Enhancing the Thermo-Stability and Anti-Biofilm Activity of Alginate Lyase by Immobilization on Low Molecular Weight Chitosan Nanoparticles

Abstract
1. Introduction
2. Results and Discussion
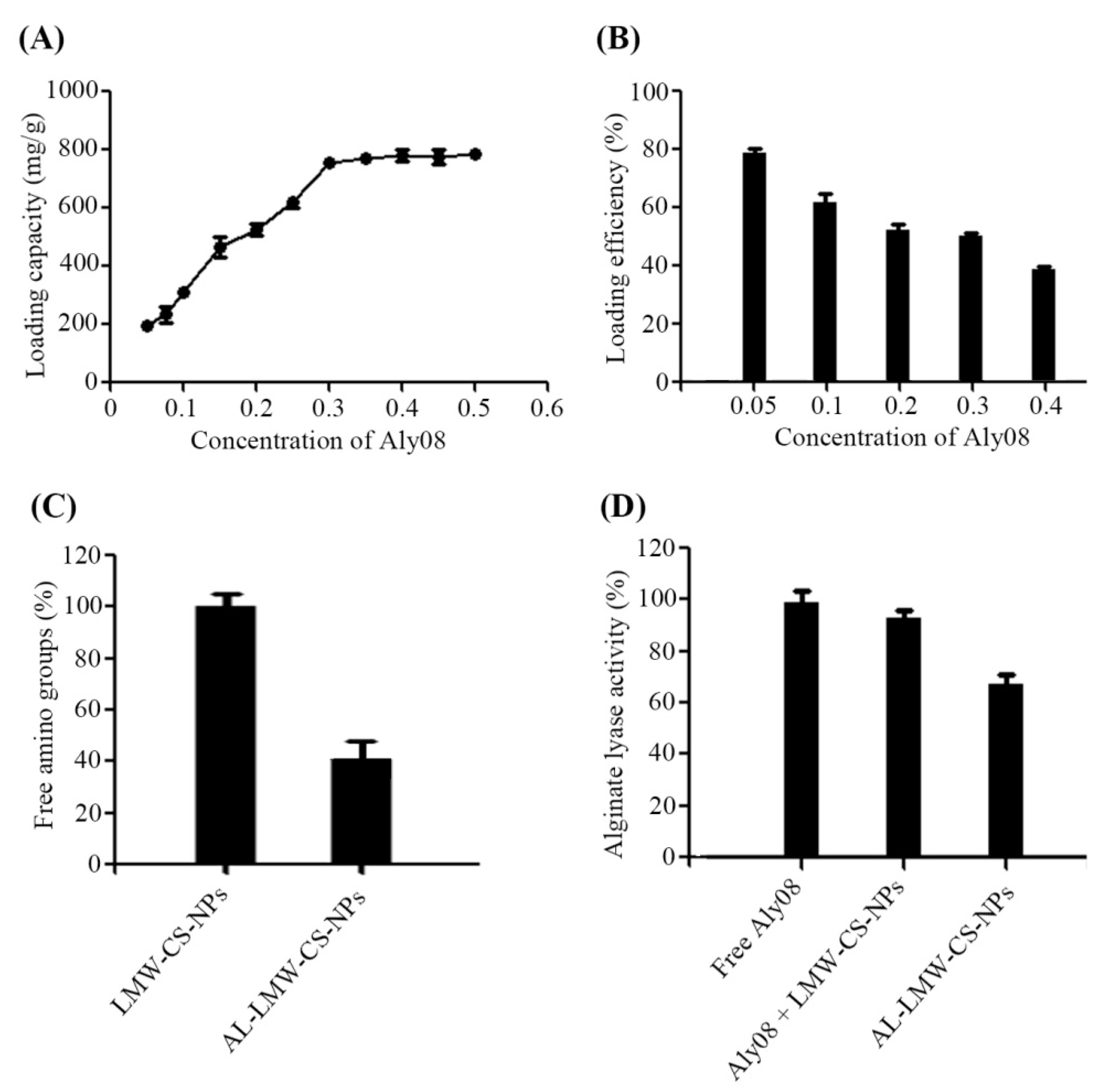
2.1. Synthesis and Determination of Chitosan Nanoparticles
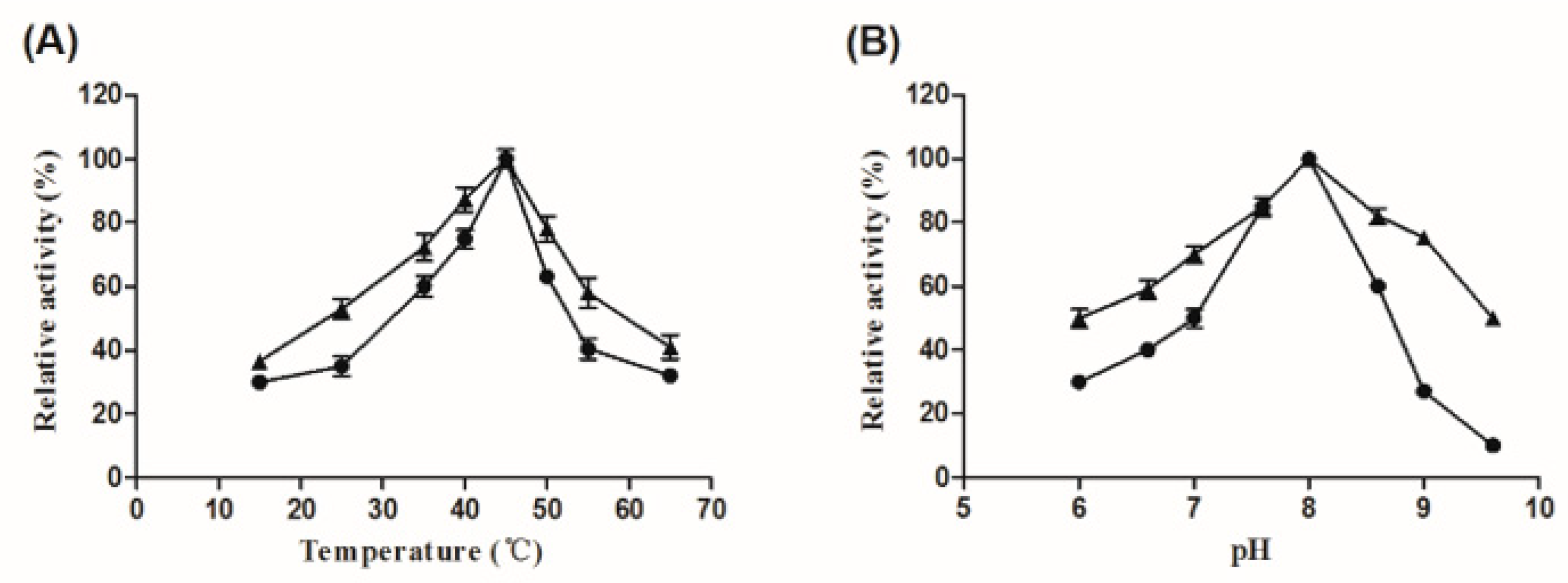
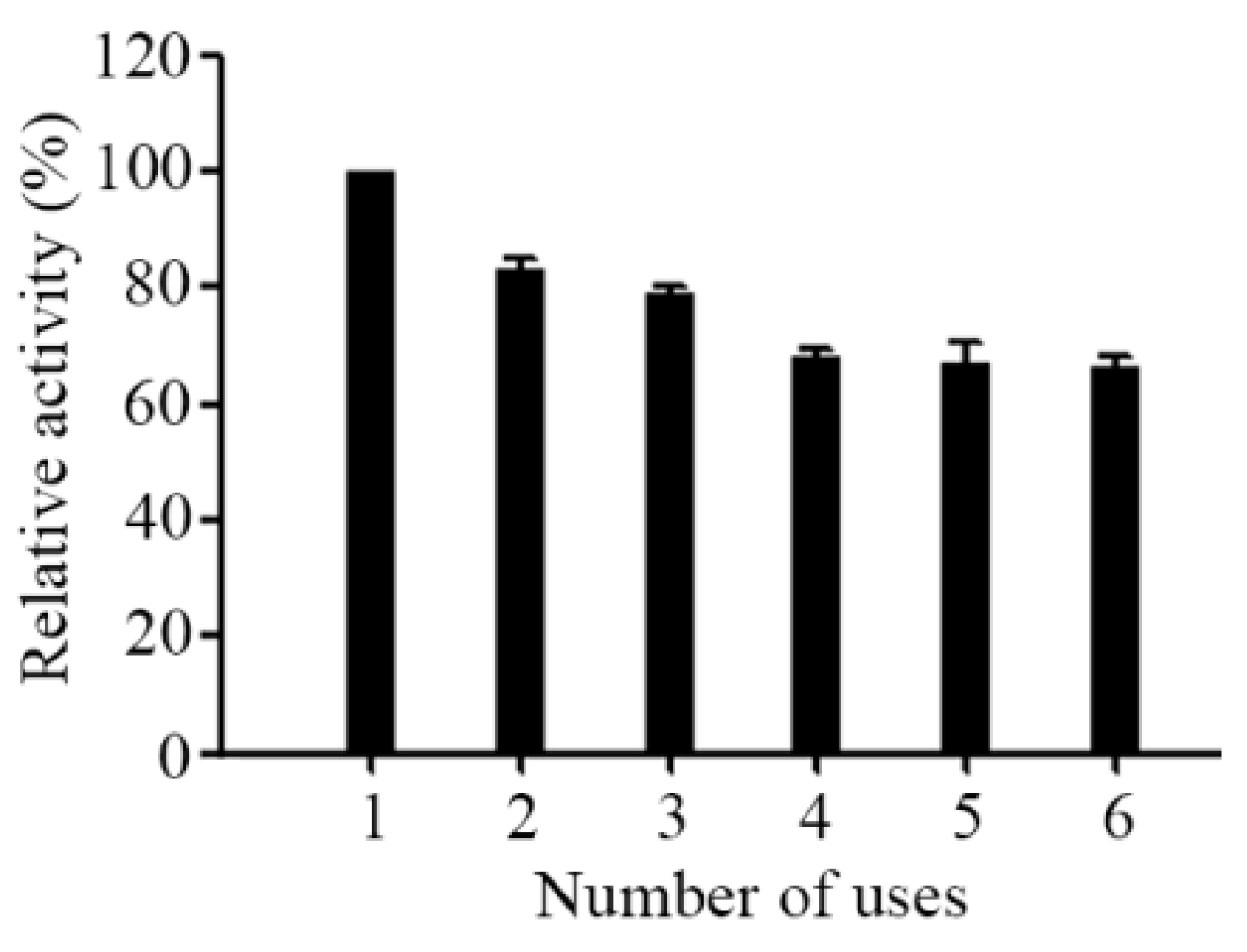
2.2. Biochemical Characterizartion of AL-LMW-CS-NPs
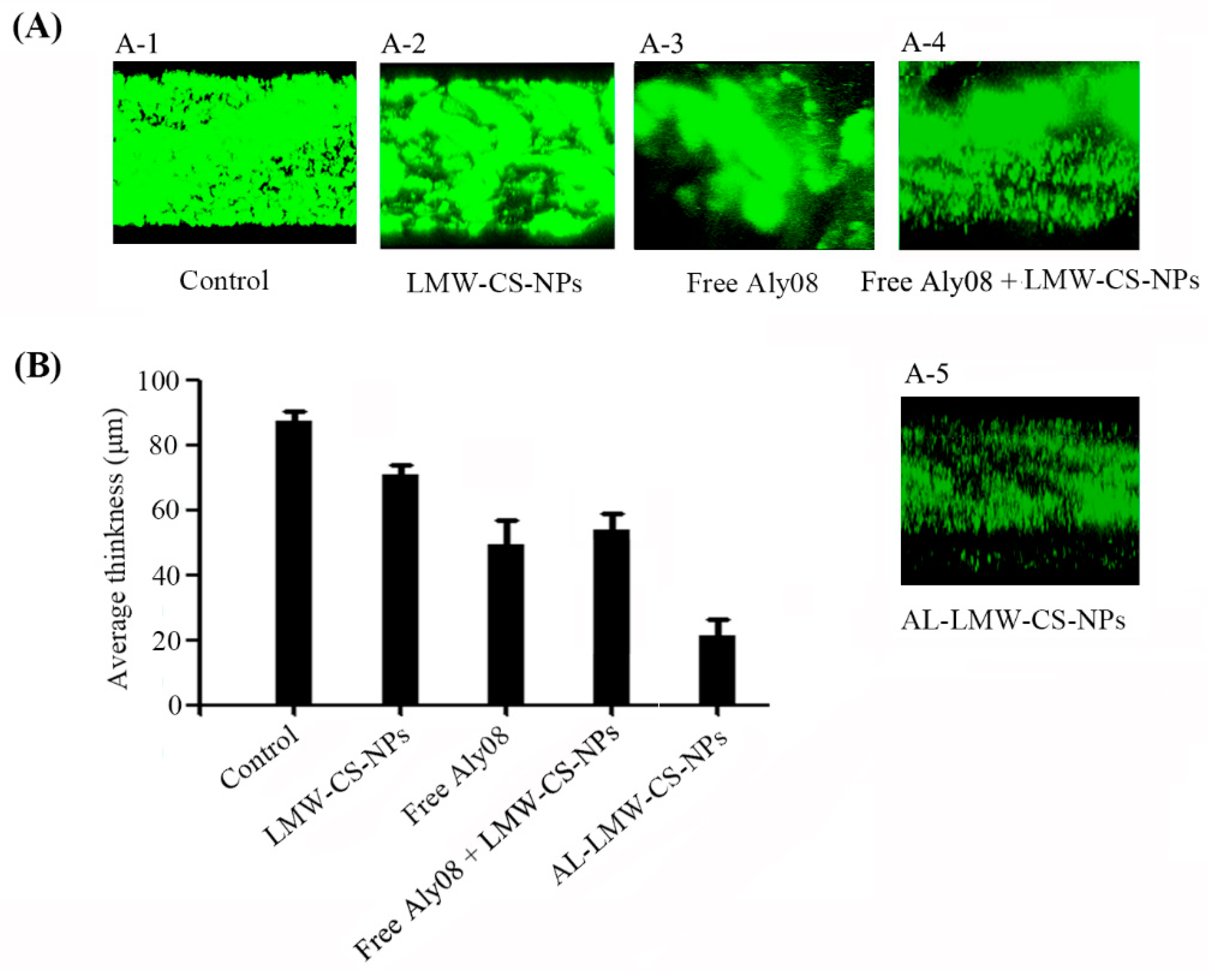
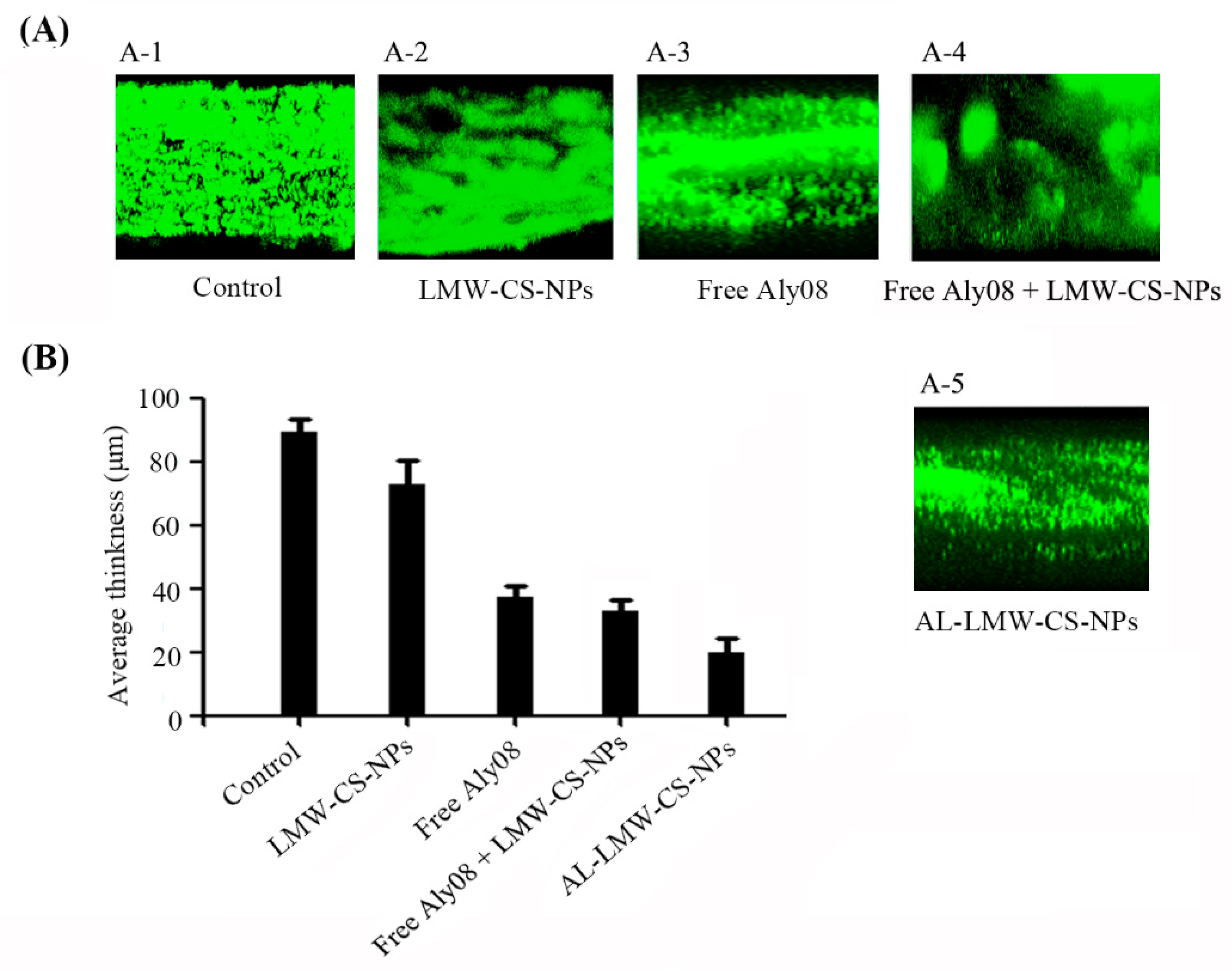
2.3. Anti-Biofilm Activity of Immobilized AL-LMW-CS-NPs
2.4. AL-LMW-CS-NPs Increases the Antibiotics Susceptibility of P. aeruginosa Biofilm
3. Materials and Methods
3.1. Materials
3.2. Preparation of Low Molecular Weight Chitosan
3.3. Synthesis of Chitosan Nanoparticles
3.4. Analysis of the Particle Characterization
3.5. Enzyme Activity and Stability
3.6. Anti-Biofilm Activity
3.7. Antibiotics Susceptibility Assay of Biofilm
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| EPS | extracellular polymeric substance |
| CS-NPs | chitosan nanoparticles |
| CF | Cystic fibrosis |
| LMW-CS | low molecular weight chitosan |
| HMW-CS | high molecular weight chitosan |
| LMW-CS-NPs | low molecular weight chitosan nanoparticles |
| HMW-CS-NP | high molecular weight chitosan nanoparticles |
| AL-LMW-CS-NPs | alginate lyase immobilized low molecular weight chitosan nanoparticles |
| TEM | Transmission electron micrograph |
| CLSM | Confocal laser scanning microscopy |
| TEM | Transmission electron microscopy |
| LC | loading capacity |
| %LE | loading efficiency |
| MIC | minimum inhibitory concentration |
| MBEC | minimum biofilm eradication concentration |
| EDAC | N-(3-Dimethylaminopropyl)-N’- ethylcarbodiimide hydrochloride |
| NHS | N-Hydroxysuccinimide |
| TPP | Tri-polyphosphate |
References
- Roy, R.; Tiwari, M.; Donelli, G.; Tiwari, V. Strategies for combating bacterial biofilms: A focus on anti-biofilm agents and their mechanisms of action. Virulence 2018, 9, 522–554. [Google Scholar] [CrossRef] [PubMed]
- Davies, D.G.; Parsek, M.R.; Pearson, J.P.; Iglewski, B.H.; Costerton, J.W.; Greenberg, E.P. The involvement of cell-to-cell signals in the development of a bacterial biofilm. Science 2018, 9, 522–554. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Yoon, S.S. Pseudomonas aeruginosa Biofilm. a Programmed Bacterial Life for Fitness. J Microbiol Biotechnol. 2017, 27, 1053–1064. [Google Scholar] [CrossRef] [PubMed]
- Sheng, G.P.; Yu, H.Q.; Li, X.Y. Extracellular polymeric substances (EPS) of microbial aggregates in biological wastewater treatment systems: a review. Biotechnol. Adv. 2010, 28, 882–894. [Google Scholar] [CrossRef] [PubMed]
- Skariyachan, S.; Sridhar, V.S.; Packirisamy, S.; Kumargowda, S.T.; Challapilli, S.B. Recent perspectives on the molecular basis of biofilm formation by Pseudomonas aeruginosa and approaches for treatment and biofilm dispersal. Folia Microbiol. (Praha) 2018, 63, 413–432. [Google Scholar] [CrossRef] [PubMed]
- Davies, D. Understanding biofilm resistance to antibacterial agents. Nat. Rev. Drug Discov. 2003, 2, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Hoyle, B.D.; Costerton, J.W. Bacterial resistance to antibiotics: the role of biofilms. Prog. Drug Res. 1991, 37, 91–105. [Google Scholar] [PubMed]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.J.; Cheng, Z. Antibiotic resistance in Pseudomonas aeruginosa: mechanisms and alternative therapeutic strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef]
- Hughes, G.; Webber, M.A. Novel approaches to the treatment of bacterial biofilm infections. B. J. Pharmacol. 2017, 174, 2237–2246. [Google Scholar] [CrossRef]
- Ryan, E.M.; Gorman, S.P.; Donnelly, R.F.; Gilmore, B.F. Recent advances in bacteriophage therapy: how delivery routes, formulation, concentration and timing influence the success of phage therapy. J. Phar. Pharmacol. 2011, 63, 1253–1264. [Google Scholar] [CrossRef]
- Xie, Y.; Chen, S.; Qian, Y.; Zhao, W.; Zhao, C. Photo-responsive membrane surface: Switching from bactericidal to bacteria-resistant property. Mater. Sci. Eng. C. Mater. Biol. Appl. 2018, 84, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Ly-Chatain, M.H. The factors affecting effectiveness of treatment in phages therapy. Front. Microbiol. 2014, 5, 51. [Google Scholar] [CrossRef] [PubMed]
- Sulakvelidze, A.; Alavidze, Z.; Morris, J.G., Jr. Bacteriophage therapy. Antimicrob. Agents Chemother. 2001, 45, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Dosler, S.; Karaaslan, E. Inhibition and destruction of Pseudomonas aeruginosa biofilms by antibiotics and antimicrobial peptides. Peptides 2014, 62, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Wnorowska, U.; Niemirowicz, K.; Myint, M.; Diamond, S.L.; Wroblewska, M.; Savage, P.B.; Janmey, P.A.; Bucki, R. Bactericidal activities of cathelicidin LL-37 and select cationic lipids against the hypervirulent Pseudomonas aeruginosa strain LESB58. Antimicrob. Agents Chemother. 2015, 59, 3808–3815. [Google Scholar] [CrossRef] [PubMed]
- Aoki, W.; Ueda, M. Characterization of antimicrobial peptides toward the development of novel antibiotics. Pharmaceuticals 2013, 6, 1055–1081. [Google Scholar] [CrossRef]
- Tan, Y.; Ma, S.; Liu, C.; Yu, W.; Han, F. Enhancing the stability and antibiofilm activity of DspB by immobilization on carboxymethyl chitosan nanoparticles. Microbiol. Res. 2015, 178, 35–41. [Google Scholar] [CrossRef]
- Jiang, P.; Li, J.; Han, F.; Duan, G.; Lu, X.; Gu, Y.; Yu, W. Antibiofilm activity of an exopolysaccharide from marine bacterium Vibrio sp. QY101. PloS ONE 2011, 6, e18514. [Google Scholar] [CrossRef]
- Rice, K.C.; Mann, E.E.; Endres, J.L.; Weiss, E.C.; Cassat, J.E.; Smeltzer, M.S.; Bayles, K.W. The cidA murein hydrolase regulator contributes to DNA release and biofilm development in Staphylococcus aureus. Proc. Natl. Acad. Sci. USA 2007, 104, 8113–8118. [Google Scholar] [CrossRef]
- Owlia, P.; Nosrati, R.; Alaghehbandan, R.; Lari, A.R. Antimicrobial susceptibility differences among mucoid and non-mucoid Pseudomonas aeruginosa isolates. GMS Hyg. Infect. Control 2014, 9, Doc13. [Google Scholar]
- Patel, K.K.; Tripathi, M.; Pandey, N.; Agrawal, A.K.; Gade, S.; Anjum, M.M.; Tilak, R.; Singh, S. Alginate lyase immobilized chitosan nanoparticles of ciprofloxacin for the improved antimicrobial activity against the biofilm associated mucoid P. aeruginosa infection in cystic fibrosis. Int. J. Pharm. 2019, 563, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Maunders, E.; Welch, M. Matrix exopolysaccharides; the sticky side of biofilm formation. FEMS Microbiol. Lett. 2017, 364, fnx120. [Google Scholar] [CrossRef] [PubMed]
- Verlee, A.; Mincke, S.; Stevens, C.V. Recent developments in antibacterial and antifungal chitosan and its derivatives. Virulence 2018, 9, 522–554. [Google Scholar] [CrossRef] [PubMed]
- Kravanja, G.; Primozic, M.; Knez, Z.; Leitgeb, M. Chitosan-based (Nano)materials for novel biomedical applications. Molecules 2019, 24, 1960. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, X.; Bi, X.; Ren, Y.; Han, Q.; Zhou, Y.; Han, Y.; Yao, R.; Li, S. Characterization of an alkaline alginate lyase with pH-stable and thermo-tolerance property. Mar drugs 2019, 17, 308. [Google Scholar] [CrossRef] [PubMed]
- De Farias, B.S.; Grundmann, D.D.R.; Rizzi, F.Z.; Martins, N.S.S.; Sant’Anna Cadaval Junior, T.R.; de Almeida Pinto, L.A. Production of low molecular weight chitosan by acid and oxidative pathways: Effect on physicochemical properties. Food Res. Int. 2019, 123, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Wattjes, J.; Niehues, A. Enzymatic production and enzymatic-mass spectrometric fingerprinting analysis of chitosan polymers with different nonrandom patterns of acetylation. J. Am. Chem. Soc. 2019, 141, 3137–3145. [Google Scholar] [CrossRef]
- Shinya, S.; Fukamizo, T. Interaction between chitosan and its related enzymes: A review. Int. J. Biol. Macromol. 2017, 104, 1422–1435. [Google Scholar] [CrossRef]
- Qin, Z.; Chen, Q.; Lin, S.; Luo, S.; Qiu, Y.; Zhao, L. Expression and characterization of a novel cold-adapted chitosanase suitable for chitooligosaccharides controllable preparation. Food chem. 2014, 150, 392–399. [Google Scholar] [CrossRef]
- Aranaz, I.; Castro, C.; Heras, A.; Acosta, N. On the ability of low molecular weight chitosan enzymatically depolymerized to produce and stabilize silver nanoparticles. Biomimetics 2018, 3, 21. [Google Scholar] [CrossRef]
- Thadathil, N.; Velappan, S.P. Recent developments in chitosanase research and its biotechnological applications: a review. Food chem. 2014, 150, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Chen, X.; Li, X.; Han, Y.; Wang, Y.; Yao, R.; Li, S. Purification and characterization of a new cold-adapted and thermo-tolerant chitosanase from marine bacterium Pseudoalteromonas sp. SY39. Molecules 2019, 24, 183. [Google Scholar] [CrossRef] [PubMed]
- Nawani, N.; Khurana, J.; Kaur, J. A thermostable lipolytic enzyme from a thermophilic Bacillus sp.: purification and characterization. Mol. Cell. Biochem. 2006, 290, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Yang, S.X.; Liu, Z.M. Rational design of alginate lyase from Microbulbifer sp. Q7 to improve thermal stability. Mar. drugs 2019, 17, 378. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Yu, Y.; Yang, S.; Shi, X.; Mou, H.; Li, L. Expression and characterization of a new polyG-specific alginate lyase from marine bacterium microbulbifer sp. Q7. Front. Microbiol. 2018, 9, 2894. [Google Scholar] [CrossRef] [PubMed]
- Xing, X.; Jia, J.Q.; Zhang, J.F.; Zhou, Z.W.; Li, J.; Wang, N.; Yu, X.Q. CALB immobilized onto magnetic nanoparticles for efficient kinetic resolution of racemic secondary alcohols: long-term stability and reusability. Molecules 2019, 24, 490. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, M.T.; Schiller, N.L. Alginate lyase (AlgL) activity is required for alginate biosynthesis in Pseudomonas aeruginosa. J. Bacteriol. 2005, 187, 3869–3872. [Google Scholar] [CrossRef]
- Alkawash, M.A.; Soothill, J.S.; Schiller, N.L. Alginate lyase enhances antibiotic killing of mucoid Pseudomonas aeruginosa in biofilms. APMIS 2006, 114, 131–138. [Google Scholar] [CrossRef]
- Lamppa, J.W.; Griswold, K.E. Alginate lyase exhibits catalysis-independent biofilm dispersion and antibiotic synergy. Antimicrob. Agents Chemother. 2013, 57, 137–145. [Google Scholar] [CrossRef]
- Taylor, P.K.; Yeung, A.T.; Hancock, R.E. Antibiotic resistance in Pseudomonas aeruginosa biofilms: towards the development of novel anti-biofilm therapies. J. Biotechnol. 2014, 191, 121–130. [Google Scholar] [CrossRef]
- Davoodbasha, M.; Lee, S.Y.; Kim, J.W. Solution plasma mediated formation of low molecular weight chitosan and its application as a biomaterial. Int. J. Biol. Macromol. 2018, 118, 1511–1517. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, L.; Chen, X.; Sun, M.; Han, Y. Design and synthesis of a chitodisaccharide-based affinity resin for chitosanases purification. Mar. drugs 2019, 17, 68. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Wang, D.; Gu, J.; Li, J.; Liu, H.; Li, F.; Han, W. Biochemical characteristics and variable alginate-degrading modes of a novel bifunctional endolytic alginate lyase. Appl. Environ. Microbiol. 2018, 118, 1511–1517. [Google Scholar] [CrossRef] [PubMed]
- Brooun, A.; Liu, S.; Lewis, K. A dose-response study of antibiotic resistance in Pseudomonas aeruginosa biofilms. Antimicro. Agents Chemother. 2000, 44, 640–646. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration(mg/mL) | Piperacillin | Ceftazidime | Amikacin |
|---|---|---|---|
| MIC | 32 | 16 | 16 |
| MBEC | >2048 | 2048 | 1024 |
| MBEC with LMW-CS-NPs | 2048 | 1024 | 512 |
| MBEC with free Aly08 | 512 | 256 | 256 |
| MBEC with free Aly08 + LMW-CS-NPs | 512 | 256 | 256 |
| MBEC with AL-LMW-CS-NPs | 256 | 64 | 64 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Wang, Y.; Li, X.; Lee, B.S.; Jung, S.; Lee, M.-S. Enhancing the Thermo-Stability and Anti-Biofilm Activity of Alginate Lyase by Immobilization on Low Molecular Weight Chitosan Nanoparticles. Int. J. Mol. Sci. 2019, 20, 4565. https://doi.org/10.3390/ijms20184565
Li S, Wang Y, Li X, Lee BS, Jung S, Lee M-S. Enhancing the Thermo-Stability and Anti-Biofilm Activity of Alginate Lyase by Immobilization on Low Molecular Weight Chitosan Nanoparticles. International Journal of Molecular Sciences. 2019; 20(18):4565. https://doi.org/10.3390/ijms20184565
Chicago/Turabian StyleLi, Shangyong, Yanan Wang, Xiao Li, Beom Suk Lee, Samil Jung, and Myeong-Sok Lee. 2019. "Enhancing the Thermo-Stability and Anti-Biofilm Activity of Alginate Lyase by Immobilization on Low Molecular Weight Chitosan Nanoparticles" International Journal of Molecular Sciences 20, no. 18: 4565. https://doi.org/10.3390/ijms20184565
APA StyleLi, S., Wang, Y., Li, X., Lee, B. S., Jung, S., & Lee, M.-S. (2019). Enhancing the Thermo-Stability and Anti-Biofilm Activity of Alginate Lyase by Immobilization on Low Molecular Weight Chitosan Nanoparticles. International Journal of Molecular Sciences, 20(18), 4565. https://doi.org/10.3390/ijms20184565