Impaired Expression of Ectonucleotidases in Ectopic and Eutopic Endometrial Tissue Is in Favor of ATP Accumulation in the Tissue Microenvironment in Endometriosis

 , , , , ,
, , , , ,  , ,
, ,
Abstract
1. Introduction
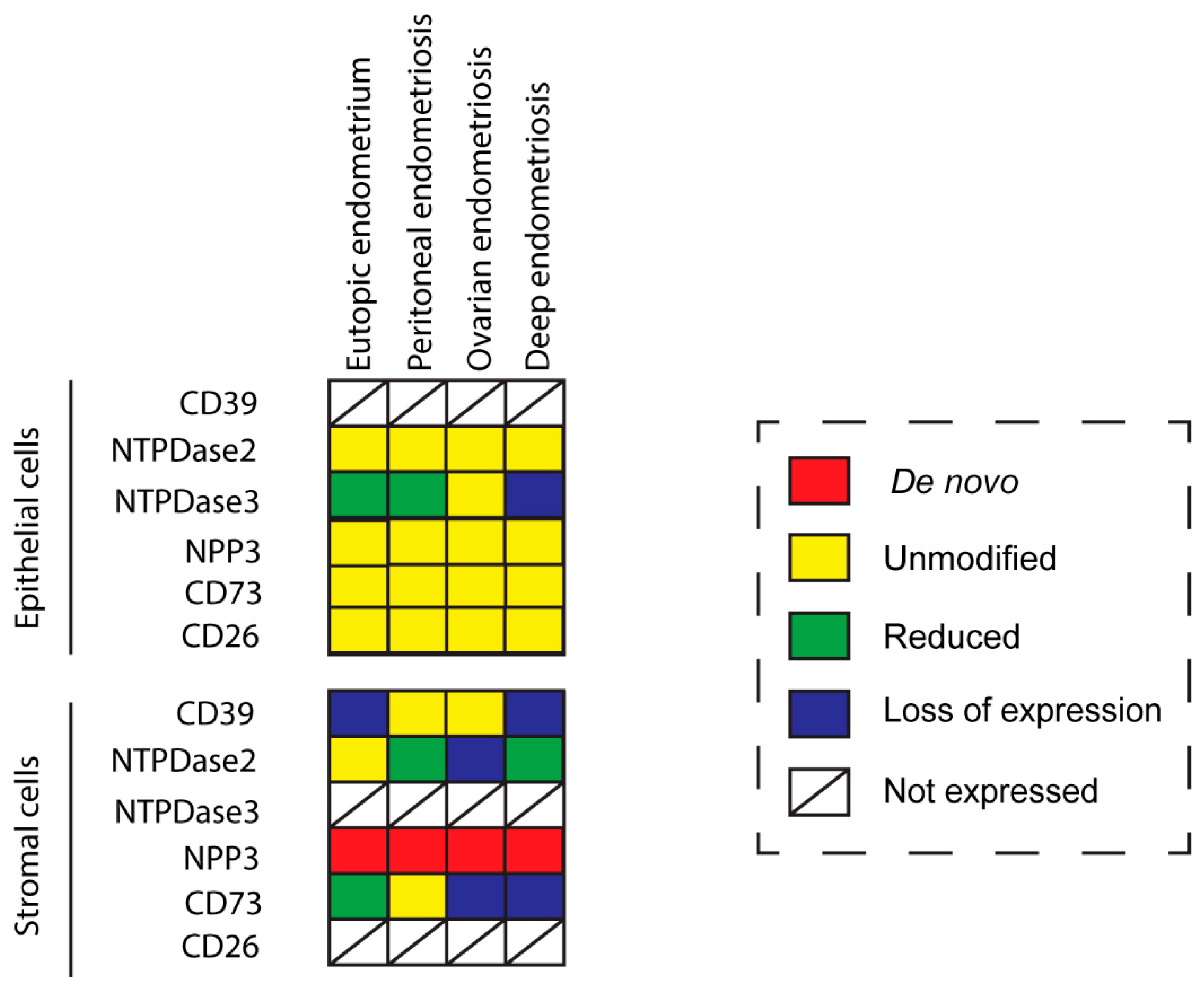
2. Results
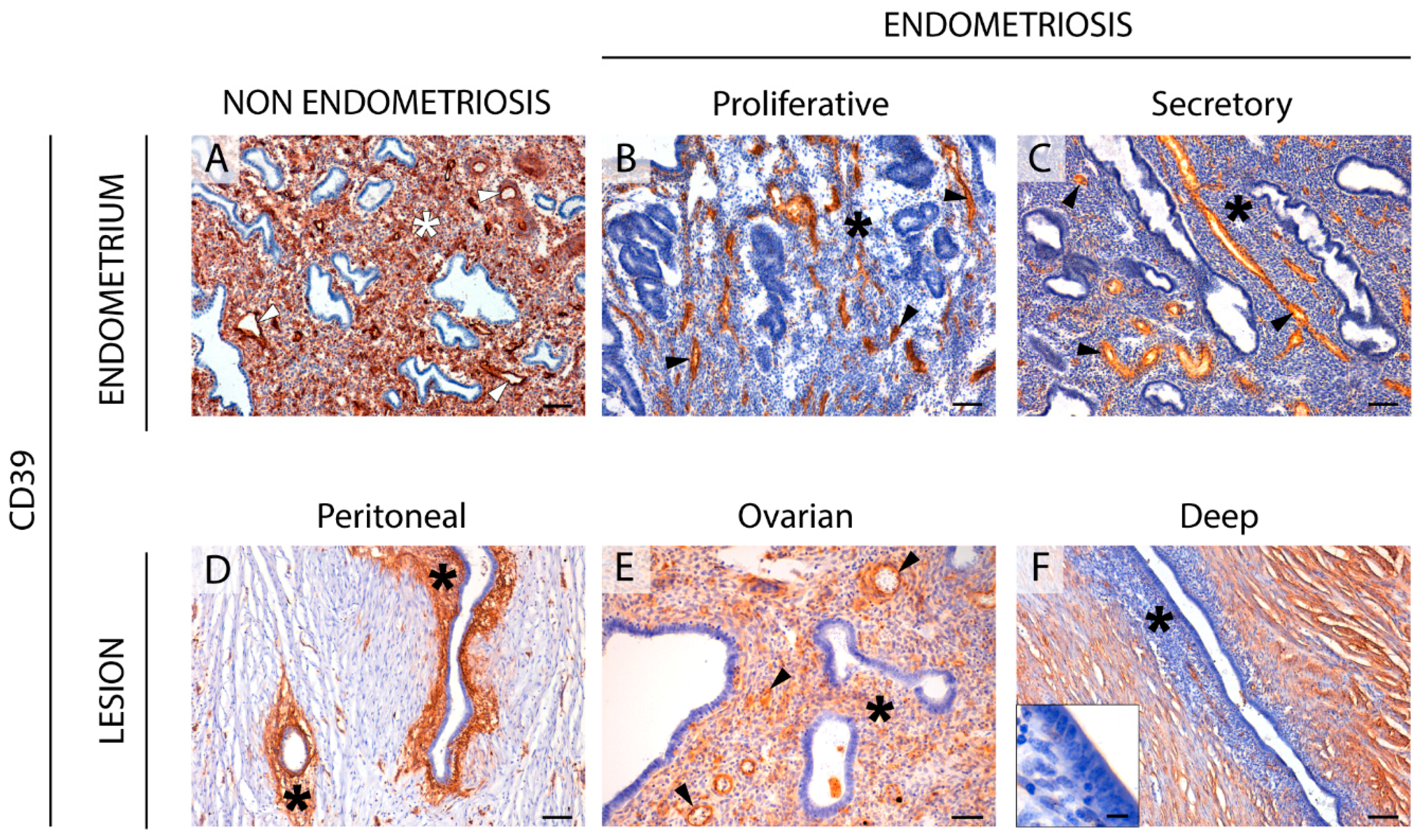
2.1. CD39 Expression in the Eutopic and Ectopic Endometrial Tissues
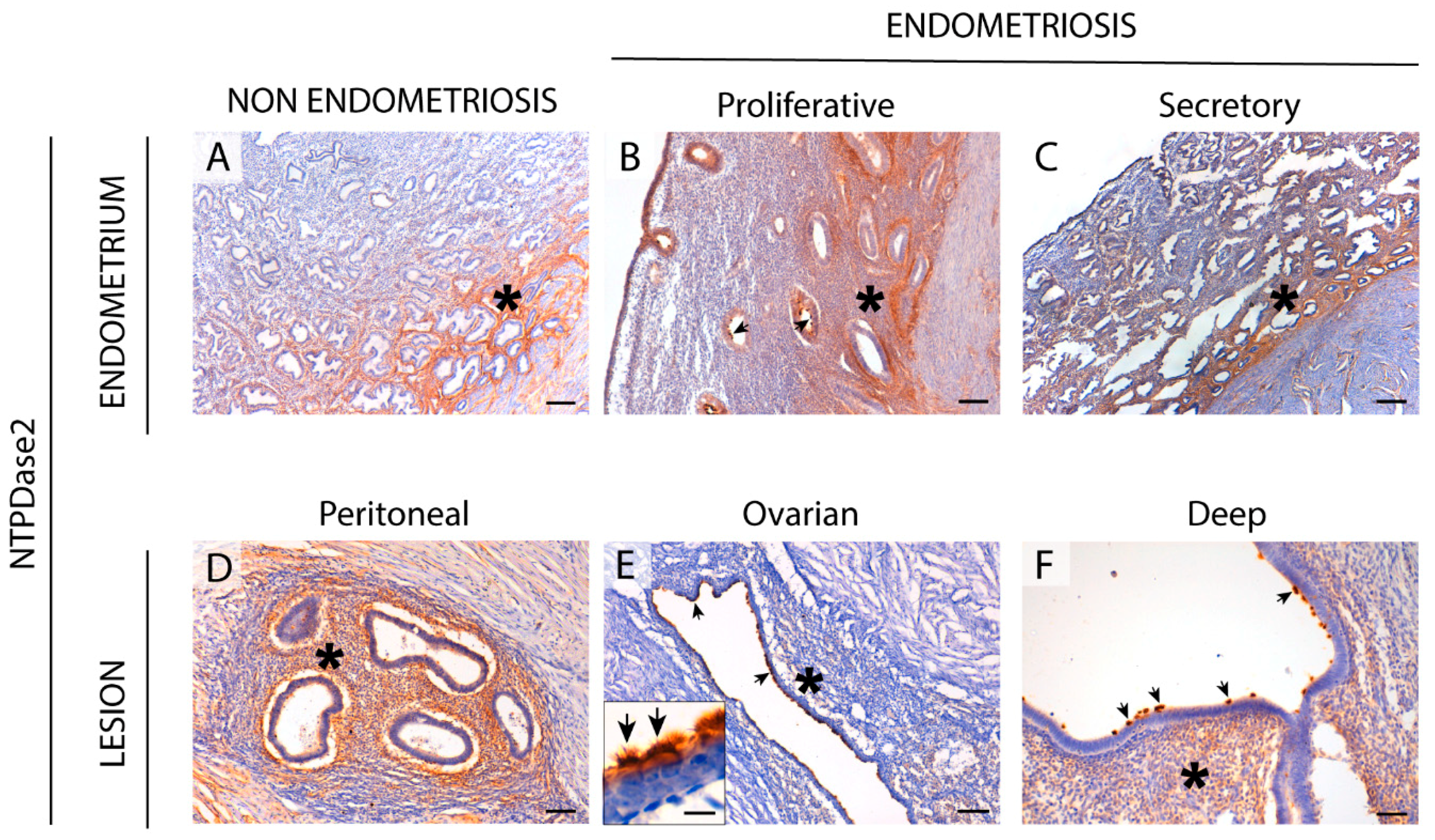
2.2. NTPDase2 Expression in the Eutopic and Ectopic Endometrial Tissues
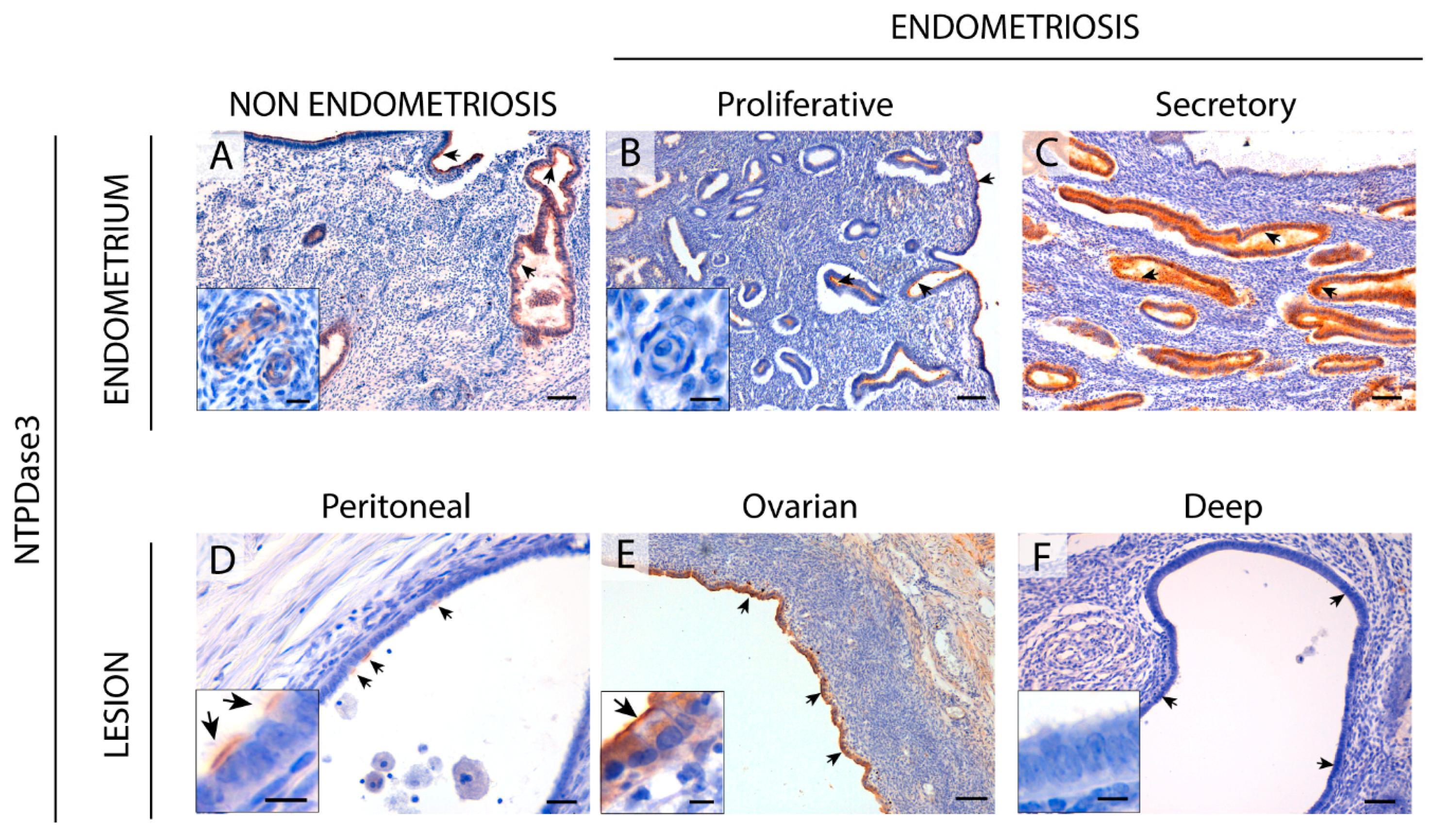
2.3. NTPDase3 Expression in the Eutopic and Ectopic Endometrial Tissues
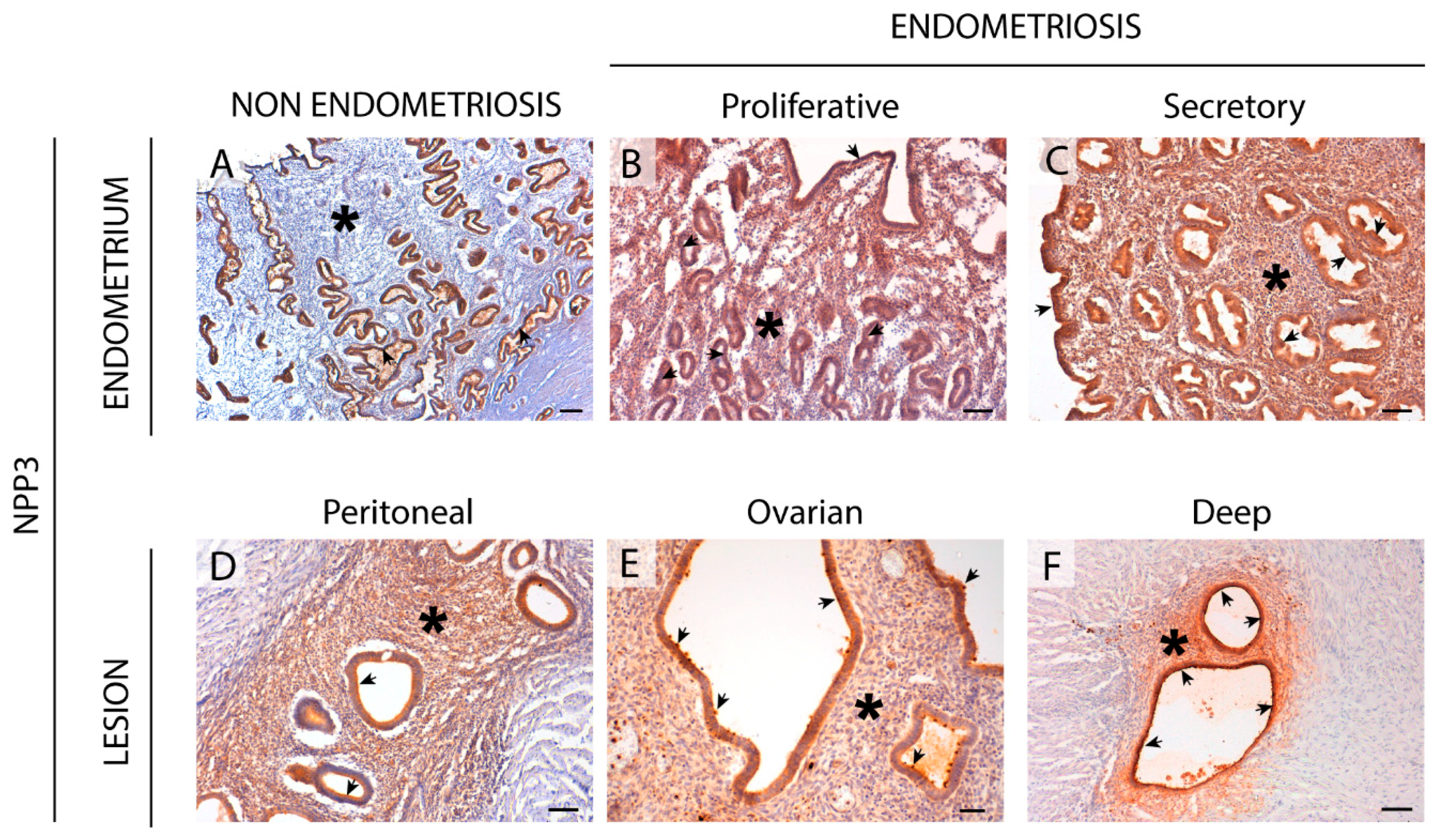
2.4. NPP3 Expression in the Eutopic and Ectopic Endometrial Tissues
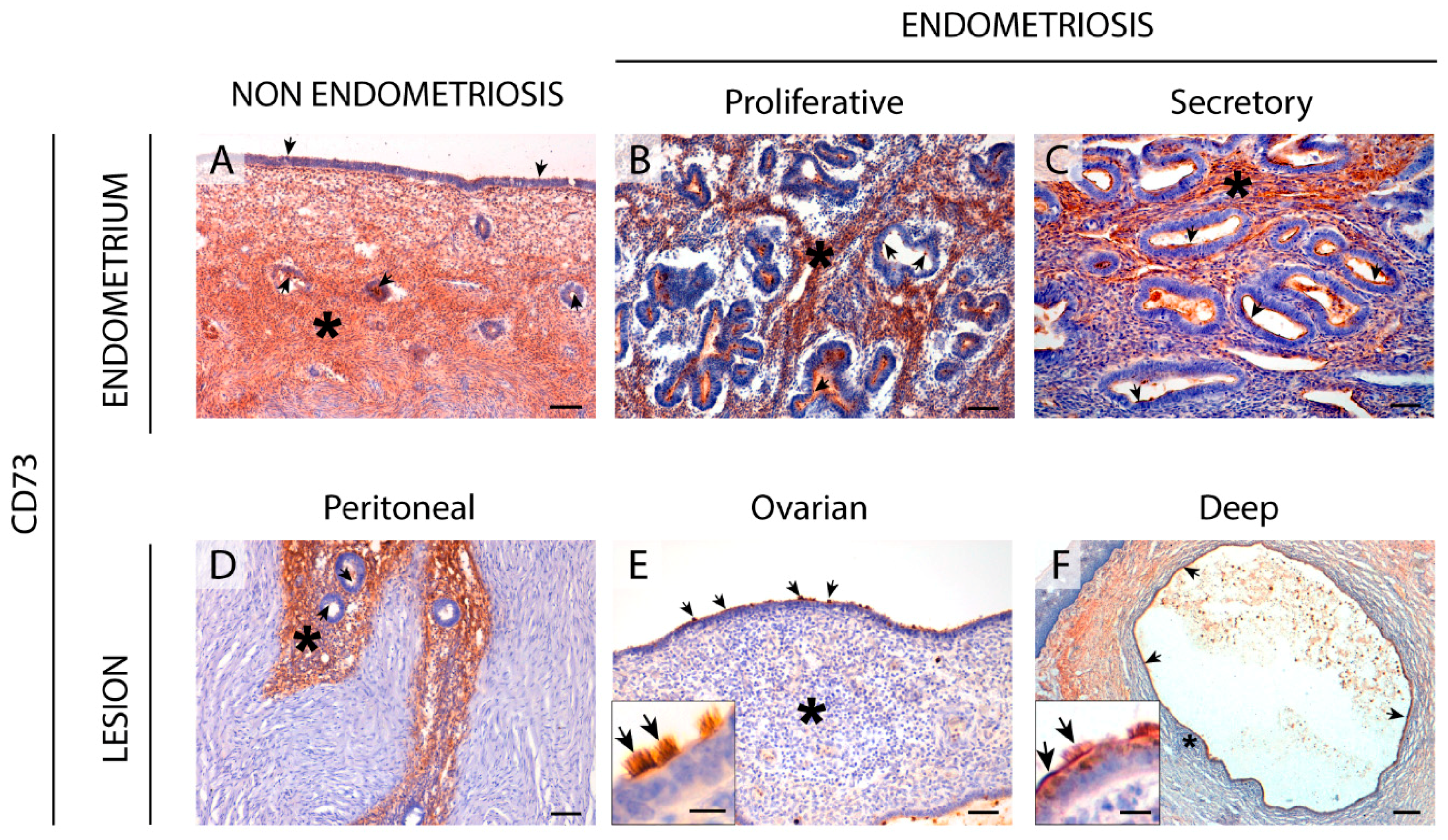
2.5. CD73 Expression in the Eutopic and Ectopic Endometrial Tissues
2.6. Enzyme Dipeptidyl Peptidase IV/CD26 Expression in the Eutopic and Ectopic Endometrial Tissues
3. Discussion
4. Materials and Methods
4.1. Samples
4.2. Antibodies
4.3. Immunolabeling Experiments
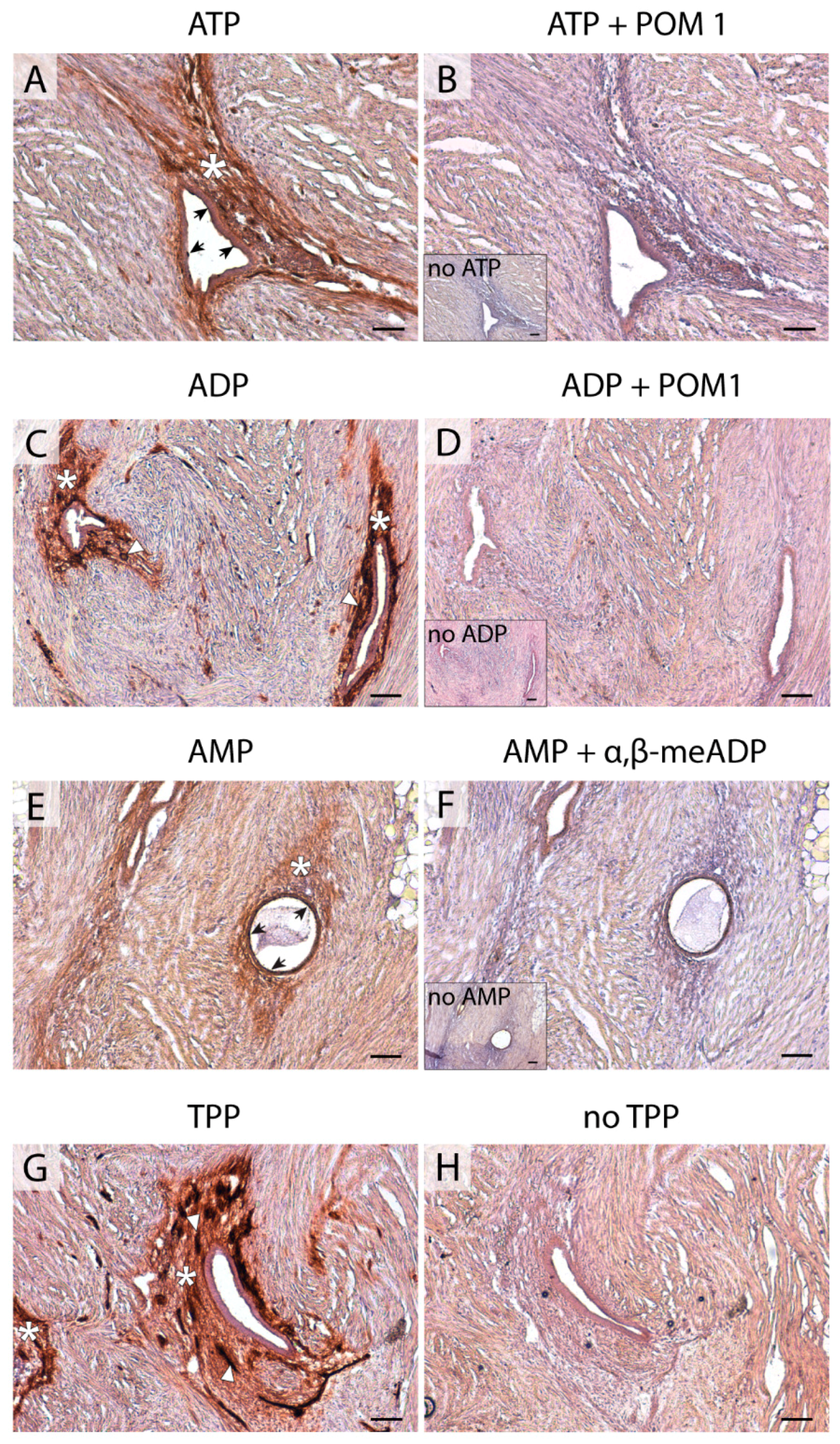
4.4. In situ ATPase, ADPase, AMPase, and TPPase Activity Experiments
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ATP | Adenosine triphosphate |
| E-NPP | Ecto-nucleotide pyrophophotase/phosphodiesterase |
| E-NTPDase | Ecto-nucleoside triphosphate diphosphohydrolase |
| ADP | Adenosine diphosphate |
| AMP | Adenosine monophosphate |
| 5′-NT | 5′-nucleotidase |
| ADA | Adenosine deaminase |
| eMSC | Endometrial mesenchymal stem cell |
| SUSD2 | Sushi domain containing 2 |
| SMA | Smooth muscle actin |
| TPP | Thiamine pyrophosphate |
| α, β-meADP | α, β-methylene-ADP |
| ATP1B4 | ATPase Na+/K+ Transporting Family Member Beta 4 |
| DPPIV | Dipeptidyl peptidase IV |
| HRP | Horseradish peroxidase |
| DAB | 3, 3′-diaminobenzidine substrate |
| AP | Alkaline phosphatase |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples Stained | NTPDase1 (CD39) | NTPDase2 | NTPDase3 | NPP3 | 5′-NT (CD73) | CD26 |
|---|---|---|---|---|---|---|
| Positive Cases/Total of Samples (% of Tissues Stained) | ||||||
| Eutopic endometrium | ||||||
| Surface epithelium | 1/14 (7.1%) | 14/14 (100%) | 7/13 (53.8%) | 12/13 (92.3%) | 9/12 (75%) | 7/14 (50%) |
| Glandular epithelium | 1/20 (5%) | 20/20 (100%) | 16/20 (80%) | 18/19 (94.7%) | 20/20 (100%) | 15/20 (75%) |
| Endometrial stromal cells | 7/20 (35%) | 20/20 (100%) | 0/20 (0%) | 15/19 (78.9%) | 12/20 (60%) | 0/20 (0%) |
| Spiral arteries | 19/20 (95%) | 3/20 (15%) | 5/20 (25%) | 16/18 (88.9%) | 3/20 (15%) | 3/19 (15.8%) |
| Peritoneal endometriosis | ||||||
| Endometrial epithelial cells | 1/7 (14.3%) | 7/7 (100%) | 7/7 (100%) | 7/7 (100%) | 5/7 (71.4%) | 4/7 (57.1%) |
| Endometrial stromal cells | 6/7 (85.7%) | 7/7 (100%) | 0/7 (0%) | 7/7 (100%) | 5/7 (71.4%) | 2/7 (28.6%) |
| Vessels of the lesion | 5/7 (71.4%) | 0/7 (0%) | 0/7 (0%) | 0/7 (0%) | 1/7 (14.28%) | 0/7 (0%) |
| Ovarian endometriosis | ||||||
| Endometrial epithelial cells | 8/28 (28.6%) | 20/28 (71.4%) | 16/28 (57.1%) | 24/28 (85.7%) | 24/28 (85.7%) | 19/28 (67.9%) |
| Endometrial stromal cells | 16/27 (59.3%) | 10/27 (37%) | 3/27 (11.1%) | 16/27 (59.3%) | 13/27 (48.1%) | 2/27 (7.4%) |
| Vessels of the lesion | 23/28 (82.1%) | 1/28 (3.6%) | 2/28 (7.1%) | 8/28 (28.6%) | 3/28 (10.7%) | 3/28 (10.7%) |
| Deep endometriosis | ||||||
| Endometrial epithelial cells | 6/28 (21.4%) | 23/26 (88.5%) | 12/27 (44.4%) | 24/28 (85.7%) | 17/28 (60.7%) | 15/28 (53.6%) |
| Endometrial stromal cells | 9/25 (36%) | 17/26 (65.4%) | 1/27 (3.7%) | 18/27 (66.7%) | 6/27 (22.2%) | 1/27 (3.7%) |
| Vessels of the lesion | 11/24 (45.8%) | 5/25 (20%) | 1/27 (3.7%) | 7/27 (25.9%) | 7/27 (25.9%) | 0/27 (0%) |
References
- Ponandai-Srinivasan, S.; Andersson, K.L.; Nister, M.; Saare, M.; Hassan, H.A.; Varghese, S.J.; Peters, M.; Salumets, A.; Gemzell-Danielsson, K.; Lalitkumar, P.G.L. Aberrant expression of genes associated with stemness and cancer in endometria and endometrioma in a subset of women with endometriosis. Hum. Reprod. 2018, 33, 1924–1938. [Google Scholar] [CrossRef] [PubMed]
- Grandi, G.; Mueller, M.D.; Papadia, A.; Kocbek, V.; Bersinger, N.A.; Petraglia, F.; Cagnacci, A.; McKinnon, B. Inflammation influences steroid hormone receptors targeted by progestins in endometrial stromal cells from women with endometriosis. J. Reprod. Immunol. 2016, 117, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Nothnick, W.; Alali, Z. Recent advances in the understanding of endometriosis: The role of inflammatory mediators in disease pathogenesis and treatment. F1000Research 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.H.; Khalaj, K.; Young, S.L.; Lessey, B.A.; Koti, M.; Tayade, C. Immune-inflammation gene signatures in endometriosis patients. Fertil. Steril. 2016, 106, 1420–1431. [Google Scholar] [CrossRef]
- Zhang, T.; De Carolis, C.; Man, G.C.W.; Wang, C.C. The link between immunity, autoimmunity and endometriosis: A literature update. Autoimmun. Rev. 2018, 17, 945–955. [Google Scholar] [CrossRef]
- Yegutkin, G.G. Enzymes involved in metabolism of extracellular nucleotides and nucleosides: Functional implications and measurement of activities. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 473–497. [Google Scholar] [CrossRef]
- Ziganshin, A.U.; Zaitcev, A.P.; Khasanov, A.A.; Shamsutdinov, A.F.; Burnstock, G. Term-dependency of P2 receptor-mediated contractile responses of isolated human pregnant uterus. Eur. J. Obstet. Gynecol. Reprod. Biol. 2006, 129, 128–134. [Google Scholar] [CrossRef]
- Aliagas, E.; Vidal, A.; Torrejon-Escribano, B.; Taco Mdel, R.; Ponce, J.; de Aranda, I.G.; Sevigny, J.; Condom, E.; Martin-Satue, M. Ecto-nucleotidases distribution in human cyclic and postmenopausic endometrium. Purinergic Signal. 2013, 9, 227–237. [Google Scholar] [CrossRef]
- Burnstock, G. Purinergic signalling in the reproductive system in health and disease. Purinergic Signal. 2014, 10, 157–187. [Google Scholar] [CrossRef]
- Bellezza, I.; Minelli, A. Adenosine in sperm physiology. Mol. Asp. Med. 2017, 55, 102–109. [Google Scholar] [CrossRef]
- Villamonte, M.L.; Torrejon-Escribano, B.; Rodriguez-Martinez, A.; Trapero, C.; Vidal, A.; Gomez de Aranda, I.; Sevigny, J.; Matias-Guiu, X.; Martin-Satue, M. Characterization of ecto-nucleotidases in human oviducts with an improved approach simultaneously identifying protein expression and in situ enzyme activity. Histochem. Cell Biol. 2018, 149, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Barrera, N.P.; Morales, B.; Villalon, M. Plasma and intracellular membrane inositol 1,4,5-trisphosphate receptors mediate the Ca(2+) increase associated with the ATP-induced increase in ciliary beat frequency. Am. J. Physiol. Cell Physiol. 2004, 287, C1114–C1124. [Google Scholar] [CrossRef] [PubMed]
- Yegutkin, G.G. Nucleotide- and nucleoside-converting ectoenzymes: Important modulators of purinergic signalling cascade. Biochim. Biophys. Acta 2008, 1783, 673–694. [Google Scholar] [CrossRef] [PubMed]
- Trapero, C.; Vidal, A.; Rodriguez-Martinez, A.; Sevigny, J.; Ponce, J.; Coroleu, B.; Matias-Guiu, X.; Martin-Satue, M. The ectonucleoside triphosphate diphosphohydrolase-2 (NTPDase2) in human endometrium: A novel marker of basal stroma and mesenchymal stem cells. Purinergic Signal. 2019, 15, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Texido, L.; Romero, C.; Vidal, A.; Garcia-Valero, J.; Fernandez Montoli, M.E.; Baixeras, N.; Condom, E.; Ponce, J.; Garcia-Tejedor, A.; Martin-Satue, M. Ecto-nucleotidases activities in the contents of ovarian endometriomas: Potential biomarkers of endometriosis. Mediat. Inflamm. 2014, 2014, 120673. [Google Scholar] [CrossRef]
- Trapero, C.; Jover, L.; Fernandez-Montoli, M.E.; Garcia-Tejedor, A.; Vidal, A.; Gomez de Aranda, I.; Ponce, J.; Matias-Guiu, X.; Martin-Satue, M. Analysis of the ectoenzymes ADA, ALP, ENPP1, and ENPP3, in the contents of ovarian endometriomas as candidate biomarkers of endometriosis. Am. J. Reprod. Immunol. 2018, 79. [Google Scholar] [CrossRef] [PubMed]
- Bono, M.R.; Fernandez, D.; Flores-Santibanez, F.; Rosemblatt, M.; Sauma, D. CD73 and CD39 ectonucleotidases in T cell differentiation: Beyond immunosuppression. FEBS Lett. 2015, 589, 3454–3460. [Google Scholar] [CrossRef]
- Aliagas, E.; Torrejon-Escribano, B.; Lavoie, E.G.; de Aranda, I.G.; Sevigny, J.; Solsona, C.; Martin-Satue, M. Changes in expression and activity levels of ecto-5′-nucleotidase/CD73 along the mouse female estrous cycle. Acta Physiol. 2010, 199, 191–197. [Google Scholar] [CrossRef]
- Mate, G.; Bernstein, L.R.; Torok, A.L. Endometriosis is a cause of infertility. Does reactive oxygen damage to gametes and embryos play a key role in the pathogenesis of infertility caused by endometriosis? Front. Endocrinol. 2018, 9, 725. [Google Scholar] [CrossRef]
- Ahn, S.H.; Monsanto, S.P.; Miller, C.; Singh, S.S.; Thomas, R.; Tayade, C. Pathophysiology and immune dysfunction in endometriosis. Biomed. Res. Int. 2015, 2015, 795976. [Google Scholar] [CrossRef]
- Faas, M.M.; Saez, T.; de Vos, P. Extracellular ATP and adenosine: The yin and yang in immune responses? Mol. Asp. Med. 2017, 55, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Gao, L.; Yi, Y.; Han, H.; Cheng, H.; Ye, X.; Ma, R.; Sun, K.; Cui, H.; Chang, X. Adenosine triphosphate regresses endometrial explants in a rat model of endometriosis. Reprod. Sci. 2016, 23, 924–930. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Niu, Y.; Feng, J.; Guo, H.; Ye, X.; Cui, H. Use of proteomic analysis of endometriosis to identify different protein expression in patients with endometriosis versus normal controls. Fertil. Steril. 2006, 86, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Liu, Y.N.; Li, Y.; Tian, L.; Ye, X.; Cui, H.; Chang, X.H. Identification of biomarkers for endometriosis using clinical proteomics. Chin. Med. J. 2015, 128, 520–527. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Zhu, L.; Tian, Y.; Zhu, T.; Huang, X.; Zhang, X. P2X3 receptor involvement in endometriosis pain via ERK signaling pathway. PLoS ONE 2017, 12, e0184647. [Google Scholar] [CrossRef]
- Yuan, M.; Ding, S.; Meng, T.; Lu, B.; Shao, S.; Zhang, X.; Yuan, H.; Hu, F. Effect of A-317491 delivered by glycolipid-like polymer micelles on endometriosis pain. Int. J. Nanomed. 2017, 12, 8171–8183. [Google Scholar] [CrossRef]
- Alimi, Y.; Iwanaga, J.; Loukas, M.; Tubbs, R.S. The clinical anatomy of endometriosis: A review. Cureus 2018, 10, e3361. [Google Scholar] [CrossRef]
- Bowser, J.L.; Blackburn, M.R.; Shipley, G.L.; Molina, J.G.; Dunner, K., Jr.; Broaddus, R.R. Loss of CD73-mediated actin polymerization promotes endometrial tumor progression. J. Clin. Investig. 2016, 126, 220–238. [Google Scholar] [CrossRef]
- Sampson, J. Peritoneal endometriosis due to the menstrual dissemination of endometrial tissue into the peritoneal cavity. Am. J. Obstet. Gynecol. 1927, 14, 422–469. [Google Scholar] [CrossRef]
- Hood, B.L.; Liu, B.; Alkhas, A.; Shoji, Y.; Challa, R.; Wang, G.; Ferguson, S.; Oliver, J.; Mitchell, D.; Bateman, N.W.; et al. Proteomics of the human endometrial glandular epithelium and stroma from the proliferative and secretory phases of the menstrual cycle. Biol. Reprod. 2015, 92, 106. [Google Scholar] [CrossRef][Green Version]
- Boggavarapu, N.R.; Lalitkumar, S.; Joshua, V.; Kasvandik, S.; Salumets, A.; Lalitkumar, P.G.; Gemzell-Danielsson, K. Compartmentalized gene expression profiling of receptive endometrium reveals progesterone regulated ENPP3 is differentially expressed and secreted in glycosylated form. Sci. Rep. 2016, 6, 33811. [Google Scholar] [CrossRef] [PubMed]
- Donate, F.; Raitano, A.; Morrison, K.; An, Z.; Capo, L.; Avina, H.; Karki, S.; Yang, P.; Ou, J.; Moriya, R.; et al. AGS16F is a novel antibody drug conjugate directed against ENPP3 for the treatment of renal cell carcinoma. Clin. Cancer Res. 2016, 22, 1989–1999. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.H.; Takeda, K. Regulation of allergic inflammation by the ectoenzyme E-NPP3 (CD203c) on basophils and mast cells. Semin. Immunopathol. 2016, 38, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Xin, A.; Qu, R.; Zhang, W.; Li, L.; Chen, J.; Lu, X.; Gu, Y.; Li, J.; Sun, X. Expression of ENPP3 in human cyclic endometrium: A novel molecule involved in embryo implantation. Reprod. Fertil. Dev. 2018, 30, 1277–1285. [Google Scholar] [CrossRef]
- Deissler, H.; Blass-Kampmann, S.; Bruyneel, E.; Mareel, M.; Rajewsky, M.F. Neural cell surface differentiation antigen gp130(RB13-6) induces fibroblasts and glioma cells to express astroglial proteins and invasive properties. FASEB J. 1999, 13, 657–666. [Google Scholar] [CrossRef]
- Thompson, J.A.; Motzer, R.J.; Molina, A.M.; Choueiri, T.K.; Heath, E.I.; Redman, B.G.; Sangha, R.S.; Ernst, D.S.; Pili, R.; Kim, S.K.; et al. Phase I Trials of Anti-ENPP3 Antibody-drug conjugates in advanced refractory renal cell carcinomas. Clin. Cancer Res. 2018, 24, 4399–4406. [Google Scholar] [CrossRef]
- Rockenbach, L.; Braganhol, E.; Dietrich, F.; Figueiro, F.; Pugliese, M.; Edelweiss, M.I.; Morrone, F.B.; Sevigny, J.; Battastini, A.M. NTPDase3 and ecto-5′-nucleotidase/CD73 are differentially expressed during mouse bladder cancer progression. Purinergic Signal. 2014, 10, 421–430. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, X.; Wu, R.; Huang, Q.; Jiang, Y.; Qin, J.; Yao, F.; Jin, G.; Zhang, Y. DPPIV promotes endometrial carcinoma cell proliferation, invasion and tumorigenesis. Oncotarget 2017, 8, 8679–8692. [Google Scholar] [CrossRef]
- Tan, C.W.; Lee, Y.H.; Tan, H.H.; Lau, M.S.; Choolani, M.; Griffith, L.; Chan, J.K. CD26/DPPIV down-regulation in endometrial stromal cell migration in endometriosis. Fertil. Steril. 2014, 102, 167–177. [Google Scholar] [CrossRef]
- Busacca, M.; Chiaffarino, F.; Candiani, M.; Vignali, M.; Bertulessi, C.; Oggioni, G.; Parazzini, F. Determinants of long-term clinically detected recurrence rates of deep, ovarian, and pelvic endometriosis. Am. J. Obstet. Gynecol. 2006, 195, 426–432. [Google Scholar] [CrossRef]
- Koninckx, P.R.; Meuleman, C.; Demeyere, S.; Lesaffre, E.; Cornillie, F.J. Suggestive evidence that pelvic endometriosis is a progressive disease, whereas deeply infiltrating endometriosis is associated with pelvic pain. Fertil. Steril. 1991, 55, 759–765. [Google Scholar] [CrossRef]
- Burnstock, G. Purinergic mechanisms and pain. Adv. Pharmacol. 2016, 75, 91–137. [Google Scholar] [CrossRef] [PubMed]
- Fung, C.Y.; Marcus, A.J.; Broekman, M.J.; Mahaut-Smith, M.P. P2X(1) receptor inhibition and soluble CD39 administration as novel approaches to widen the cardiovascular therapeutic window. Trends Cardiovasc. Med. 2009, 19, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Wachstein, M.; Meisel, E. Histochemistry of hepatic phosphatases of a physiologic pH; with special reference to the demonstration of bile canaliculi. Am. J. Clin. Pathol. 1957, 27, 13–23. [Google Scholar] [CrossRef]
- Martín-Satué, M.; Rodríguez-Martínez, A.; Trapero, C. In situ identification of ectoenzymes involved in the hydrolysis of extracellular nucleotides. In Immunohistochemistry; IntechOpen: London, UK, 2019. [Google Scholar]








| NTPDase1 (CD39) | NTPDase2 | NTPDase3 | NPP3 | 5′-NT (CD73) | CD26 | |
|---|---|---|---|---|---|---|
| Proliferative endometrium | ||||||
| Surface epithelium | - | +++ | + | ++ | ++ | - |
| Glandular epithelium | ||||||
| Functional layer | - | +++ | ++ | ++ | +++ | ++/- 1 |
| Basal layer | - | +++ | + | + | +++ | +++ |
| Endometrial stromal cells | - | +++ | - | +++ | ++ | - |
| Spiral arteries | +++ | - | - | + | - | - |
| Secretory endometrium | ||||||
| Surface epithelium | - | +++ | +/- 1 | +++ | +++ | + |
| Glandular epithelium | ||||||
| Functional layer | - | +++ | +++ | +++ | ++ | +++ |
| Basal layer | - | +++ | +++ | +++ | +++ | ++ |
| Endometrial stromal cells | - | +++ | - | + | ++ | - |
| Spiral arteries | +++ | - | - | + | - | - |
| Atrophic endometrium | ||||||
| Surface epithelium | - | +++ | - | + | + | ++/- 1 |
| Glandular epithelium | - | +++ | - | ++ | ++ | - |
| Endometrial stromal cells | ++ | +++ | - | ++ | - | - |
| Vessels | ++ | - | - | + | - | - |
| NTPDase1 (CD39) | NTPDase2 | NTPDase3 | NPP3 | 5′-NT (CD73) | CD26 | |
|---|---|---|---|---|---|---|
| Peritoneal endometriosis | ||||||
| Endometrial epithelial cells | - | +++ | + | +++ | +++ | +++ |
| Endometrial stromal cells | +++ | ++ | - | ++ | ++ | - |
| Vessels of the lesion | +++ | - | - | - | - | - |
| Ovarian endometriosis | ||||||
| Endometrial epithelial cells | - | +++ | +++ | +++ | +++ | +++ |
| Endometrial stromal cells | ++ | - | - | + | - | - |
| Vessels of the lesion | +++ | - | - | - | - | - |
| Deep endometriosis | ||||||
| Endometrial epithelial cells | - | +++ | - | ++ | +++ | +++ |
| Endometrial stromal cells | - | ++ | - | + | - | - |
| Vessels of the lesion | - | - | - | - | - | - |
| Type of Endometrium | Number of Cases | Age (years) ± Standard Deviation | Average (Range) |
|---|---|---|---|
| Proliferative | 12 | 44.33 ± 3.11 | 39–51 |
| Secretory | 10 | 45.00 ± 5.21 | 38–53 |
| Atrophic | 3 | 43.00 ± 4.00 | 39–47 |
| Type of Lesion | |||
| Peritoneal | 7 | 40.29 ± 4.96 | 35–46 |
| Ovarian | 27 | 43.42 ± 8.36 | 23–57 |
| Deep | 21 | 38.00 ± 5.73 | 27–47 |
| Antibody Specificity | Name/Clone | Source | Supplier | Dilution |
|---|---|---|---|---|
| NTPDase1 (CD39) | BU-61 | Mouse | Ancell (188-820) | 1:500 |
| NTPDase2 | - | Rabbit | Enzo (ALX-215-045) | 1:100 |
| NTPDase2 | H9s | Mouse | http://ectonucleotidases-ab.com | 1:400 |
| NTPDase3 | B3S10 | Mouse | http://ectonucleotidases-ab.com | 1:500 |
| NPP3 | NP4D6 | Mouse | Abcam (ab90754) | 1:100 |
| 5′-nucleotidase (CD73) | 4G4 | Mouse | Abcam (ab81720) | 1:50 |
| CD26 | 202-36 | Mouse | Abcam (ab3154) | 1:100 |
| CD26 | 202-36 | Mouse | NovusBio (NBP2-44571) | 1:100 |
| SUSD2 | - | Rabbit | Abcam (ab121214) | 1:400 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trapero, C.; Vidal, A.; Fernández-Montolí, M.E.; Coroleu, B.; Tresserra, F.; Barri, P.; Gómez de Aranda, I.; Sévigny, J.; Ponce, J.; Matias-Guiu, X.; et al. Impaired Expression of Ectonucleotidases in Ectopic and Eutopic Endometrial Tissue Is in Favor of ATP Accumulation in the Tissue Microenvironment in Endometriosis. Int. J. Mol. Sci. 2019, 20, 5532. https://doi.org/10.3390/ijms20225532
Trapero C, Vidal A, Fernández-Montolí ME, Coroleu B, Tresserra F, Barri P, Gómez de Aranda I, Sévigny J, Ponce J, Matias-Guiu X, et al. Impaired Expression of Ectonucleotidases in Ectopic and Eutopic Endometrial Tissue Is in Favor of ATP Accumulation in the Tissue Microenvironment in Endometriosis. International Journal of Molecular Sciences. 2019; 20(22):5532. https://doi.org/10.3390/ijms20225532
Chicago/Turabian StyleTrapero, Carla, August Vidal, Maria Eulàlia Fernández-Montolí, Buenaventura Coroleu, Francesc Tresserra, Pere Barri, Inmaculada Gómez de Aranda, Jean Sévigny, Jordi Ponce, Xavier Matias-Guiu, and et al. 2019. "Impaired Expression of Ectonucleotidases in Ectopic and Eutopic Endometrial Tissue Is in Favor of ATP Accumulation in the Tissue Microenvironment in Endometriosis" International Journal of Molecular Sciences 20, no. 22: 5532. https://doi.org/10.3390/ijms20225532
APA StyleTrapero, C., Vidal, A., Fernández-Montolí, M. E., Coroleu, B., Tresserra, F., Barri, P., Gómez de Aranda, I., Sévigny, J., Ponce, J., Matias-Guiu, X., & Martín-Satué, M. (2019). Impaired Expression of Ectonucleotidases in Ectopic and Eutopic Endometrial Tissue Is in Favor of ATP Accumulation in the Tissue Microenvironment in Endometriosis. International Journal of Molecular Sciences, 20(22), 5532. https://doi.org/10.3390/ijms20225532