Withania Somnifera (Ashwagandha) and Withaferin A: Potential in Integrative Oncology




Abstract
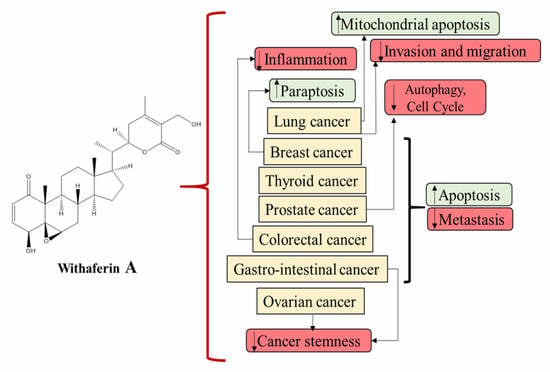
1. Introduction
2. History of WS
3. WS Extracts and their Anticancer Activity
4. Active Components in WS Extract
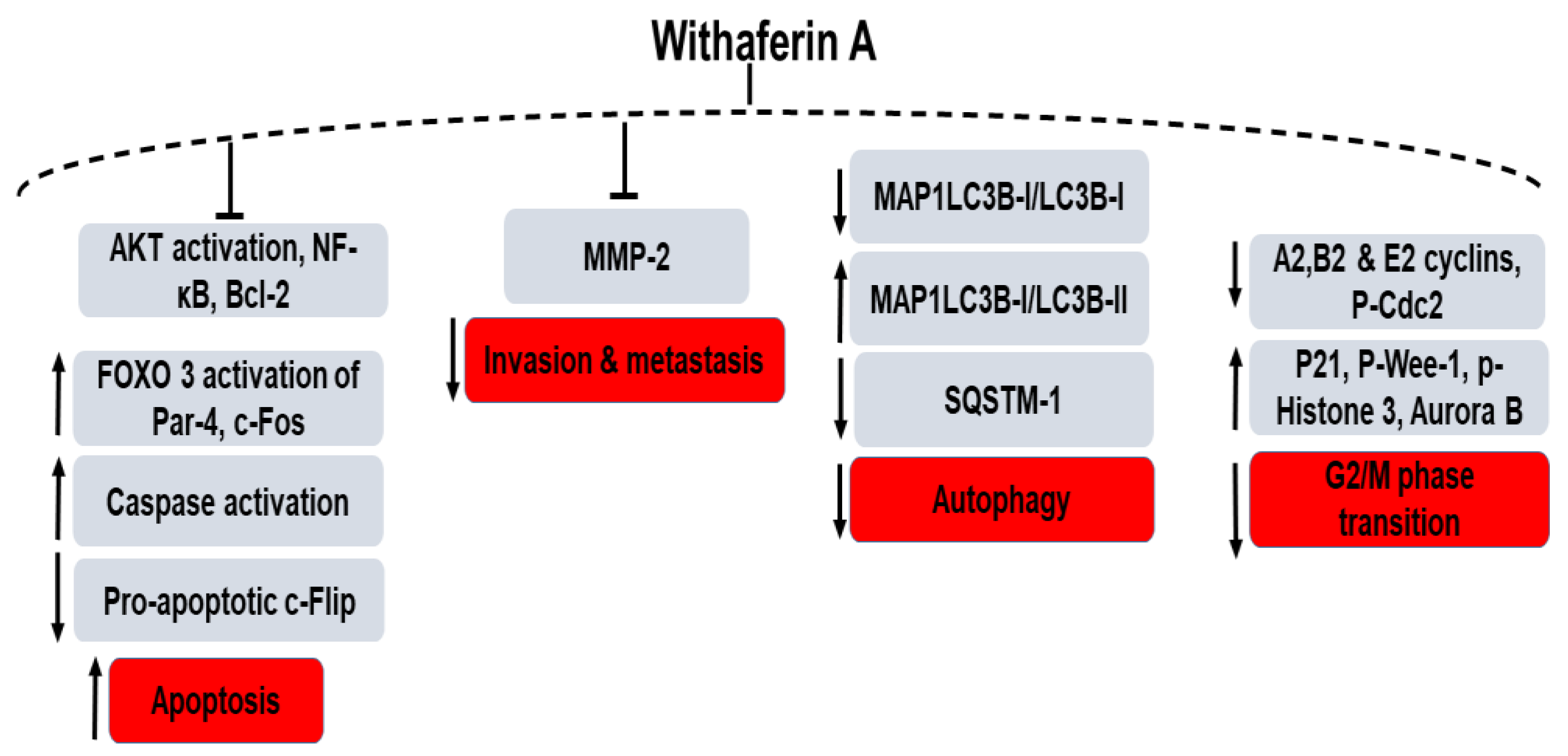
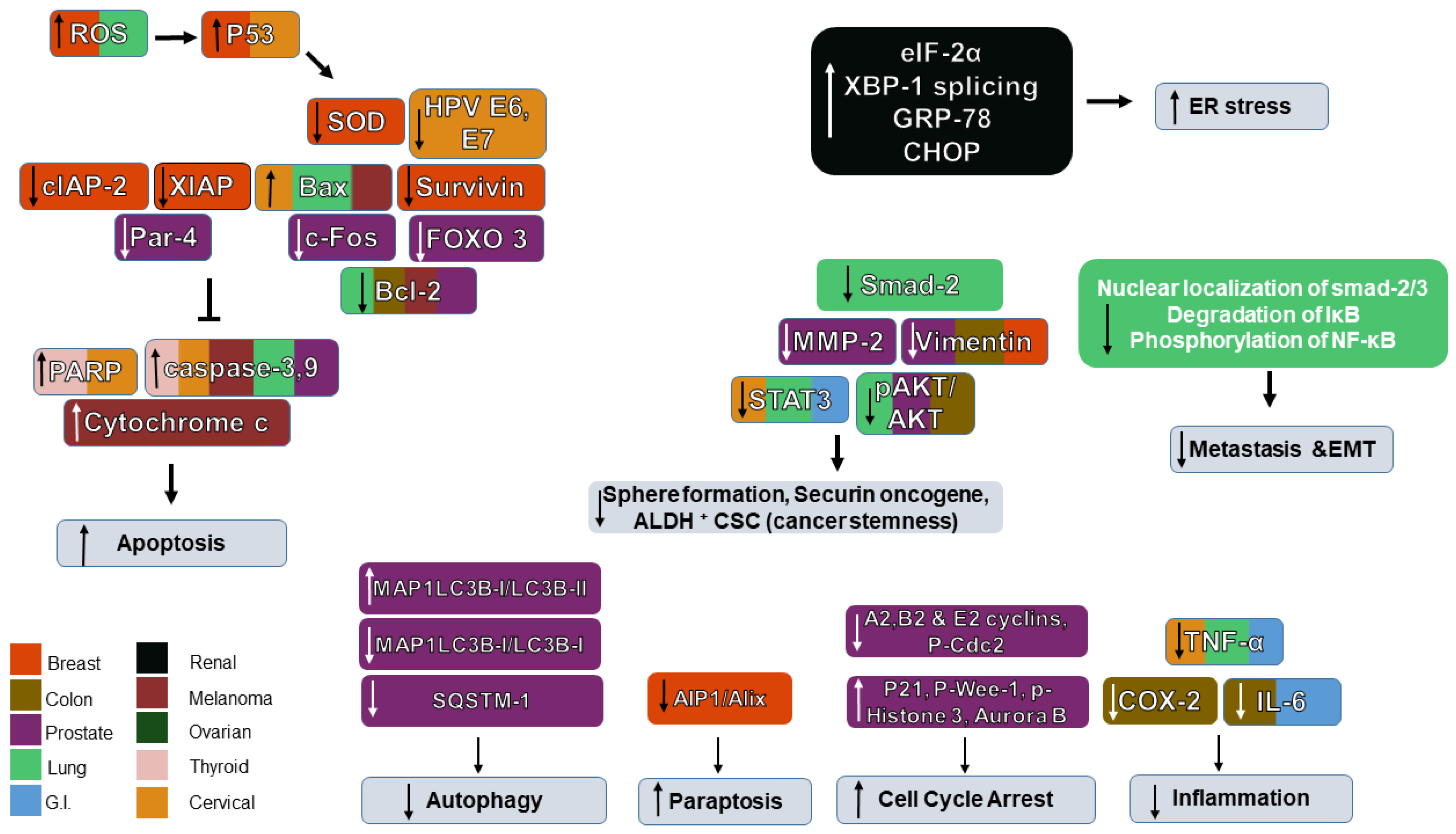
5. Role of WS and WFA in Various Cancers
5.1. Breast Cancer
5.2. Colorectal Cancer
5.3. Prostate Cancer
5.4. Lung Cancer
5.5. Ovarian Cancer
5.6. Other Cancers
6. WS and Cancer Chemotherapy–Induced Toxicities.
7. Concluding Remarks
Funding
Acknowledgments
Conflicts of Interest
References
- Farooqui, A.A.; Farooqui, T.; Madan, A.; Ong, J.H.; Ong, W.Y. Ayurvedic Medicine for the Treatment of Dementia: Mechanistic Aspects. Evid. -Based Complement. Altern. Med. 2018, 2018, 2481076. [Google Scholar] [CrossRef]
- Pratte, M.A.; Nanavati, K.B.; Young, V.; Morley, C.P. An alternative treatment for anxiety: A systematic review of human trial results reported for the Ayurvedic herb ashwagandha (Withania somnifera). J. Altern. Complement. Med. 2014, 20, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.H.; Narsimhamurthy, K.; Singh, G. Neuronutrient impact of Ayurvedic Rasayana therapy in brain aging. Biogerontology 2008, 9, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Rege, N.N.; Thatte, U.M.; Dahanukar, S.A. Adaptogenic properties of six rasayana herbs used in Ayurvedic medicine. Phytother. Res. 1999, 13, 275–291. [Google Scholar] [CrossRef]
- Montalvan, V.; Gallo, M.; Rojas, E. A 25 years-old woman with a postvaccine thalamic pseudotumoral lesion. Rev. Clin. Esp. 2015, 215, 468–472. [Google Scholar]
- Mirjalili, M.H.; Moyano, E.; Bonfill, M.; Cusido, R.M.; Palazon, J. Steroidal lactones from Withania somnifera, an ancient plant for novel medicine. Molecules 2009, 14, 2373–2393. [Google Scholar] [CrossRef]
- Rai, M.; Jogee, P.S.; Agarkar, G.; Dos Santos, C.A. Anticancer activities of Withania somnifera: Current research, formulations, and future perspectives. Pharm. Biol. 2016, 54, 189–197. [Google Scholar] [CrossRef]
- Subbaraju, G.V.; Vanisree, M.; Rao, C.V.; Sivaramakrishna, C.; Sridhar, P.; Jayaprakasam, B.; Nair, M.G. Ashwagandhanolide, a bioactive dimeric thiowithanolide isolated from the roots of Withania somnifera. J. Nat. Prod. 2006, 69, 1790–1792. [Google Scholar] [CrossRef]
- Gannon, J.M.; Brar, J.; Rai, A.; Chengappa, K.N.R. Effects of a standardized extract of Withania somnifera (Ashwagandha) on depression and anxiety symptoms in persons with schizophrenia participating in a randomized, placebo-controlled clinical trial. Ann. Clin. Psychiatry 2019, 31, 123–129. [Google Scholar]
- Ziegenfuss, T.N.; Kedia, A.W.; Sandrock, J.E.; Raub, B.J.; Kerksick, C.M.; Lopez, H.L. Effects of an Aqueous Extract of Withania somnifera on Strength Training Adaptations and Recovery: The STAR Trial. Nutrients 2018, 10, 1807. [Google Scholar] [CrossRef]
- Nasimi Doost Azgomi, R.; Nazemiyeh, H.; Sadeghi Bazargani, H.; Fazljou, S.M.B.; Nejatbakhsh, F.; Moini Jazani, A.; Ahmadi Asrbadr, Y.; Zomorrodi, A. Comparative evaluation of the effects of Withania somnifera with pentoxifylline on the sperm parameters in idiopathic male infertility: A triple-blind randomised clinical trial. Andrologia 2018, 50, e13041. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.K.; Basu, I.; Singh, S. Efficacy and Safety of Ashwagandha Root Extract in Subclinical Hypothyroid Patients: A Double-Blind, Randomized Placebo-Controlled Trial. J. Altern. Complement. Med. 2018, 24, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, D.; Bhattacharyya, S.; Bose, S. Efficacy and Safety of Ashwagandha (Withania somnifera (L.) Dunal) Root Extract in Improving Memory and Cognitive Functions. J. Diet. Suppl. 2017, 14, 599–612. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, D.; Bhattacharyya, S.; Joshi, K. Body Weight Management in Adults Under Chronic Stress Through Treatment With Ashwagandha Root Extract: A Double-Blind, Randomized, Placebo-Controlled Trial. J. Evid. -Based Complement. Altern. Med. 2017, 22, 96–106. [Google Scholar] [CrossRef]
- Devi, P.U.; Sharada, A.C.; Solomon, F.E.; Kamath, M.S. In vivo growth inhibitory effect of Withania somnifera (Ashwagandha) on a transplantable mouse tumor, Sarcoma 180. Indian J. Exp. Biol. 1992, 30, 169–172. [Google Scholar]
- Vanden Berghe, W.; Sabbe, L.; Kaileh, M.; Haegeman, G.; Heyninck, K. Molecular insight in the multifunctional activities of Withaferin A. Biochem. Pharmacol. 2012, 84, 1282–1291. [Google Scholar] [CrossRef]
- Scartezzini, P.; Speroni, E. Review on some plants of Indian traditional medicine with antioxidant activity. J. Ethnopharmacol. 2000, 71, 23–43. [Google Scholar] [CrossRef]
- Devi, P.U. Withania somnifera Dunal (Ashwagandha): Potential plant source of a promising drug for cancer chemotherapy and radiosensitization. Indian J. Exp. Biol. 1996, 34, 927–932. [Google Scholar]
- Kakar, S.S.; Ratajczak, M.Z.; Powell, K.S.; Moghadamfalahi, M.; Miller, D.M.; Batra, S.K.; Singh, S.K. Withaferin a alone and in combination with cisplatin suppresses growth and metastasis of ovarian cancer by targeting putative cancer stem cells. PLoS ONE 2014, 9, e107596. [Google Scholar] [CrossRef]
- Mondal, A.; Gandhi, A.; Fimognari, C.; Atanasov, A.G.; Bishayee, A. Alkaloids for cancer prevention and therapy: Current progress and future perspectives. Eur. J. Pharmacol. 2019, 858, 172472. [Google Scholar] [CrossRef]
- Kurup, P.A. The antibacterial principle of Withania somnifera. I. Isoation and antibacterial activity. Antibiot. Chemother. 1958, 8, 511–515. [Google Scholar]
- Malhotra, C.L.; Das, P.K.; Dhalla, N.S. Studies on Withania ashwagandha. (Part I): Effect of total extract on central nervous system and smooth muscles. Indian J. Physiol. Pharmacol. 1960, 4, 35–48. [Google Scholar] [PubMed]
- Malhotra, C.L.; Das, P.K.; Dhalla, N.S. Studies on Withania ashwagandha. (Part II): Effect of total extract on cardiovascular system, respiration and skeletal muscle. Indian J. Physiol. Pharmacol. 1960, 4, 49–64. [Google Scholar] [PubMed]
- Kohlmuenzer, S.; Krupinska, J. Chemotherapeutic properties of substances isolated from leaves of Withania somnifera Dunal. Acta Physiol. Pol. 1960, 11, 778–780. [Google Scholar]
- Dhalla, N.S.; Sastry, M.S.; Malhotra, C.L. Chemical studies of the leaves of Withania somnifera. J. Pharm. Sci. 1961, 50, 876–877. [Google Scholar] [CrossRef]
- Menssen, H.G.; Stapel, G. A C28-steroidlacton from the roots of Withania somnifera (author’s transl). Planta Med. 1973, 24, 8–12. [Google Scholar]
- Chakraborti, S.K.; De, B.K.; Bandyopadhyay, T. Variations in the antitumour constituents of withania somnifera dunal. Experientia 1974, 30, 852–853. [Google Scholar] [CrossRef]
- Devi, P.U.; Sharada, A.C.; Solomon, F.E. Antitumor and radiosensitizing effects of Withania somnifera (Ashwagandha) on a transplantable mouse tumor, Sarcoma-180. Indian J. Exp. Biol. 1993, 31, 607–611. [Google Scholar]
- Devi, P.U.; Akagi, K.; Ostapenko, V.; Tanaka, Y.; Sugahara, T. Withaferin A: A new radiosensitizer from the Indian medicinal plant Withania somnifera. Int. J. Radiat. Biol. 1996, 69, 193–197. [Google Scholar] [CrossRef]
- Kuttan, G. Use of Withania somnifera Dunal as an adjuvant during radiation therapy. Indian J. Exp. Biol. 1996, 34, 854–856. [Google Scholar]
- Mulabagal, V.; Subbaraju, G.V.; Rao, C.V.; Sivaramakrishna, C.; Dewitt, D.L.; Holmes, D.; Sung, B.; Aggarwal, B.B.; Tsay, H.S.; Nair, M.G. Withanolide sulfoxide from Aswagandha roots inhibits nuclear transcription factor-kappa-B, cyclooxygenase and tumor cell proliferation. Phytother. Res. 2009, 23, 987–992. [Google Scholar] [CrossRef] [PubMed]
- Caputi, F.F.; Acquas, E.; Kasture, S.; Ruiu, S.; Candeletti, S.; Romualdi, P. The standardized Withania somnifera Dunal root extract alters basal and morphine-induced opioid receptor gene expression changes in neuroblastoma cells. BMC Complement. Altern. Med. 2018, 18, 9. [Google Scholar] [CrossRef] [PubMed]
- Henley, A.B.; Yang, L.; Chuang, K.L.; Sahuri-Arisoylu, M.; Wu, L.H.; Bligh, S.W.; Bell, J.D. Withania somnifera Root Extract Enhances Chemotherapy through ‘Priming’. PLoS ONE 2017, 12, e0170917. [Google Scholar] [CrossRef] [PubMed]
- Sudeep, H.V.; Gouthamchandra, K.; Venkatesh, B.J.; Prasad, K.S. Viwithan, a Standardized Withania somnifera Root Extract Induces Apoptosis in Murine Melanoma Cells. Pharmacogn. Mag. 2018, 13, S801–S806. [Google Scholar]
- Khazal, K.F.; Hill, D.L.; Grubbs, C.J. Effect of Withania somnifera root extract on spontaneous estrogen receptor-negative mammary cancer in MMTV/Neu mice. Anticancer Res. 2014, 34, 6327–6332. [Google Scholar]
- Turrini, E.; Calcabrini, C.; Sestili, P.; Catanzaro, E.; De Gianni, E.; Diaz, A.R.; Hrelia, P.; Tacchini, M.; Guerrini, A.; Canonico, B.; et al. Withania somnifera Induces Cytotoxic and Cytostatic Effects on Human T Leukemia Cells. Toxins (Basel) 2016, 8, 147. [Google Scholar] [CrossRef]
- Devi, P.U.; Sharada, A.C.; Solomon, F.E. In vivo growth inhibitory and radiosensitizing effects of withaferin A on mouse Ehrlich ascites carcinoma. Cancer Lett. 1995, 95, 189–193. [Google Scholar] [CrossRef]
- Mohan, R.; Hammers, H.J.; Bargagna-Mohan, P.; Zhan, X.H.; Herbstritt, C.J.; Ruiz, A.; Zhang, L.; Hanson, A.D.; Conner, B.P.; Rougas, J.; et al. Withaferin A is a potent inhibitor of angiogenesis. Angiogenesis 2004, 7, 115–122. [Google Scholar] [CrossRef]
- Halder, B.; Singh, S.; Thakur, S.S. Withania somnifera Root Extract Has Potent Cytotoxic Effect against Human Malignant Melanoma Cells. PLoS ONE 2015, 10, e0137498. [Google Scholar]
- Dar, P.A.; Mir, S.A.; Bhat, J.A.; Hamid, A.; Singh, L.R.; Malik, F.; Dar, T.A. An anti-cancerous protein fraction from Withania somnifera induces ROS-dependent mitochondria-mediated apoptosis in human MDA-MB-231 breast cancer cells. Int. J. Biol. Macromol. 2019, 135, 77–87. [Google Scholar] [CrossRef]
- Naidoo, D.B.; Chuturgoon, A.A.; Phulukdaree, A.; Guruprasad, K.P.; Satyamoorthy, K.; Sewram, V. Withania somnifera modulates cancer cachexia associated inflammatory cytokines and cell death in leukaemic THP-1 cells and peripheral blood mononuclear cells (PBMC’s). BMC Complement. Altern. Med. 2018, 18, 126. [Google Scholar] [CrossRef] [PubMed]
- Kaileh, M.; Vanden Berghe, W.; Heyerick, A.; Horion, J.; Piette, J.; Libert, C.; De Keukeleire, D.; Essawi, T.; Haegeman, G. Withaferin a strongly elicits IkappaB kinase beta hyperphosphorylation concomitant with potent inhibition of its kinase activity. J. Biol. Chem. 2007, 282, 4253–4264. [Google Scholar] [CrossRef] [PubMed]
- Alfaifi, M.Y.; Saleh, K.A.; El-Boushnak, M.A.; Elbehairi, S.E.; Alshehri, M.A.; Shati, A.A. Antiproliferative Activity of the Methanolic Extract of Withania Somnifera Leaves from Faifa Mountains, Southwest Saudi Arabia, against Several Human Cancer Cell Lines. Asian Pac. J. Cancer Prev. 2016, 17, 2723–2726. [Google Scholar] [PubMed]
- Shah, N.; Kataria, H.; Kaul, S.C.; Ishii, T.; Kaur, G.; Wadhwa, R. Effect of the alcoholic extract of Ashwagandha leaves and its components on proliferation, migration, and differentiation of glioblastoma cells: Combinational approach for enhanced differentiation. Cancer Sci. 2009, 100, 1740–1747. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, W.; Mofed, D.; Zekri, A.R.; El-Sayed, N.; Rahouma, M.; Sabet, S. Antioxidant activity and apoptotic induction as mechanisms of action of Withania somnifera (Ashwagandha) against a hepatocellular carcinoma cell line. J. Int. Med. Res. 2018, 46, 1358–1369. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.; Kaur, G. Aqueous extract from the Withania somnifera leaves as a potential anti-neuroinflammatory agent: A mechanistic study. J. Neuroinflamm. 2016, 13, 193. [Google Scholar] [CrossRef]
- Wadhwa, R.; Singh, R.; Gao, R.; Shah, N.; Widodo, N.; Nakamoto, T.; Ishida, Y.; Terao, K.; Kaul, S.C. Water extract of Ashwagandha leaves has anticancer activity: Identification of an active component and its mechanism of action. PLoS ONE 2013, 8, e77189. [Google Scholar] [CrossRef]
- Kataria, H.; Kumar, S.; Chaudhary, H.; Kaur, G. Withania somnifera Suppresses Tumor Growth of Intracranial Allograft of Glioma Cells. Mol. Neurobiol. 2016, 53, 4143–4158. [Google Scholar] [CrossRef]
- Shah, N.; Singh, R.; Sarangi, U.; Saxena, N.; Chaudhary, A.; Kaur, G.; Kaul, S.C.; Wadhwa, R. Combinations of Ashwagandha leaf extracts protect brain-derived cells against oxidative stress and induce differentiation. PLoS ONE 2015, 10, e0120554. [Google Scholar] [CrossRef]
- Srivastava, A.N.; Ahmad, R.; Khan, M.A. Evaluation and Comparison of the In Vitro Cytotoxic Activity of Withania somnifera Methanolic and Ethanolic Extracts against MDA-MB-231 and Vero Cell Lines. Sci. Pharm. 2016, 84, 41–59. [Google Scholar] [CrossRef]
- Maliyakkal, N.; Appadath Beeran, A.; Balaji, S.A.; Udupa, N.; Ranganath Pai, S.; Rangarajan, A. Effects of Withania somnifera and Tinospora cordifolia extracts on the side population phenotype of human epithelial cancer cells: Toward targeting multidrug resistance in cancer. Integr. Cancer Ther. 2015, 14, 156–171. [Google Scholar] [CrossRef] [PubMed]
- Vareed, S.K.; Bauer, A.K.; Nair, K.M.; Liu, Y.; Jayaprakasam, B.; Nair, M.G. Blood-brain barrier permeability of bioactive withanamides present in Withania somnifera fruit extract. Phytother. Res. 2014, 28, 1260–1264. [Google Scholar] [CrossRef] [PubMed]
- Abutaha, N. In vitro antiproliferative activity of partially purified Withania somnifera fruit extract on different cancer cell lines. J. BUON 2015, 20, 625–630. [Google Scholar] [PubMed]
- Tao, S.; Tillotson, J.; Wijeratne, E.M.K.; Xu, Y.M.; Kang, M.; Wu, T.; Lau, E.C.; Mesa, C.; Mason, D.J.; Brown, R.V.; et al. Withaferin A Analogs That Target the AAA+ Chaperone p97. ACS Chem. Biol. 2015, 10, 1916–1924. [Google Scholar] [CrossRef] [PubMed]
- Motiwala, H.F.; Bazzill, J.; Samadi, A.; Zhang, H.; Timmermann, B.N.; Cohen, M.S.; Aube, J. Synthesis and Cytotoxicity of Semisynthetic Withalongolide A Analogues. ACS Med. Chem. Lett. 2013, 4, 1069–1073. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Dabrosin, C.; Yin, X.; Fuster, M.M.; Arreola, A.; Rathmell, W.K.; Generali, D.; Nagaraju, G.P.; El-Rayes, B.; Ribatti, D.; et al. Broad targeting of angiogenesis for cancer prevention and therapy. Semin. Cancer Biol. 2015, 35, S224–S243. [Google Scholar] [CrossRef]
- Srinivasan, S.; Ranga, R.S.; Burikhanov, R.; Han, S.S.; Chendil, D. Par-4-dependent apoptosis by the dietary compound withaferin A in prostate cancer cells. Cancer Res. 2007, 67, 246–253. [Google Scholar] [CrossRef]
- Malik, F.; Kumar, A.; Bhushan, S.; Khan, S.; Bhatia, A.; Suri, K.A.; Qazi, G.N.; Singh, J. Reactive oxygen species generation and mitochondrial dysfunction in the apoptotic cell death of human myeloid leukemia HL-60 cells by a dietary compound withaferin A with concomitant protection by N-acetyl cysteine. Apoptosis 2007, 12, 2115–2133. [Google Scholar] [CrossRef]
- Stan, S.D.; Hahm, E.R.; Warin, R.; Singh, S.V. Withaferin A causes FOXO3a- and Bim-dependent apoptosis and inhibits growth of human breast cancer cells in vivo. Cancer Res. 2008, 68, 7661–7669. [Google Scholar] [CrossRef]
- Mandal, C.; Dutta, A.; Mallick, A.; Chandra, S.; Misra, L.; Sangwan, R.S.; Mandal, C. Withaferin A induces apoptosis by activating p38 mitogen-activated protein kinase signaling cascade in leukemic cells of lymphoid and myeloid origin through mitochondrial death cascade. Apoptosis 2008, 13, 1450–1464. [Google Scholar] [CrossRef]
- Li, X.; Zhu, F.; Jiang, J.; Sun, C.; Zhong, Q.; Shen, M.; Wang, X.; Tian, R.; Shi, C.; Xu, M.; et al. Simultaneous inhibition of the ubiquitin-proteasome system and autophagy enhances apoptosis induced by ER stress aggravators in human pancreatic cancer cells. Autophagy 2016, 12, 1521–1537. [Google Scholar] [CrossRef] [PubMed]
- Chang, E.; Pohling, C.; Natarajan, A.; Witney, T.H.; Kaur, J.; Xu, L.; Gowrishankar, G.; D’souza, A.L.; Murty, S.; Schick, S.; et al. AshwaMAX and Withaferin A inhibits gliomas in cellular and murine orthotopic models. J. Neuro. Oncol. 2016, 126, 253–264. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Miao, Y.; Liu, S. Withaferin A induces apoptosis by ROS-dependent mitochondrial dysfunction in human colorectal cancer cells. Biochem. Biophys. Res. Commun. 2018, 503, 2363–2369. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Li, T.; Bai, Z.; Yang, Y.; Liu, X.; Zhan, J.; Shi, B. Breast cancer intrinsic subtype classification, clinical use and future trends. Am. J. Cancer Res. 2015, 5, 2929–2943. [Google Scholar]
- Samanta, S.K.; Sehrawat, A.; Kim, S.H.; Hahm, E.R.; Shuai, Y.; Roy, R.; Pore, S.K.; Singh, K.B.; Christner, S.M.; Beumer, J.H.; Davidson, N.E.; Singh, S.V. Disease Subtype-Independent Biomarkers of Breast Cancer Chemoprevention by the Ayurvedic Medicine Phytochemical Withaferin A. J. Natl. Cancer Inst. 2017, 109, 6. [Google Scholar] [CrossRef]
- Hahm, E.R.; Lee, J.; Huang, Y.; Singh, S.V. Withaferin a suppresses estrogen receptor-alpha expression in human breast cancer cells. Mol. Carcinog. 2011, 50, 614–624. [Google Scholar] [CrossRef]
- Szarc Vel Szic, K.; Op De Beeck, K.; Ratman, D.; Wouters, A.; Beck, I.M.; Declerck, K.; Heyninck, K.; Fransen, E.; Bracke, M.; De Bosscher, K.; et al. Pharmacological levels of Withaferin A (Withania somnifera) trigger clinically relevant anticancer effects specific to triple negative breast cancer cells. PLoS One 2014, 9, e87850. [Google Scholar] [CrossRef]
- Szarc Vel Szic, K.; Declerck, K.; Crans, R.A.J.; Diddens, J.; Scherf, D.B.; Gerhauser, C.; Vanden Berghe, W. Epigenetic silencing of triple negative breast cancer hallmarks by Withaferin A. Oncotarget 2017, 8, 40434–40453. [Google Scholar] [CrossRef] [PubMed]
- Mohan, R.; Bargagna-Mohan, P. The Use of Withaferin A to Study Intermediate Filaments. Methods Enzymol. 2016, 568, 187–218. [Google Scholar]
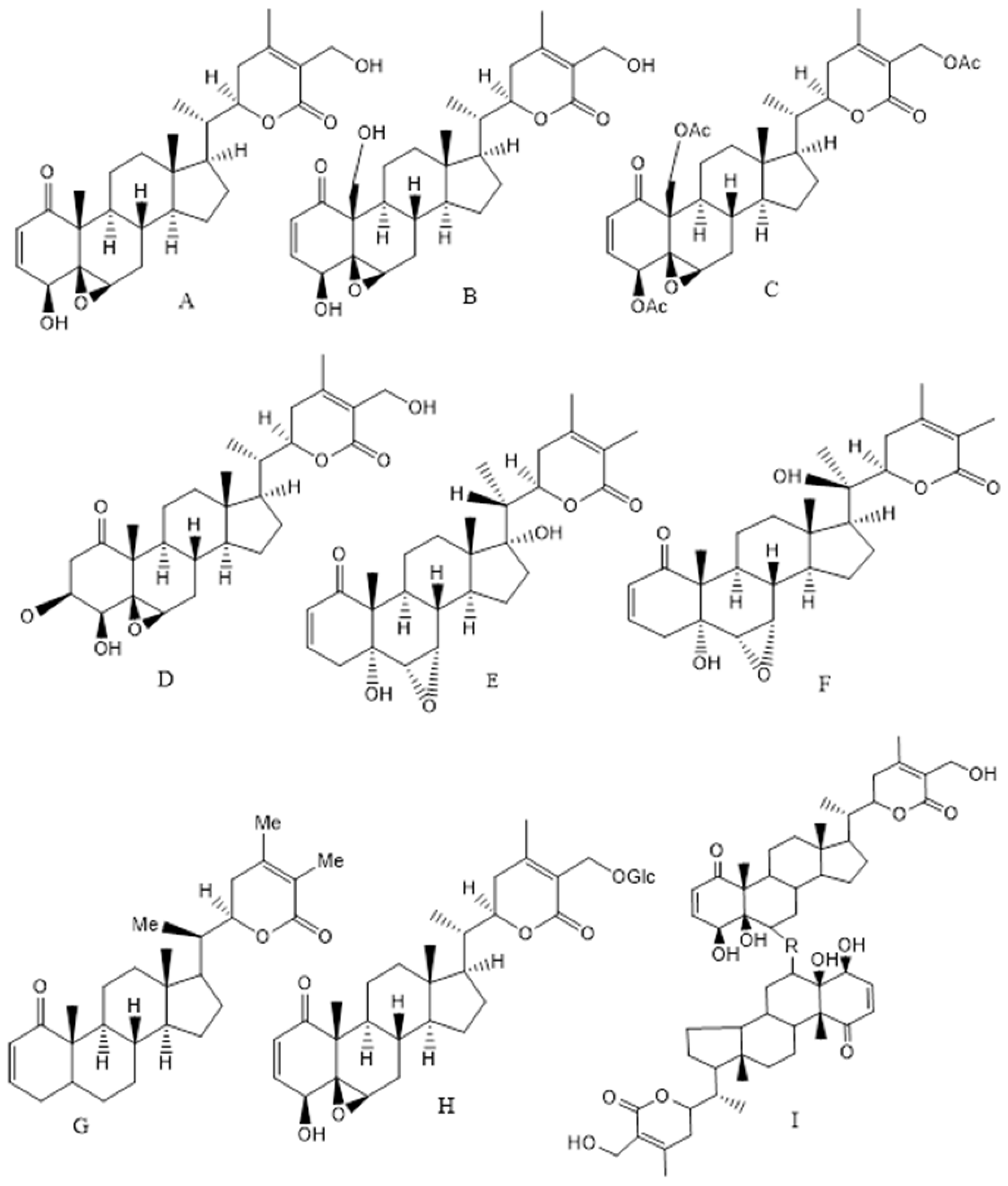
- Yang, Z.; Garcia, A.; Xu, S.; Powell, D.R.; Vertino, P.M.; Singh, S.; Marcus, A.I. Withania somnifera root extract inhibits mammary cancer metastasis and epithelial to mesenchymal transition. PLoS ONE 2013, 8, e75069. [Google Scholar] [CrossRef]
- Widodo, N.; Kaur, K.; Shrestha, B.G.; Takagi, Y.; Ishii, T.; Wadhwa, R.; Kaul, S.C. Selective killing of cancer cells by leaf extract of Ashwagandha: Identification of a tumor-inhibitory factor and the first molecular insights to its effect. Clin. Cancer Res. 2007, 13, 2298–2306. [Google Scholar] [CrossRef] [PubMed]
- Hahm, E.R.; Moura, M.B.; Kelley, E.E.; Van Houten, B.; Shiva, S.; Singh, S.V. Withaferin A-induced apoptosis in human breast cancer cells is mediated by reactive oxygen species. PLoS ONE 2011, 6, e23354. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, K.; De, S.; Das, S.; Mukherjee, S.; Sengupta Bandyopadhyay, S. Withaferin A Induces ROS-Mediated Paraptosis in Human Breast Cancer Cell-Lines MCF-7 and MDA-MB-231. PLoS ONE 2016, 11, e0168488. [Google Scholar] [CrossRef] [PubMed]
- Widodo, N.; Priyandoko, D.; Shah, N.; Wadhwa, R.; Kaul, S.C. Selective killing of cancer cells by Ashwagandha leaf extract and its component Withanone involves ROS signaling. PLoS ONE 2010, 5, e13536. [Google Scholar] [CrossRef]
- Sehrawat, A.; Samanta, S.K.; Hahm, E.R.; St Croix, C.; Watkins, S.; Singh, S.V. Withaferin A-mediated apoptosis in breast cancer cells is associated with alterations in mitochondrial dynamics. Mitochondrion 2019. [Google Scholar] [CrossRef]
- Hahm, E.R.; Singh, S.V. Withaferin A-induced apoptosis in human breast cancer cells is associated with suppression of inhibitor of apoptosis family protein expression. Cancer Lett. 2013, 334, 101–108. [Google Scholar] [CrossRef]
- Sameer, A.S. Colorectal cancer: molecular mutations and polymorphisms. Front. Oncol. 2013, 3, 114. [Google Scholar] [CrossRef]
- Choi, B.Y.; Kim, B.W. Withaferin-A Inhibits Colon Cancer Cell Growth by Blocking STAT3 Transcriptional Activity. J. Cancer Prev. 2015, 20, 185–192. [Google Scholar] [CrossRef]
- Koduru, S.; Kumar, R.; Srinivasan, S.; Evers, M.B.; Damodaran, C. Notch-1 inhibition by Withaferin-A: A therapeutic target against colon carcinogenesis. Mol. Cancer Ther. 2010, 9, 202–210. [Google Scholar] [CrossRef]
- Chandrasekaran, B.; Pal, D.; Kolluru, V.; Tyagi, A.; Baby, B.; Dahiya, N.R.; Khafateh, Y.; Houda, A.; Ankem, M.K.; Sharma, A.K.; et al. The Chemopreventive effect of Withaferin A on spontaneous and Inflammation-associated colon carcinogenesis models. Carcinogenesis 2018. [Google Scholar] [CrossRef]
- Roy, R.V.; Suman, S.; Das, T.P.; Luevano, J.E.; Damodaran, C. Withaferin A, a steroidal lactone from Withania somnifera, induces mitotic catastrophe and growth arrest in prostate cancer cells. J. Nat. Prod. 2013, 76, 1909–1915. [Google Scholar] [CrossRef] [PubMed]
- Suman, S.; Das, T.P.; Moselhy, J.; Pal, D.; Kolluru, V.; Alatassi, H.; Ankem, M.K.; Damodaran, C. Oral administration of withaferin A inhibits carcinogenesis of prostate in TRAMP model. Oncotarget 2016, 7, 53751–53761. [Google Scholar] [CrossRef] [PubMed]
- Moselhy, J.; Suman, S.; Alghamdi, M.; Chandarasekharan, B.; Das, T.P.; Houda, A.; Ankem, M.; Damodaran, C. Withaferin A Inhibits Prostate Carcinogenesis in a PTEN-deficient Mouse Model of Prostate Cancer. Neoplasia 2017, 19, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Das, T.P.; Suman, S.; Alatassi, H.; Ankem, M.K.; Damodaran, C. Inhibition of AKT promotes FOXO3a-dependent apoptosis in prostate cancer. Cell Death Dis. 2016, 7, e2111. [Google Scholar] [CrossRef]
- Rah, B.; Ur Rasool, R.; Nayak, D.; Yousuf, S.K.; Mukherjee, D.; Kumar, L.D.; Goswami, A. PAWR-mediated suppression of BCL2 promotes switching of 3-azido withaferin A (3-AWA)-induced autophagy to apoptosis in prostate cancer cells. Autophagy 2015, 11, 314–331. [Google Scholar] [CrossRef]
- Rah, B.; Amin, H.; Yousuf, K.; Khan, S.; Jamwal, G.; Mukherjee, D.; Goswami, A. A novel MMP-2 inhibitor 3-azidowithaferin A (3-azidoWA) abrogates cancer cell invasion and angiogenesis by modulating extracellular Par-4. PLoS ONE 2012, 7, e44039. [Google Scholar] [CrossRef]
- Nishikawa, Y.; Okuzaki, D.; Fukushima, K.; Mukai, S.; Ohno, S.; Ozaki, Y.; Yabuta, N.; Nojima, H. Withaferin A Induces Cell Death Selectively in Androgen-Independent Prostate Cancer Cells but Not in Normal Fibroblast Cells. PLoS ONE 2015, 10, e0134137. [Google Scholar] [CrossRef]
- Coudray, N.; Ocampo, P.S.; Sakellaropoulos, T.; Narula, N.; Snuderl, M.; Fenyo, D.; Moreira, A.L.; Razavian, N.; Tsirigos, A. Classification and mutation prediction from non-small cell lung cancer histopathology images using deep learning. Nat. Med. 2018, 24, 1559–1567. [Google Scholar] [CrossRef]
- Hsu, J.H.; Chang, P.M.; Cheng, T.S.; Kuo, Y.L.; Wu, A.T.; Tran, T.H.; Yang, Y.H.; Chen, J.M.; Tsai, Y.C.; Chu, Y.S.; Huang, T.H.; Huang, C.F.; Lai, J.M. Identification of Withaferin A as a Potential Candidate for Anti-Cancer Therapy in Non-Small Cell Lung Cancer. Cancers (Basel) 2019, 11, 1003. [Google Scholar] [CrossRef]
- Nakano, T.; Kanai, Y.; Amano, Y.; Yoshimoto, T.; Matsubara, D.; Shibano, T.; Tamura, T.; Oguni, S.; Katashiba, S.; Ito, T.; Murakami, Y.; Fukayama, M.; Murakami, T.; Endo, S.; Niki, T. Establishment of highly metastatic KRAS mutant lung cancer cell sublines in long-term three-dimensional low attachment cultures. PLoS ONE 2017, 12, e0181342. [Google Scholar] [CrossRef]
- Cai, Y.; Sheng, Z.Y.; Chen, Y.; Bai, C. Effect of Withaferin A on A549 cellular proliferation and apoptosis in non-small cell lung cancer. Asian Pac. J. Cancer Prev. 2014, 15, 1711–1714. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, L.; Liang, T.; Tian, X.D.; Liu, Y.; Zhang, T. Withaferin A induces mitochondrial-dependent apoptosis in non-small cell lung cancer cells via generation of reactive oxygen species. J. BUON 2017, 22, 244–250. [Google Scholar] [PubMed]
- Kyakulaga, A.H.; Aqil, F.; Munagala, R.; Gupta, R.C. Withaferin A inhibits Epithelial to Mesenchymal Transition in Non-Small Cell Lung Cancer Cells. Sci. Rep. 2018, 8, 15737. [Google Scholar] [CrossRef] [PubMed]
- Kunimasa, K.; Nagano, T.; Shimono, Y.; Dokuni, R.; Kiriu, T.; Tokunaga, S.; Tamura, D.; Yamamoto, M.; Tachihara, M.; Kobayashi, K.; et al. Glucose metabolism-targeted therapy and withaferin A are effective for epidermal growth factor receptor tyrosine kinase inhibitor-induced drug-tolerant persisters. Cancer Sci. 2017, 108, 1368–1377. [Google Scholar] [CrossRef]
- Kakar, S.S.; Worth, C.A.; Wang, Z.; Carter, K.; Ratajczak, M.; Gunjal, P. DOXIL when combined with Withaferin A (WFA) targets ALDH1 positive cancer stem cells in ovarian cancer. J. Cancer Stem Cell Res. 2016, 4. [Google Scholar] [CrossRef]
- Kim, G.; Kim, T.H.; Hwang, E.H.; Chang, K.T.; Hong, J.J.; Park, J.H. Withaferin A inhibits the proliferation of gastric cancer cells by inducing G2/M cell cycle arrest and apoptosis. Oncol. Lett. 2017, 14, 416–422. [Google Scholar] [CrossRef]
- Ting, L.-L.; Chou, A.S.-B.; Hsieh, C.-H.; Hsiung, S.-C.; Pang, S.-T.; Liao, S.-K. Withaferin A targeting both cancer stem cells and metastatic cancer stem cells in the UP-LN1 carcinoma cell model. J. Cancer Metastasis Treat. 2016, 2, 29–40. [Google Scholar]
- Cohen, S.M.; Mukerji, R.; Timmermann, B.N.; Samadi, A.K.; Cohen, M.S. A novel combination of withaferin A and sorafenib shows synergistic efficacy against both papillary and anaplastic thyroid cancers. Am. J. Surg. 2012, 204, 895–901. [Google Scholar] [CrossRef]
- Munagala, R.; Kausar, H.; Munjal, C.; Gupta, R.C. Withaferin A induces p53-dependent apoptosis by repression of HPV oncogenes and upregulation of tumor suppressor proteins in human cervical cancer cells. Carcinogenesis 2011, 32, 1697–1705. [Google Scholar] [CrossRef]
- Mayola, E.; Gallerne, C.; Esposti, D.D.; Martel, C.; Pervaiz, S.; Larue, L.; Debuire, B.; Lemoine, A.; Brenner, C.; Lemaire, C. Withaferin A induces apoptosis in human melanoma cells through generation of reactive oxygen species and down-regulation of Bcl-2. Apoptosis 2011, 16, 1014–1027. [Google Scholar] [CrossRef]
- Choi, M.J.; Park, E.J.; Min, K.J.; Park, J.W.; Kwon, T.K. Endoplasmic reticulum stress mediates withaferin A-induced apoptosis in human renal carcinoma cells. Toxicol. In Vitro 2011, 25, 692–698. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Katiyar, S.P.; Sundar, D.; Kaul, Z.; Miyako, E.; Zhang, Z.; Kaul, S.C.; Reddel, R.R.; Wadhwa, R. Withaferin-A kills cancer cells with and without telomerase: Chemical, computational and experimental evidences. Cell Death Dis. 2017, 8, e2755. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Guo, R.; Gan, L.; Lau, W.B.; Cao, X.; Zhao, J.; Ma, X.; Christopher, T.A.; Lopez, B.L.; Wang, Y. Withaferin A inhibits apoptosis via activated Akt-mediated inhibition of oxidative stress. Life Sci. 2018, 211, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Vedi, M.; Sabina, E.P. Assessment of hepatoprotective and nephroprotective potential of withaferin A on bromobenzene-induced injury in Swiss albino mice: Possible involvement of mitochondrial dysfunction and inflammation. Cell Biol. Toxicol. 2016, 32, 373–390. [Google Scholar] [CrossRef] [PubMed]
- Palliyaguru, D.L.; Chartoumpekis, D.V.; Wakabayashi, N.; Skoko, J.J.; Yagishita, Y.; Singh, S.V.; Kensler, T.W. Withaferin A induces Nrf2-dependent protection against liver injury: Role of Keap1-independent mechanisms. Free Radic. Biol. Med. 2016, 101, 116–128. [Google Scholar] [CrossRef] [PubMed]
- Tiruveedi, V.L.; Bale, S.; Khurana, A.; Godugu, C. Withaferin A, a novel compound of Indian ginseng (Withania somnifera), ameliorates Cerulein-induced acute pancreatitis: Possible role of oxidative stress and inflammation. Phytother. Res. 2018. [Google Scholar] [CrossRef]
- Bale, S.; Pulivendala, G.; Godugu, C. Withaferin A attenuates bleomycin-induced scleroderma by targeting FoxO3 and NF-κβ signaling: Connecting fibrosis and inflammation. Biofactors 2018, 44, 507–517. [Google Scholar] [CrossRef]
- Anwar, M.F.; Yadav, D.; Rastogi, S.; Arora, I.; Khar, R.K.; Chander, J.; Samim, M. Modulation of liver and kidney toxicity by herb Withania somnifera for silver nanoparticles: A novel approach for harmonizing between safety and use of nanoparticles. Protoplasma 2015, 252, 547–558. [Google Scholar] [CrossRef]
- Biswal, B.M.; Sulaiman, S.A.; Ismail, H.C.; Zakaria, H.; Musa, K.I. Effect of Withania somnifera (Ashwagandha) on the development of chemotherapy-induced fatigue and quality of life in breast cancer patients. Integr. Cancer Ther. 2013, 12, 312–322. [Google Scholar] [CrossRef]
- World Health Organization. WHO Traditional Medicine Strategy 2014–2023; WHO: Geneva, Switzerland, 2013; Available online: https://www.who.int/medicines/publications/traditional/trm_strategy14_23/en/ (accessed on 1 December 2013).
- Zhang, A.L.; Xue, C.C.; Fong, H.H.S. Integration of herbal medicine into evidence-based clinical practice. In Herbal Medicine: Biomolecular and Clinical Aspects, 2nd ed.; Benzie, I.F.F., Wachtel-Galor, S., Eds.; CRC Press, Taylor Francis: Boca Raton, FL, USA, 2011. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disease, n of Subjects, Dose/Duration | Efficacy | Safety |
|---|---|---|
| Schizophrenia, n = 66, 1000 mg bid, 12 weeks | Medium effect sizes of favoring WS extracts (WSE) over placebo | Adverse events were mild and transient [9] |
| STAR (Strength Training Adaptations and Recovery) Trial, n = 19 (S500 vs. placebo), 500 mg/d, 12 weeks | Improved upper and lower-body strength, and supported a favorable distribution of body mass | Well tolerated clinically [10] |
| Idiopathic male infertility n = 50, 5 g/d, 90 days | Improves sperms parameters in idiopathic male infertility | Without causing adverse effects [11] |
| Subclinical Hypothyroid, n = 25, 600 mg daily | Compared to placebo, WS normalized serum TSH, serum triiodothyronine (T3) and thyroxine (T4) levels significantly in a subclinical double-blinded, randomized placebo-controlled trial. | Well tolerated [12] |
| Mild cognitive impairment (MCI), n = 50, 300 mg bid, 8 weeks | Improvement in overall cognitive function of adult patients with MCI | Effective in WS pre-treatment vs. post treatment [13] |
| Body weight management under chronic stress, n = 52, 300 mg bid, 6 weeks | Significant improvements in both primary and secondary measures | Well tolerated [14] |
| Extractant | Disease | Pathways or as Adjuvants |
|---|---|---|
| Whole Plant | ||
| Methanol (75%) | Increase bone marrow cellularity; stem cell proliferation | Increase in total WBC (white blood cell) count; adjuvant during radiation therapy [30] |
| WS Root Extract | ||
| Methanol (withanolide sulfoxide) | Gastric (AGS), breast (MCF-7), colon (HCT-116) | Suppressed TNF (tumor necrosis factor)-induced NF-kappa B activation [31] |
| Methanol | Neuroblastoma cells | alters basal and morphine-induced opioid receptor gene expression [32] |
| Methanol | Colon cancer | Chemotherapy through ‘Priming’ increases reactive oxygen species (ROS) [33] |
| Alcohol | Murine B16F1 melanoma | apoptosis through suppression of intrinsic pathway for carcinogenesis [34] |
| Ethanol | Spontaneous Estrogen Receptor-negative Mammary Cancer in MMTV/Neu Mice | significant decrease in CCL2 levels in mammary tumors [35] |
| Alcoholic extract | V79 Chinese hamster cells | Radiosensitizer [18] |
| DMSO | Human T leukemia cells | Immunogenic cell death; pro-apoptotic mechanism involves Ca2+ accumulation and generation of ROS [36] |
| Aqueous | Mouse Ehrlich ascites carcinoma | 7.5 Gy gamma radiation combination synergistically [37] |
| WFA | HUVEC cells | Inhibition of NF-kappa B by interference with the ubiquitin-mediated proteasome pathway by increasing levels of poly-ubiquitinated proteins [38] |
| Water | Human malignant melanoma cells | Apoptotic body and nuclear blebbing [39] |
| Water | human MDA-MB-231 breast cancer cells | ROS-dependent mitochondria-mediated apoptosis [40] |
| Water | leukemic THP-1 cells and peripheral blood mononuclear cells (PBMCs) | Modulation of cancer cachexia associated inflammatory cytokines and cell death [41] |
| Leaves | ||
| Methanol | Inflammatory disorders/cancer | Inhibition of NF-kappa B by preventing TNF-induced activation of Ikappa B kinase beta via thiol alkylation-sensitive redox mechanism [42] |
| Methanol | Breast, colon and liver cancer cell lines | Cell cycle arrest at S phase, increase in caspase 3 activity with induction of cell apoptosis [43] |
| Alcoholic | Glioma and YKG1 cell lines | induce senescence-like growth arrest and differentiation in glioma cells [44] |
| Water | Hepatocellular carcinoma | cell cycle arrest at G0/G1 and G2/M phases [45] |
| Water | Neuroinflammation | Microglial inactivation and migration via cell cycle arrest at G0/G1 and G2/M phase [46] |
| Water | Sarcoma, breast cell lines | activation of tumor suppressor proteins p53 and pRB, decrease in cyclin B1 and increase in cyclin D1, downregulation of MMP-3 and -9 [47] |
| Water | Glioma | Suppression of Tumor Growth of Intracranial Allograft of Glioma Cells by GFAP reduction and upregulation of mortalin and NCAM expression [48] |
| Alcohol | glioblastoma and neuroblastoma cells | oxidative stress and induction of differentiation [49] |
| Stems | ||
| Methanol, ethanol and aqueous | Breast cancer (MDA-MB-231) and Vero cell lines | Cytotoxicity of the extracts were found and WFA was found to be the active component in both extracts [50] |
| Ethanol | HeLa, A549, BT474, MDA-MB-231, MDA-MB-453, T47D, MDA-MB-435S, G361, and WM 266.4 cells | Targeting Multidrug Resistance [51] |
| Fruit | ||
| Methanol | Neurological disorders | BBB (blood-brain-barrier) permeability [52] |
| Methanol | HepG2 | changes in the chromatin structure (fragmentation, uniform condensation) [53] |
| Cancer Type | Mechanism of Action |
|---|---|
| Prostate cancer | Par-4-Dependent Apoptosis [57] |
| Myeloid leukemia HL-60 cells | Early ROS generation and mitochondrial dysfunction [58] |
| Breast cancer | FOXO3a (Forkhead box O3)- and Bim-dependent apoptosis [59] |
| Leukemic cells of lymphoid and myeloid origin | Mitochondrial apoptosis by activating p38 MAPK cascade [60] |
| Pancreatic cancer cells | Induction of proteasome inhibition and promotion the accumulation of ubiquitinated proteins, resulting in ER stress-mediated apoptosis [61] |
| Gliobastoma multiforme (GBM) | Orthotopic mouse model showed GBM neurosphere collapsed at nM concentrations [62] |
| Colorectal cancer cells | ROS-dependent mitochondrial dysfunction-mediated apoptosis [63] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dutta, R.; Khalil, R.; Green, R.; Mohapatra, S.S.; Mohapatra, S. Withania Somnifera (Ashwagandha) and Withaferin A: Potential in Integrative Oncology. Int. J. Mol. Sci. 2019, 20, 5310. https://doi.org/10.3390/ijms20215310
Dutta R, Khalil R, Green R, Mohapatra SS, Mohapatra S. Withania Somnifera (Ashwagandha) and Withaferin A: Potential in Integrative Oncology. International Journal of Molecular Sciences. 2019; 20(21):5310. https://doi.org/10.3390/ijms20215310
Chicago/Turabian StyleDutta, Rinku, Roukiah Khalil, Ryan Green, Shyam S Mohapatra, and Subhra Mohapatra. 2019. "Withania Somnifera (Ashwagandha) and Withaferin A: Potential in Integrative Oncology" International Journal of Molecular Sciences 20, no. 21: 5310. https://doi.org/10.3390/ijms20215310
APA StyleDutta, R., Khalil, R., Green, R., Mohapatra, S. S., & Mohapatra, S. (2019). Withania Somnifera (Ashwagandha) and Withaferin A: Potential in Integrative Oncology. International Journal of Molecular Sciences, 20(21), 5310. https://doi.org/10.3390/ijms20215310