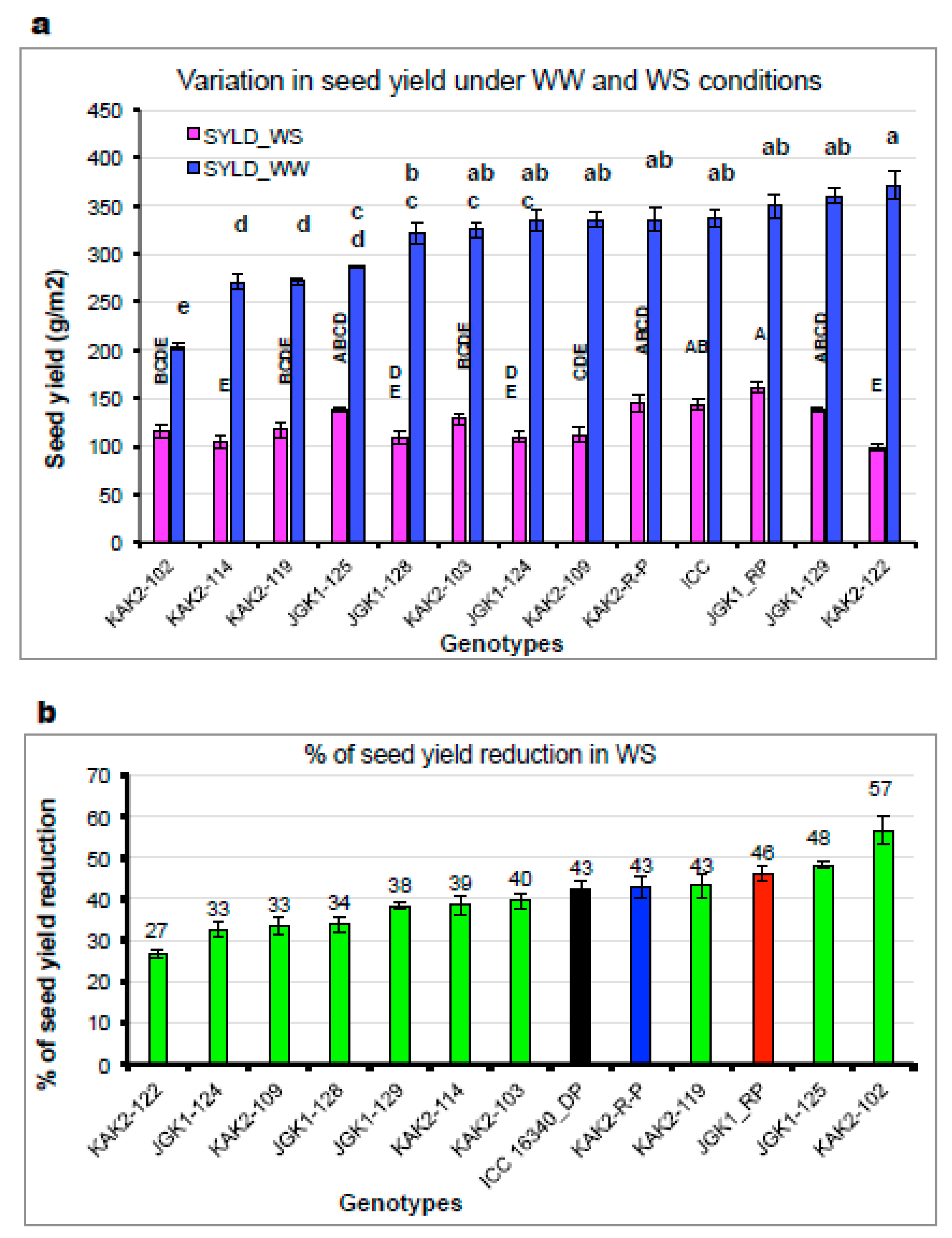
2.2.4. Evaluation in the Field Conditions (Experiment 5)
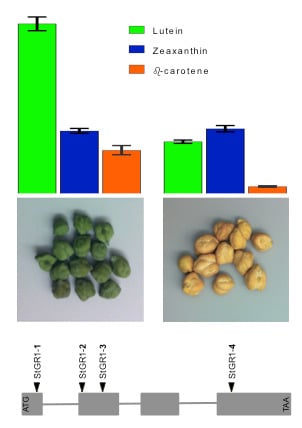
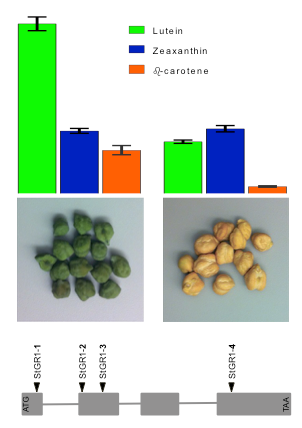
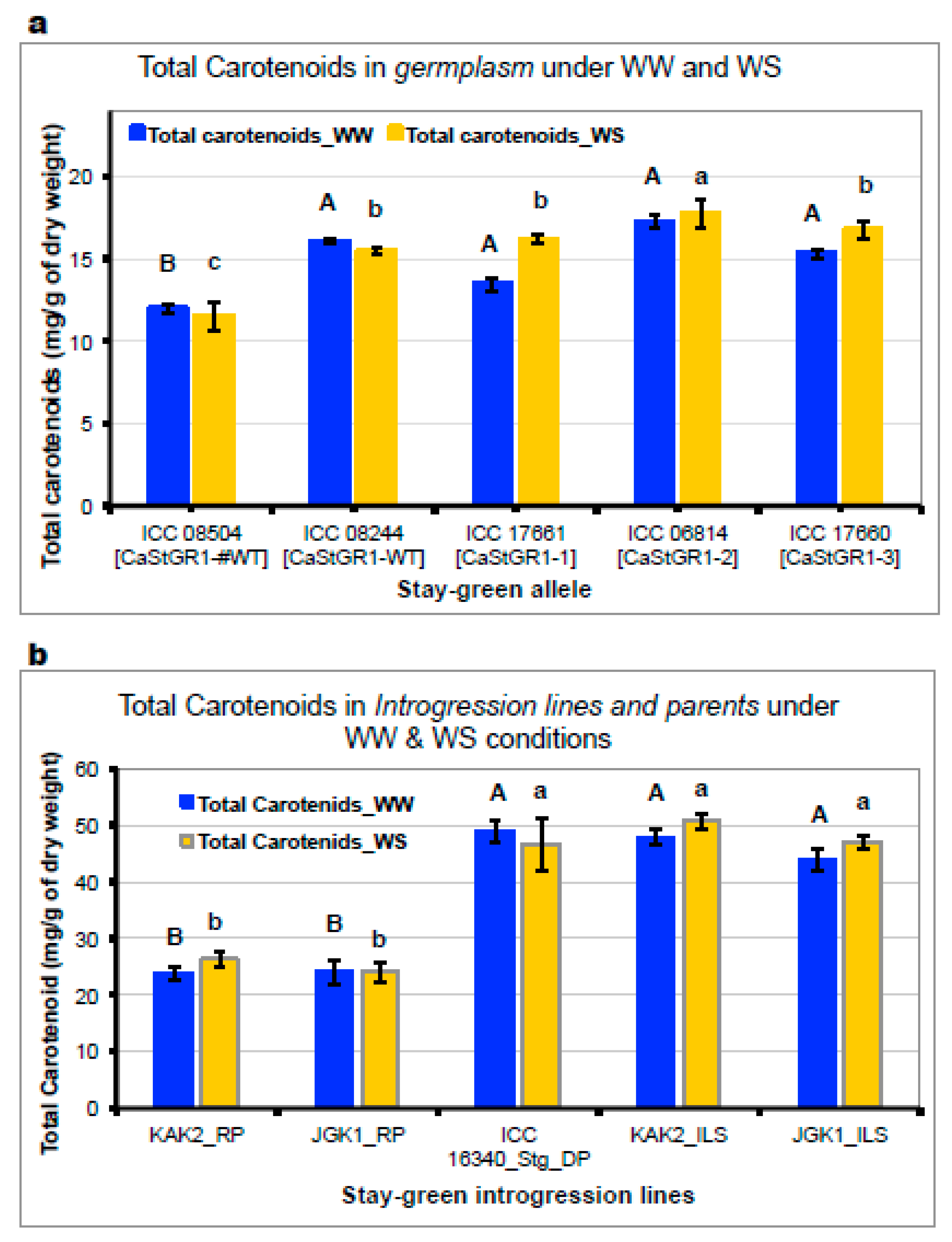
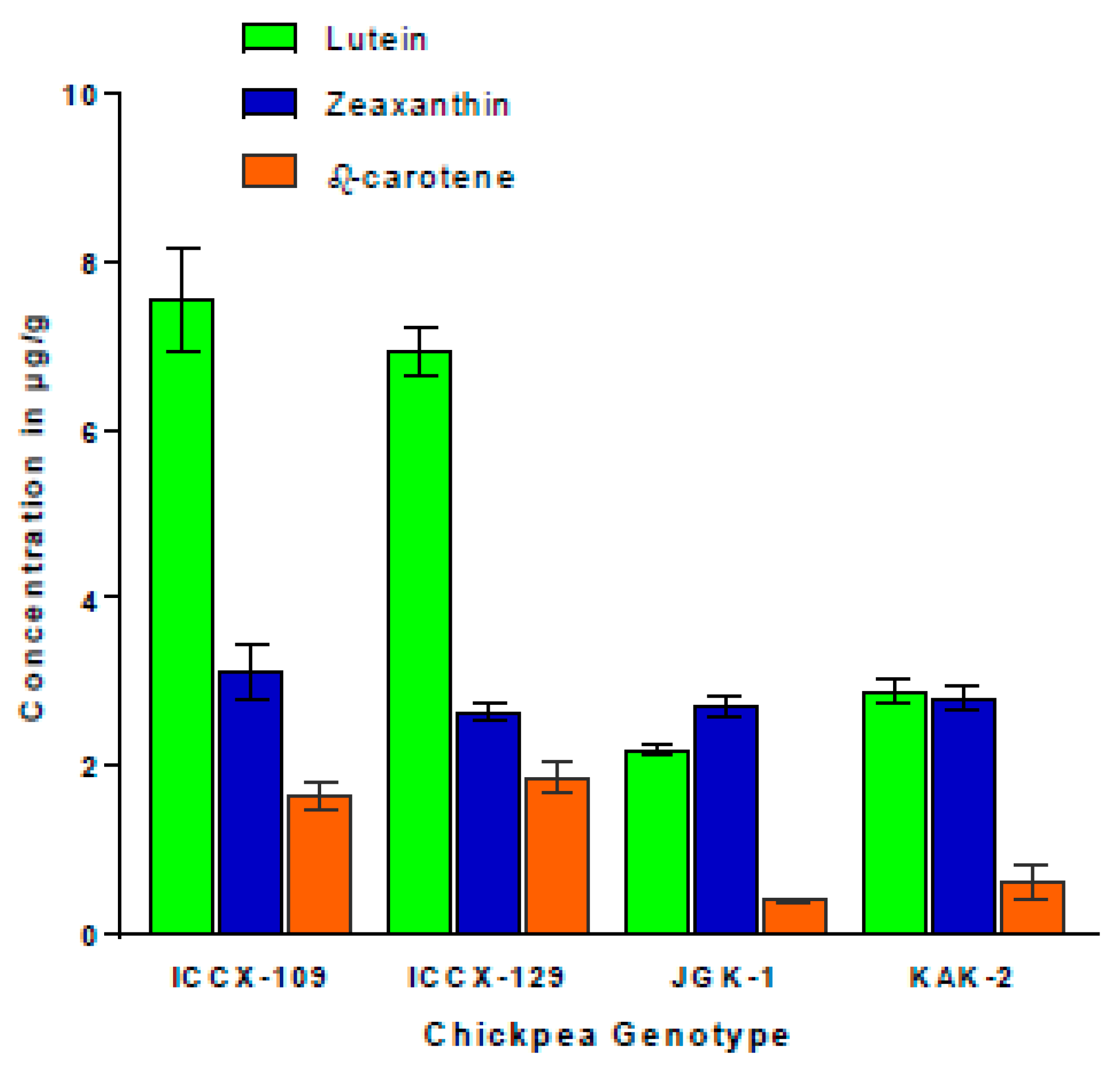
2.2.5. Leaf Pigments Content Under WW and WS Conditions (Experiments 1c, 3a,b,c)
3. Discussion
3.1. Identification of ‘Cosmetic Stay-Green’ Allele in Chickpea
3.2. Green Cotyledon Trait as a Vavilovian Homologous Series of Variation
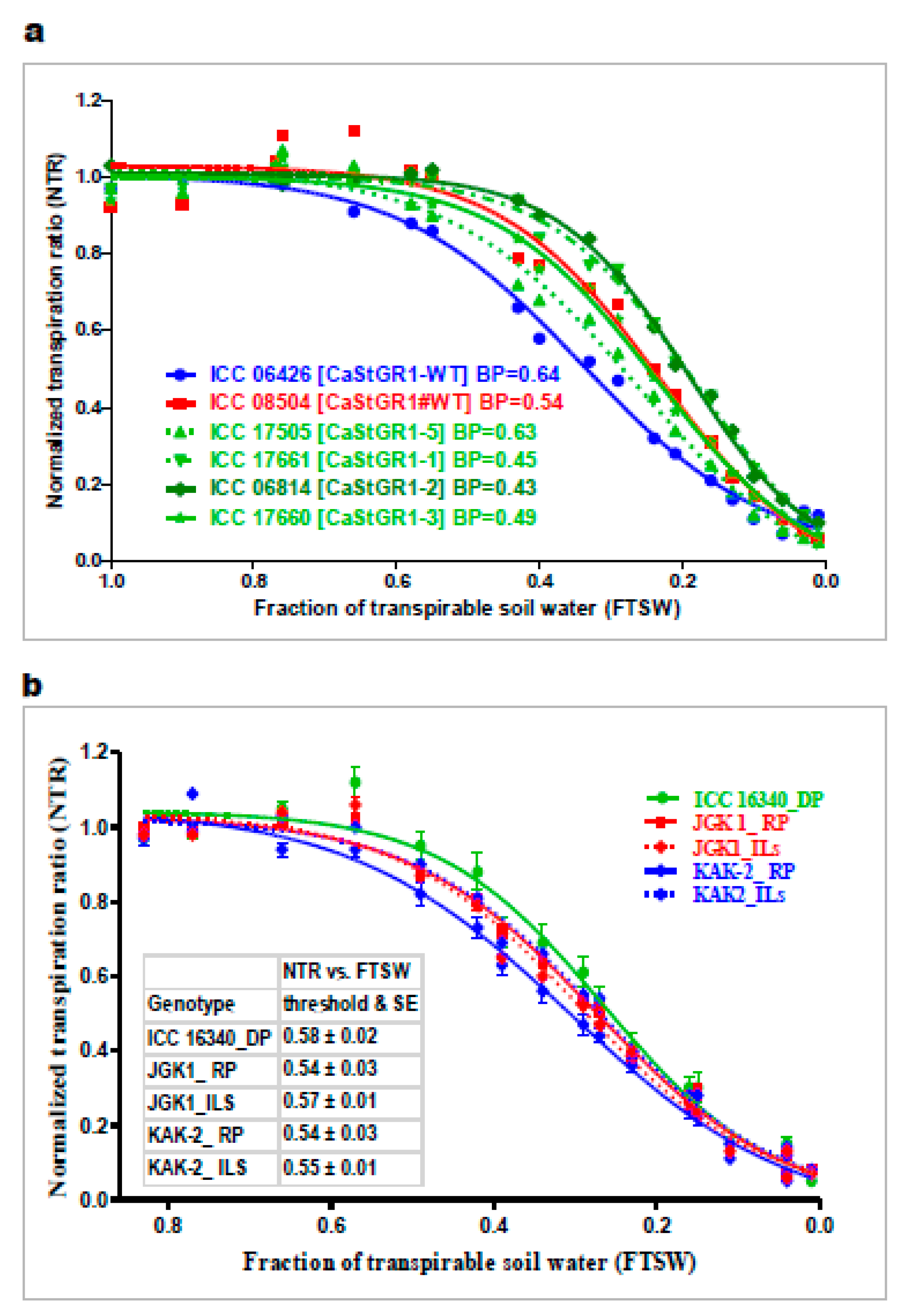
3.3. Stay-Green Alleles do not Affect the Plant Responsiveness to Soil and Atmospheric Drought
3.3.1. Plant Responsiveness to Soil and Atmospheric Drought
3.3.2. Plant Water-Use Related Traits and Agronomic Performance
3.3.3. Stay-Green Alleles Extend Retention of Chlorophyll and Provitaminogenic Carotenoids in Grains and Leaves
4. Materials and Methods
4.1. Plant Material: Chickpea Germplasm and Breeding Lines
4.2. Molecular Characterization of Candidate Gene and Genome:
4.3. Plant Growth Conditions for Physiological Assays (Experiments Listed in Table 3)
4.3.1. Experiments Conducted in Glass-House (Experiment 1, 2, 3a and b)
4.3.2. Experiments Conducted at LeasyField (Experiment 3c)
4.3.3. Experiments Conducted at LeasyScan (Experiment 4)
4.3.4. Experiments Conducted in Field (Experiment 5)
4.4. Physiological Assays
4.4.1. Experiments to Test Plant Responsiveness to Soil Drought (Experiment 1b and 1c)
4.4.2. Experiments to Test Plant Responsiveness to Atmospheric Drought (Experiment 2a,b,c)
4.4.3. Experiments to Test Plant Baseline Agronomic Features and Water-Use Related Traits in Lysimetric Facility (Experiment 3a,b,c)
4.4.4. Experiments to Assess Plant Canopy at LeasyScan (Experiment 4)
4.4.5. Agronomic Evaluation of ILs in Field Settings (Experiment 5)
HI = (Seed weight/total shoot biomass weight) × 100 [%].
4.5. Chlorophyll and Carotenoid Estimation in Leaves and Seeds (Measured in Experiment 1 and 3)
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. Chickpea Value Chain Food Loss Analysis: Causes and Solutions; FAO: Rome, Italy, 2017. [Google Scholar]
- Hajjarpoor, A.; Vadez, V.; Soltani, A.; Gaur, P.; Whitbread, A.; Suresh Babu, D.; Kholová, J. Characterization of the main chickpea cropping systems in India using a yield gap analysis approach. Field Crops Res. 2018, 223, 93–104. [Google Scholar] [CrossRef]
- FAOSTAT. Food and Agricultural Commodities Production; FAOSTAT: Rome, Italy, 2014. [Google Scholar]
- Tharanya, M.; Kholova, J.; Sivasakthi, K.; Thirunalasundari, T.; Vadez, V. Pearl Millet. In Water-Conservation Traits to Increase Crop Yields in Water-Deficit Environments: Case Studies; Springer: Cham, Switherland, 2017; pp. 73–83. [Google Scholar]
- Borrell, A.K.; Hammer, G.L. Nitrogen dynamics and the physiological basis of stay-green in Sorghum. Crop Sci. 2000, 40, 1295–1307. [Google Scholar] [CrossRef]
- Burgess, M.G.; Rush, C.M.; Piccinni, G.; Schuster, G. Relationship between charcoal rot, the stay-green trait, and irrigation in grain sorghum. Phytopathology 2002, 92, S10. [Google Scholar]
- Jordan, D.R.; Hunt, C.H.; Cruickshank, A.W.; Borrell, A.K.; Henzell, R.G. The relationship between the stay-green trait and grain yield in elite sorghum hybrids grown in a range of environments. Crop Sci. 2012, 52, 1153–1161. [Google Scholar] [CrossRef]
- Kholová, J.; Tharanya, M.; Sivasakthi, K.; Srikanth, M.; Rekha, B.; Hammer, G.L.; McLean, G.; Deshpande, S.; Hash, C.T.; Craufurd, P.; et al. Modelling the effect of plant water use traits on yield and stay-green expression in sorghum. Funct. Plant Biol. 2014, 1, 1019–1034. [Google Scholar] [CrossRef]
- McBee, G.G.; Waskom, R.M.; Miller, F.R.; Creelman, R.A. Effect of senescence and nonsenescence on carbohydrates in sorghum during late kernel maturity states. Crop Sci. 1983, 23, 372–376. [Google Scholar] [CrossRef]
- Vadez, V.; Kholová, J.; Yadav, R.S.; Hash, C.T. Small temporal differences in water uptake among varieties of pearl millet (Pennisetum glaucum (L.) R Br) are critical for grain yield under terminal drought. Plant Soil 2013, 371, 447–462. [Google Scholar] [CrossRef]
- Crafts-Brandner, S.J.; Below, F.E.; Wittenbach, V.A.; Harper, J.E.; Hageman, R.H. Differential Senescence of Maize Hybrids following Ear Removal. Plant Physiol. 1984, 74, 368–373. [Google Scholar] [CrossRef]
- Gentinetta, E.; Ceppi, D.; Lepori, C.; Perico, G.; Motto, M.; Salamini, F. A major gene for delayed senescence in maize. Pattern of photosynthates accumulation and inheritance. Plant Breed. 1986, 97, 193–203. [Google Scholar] [CrossRef]
- Rajcan, I.; Tollenaar, M. Source: Sink ratio and leaf senescence in maize. I. Dry matter accumulation and partitioning during grain filling. Field Crops Res. 1999, 60, 245–253. [Google Scholar] [CrossRef]
- Zheng, H.J.; Wu, A.Z.; Zheng, C.; Dong, S.T. QTL mapping of maize (Zea mays) stay-green traits and their relationship to yield. Plant Breed. 2009, 128, 54–62. [Google Scholar] [CrossRef]
- Adu, M.O.; Sparkes, D.L.; Parmar, A.; Yawson, D.O.; Science, B. ‘Stay Green’ in Wheat: Comparative Study of Modern Bread Wheat and Ancient Wheat Cultivars. J. Agric. Biol. Sci. 2011, 6, 16–24. [Google Scholar]
- Bogard, M.; Jourdan, M.; Allard, V.; Martre, P.; Perretant, M.R.; Ravel, C.; Heumez, E.; Orford, S.; Snape, J.; Gaju, O.; et al. Anthesis date mainly explained correlations between post-anthesis leaf senescence, grain yield, and grain protein concentration in a winter wheat population segregating for flowering time QTLs. J. Exp. Bot. 2011, 62, 3621–3636. [Google Scholar] [CrossRef] [PubMed]
- De Souza Luche, H.; da Silva, J.A.G., II; da Maia, L.C., III; de Oliveira, A.C., III. Stay-green: A potentiality in plant breeding. Stay-green: A potentiality in plant breeding Stay-green: Uma potencialidade no melhoramento genético de plantas. J. Crop Prod. 2015, 45, 1755–1760. [Google Scholar]
- Christopher, J.T.; Manschadi, A.M.; Hammer, G.L.; Borrell, A.K. Developmental and physiological traits associated with high yield and stay-green phenotype in wheat. Aust. J. Agric. Res. 2008, 59, 354–364. [Google Scholar] [CrossRef]
- Lopes, M.S.; Reynolds, M.P. Stay-green in spring wheat can be determined by spectral reflectance measurements (normalized difference vegetation index) independently from phenology. J. Exp. Bot. 2012, 63, 3789–3798. [Google Scholar] [CrossRef]
- Fu, J.-D.; Yan, Y.-F.; Lee, B.-W. Physiological characteristics of a functional stay-green rice “SNU-SG1” during grain-filling period. J. Crop Sci. Biotechnol. 2009, 12, 47–52. [Google Scholar] [CrossRef]
- Hoang, T.B.; Kobata, T. Stay-green in rice (Oryza sativa L.) of drought-prone areas in desiccated soils. Plant Prod. Sci. 2009, 12, 397–408. [Google Scholar] [CrossRef]
- Mondal, W.A.; Dey, B.B.; Choudhuri, M.A. Proline accumulation as a reliable indicator of monocarpic senescence in rice cultivars. Experientia 1985, 41, 346–348. [Google Scholar] [CrossRef]
- Wada, Y.; Wada, G. Varietal difference in leaf senescence during ripening period of advanced indica rice. Jap. J. Crop Sci. 1991, 60, 529–553. [Google Scholar] [CrossRef]
- Borrell, A.K.; Mullet, J.E.; George-Jaeggli, B.; van Oosterom, E.J.; Hammer, G.L.; Klein, P.E.; Jordan, D.R. Drought adaptation of stay-green sorghum is associated with canopy development, leaf anatomy, root growth, and water uptake. J. Exp. Bot. 2014, 65, 6251–6263. [Google Scholar] [CrossRef]
- Borrell, A.K.; van Oosterom, E.J.; Mullet, J.E.; George-Jaeggli, B.; Jordan, D.R.; Klein, P.E.; Hammer, G.L. Stay-green alleles individually enhance grain yield in sorghum under drought by modifying canopy development and water uptake patterns. New Phytol. 2014, 203, 817–830. [Google Scholar] [CrossRef] [PubMed]
- Kholová, J.; McLean, G.; Vadez, V.; Craufurd, P.; Hammer, G.L. Drought stress characterization of post-rainy season (rabi) sorghum in India. Field Crops Res. 2013, 141, 38–46. [Google Scholar] [CrossRef]
- Van Oosterom, E.J.; Borrell, A.K.; Chapman, S.C.; Broad, I.J.; Hammer, G.L. Functional dynamics of the nitrogen balance of sorghum. I. N demand of vegetative plant parts. Field Crop Res. 2010, 115, 19–28. [Google Scholar] [CrossRef]
- Van Oosterom, E.J.; Chapman, S.C.; Borrell, A.K.; Broad, I.J.; Hammer, G.L. Functional dynamics of the nitrogen balance of sorghum. II. Grain filling period. Field Crops Res. 2010, 115, 29–38. [Google Scholar] [CrossRef]
- Thomas, H.; Ougham, H. The stay-green trait. J. Exp. Bot. 2014, 65, 3889–3900. [Google Scholar] [CrossRef]
- Ashokkumar, K.; Tar’an, B.; Diapari, M.; Arganosa, G.; Warkentin, T.D. Effect of Cultivar and Environment on Carotenoid Profile of Pea and Chickpea. Crop Sci. 2014, 54, 2225–2235. [Google Scholar] [CrossRef]
- Hortensteiner, S. Stay-green regulates chlorophyll and chlorophyll-binding protein degradation during senescence. Trends Plant Sci. 2009, 14, 155–162. [Google Scholar] [CrossRef]
- Segev, A.; Badani, H.; Kapulnik, Y.; Shomer, I.; Oren-Shamir, M.; Galili, S. Determination of Polyphenols, Flavonoids, and Antioxidant Capacity in Colored Chickpea (Cicer arietinum L.). J. Food Sci. 2010, 75, 115–119. [Google Scholar] [CrossRef]
- Zhou, C.; Han, L.; Pislariu, C.; Nakashima, J.; Fu, C.; Jiang, Q.; Quan, L.; Blancaflor, E.B.; Tang, Y.; Bouton, J.H.; et al. From model to crop: Functional analysis of a STAY-GREEN gene in the model legume Medicago truncatula and effective use of the gene for alfalfa improvement. Plant Physiol. 2011, 157, 483–1496. [Google Scholar] [CrossRef]
- Luo, Z.; Zhang, J.; Li, J.; Yang, C.; Wang, T.; Ouyang, B.; Li, H.; Giovannoni, J.; Ye, Z. A STAY-GREEN protein SlSGR1 regulates lycopene and β-carotene accumulation by interacting directly with SlPSY1 during ripening processes in tomato. New Phytol. 2013, 198, 442–452. [Google Scholar] [CrossRef] [PubMed]
- Meier, S.; Tzfadia, O.; Vallabhaneni, R.; Gehring, C.; Wurtzel, E.T. A transcriptional analysis of carotenoid, chlorophyll and plastidial isoprenoid biosynthesis genes during development and osmotic stress responses in Arabidopsis thaliana. BMC Syst. Biol. 2011, 5, 77. [Google Scholar] [CrossRef] [PubMed]
- Mur, L.A.J.; Aubry, S.; Mondhe, M.; Kingston-Smith, A.; Gallagher, J.; Timms-Taravella, E.; James, C.; Papp, I.; Hörtensteiner, S.; Thomas, H.; et al. Accumulation of chlorophyll catabolites photosensitizes the hypersensitive response elicited by Pseudomonas syringae in Arabidopsis. New Phytol. 2010, 188, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Ross, C. Vitamin A. In Encyclopedia of Dietary Supplements, 2nd ed.; Informa Healthcare: London, UK; New York, NY, USA, 2010; pp. 778–791. [Google Scholar]
- Ross, A. Vitamin A and Carotenoids. In Modern Nutrition in Health and Disease, 10th ed.; Lippincott Williams & Wilkins: Baltimore, MD, USA, 2006; pp. 351–375. [Google Scholar]
- Rezaei, M.K.; Deokar, A.; Arganosa, G.; Roorkiwal, M.; Pandey, S.K.; Warrkentin, T.D.; Varshney, R.K.; Tar’an, B. Mapping Quantitative Trait Loci for Carotenoid Concentration in Three F2 Populations of Chickpea. Plant Genome 2019, 12, 1–12. [Google Scholar]
- Date, S.G. The Study of the Effect of Various Methods of Harvesting and Curing on the Color and Viability of Lima Bean Seed. Master’s Thesis, Utah State University, Logan, UT, USA, 1962. [Google Scholar]
- Davis, J.; Myers, J.R.; McClean, P.; Lee, R. Staygreen (sgr), a candidate gene for the Persistent Color phenotype in common bean. Acta Hortic. 2010, 859, 99–102. [Google Scholar] [CrossRef]
- Fery, R.L.; Dukes, P.D. Genetic analysis of the green cotyledon trait in southern pea (Vigna unguiculata (L.) Walp.). J. Am. Soc. Hortic. Sci. 1994, 119, 1054–1056. [Google Scholar] [CrossRef]
- Mae, T.; Ohira, K. Origin of the nitrogen a growing rice leaf and its relation to nitrogen nutrition. Jpn. J. Soil Sci. Plant Nutr. 1983, 54, 401–405. [Google Scholar]
- Morita, K. Release of nitrogen from chloroplasts during senescence in rice (Oryza sativa L.). Ann. Bot. 1980, 46, 297–302. [Google Scholar] [CrossRef]
- Sage, R.F.; Pearcy, R.W. The Nitrogen Use Efficiency of C3 and C4 Plants: I. Leaf Nitrogen, Growth, and Biomass Partitioning in Chenopodium album (L.) and Amaranthus retroflexus (L.). Plant Physiol. 1987, 84, 954–958. [Google Scholar] [CrossRef]
- Tucker, C.L. Inheritance of white and green seed coat colors in lima beans. Proc. Amer. Soc. Hortic. Sci. 1965, 87, 286–287. [Google Scholar]
- Tucker, C.L.; Sanches, R.L.; Harding, J. Effects of a gene for cotyledon color in lima beans, Phaseolus lunatus. Crop Sci. 1967, 87, 262–263. [Google Scholar] [CrossRef]
- Armstead, I.; Donnison, I.; Aubry, S.; Harper, J.; Hortensteiner, S.; James, C.; Mani, J.; Moffet, M.; Ougham, H.; Roberts, L.; et al. Cross-species identification of Mendel’s I locus. Science 2007, 315, 73. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Morita, R.; Nishimura, M.; Yamaguchi, H.; Kusaba, M. Mendel’s green cotyledon gene encodes a positive regulator of the chlorophyll-degradation pathway. Proc. Natl. Acad. Sci. USA 2007, 104, 14169–14174. [Google Scholar] [CrossRef] [PubMed]
- Varshney, R.K.; Song, C.; Saxena, R.K.; Azam, S.; Yu, S.; Sharpe, A.G.; Cannon, S.; Baek, J.; Rosen, B.D.; Tar’an, B.; et al. Draft genome sequence of chickpea (Cicer arietinum) provides a resource for trait improvement. Nat. Biotechnol. 2013, 31, 240–246. [Google Scholar]
- Jiang, H.; Li, M.; Liang, N.; Yan, H.; Wei, Y.; Xu, X.; Liu, J.; Xu, Z.; Chen, F.; Wu, G. Molecular cloning and function analysis of the stay green gene in rice. Plant J. 2007, 52, 197–209. [Google Scholar] [CrossRef]
- Myers, J.R.; Aljadi, M.; Brewer, L. The Importance of Cosmetic Stay Green in Specialty Crops. Plant Breed. Rev. 2018, 219. [Google Scholar] [CrossRef]
- Sakuraba, Y.; Schelbert, S.; Park, S.Y.; Han, S.H.; Lee, B.D.; Andrès, C.B.; Paek, N.C. STAY-GREEN and chlorophyll catabolic enzymes interact at light-harvesting complex II for chlorophyll detoxification during leaf senescence in Arabidopsis. Plant Cell 2012, 24, 507–518. [Google Scholar] [CrossRef]
- Muchero, W.; Roberts, P.A.; Diop, N.N.; Drabo, I.; Cisse, N.; Close, T.J.; Muranaka, S.; Boukar, O.; Ehlers, J.D. Genetic architecture of delayed senescence, biomass, and grain yield under drought stress in cowpea. PLoS ONE 2013, 8, e70041. [Google Scholar] [CrossRef]
- Singh, U.; Pundir, R.P.S. Amino acid composition and protein content of chickpea and its wild relatives. Int. Chickpea Newsl. 1991, 25, 19–20. [Google Scholar]
- Smartt, J. Vavilov’s law of homologous serious and de nova crop plant domestication. Biol. J Linnean Society. 1990, 39, 27–38. [Google Scholar] [CrossRef]
- Kholova, J.; Zindy, P.; Malayee, S.; Baddam, R.; Murugesan, T.; Kaliamoorthy, S.; Hash, C.T.; Votrubová, O.; Soukup, A.; Kocová, M.; et al. Component traits of plant water use are modulated by vapour pressure deficit in pearl millet (Pennisetum glaucum(L.) R.Br.). Funct. Plant Biol. 2016, 43, 423–437. [Google Scholar] [CrossRef]
- Vadez, V.; Halilou, O.; Hissene, H.M.; Sibiry-Traore, P.; Sinclair, T.R.; Soltani, A. Mapping Water Stress Incidence and Intensity, Optimal Plant Populations, and Cultivar Duration for African Groundnut Productivity Enhancement. Front. Plant Sci. 2017, 8, 432. [Google Scholar] [CrossRef] [PubMed]
- Vadez, V.; Kholová, J.; Medina, S.; Kakkera, A.; Anderberg, H. Transpiration efficiency: New insight on an old story. J. Exp. Bot. 2014, 65, 6141–6153. [Google Scholar] [CrossRef] [PubMed]
- Vadez, V.; Ratnakumar, P. High transpiration efficiency increases pod yield under intermittent drought in dry and hot atmospheric conditions but less so under wetter and cooler conditions in groundnut (Arachis hypogaea (L.)). Field Crops Res. 2016, 193, 16–23. [Google Scholar] [CrossRef]
- Sivasakthi, K.; Zaman-Allah, M.; Tharanya, M.; Kholova, J.; Thirunalasundari, T.; Vadez, V. Chickpea. In Water-Conservation Traits to Increase Crop Yields in Water-Deficit Environments: Case Studies; Springer: Cham, Switherland, 2017; pp. 35–45. [Google Scholar]
- Tharanya, M.; Kholová, J.; Sivasakthi, K.; Seghal, D.; Hash, C.T.; Raj, B.; Srivastava, R.K.; Baddam, R.; Thirunalasundari, T.; Yadav, R.; et al. Quantitative trait loci (QTLs) for water use and crop production traits co-locate with major QTL for tolerance to water deficit in a fine-mapping population of pearl millet (Pennisetum glaucum L. R.Br.). Theor. Appl. Genet. 2018, 131, 1509. [Google Scholar] [CrossRef]
- Tharanya, M.; Sivasakthi, K.; Gloria, B.; Kholova, J.; Thirunalasundari, T.; Vadez, V. Pearl millet [Pennisetum glaucum (L.) R. Br.] contrasting for the transpiration response to vapour pressure deficit also differ in their dependence on the symplastic and apoplastic water transport pathways. Funct. Plant Biol. 2018, 45. [Google Scholar] [CrossRef]
- Vadez, V.; Kholová, J.; Hummel, G.; Zhokhavets, U.; Gupta, S.K.; Hash, C.T. LeasyScan: A novel concept combining 3D imaging and lysimetry for high-throughput phenotyping of traits controlling plant water budget. J. Exp. Bot. 2015, 66, 5581–5593. [Google Scholar] [CrossRef]
- Von Wettberg, E.J.; Chang, P.L.; Greenspan, A.; Carrasquila-Garcia, N.; Basdemir, F.; Moenga, S.; Bedada, G.; Dacosta-Calheiros, E.; Moriuchi, K.S.; Balcha, L.; et al. Ecology and community genomics of an important crop wild relative as a prelude to agricultural innovation. Nat. Commun. 2018, 9, 649. [Google Scholar] [CrossRef]
- Vadez, V.; Deshpande, S.; Kholova, J.; Ramu, P.; Hash, C.T. Molecular breeding for stay-green: Progress and challenges in sorghum. In Genomic Applications to Crop Breeding: Vol. 2. Improvement for Abiotic Stress, Quality and Yield Improvement; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013; pp. 125–141. [Google Scholar]
- Vadez, V.; Kholová, J.; Zaman-Allah, M.; Belko, N. Water: The most important “molecular” component of water stress tolerance research. Funct. Plant Biol. 2013, 40, 1310–1322. [Google Scholar] [CrossRef]
- Vadez, V.; Rao, J.S.; Bhatnagar-Mathur, P.; Sharma, K.K. DREB1A promotes root development in deep soil layers and increases water extraction under water stress in groundnut. Plant Biol. 2013, 15, 45–52. [Google Scholar] [CrossRef]
- Zaman-Allah, M.; Jenkinson, D.M.; Vadez, V. A conservative pattern of water use, rather than deep or profuse rooting, is critical for the terminal drought tolerance of chickpea. J. Exp. Bot. 2011, 62, 4239–4252. [Google Scholar] [CrossRef] [PubMed]
- Zaman-Allah, M.; Jenkinson, D.M.; Vadez, V. Chickpea genotypes contrasting for seed yield under terminal drought stress in the field differ for traits related to the control of water use. Funct. Plant Biol. 2011, 38, 270–281. [Google Scholar] [CrossRef]
- Balazadeh, S. Stay-Green not always stay-green. Mol. Plant. 2014, 7, 1264–1266. [Google Scholar] [CrossRef] [PubMed]
- Thomas, H.; Howarth, C.M. Five ways to stay green. J. Exp. Bot. 2000, 51, 329–337. [Google Scholar] [CrossRef]
- Borrell, A.K. Stay-green alleles individually enhance grain yield in sorghum. New Phytol. 2014, 203, 817–830. [Google Scholar] [CrossRef]
- Kamal, N.M.; Serag, Y.; Gorafi, A.; Tsujimoto, H.; Ghanim, A.M.A. Stay-Green QTLs Response in Adaptation to Post-Flowering Drought Depends on the Drought Severity. BioMed Res. Int. 2018, 2018, 1–15. [Google Scholar] [CrossRef]
- Abbo, S.; Bonfil, D.J.; Berkovitch, Z.; Reifen, R. Towards enhancing lutein concentration in chickpea, cultivar and management effects. Plant Breed. 2010, 129, 407–411. [Google Scholar] [CrossRef]
- Rezaei, M.K.; Deokar, A.; Tar’an, B. Identification and Expression Analysis of Candidate Genes Involved in Carotenoid Biosynthesis in Chickpea Seeds. Front. Plant Sci. 2016, 7, 1867. [Google Scholar] [CrossRef]
- Vadez, V.; Rao, J.S.; Kholova, J.; Krishnamurthy, L.; Kashiwagi, J.; Ratnakumar, P.; Sharma, K.K.; Bhatnagar-Mathur, P.; Basu, P.S. Roots research for legume tolerance to drought: Quo vadis? J. Food Legumes 2008, 21, 77–85. [Google Scholar]
- Sharma, S.B.; Sahrawat, K.L.; Burford, J.R.; Rupela, O.P.; Kumar Rao, J.V.D.K.; Sithanantham, S. Effects of Soil Solarization on Pigeonpea and Chickpea. Research Bulletin No. 11. Int. Crops Res. Inst. Semi-Arid Trop. 1988, 11, 1–23. [Google Scholar]
- Belko, N.; Zaman-allah, M.; Diop, N.N.; Cisse, N.; Zombre, G.; Ehlers, J.D.; Vadez, V. Restriction of transpiration rate under high vapour pressure deficit and non-limiting water conditions is important for terminal drought tolerance in cowpea. Plant Biol. 2012, 15, 304–316. [Google Scholar] [CrossRef] [PubMed]
- Kholová, J.; Hash, C.T.; Kakkera, A.; Kočová, M.; Vadez, V. Constitutive water conserving mechanisms are correlated with the terminal drought tolerance of pearl millet [Pennisetum glaucum (L.) R. Br.]. J. Exp. Bot. 2010, 61, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Vadez, V.; Sinclair, T.R. Leaf ureide degradation and N2 fixation tolerance to water deficit in soybean. J. Exp. Bot. 2001, 52, 153–159. [Google Scholar] [PubMed]
- Vadez, V.; Deshpande, S.P.; Kholová, J.; Hammer, G.L.; Borrell, A.K.; Talwar, H.S.; Hash, C.T. Stay-green quantitative trait loci’s effects on water extraction, transpiration efficiency and seed yield depend on recipient parent background. Funct. Plant Biol. 2011, 38, 553–566. [Google Scholar] [CrossRef]
- Vadez, V.; Krishnamurthy, L.; Hash, C.T.; Upadhyaya, H.D.; Borrell, A.K. Yield, transpiration efficiency, and water-use variations and their interrelationships in the sorghum reference collection. Crop Pasture Sci. 2011, 62, 645–655. [Google Scholar] [CrossRef]
- Wellburn, A.R. The Spectral Determination of Chlorophylls a and b, as well as Total Carotenoids, Using Various Solvents with Spectrophotometers of Different Resolution. J. Plant Physiol. 1994, 144, 3307–3313. [Google Scholar] [CrossRef]
- Kurilich, A.C.; Juvik, J.A. Quantification of carotenoid and tocopherol antioxidants in Zea mays. J. Agric. Food Chem. 1999, 47, 1948–1955. [Google Scholar] [CrossRef]
- Kholová, J.; Hash, C.T.; Kumar, P.L.; Yadav, S.R.; Kočová, M.; Vadez, V. Terminal drought-tolerant pearl millet [Pennisetum glaucum (L.) R. Br.] have high leaf ABA and limit transpiration at high vapor pressure deficit. J. Exp. Bot. 2010, 61, 1431–1440. [Google Scholar] [CrossRef]
- Sivasakthi, K.; Thudi, M.; Tharanya, M.; Kale, S.M.; Kholová, J.; Halime, M.H.; Jaganathan, D.; Baddam, R.; Thirunalasundari, T.; Gaur, P.M.; et al. Plant vigour QTLs co-map with an earlier reported QTL hotspot for drought tolerance while water saving QTLs map in other regions of the chickpea genome. BMC Plant Biol. 2018, 18, 29. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cotyledon Color | Allele | Nucleotide Variation in CaStGR1 | Number of Genotypes |
|---|---|---|---|
| green | CaStGR1-1 | 1-bp “g“ del in exon 1 | 7 |
| green | CaStGR1-2 | 8-bp “ctaggttg“ deletion in exon 2 | 5 |
| green | CaStGR1-3 | 1-bp “c“ deletion in exon 2 | 10 |
| green | CaStGR1-4 | 1-bp “g“ del in exon 4 | 6 |
| green | CaStGR1-5 | entire gene deleted | 11 |
| Yellow/Tan | CaStGR1 WT | “Wild Type” | 6 |
| Genotypes | NTR-FTSW Thresholds and Std. Error | Slope 1 and Std. Error | Slope 2 and Std. Error |
|---|---|---|---|
| ICC 16340_Stg-D-P | 0.58 ± 0.02 | 1.92 ± 0.08 | −0.59 ± 0.23 |
| KAK2_R-P | 0.54 ± 0.03 | 1.72 ± 0.05 | 0.20 ± 0.26 |
| JGK 1_R-P | 0.54 ± 0.03 | 1.88 ± 0.06 | −0.12 ± 0.24 |
| ICCX-060119-107 (KAK2) | 0.54 ± 0.03 | 1.85 ± 0.08 | −0.10 ± 0.22 |
| ICCX-060119-113 (KAK2) | 0.48 ± 0.03 | 1.98 ± 0.09 | 0.06 ± 0.17 |
| ICCX-060119-116 (KAK2) | 0.58 ± 0.03 | 1.64 ± 0.07 | 0.01 ± 0.19 |
| ICCX-060119-123 (KAK2) | 0.62 ± 0.05 | 1.69 ± 0.09 | −0.32 ± 0.53 |
| ICCX-060121-125 (JGK1) | 0.51 ± 0.02 | 1.87 ± 0.06 | −0.01 ± 0.17 |
| ICCX-060121-128 (JGK1) | 0.60 ± 0.03 | 1.63 ± 0.06 | 0.05 ± 0.28 |
| ICCX-060121-129 (JGK1) | 0.55 ± 0.03 | 1.72 ± 0.07 | 0.14 ± 0.18 |
| Genotypes | TR Response to VPD at Outdoor | TR Response to VPD at Growth Chamber | |||
|---|---|---|---|---|---|
| KAK-2 Background | Mean TR & SE LSD (0.01) = 0.09 | Slope at high VPD & SE LSD (0.01) = 0.59 | Mean TR & SE LSD (0.001) = 0.35 | Slope & SE LSD (0.01) = 0.04 | R2 |
| ICC 16340_Stay-green_Donor Parent | 1.31 ± 0.04a | 5.48 ± 0.18a | 2.02 ± 0.06a | 0.52 ± 0.06a | 0.94 |
| KAK2_Recurrent parent | 0.95 ± 0.02b | 4.18 ± 0.08b | 1.45 ± 0.06b | 0.41 ± 0.04b | 0.94 |
| ICCX-060119-107 (KAK2 Background) | 0.98 ± 0.04b | 4.16 ± 0.18b | 1.37 ± 0.04b | 0.43 ± 0.04b | 0.95 |
| ICCX-060119-113 (KAK2 Background) | 1.04 ± 0.02b | 4.23 ± 0.06b | 1.49 ± 0.06b | 0.46 ± 0.04b | 0.95 |
| ICCX-060119-116 (KAK2 Background) | 0.94 ± 0.04b | 4.19 ± 0.20b | 1.45 ± 0.08b | 0.47 ± 0.05b | 0.94 |
| ICCX-060119-123 (KAK2 Background) | 1.03 ± 0.02b | 4.16 ± 0.16b | 1.63 ± 0.04b | 0.40 ± 0.05b | 0.91 |
| JGK-1 Background | Mean TR & SE LSD (0.001) = 0.12 | Slope at high VPD & SE LSD (0.01) = 0.46 | Mean TR & SE LSD (0.01) = 0.25 | Slope & SE LSD (0.01) = 0.033 | R2 |
| ICC 16340_Stay-green_Donor Parent | 1.31 ± 0.04a | 5.48 ± 0.18a | 2.02 ± 0.06a | 0.52 ± 0.06a | 0.94 |
| JGK 1_Recurrent parent | 1.14 ± 0.04b | 4.77 ± 0.16b | 1.70 ± 0.06b | 0.45 ± 0.04b | 0.95 |
| ICCX-060121-125 (JGK1 Background) | 0.99 ± 0.03bc | 4.09 ± 0.01c | 1.53 ± 0.06b | 0.42 ± 0.04b | 0.95 |
| ICCX-060121-128 (JGK1 Background) | 0.90 ± 0.03c | 3.82 ± 0.08c | 1.52 ± 0.08b | 0.38 ± 0.04b | 0.94 |
| ICCX-060121-129 (JGK1 Background) | 0.90 ± 0.01c | 3.78 ± 0.05c | 1.52 ± 0.05b | 0.37 ± 0.04b | 0.94 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sivasakthi, K.; Marques, E.; Kalungwana, N.; Carrasquilla-Garcia, N.; Chang, P.L.; Bergmann, E.M.; Bueno, E.; Cordeiro, M.; Sani, S.G.A.S.; Udupa, S.M.; et al. Functional Dissection of the Chickpea (Cicer arietinum L.) Stay-Green Phenotype Associated with Molecular Variation at an Ortholog of Mendel’s I Gene for Cotyledon Color: Implications for Crop Production and Carotenoid Biofortification. Int. J. Mol. Sci. 2019, 20, 5562. https://doi.org/10.3390/ijms20225562
Sivasakthi K, Marques E, Kalungwana N, Carrasquilla-Garcia N, Chang PL, Bergmann EM, Bueno E, Cordeiro M, Sani SGAS, Udupa SM, et al. Functional Dissection of the Chickpea (Cicer arietinum L.) Stay-Green Phenotype Associated with Molecular Variation at an Ortholog of Mendel’s I Gene for Cotyledon Color: Implications for Crop Production and Carotenoid Biofortification. International Journal of Molecular Sciences. 2019; 20(22):5562. https://doi.org/10.3390/ijms20225562
Chicago/Turabian StyleSivasakthi, Kaliamoorthy, Edward Marques, Ng’andwe Kalungwana, Noelia Carrasquilla-Garcia, Peter L. Chang, Emily M. Bergmann, Erika Bueno, Matilde Cordeiro, Syed Gul A.S. Sani, Sripada M. Udupa, and et al. 2019. "Functional Dissection of the Chickpea (Cicer arietinum L.) Stay-Green Phenotype Associated with Molecular Variation at an Ortholog of Mendel’s I Gene for Cotyledon Color: Implications for Crop Production and Carotenoid Biofortification" International Journal of Molecular Sciences 20, no. 22: 5562. https://doi.org/10.3390/ijms20225562
APA StyleSivasakthi, K., Marques, E., Kalungwana, N., Carrasquilla-Garcia, N., Chang, P. L., Bergmann, E. M., Bueno, E., Cordeiro, M., Sani, S. G. A. S., Udupa, S. M., Rather, I. A., Rouf Mir, R., Vadez, V., Vandemark, G. J., Gaur, P. M., Cook, D. R., Boesch, C., von Wettberg, E. J. B., Kholova, J., & Penmetsa, R. V. (2019). Functional Dissection of the Chickpea (Cicer arietinum L.) Stay-Green Phenotype Associated with Molecular Variation at an Ortholog of Mendel’s I Gene for Cotyledon Color: Implications for Crop Production and Carotenoid Biofortification. International Journal of Molecular Sciences, 20(22), 5562. https://doi.org/10.3390/ijms20225562