Abstract
Parkinson’s disease is a progressive neurodegenerative disorder resulting from the degeneration of pigmented dopaminergic neurons in the substantia nigra pars compacta. It induces a series of functional modifications in the circuitry of the basal ganglia nuclei and leads to severe motor disturbances. The amino acid glutamate, as an excitatory neurotransmitter, plays a key role in the disruption of normal basal ganglia function regulated through the interaction with its receptor proteins. It has been proven that glutamate receptors participate in the modulation of neuronal excitability, transmitter release, and long-term synaptic plasticity, in addition to being related to the altered neurotransmission in Parkinson’s disease. Therefore, they are considered new targets for improving the therapeutic strategies used to treat Parkinson’s disease. In this review, we discuss the biological characteristics of these receptors and demonstrate the receptor-mediated neuroprotection in Parkinson’s disease. Pharmacological manipulation of these receptors during anti-Parkinsonian processes in both experimental studies and clinical trials are also summarized.
1. Introduction
Parkinson’s disease (PD) is a debilitating neurodegenerative disorder which is second to Alzheimer’s disease as the most common age-related disease. The clinical symptoms of PD include motor disturbances (such as resting tremor, bradykinesia), rapid eye movement behavior disorder, as well as autonomic and cognitive impairment [1]. The pathology underlying PD comprises the degeneration of dopaminergic neurons in the substantia nigra pars compacta (SNc) and the accumulation of intracytoplasmic inclusions, which are known as Lewy bodies in these neurons [2]. Therefore, dopamimetic drugs, including the dopamine precursor levodopa (l-3,4-dihydroxyphenylalanine, l-DOPA), and dopamine receptor agonists are currently considered as the only standard therapy for treating Parkinsonian symptoms [3]. Although these treatments ameliorate the motor signs of PD for several years in most patients, prolonged therapy frequently leads to the development of motor complications, known as L-DOPA-induced-dyskinesia (LID), such as choreic or larger amplitude choreo-athetotic movements, dystonia, and ballism [4]. It is known that different neurotransmitter systems in the human brain and central nervous system (CNS) are involved in the pathophysiology of PD and LID. Among these, glutamate takes up 40% of all synapses and plays an important role in the mediation of basal ganglia circuitry in continuous feedback, leading to the dopaminergic denervation of the striatum [5]. Moreover, an increasing body of evidence has demonstrated the contribution of glutamatergic transmission to the processes of PD and LID [4]. It was also determined that the concentration of serum glutamate in PD patients is higher than that in healthy subjects [6]. Therefore, it has been suggested that pharmacological therapies with the potential to restore normal glutamatergic functions show promise as therapeutic interventions by reversing the severe motor complications that derive from the current dopamine replacement strategies.
Glutamate plays a critical role in brain function through multiple receptor proteins which are primarily located on pre- and post-synaptic neurons in virtually all areas of the CNS. Figure 1 shows that glutamate receptors were originally classified into two major classes of ionotropic (iGluRs) and metabotropic receptors (mGluRs), according to pharmacological means. The iGluRs are multimeric ion channels and are responsible for fast excitatory transmission in the mammalian CNS. Through binding the presynaptically released glutamate, iGluRs transduce signals into the excitation of postsynaptic neurons on a millisecond timescale. This process generates a synaptic current crucial to brain function and regulates learning and memory. iGluRs can be further classified into N-methyl-d-aspartate (NMDA) receptors, α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors, and kainate receptors. As members of the G-protein receptors superfamily, the mGluRs mediate slow glutamate responses, which contributes to long-lasting changes in synaptic activity [7]. The mGluRs family can be further divided into eight receptor subtypes. Based on their sequence homology, signal transduction mechanisms, and pharmacological profile, these subtypes are classified into three groups. Group I receptors include mGluR1 and mGluR5, which are linked to phospholipase C-mediated polyphosphoinositide hydrolysis, while group II includes mGluR2 and mGluR3, which in recombinant systems are negatively coupled to adenylate cyclase. Group III consists of mGluR4, −6, −7, and −8, which are also negatively coupled to adenylate cyclase or linked to ion channels [8]. In the pathophysiology of PD, there are regulatory alterations of glutamate receptors in specific loci in the basal ganglia. In addition, glutamate receptors are also changed in the process of LID, such as the increased specific binding of NMDA receptors and decreased mGlu2/3 receptors after levodopa treatment in 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-lesioned monkeys [9].
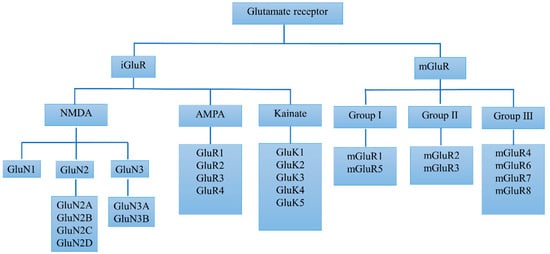
Figure 1.
Classification of glutamate receptors.
Therefore, a comprehensive understanding of glutamate receptors, concerning both the pathophysiology and the treatment targets of PD, may contribute to the development of novel therapeutic approaches to PD. In this review, we discuss the distribution of these different subtypes of glutamate receptors and their neuroprotective properties, as well as evidence of the pharmacological manipulation of these receptors in PD.
2. Basic Biology and Alterations of Glutamate Receptors in PD
Most studies of these receptors are initially focused on their distribution, structure, and complex subunits, as well as functions. However, these factors would change due to dopamine depletion in PD.
2.1. Basic Biology and Alterations of iGluRs in PD
NMDA receptors have been the subject of intense study and are the best characterized of the iGluRs. They are widely expressed in the basal ganglia and directly mediate the excitation of neurons in the striatum, globus pallidus, subthalamic nucleus, substantia nigra reticulata, and substantia nigra compacta [10]. These receptors consist of seven subunits: GluN1, GluN2A, −B, −C, and −D, as well as GluN3A, −B, and −C, which assemble in the form of heterodimers or heterotrimers. Because of their high permeability to calcium ions and their ability to trigger a cascade of downstream calcium-dependent signal-transduction processes of physiological and pathophysiological changes, NMDA receptors play an important role in the regulation of excitatory synaptic transmission. Ligands need to occupy both the glutamate site and the cofactor glycine site for the activation of an NMDA receptor [11]. A glutamate site is a binding pocket on the GluN2A and GluN2B subunits formed through interactions between the N-terminus of the receptor and the extracellular loops. Cofactor glycine binding sites exist in the homologous region of the GluN1 and GluN3 subunits and form an excitatory glycine receptor upon heteromeric assembly. At normal resting membrane potential, the ion channel is blocked by the ambient extracellular magnesium through their binding to a site within the pore, which makes the sites of glutamate and glycine appear occupied [12]. On the other hand, due to the voltage-dependent property of the channel induced by the magnesium blockade, excessive stimulation of NMDA receptors by agonists, such as glutamate and glycine, could induce the depolarization of neurons and lead to the extrusion of Mg2+ from the channel. This allows the ions to flow inward, especially for a Ca2+ influx, by opening the ion channel pore. The accumulation of Ca2+ in cytoplasmic material triggers a series of Ca2+-dependent enzymes and pathophysiological changes in intracellular biochemistry, which may lead to neuronal damage and death. This biofunction on the regulation of both cellular excitability and biochemistry indicates that NMDA receptors play a major role in excitatory synaptic transmission, plasticity, and excitotoxicity [13,14]. It was reported that both PD-induced dopamine depletion and L-DOPA treatment lead to the redistribution of NMDA receptor subunits. For example, the GluN2A subunits and GluN2A/GluN2B subunit ratio were increased in PD patients as well as in levodopa-treated dyskinetic rats and monkeys [15]. Despite the different sensitivities of subunits to dopamine depletion, the expression levels of GluN1 and GluN2B were augmented in the distinct surface compartment of PD rats [16]. A recent study also showed the increased GluN2D subunit in the striatum of L-DOPA-treated Parkinsonian rats [17]. A growing body of literature has shown substantially increased NMDA-sensitive glutamate binding in the striatum and nucleus accumbent in both experimental models of PD and tissue of PD patients, which may accelerate the degenerative process [18]. Therefore, considerable evidence shows that NMDA receptors are viable targets for the treatment of PD and that their antagonists can provide anti-Parkinsonian benefits in various animal models of PD.
AMPA receptors are postsynaptic ion channels that regulate the majority of the fast excitatory amino acid neurotransmitters in the brain. They are cation-selective heterooligomers assembled in various combinations of the subunits GluR1, −2, −3, and −4 (or GluR A, −B, −C, and −D). AMPA receptor subunits are expressed abundantly and localized in different sites within neuronal cell bodies and processed in the cerebral cortex, basal ganglia, limbic system, thalamus, cerebellum, and brainstem, with precise patterns in the different brain regions [19]. In contrast to NMDA receptors, AMPA receptors are weakly permeable to external Ca2+. When gated open by synaptically released glutamate, AMPA receptors mediate the inflow of Na+ (and in some cases Ca2+) into neurons, while they cause the efflux of K+. Although the ion channels assembled by the homomeric or heteromeric combinations of GluR1 and GluR3 show an appreciable Ca2+-permeability, the incorporation of a GIuR2 subunit suppresses Ca 2+ permeability [20]. It has also been demonstrated that the activation of AMPA receptors increases the influx of extracellular Zn2+ into nigral dopaminergic neurons, which leads to movement disorder [21]. This suggests that Ca 2+ and Zn2+-permeable GluR2-lacking AMPA receptors are particularly important for the treatment of PD. In the 6-OHDA rat model, AMPA receptor-mediated excitatory transmission was enhanced, which increased the release of glutamate [22]. Also, the elevated expressions of AMPA receptors in Parkinsonian monkeys induced using MPTP and dopomimetic treatment have been observed in subregions of the striatum [23]. In addition, a previous study noted the upregulation of AMPA glutamate receptors in the lateral putamen of advanced PD patients experiencing levodopa-induced motor complications (both abnormal involuntary movements and “wearing-off” phenomenon) when compared to others without motor complications [24]. Therefore, antagonists of AMPA receptors could show potential anti-Parkinsonian effects.
The progress in the study of the characteristics and functions of kainate (KA) receptors is lagging more than that of NMDA and AMPA receptors due to the lack of selective kainite receptor (KAR) antagonists and agonists. Kainate receptors are also ligand-gated channels with permeability to cations. They consist of subunits of Gluk1, −2, −3 (previously called GluR5, −6, and −7), and −4, −5 (previously known as KA1 and KA2). KA receptors are highly expressed in the CNS, with the distribution of all subunits occurring in the neocortex [25]. Cells from different regions express different subunits of KA receptors, which is consistent with the fact that most GluK2 and GluK5 subunits are distributed in the rodent cortex [26]. According to the measurement of affinity for kainate receptors, the affinity of GluK1, GluK2, and GluK3 subunits is low, while that of subunits GluK4 and GluK5 is high [27]. Among all the subtypes, GluK4 and GluK5 assemble homomeric or heteromeric receptors with subunits of GluK1 to GluK3 to form functional channels. This is because subunits with a high affinity cannot form functional channels, so they co-assemble with low-affinity subtypes. Although kainate receptors do not cross-assemble with AMPA receptors, there are still many common characteristics between them. However, kainate receptors, found in both intracellular and extrasynaptic distribution in the striatum, share more common features with metabotropic receptors than with ionotropic receptors, which suggests that kainate receptors could exert functions of metabotropic receptors in the striatum [28]. KARs exist in fewer synapses and physiological studies have suggested that KARs mainly contribute to the modulation of synaptic transmission, which is different from the functions of NMDA and AMPA receptors. The latter are the main postsynaptic targets for glutamate that is released synaptically. For example, it was reported that presynaptic KAR activation inhibits GABAergic outflow from the striatum in the rat globus pallidus [29], which may be helpful for improving the treatment of PD, since increased GABAergic activity from the striatum to the globus pallidus is known as one of the cardinal features of PD pathophysiology. KARs also contribute to both short- and long-term synaptic plasticity in the hippocampus, as well as the sensory cortex [30,31]. It is worth noting that loss of parkin function leads to the increased expression of KAR, which may have a pathogenetic role in parkin-related autosomal recessive juvenile parkinsonism [32].
2.2. Basic Biology and Alterations of mGluRs in PD
The group I mGluRs consists of mGluR1 and mGluR5, with the main distribution in the postsynaptic position of dendrites and spines. In the primate striatum, they are also found in the presynaptic sites of nigrostriatal dopamine terminals [33]. The expressions of mGluR1 in cerebellar Purkinje cells, striatonigral, and striatopallidal projection neurons and striatal interneurons are most intensive [34]. A longitudinal positron emission tomography study showed the dynamic changes in the expression of mGluR1 that accompany pathological progression in a PD model of rats which mimics clinical pathology [35]. Intense expression of mGluR5 has been found predominantly in telencephalic regions, such as the cerebral cortex, hippocampus, subiculum, main and accessory olfactory bulbs, anterior olfactory nucleus, olfactory tubercle, striatum, nucleus accumbens, and lateral septal nucleus [36,37]. Recently, a positron emission tomography study showed that the expression of mGluR5 was increased in strategic dopaminergic brain regions of PD patients [38]. Moreover, the genetic knockdown of mGluR5 was determined to decrease LID in an aphakia mouse model of PD [39]. In addition, the binding potential of mGluR5 receptor was also decreased in the 6-OHDA rat model of PD [40]. In physiological conditions, the acute regulation of mGlu5 receptor signaling in cortical astrocytes causes oscillatory changes of Ca2+ and the synaptic release of neurotransmitters, as well as the programming of transcriptional events [41]. This effect on Ca2+ may bring about the interaction between mGlu5 receptors and NMDA receptors. It was reported that mGlu5 receptors enhanced NR2B-containing NMDA receptor activity in the rat hippocampus through the phosphorylation of NR2B [42]. Therefore, a negative allosteric modulator of group I mGluRs, especially for mGlu5 receptors, would be promising to show anti-Parkinsonian effects through the reduction of the excitatory drive in the overstimulation basal ganglia nuclei.
Group II mGluRs, including mGluR2 and mGluR3, are differentially distributed. The expression of mGluR2 is less than that of mGluR3in the CNS. mGluR2 is mainly expressed in Golgi cells in the cerebellar cortex, mitral cells of the accessory olfactory bulb, the external part of the anterior olfactory nucleus, and some neurons in the entorhinal and parasubicular cortices [43]. mGluR3 is widely distributed in the olfactory tubercle, dentate gyrus, cerebral cortex, nucleus accumbens, lateral septal nucleus, striatum, amygdaloid nuclei, cerebellar cortex, and substantia nigra pars reticulata [44]. Functionally, the activation of group II mGluRs regulates corticostriatal synapses and inhibits the generation of thalamus-derived glutamate as well as dopamine from the ventral midbrain, through a direct or indirect presynaptic manner [45]. A recent study showed that expressions of mGluR2/3 protein decreased in 6-OHDA-lesioned rats [46]. A mechanism study also revealed that the activation of mGluR2/3 restored the clearance of extracellular glutamate that was destroyed in astrocytes [47]. Therefore, it has been suggested that selective agonists or positive allosteric modulators of group II mGluR may contribute to PD treatment, due to their contribution to the attenuation of glutamate levels [48].
Group III receptors consist of mGluR4, −6, −7, and −8, which are distributed throughout the basal ganglia circuitry. Among group III mGluRs, the expression of mGluR7 is the highest, with extensive distribution in the hippocampus, thalamus, neocortex, amygdala, hypothalamus, and locus coeruleus, whereas that of mGluR6 is predominantly limited to the retina [49,50]. The location pattern of mGluR8 in the CNS has been found at the presynaptic level in the cerebellum, olfactory bulb, hippocampus, and cortical areas, which is also more restricted than that of mGluR7 [51]. Although mGluR4 is mainly distributed in the cerebellum, it also has been proved to be located in other areas, such as the cerebral cortex, olfactory bulb, hippocampus, lateral septum, septofimbrial nucleus, striatum, thalamic nuclei, lateral mammillary nucleus, pontine nuclei, and dorsal horn [52]. In line with the intense expression, mice lacking mGluR4 show a clear impairment in their ability to learn complex motor tasks [53]. Moreover, the activation of mGluR4 may have a neuroprotective effect through the inhibition of glutamate production in the substantia nigra and the reduction of inflammatory effects [54]. Positive allosteric modulators of mGluR4 have been proposed for the symptomatic management of PD and they have been shown to induce a reduction of motor symptoms in animal models of PD. For example, studies have demonstrated that positive allosteric modulators of mGluR4 provide functional neuroprotection against nigrostriatal impairment induced by 6-OHDA in rats and MPTP in macaques [55,56]. Although studies of the anti-Parkinsonian and neuroprotective properties of mGluR6 and mGluR7 are fewer than those addressing mGluR4, these receptor subunits have been proven to increase the proliferation of neural stem cells (NSCs) by promoting cell cycle progression and inhibiting the apoptosis of NSCs [57,58]. NSCs are able to self-renew as well as practice differentiation into the neurons and glia of the nervous system. Many studies have transplanted NSCs to promote the restoration of function after spinal cord injury [59]. Therefore, targeting mGluR6 and mGluR7 on NSCs may also be benefit for the stem cell-based strategy for the treatment of PD.
3. Key Targets of Glutamate Receptors in PD Treatment
The treatment of PD through targeting glutamate receptors can be summarized in three observations: (1) the improvement of PD motor symptoms, (2) the increase of the anti-Parkinsonian efficiency of dopaminergic agents, and (3) the protection of nigral neurons.
3.1. Key Targets of iGluRs in PD Treatment
The antagonists of NMDA reporters include competitive antagonists (e.g., SDZ 220–581, MDL 100, 453), noncompetitive antagonists (e.g., MK-801, dextrorphan, PD 174494, CP-101, 606) as well as glycine site antagonists (e.g., MRZ 2/570, L-701,324, 7-chlorokynurenate, (R)-HA-966) [60]. Plenty of studies have demonstrated that NMDA receptor antagonists can attenuate catalepsy and counteract Parkinsonian rigidity induced by dopamine receptor antagonists in rats as well as akinesia and other motor complications in monoamine-depleted rodents [61,62,63,64]. Similar effects were also shown in an ifenprodil-treated MPTP-lesioned marmoset model [65]. In addition, subthreshold doses of several NMDA receptor antagonists have been determined to synergistically increase the anti-Parkinsonian efficiency of levodopa and other drugs, such as 7-nitroindazole and opioid glycopeptide lactomorphin, as well as prophylactically prevent the appearance of motor response alterations associated with chronic levodopa administration LID both in rat and primate models of PD [66,67,68]. This suggests that these drugs may act as adjunct therapies for improving the efficacy and tolerability of dopaminergic therapies, as well as the combination between different antagonists. On the other hand, the extensive expression of NMDA receptors and widespread excitation stimulated by NMDA has resulted in concern that the global inhibition of these receptors could lead to various unwanted side effects, such as ataxia, impaired learning, and psychosis [69]. Therefore, more and more research is focused on the screening of a series of NMDA subunit-selective drugs. NR2B was demonstrated to be heavily distributed in the striatum and other basal ganglia regions [70,71]. This suggests that therapy targeting NR2B-containing receptors can contribute to more specific effects influencing the NMDA receptor regulation in brain regions related to PD pathophysiology. This was determined by Ifenprodil and traxoprodil (CP-101, 606), antagonists of the NR2B subunit-containing NMDA receptor, which were shown to ameliorate Parkinsonian symptoms and reduce LID in rodents and MPTP-lesioned monkeys [72,73]. The combination therapy of NR2B-selective antagonists with other drugs was also applied. For example, the combination of the GluN2B-selective antagonist Radiprodil and the A2A antagonist Tozadenant led to significant motor improvement both in 6-OHDA-lesioned rats and MPTP-lesioned marmosets [74,75]. In addition, a series of photo-switchable antagonists for the modulation of NMDA receptors have been synthesized by researchers. Among these, PNRA showed selectivity for GluN2A and GluN2C rather than GluN2B, which should be addressed in further studies in more advanced biological settings for potential application in PD treatment [76].
The anti-Parkinsonian properties of AMPA antagonists have been evaluated in different animal PD models. 2,3-Dihydroxy-6-nitro-7-sulfamoyl-benzo[f]quinoxaline-2,3-dione (NBQX), a selective antagonist of AMPA receptors, has been reported to suppress muscular rigidity in monoamine-depleted rats and produce clinically apparent improvement motor deficits in MPTP-lesioned aged Rhesus monkeys [77]. However, many studies have reported that AMPA receptor antagonists are not effective on anti-Parkinsonian action in animal models when given alone. For example, NBQX fails to reverse motor deficits in 6-OHDA-lesioned rats when administered alone [78,79]. Another interesting finding in the same research was that, during co-administration, AMPA receptor antagonists such as NBQX synergistically ameliorated Parkinsonian symptomatology with levodopa in 6-OHDA-lesioned rats and MPTP-lesioned common marmoset. This suggests that AMPA receptor antagonists may be used as adjuvants of levodopa for the improvement of treatment efficacy. In addition, the development of LID promotes the expression and activity of AMPA receptors, which suggests that the blockage of AMPA receptors may alleviate LID therapeutically [24,80]. For example, after administration of the noncompetitive AMPA receptor antagonist LY300164 (talampanel) for levodopa-treated Parkinsonian monkeys lesioned with MPTP, the motor activities were potentiated and LIDs were decreased [23]. In addition, although perampanel, a novel AMPA receptor antagonist, was determined to penetrate the blood–brain barrier, adjunctive Perampanel had no effect on improving LID in PD patients [81,82].
At present, several evaluations of pharmacological properties have identified limited compounds that are promising as selective antagonists of kainate receptors, when most target GluK5. For example, compounds of the quinoxalinedione family, such as CNQX and NBQX, play the role of competitive antagonists with affinity for both native and recombinant kainate receptors [26]. Usually, these antagonists are shared with AMPA receptors and are far more effective on AMPA receptors than that on KA receptors. Therefore, some pyrrolyl-quinoxalinedione derivatives with a more potent affinity for kainate receptors have been developed. Take LU97175 for example—it was more selective towards kainate receptors, especially for the sites of kainite subunits with low affinity (i.e., GluK1 to GluK3), which showed anticonvulsant effects without the induction of motor dysfunction [83]. In addition, glutamate receptor antagonists from a new series of 6-substituted decahydroisoquinoline, such as LY382884 and LY377770, display higher affinities with kainate receptors containing the GluK1 subunit than with AMPA and NMDA receptors [84,85]. In addition, due to the global distribution of AMPA/NMDA receptors, the use of AMPA/NMDA receptor antagonists in humans may lead to severe side effects, which is worse because of their fast excitatory effects. In contrast, KARs mainly affect slower excitatory modulation, which is more like the function of mGluRs [29]. This may be achieved through the mediated role of KARs in overactive striatopallidal GABAergic synapses and glutamatergic subthalamopallidal transmission [29], suggesting that KARs represent a huge potential target for the development of a novel therapeutic strategy for PD. However, almost all of the abovementioned studies on GluK1-selective antagonists with promising pharmacological efficiency focus on animal models of pain, migraine, epilepsy, stroke, and anxiety, rather than that of PD. Therefore, direct evidence of the effect of KA receptor antagonists on anti-Parkinsonian action is urgently needed. In addition, novel selective antagonists of other subunits of KA receptors need further development.
3.2. Key Targets of mGluRs in PD Treatment
mGluR5 receptors have been reported to be efficiently suppressed by a series of compounds such as phenylpyridine derivatives (2-methyl-6-(phenylethynyl)-pyridine (MPEP), 3-[(2-methyl-1,3-thiazol-4-yl)ethynyl]pyridine (MTEP), AFQ056/mavoglurant, fenobam, and ADX48621/dipraglurant), which are systemically active [86]. Consistent with this hypothesis, several studies have proven that these antagonists of the mGluR5 receptor exert anti-Parkinsonian effects in different animal models of PD. For example, both MPEP-treated and mGluR5 knockout mice displayed a higher survival probability and a reduced extent of nigrostriatal damage induced by the administration of the dopaminergic neurotoxin MPTP, which was also mimicked by SIB1893 [87]. Furthermore, MPEP also alleviated LID in 6-OHDA-lesioned rats [88]. With higher solubility than MPEP, MTEP was also proven to significantly reduce the degeneration of catecholaminergic neurons in MPTP-treated monkeys and ameliorate LID in an MPTP-lesioned macaque model of PD [89,90]. Taken together, these data obtained from rodent and monkey models suggest that antagonists of mGluR5 are potential drugs that can reduce the degeneration of monoaminergic neurons in PD. The lower dosage combination of MPEP and the NMDA receptor antagonist MK-801 reversed motor deficits in 6-OHDA-lesioned rats, which suggests that the simultaneous blockade of mGluR5 and NMDA receptors may have more beneficial effects on PD treatment [91]. As a derivative of MPEP, Mavoglurant could increase the anti-Parkinsonian duration of levodopa in MPTP-lesioned monkeys [92]. Compared to MTEP and MPEP, Dipraglurant showed more promising effects in reducing overall LID severity and peak-dose dyskinesia in macaques [93,94]. In contrast to mGluR5, fewer studies focused on the assessment of mGluR1 as a potential target of anti-Parkinsonian actions. In a study comparing the efficiency of the mGluR5 antagonist MTEP and the mGluR1 antagonist (3-ethyl-2-methyl-quinoline-6-yl)-(4-methoxy-cyclohexyl)-methanone-methanesulfonate (EMQMCM) in different models of PD, the results showed that it was MTEP, but not EMQMCM, that inhibited levodopa-induced rotations and alleviated LIDs. This suggested that mGluR1 may not be an effective target for the symptomatic treatment of PD [95]. There is not a consensus about the exact mechanisms of the neuroprotective effects brought about by the antagonists of mGluR5. Except for the abovementioned interaction with NMDA receptors, the suppression of glial mGluR5 may decrease inflammatory damage and relieve MPTP-induced toxicity to midbrain dopaminergic neurons [96].
In contrast to mGluR5, it has been suggested that selective agonists or positive allosteric modulators of group II mGluR may contribute to PD treatment due to their presynaptic reduction of the corticostriatal transmission of glutamate, which is overstimulated in PD models. Consistent with this hypothesis, mGluR2/3 agonists (S)-(+)-alpha-amino-4-carboxy-2-methylbenzeneacetic acid (LY379268) attenuated akinesia in phencyclidine- and amphetamine-treated mouse models [97]. In addition, the inhibition of group II mGluRs using the antagonist LY341495 increased the evoked excitatory postsynaptic currents in rat SNc neurons [98]. Consistent with these observations, the group II mGluR agonists LY379268 and (2R,4R)-4-aminopyrrolidine-2,4-dicarboxylate (2R,4R-APDC) were proven to display neuroprotective effects through the reduction of the extent toxicity in a 6-OHDA-lesioned rodent PD model [99]. Similar results were observed in the combination treatment of LY379268 and the mGluR2/3 receptor agonist (2S,2’R,3’R)-2-(2’,3’-dicarboxycyclopropyl) glycine (DCG-IV) on 6-OHDA-lesioned rats with reduced corticostriatal transmission, providing support for group II mGluR agonists as potential neuroprotective drugs for PD treatment [100].
Similar to the findings for group II mGluR, accumulating evidence suggests that mGlu4 receptor activation may be beneficial for the treatment of PD. Systemic or intrapleural administration of the mGluR4 agonist Phenyl-7-(hydroxyamino) cyclopropopa[b] chrome-1a-carboxamide (PHCCC) contributed to the reduction of the extent of nigrostriatal toxicity induced by MPTP in wildtype mice, but not in mGluR4-deficient mice. This supports the potential of the selective activation of mGluR4 as a therapeutic strategy for the treatment of PD [101]. In addition, the mGluR4 positive allosteric modulator VU0364770 could potentiate the motor stimulation of a subthreshold L-DOPA dosage in 6-OHDA-lesioned rats [102]. Another mGluR4 positive allosteric modulator, VU0652957 (VU2957, valiglurax), has been processed to fabricate a spray-dried dispersion formulation for clinical application [103]. However, in contrast to mGluR4, the studies on the anti-Parkinsonian and neuroprotective properties of other group III mGluR subtypes are limited. One study of 6-OHDA-lesion rats showed that the intranigral infusion of the mGluR8-selective agonist (S)-3,4-dicarboxyphenylglycine (DCPG) induced mild catalepsy, which suggested that mGluR8 activation can reduce the anti-Parkinsonian effects induced by other group III mGluRs in the substantia nigra [104]. Therefore, subtype-selective agonists should be considered when taking group III mGluRs as targets for the treatment of PD.
4. Clinical Trials Targeting Glutamate Receptors in PD
Though the obtained preclinical evaluations indicated the potential efficacy of targeting glutamate receptors for the treatment of parkinsonism, clinical trials in PD patients are still relatively insufficient and still under development. As of now, only the weak NMDA antagonist amantadine has become widely applied for the treatment of dyskinesias. The pharmacology of amantadine is complex and lacks a well-defined mechanism of action, yet it does exert an anti-Parkinsonian effect through “therapeutic” (i.e., low micromolar) concentrations on NMDA antagonist activity [105]. In a large retrospective series involving 836 Parkinsonian patients, amantadine treatment was shown to improve the survival of patients, which suggests its neuroprotective properties through NMDA receptor antagonism [106]. In addition, another uncompetitive NMDA receptor antagonist, dextromethorphan/quinidine, showed clinical benefits for the treatment of LID in a study involving 13 PD patients [107]. However, clinical treatment with other NMDA antagonists for PD patients is limited. One reason for this is that several NMDA antagonists with promising preclinical results failed in clinical trials owing to intolerable side effects, as complete glutamate antagonism may cause adverse cognitive effects. Therefore, comprehensive investigations of the characteristics of NMDA receptor subtypes and the use of more specific antagonists that do not result in a global receptor blockade could improve therapeutic efficacy while preventing the presence of side effects [108]. However, a recent study on the NR2B selective NMDA receptor antagonist MK-0657 suggested that a single dose of MK-0657 failed to improve LID and motor symptoms in PD patients [109]. In comparison to NMDA receptors, there are fewer clinical reports of targeting AMPA receptors for the treatment of PD. Perampanel, a selective AMPA receptor antagonist, has been used in two multicenter randomized, double-blind, placebo-controlled, parallel-group phase III studies. Although it showed better toleration and safety, no clinically significant improvement of levodopa-induced motor fluctuations was found [110,111,112]. In addition, talampanel has also been used in clinical studies to evaluate the efficacy and tolerability in levodopa-treated patients with PD, but no available data were obtained (NCT00108667). Although several drugs targeting AMPA receptors in recent clinical studies have assessed the efficacy of treating LID, the results of these studies are unavailable. More trials concerning the potential of AMPA receptor antagonists as adjunct therapies for improving levodopa treatment are required. In addition, a positive finding is that some mGluR5 antagonists, such as AFQ056-mavoglurant and ADX-48621-dipraglurant, have been assessed in human trials as anti-dyskinetic drugs. The obtained data prove the safety and good tolerance of these drugs, while no worsening PD motor symptoms occurred [94,113]. However, two randomized phase II studies showed that mavoglurant failed to improve LID in PD patients [114]. Therefore, further investigations with a larger number of patients are urgently need so that mGluR5 could be considered a potential target for PD treatment.
5. Conclusions
In the present review, we discussed the basic biological characteristics of glutamate receptors and their alterations in PD development and anti-Parkinsonian processes (summarized in Table 1). Glutamate receptors are distributed in almost all neural cell types. Subunit composition and expression variants significantly differ between different cell types and brain regions. Furthermore, the expression levels of subunits and variants change with the development of PD. Antagonists of NMDA receptors display beneficial effects on reversing motor symptoms, reducing LIDs, and slowing progressive neurodegeneration in preclinical PD models. While the results of targeting AMPA receptors are complex, their antagonists are effective in the treatment of LID, and their agonist displays the potential for neuroprotection. Pharmacological regulation of mGluRs shows even more potential for PD treatment because of their ability to fine-tune neurotransmission. The inhibition of mGluR5 and the activation of group II mGluRs, as well as mGluR4, have exhibited pharmacological efficiency in different animal models of PD. These therapy strategies (using the antagonists and agonist) can reverse motor disorder and provide a neuroprotective effect. However, in clinical practice, treatments targeting these glutamate receptors face some challenges due to the constant failure of previous treatments in PD [115]. Although the reasons for past failures require further discussion, some interesting possibilities have been investigated, such as ignoring the disease stage of patients [116], a lack of a basic disease mechanism, a lack of animal models with precise replicas of etiopathogenesis, uncertainty concerning drug dosing and binding site, and a lack of proper clinical trial methodology [117]. Moreover, some receptors act as a “double-edged sword”. Taking NMDA receptors as an example, their antagonists are usually used to target NMDA reporters. However, a recent study showed the reduction of the NMDA receptor co-agonist D-serine and the NMDA receptor subunits GluN1 and GluN2B in MPTP-lesioned macaques and the cerebrospinal fluid of PD patients [118]. Therefore, future investigations should consider the balance between receptor-related pro-death and pro-survival effects. In addition, traditional treatment mainly focuses on the improvement of motor dysfunction and ignores non-motor symptoms in PD, such as anxiety, apathy, cognitive dysfunction, and depression [119].

Table 1.
Alterations of glutamate receptors during PD and anti-Parkinsonian process.
For all the above-outlined receptors, the selectivity of novel compounds is still the key procedure under development. This requires a further understanding of the precise roles of glutamate receptors in the regulation of basal ganglia circuitry both in normal physiological and pathophysiological conditions, as well as the development of the more selective compounds for certain subtypes so that rational designs for combination therapy can be formulated. Further promising studies on primate Parkinson’s disease models are necessary to determine the potential of clinical trials when combination therapy is employed to maximize efficacy and avoid the induction of unexpected side effects.
Author Contributions
Conceptualization, K.K.-L.Y.; Literature collection and preparation, Z.Z. (Zhang Zhang) and K.L.; Writing—original draft preparation, Z.Z. (Zhu Zhang) and S.Z.; Writing—review and editing, P.F., J.K.-S.K., and K.K.-L.Y. All authors read and approved the final manuscript.
Funding
This work was supported in part by the National Natural Science Foundation of China (Grant No. 81703728).
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| PD | Parkinson’s disease |
| SNc | substantia nigra pars compacta |
| L-DOPA | l-3,4-dihydroxyphenylalanine |
| CNS | central nervous system |
| iGluRs | ionotropic glutamate receptors |
| mGluRs | metabotropic glutamate receptors |
| NMDA | N-methyl-d-aspartate |
| AMPA | α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid |
| KA | kainate |
| KA receptor | KAR |
| NSCs | neural stem cells |
| 6-OHDA | 6-hydroxydopamine |
| MPTP | 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine |
| DHPG | 3,5-dihydroxyphenylglycine |
| LID | levodopa-induced dyskinesia |
| NBQX | 2,3-dihydroxy-6-nitro-7-sulfamoyl-benzo[f]quinoxaline-2,3-dione |
| MPEP | 2-methyl-6-(phenylethynyl)-pyridine |
| MTEP | 3-[(2-methyl-1,3-thiazol-4-yl) ethynyl] pyridine |
| EMQMCM | (3-ethyl-2-methyl-quinoline-6-yl)-(4-methoxy-cyclohexyl)-methanone methanesulfonate |
| 2R,4R-APDC | (2R,4R)-4-aminopyrrolidine-2,4-dicarboxylate |
| PHCCC | Phenyl-7-(hydroxyamino) cyclopropopa[b] chrome-1a-carboxamide |
| DCPG | (S)-3,4-dicarboxyphenylglycine |
| NAMs | Negative allosteric modulators |
| PAMs | Positive allosteric modulators |
References
- De, P.E.; Lees, A.J.; Holton, J.L.; Warner, T.T. Prognosis and neuropathologic correlation of clinical subtypes of Parkinson disease. JAMA Neurol. 2019, 76, 470–479. [Google Scholar]
- Dickson, D.W. Neuropathology of Parkinson disease. Parkinsonism Relat. Disord. 2018, 46, S30–S33. [Google Scholar] [CrossRef]
- Chen, J.J.; Swope, D.M. Pharmacotherapy for Parkinson’s disease. Pharmacotherapy 2007, 27, 161S–173S. [Google Scholar] [CrossRef]
- Bastide, M.F.; Meissner, W.G.; Picconi, B.; Fasano, S.; Fernagut, P.O.; Feyder, M.; Francardo, V.; Alcacer, C.; Ding, Y.; Brambilla, R.; et al. Pathophysiology of L-dopa-induced motor and non-motor complications in Parkinson’s disease. Prog. Neurobiol. 2015, 132, 96–168. [Google Scholar] [CrossRef]
- Blandini, F.; Porter, R.H.; Greenamyre, J.T. Glutamate and Parkinson’s disease. Mol. Neurobiol. 1996, 12, 73–94. [Google Scholar] [CrossRef]
- Mironova, Y.S.; Zhukova, I.A.; Zhukova, N.G.; Alifirova, V.M.; Izhboldina, O.P.; Latypova, A.V. Parkinson’s disease and glutamate excitotoxicity. Zhurnal nevrologii i psikhiatrii imeni SS Korsakova 2018, 118, 50–54. [Google Scholar] [CrossRef]
- Carrillo, M.P.; Silva, A.D.; Villaseñor, A.K. Glutamate in Parkinson’s disease: Role of antiglutamatergic drugs. Basal Ganglia 2013, 3, 147–157. [Google Scholar] [CrossRef]
- Pin, J.P.; Duvoisin, R. The metabotropic glutamate receptors: Structure and functions. Neuropharmacology 1995, 34, 1–26. [Google Scholar] [CrossRef]
- Jourdain, V.A.; Morin, N.; Grégoire, L.; Morissette, M.; Di, P.T. Changes in glutamate receptors in dyskinetic parkinsonian monkeys after unilateral subthalamotomy. J. Neurosurg. 2015, 123, 1383–1393. [Google Scholar] [CrossRef]
- Götz, T.; Kraushaar, U.; Geiger, J.; Lübke, J.; Berger, T.; Jonas, P. Functional properties of AMPA and NMDA receptors expressed in identified types of basal ganglia neurons. J. Neurosci. 1997, 17, 204–215. [Google Scholar] [CrossRef]
- Mothet, J.P.; Matildé, L.B.; Billard, J.M. Time and space profiling of NMDA receptor co-agonist functions. J. Neurochem. 2015, 135, 210–225. [Google Scholar] [CrossRef]
- Seeburg, P.H.; Burnashev, N.; Köhr, G.; Kuner, T.; Sprengel, R.; Monyer, H. The NMDA receptor channel: Molecular design of a coincidence detector. Recent Prog. Horm. Res. 1995, 50, 19–34. [Google Scholar]
- Mody, I.; MacDonald, J.F. NMDA receptor-dependent excitotoxicity: The role of intracellular Ca2+ release. Trends Pharm. Sci. 1995, 16, 356–359. [Google Scholar] [CrossRef]
- Plutino, S.; Sciaccaluga, M.; Fucile, S. Extracellular mild acidosis decreases the Ca2+ permeability of the human NMDA receptors. Cell Calcium 2019, 80, 63–70. [Google Scholar] [CrossRef]
- Mellone, M.; Stanic, J.; Hernandez, L.F.; Iglesias, E.; Zianni, E.; Longhi, A.; Prigent, A.; Picconi, B.; Calabresi, P.; Hirsch, E.C.; et al. NMDA receptor GluN2A/GluN2B subunit ratio as synaptic trait of levodopa-induced dyskinesias: From experimental models to patients. Front. Cell. Neurosci. 2015, 9, 245. [Google Scholar] [CrossRef]
- Gan, J.; Qi, C.; Mao, L.M.; Liu, Z. Changes in surface expression of N-methyl-D-aspartate receptors in the striatum in a rat model of Parkinson’s disease. Drug Des. Dev. Ther. 2014, 8, 165. [Google Scholar]
- Mellone, M.; Zianni, E.; Stanic, J.; Campanelli, F.; Marino, G.; Ghiglieri, V.; Longhi, A.; Thiolot, M.L.; Li, Q.; Calabresi, P.; et al. NMDA receptor GluN2D subunit participates to levodopa-induced dyskinesia pathophysiology. Neurobiol. Dis. 2019, 121, 338–349. [Google Scholar] [CrossRef]
- Guo, H.; Camargo, L.M.; Yeboah, F.; Digan, M.E.; Niu, H.; Pan, Y.; Reiling, S.; Soler-Llavina, G.; Weihofen, W.A.; Wang, H.R.; et al. A NMDA-receptor calcium influx assay sensitive to stimulation by glutamate and glycine/D-serine. Sci. Rep. 2017, 7, 11608. [Google Scholar] [CrossRef]
- Martin, L.J.; Blackstone, C.D.; Levey, A.I.; Huganir, R.L.; Price, D.L. AMPA glutamate receptor subunits are differentially distributed in rat brain. Neuroscience 1993, 53, 327–358. [Google Scholar] [CrossRef]
- Hollmann, M.; Hartley, M.; Heinemann, S. Ca2+ permeability of KA-AMPA--gated glutamate receptor channels depends on subunit composition. Science 1991, 252, 851–853. [Google Scholar] [CrossRef]
- Tamano, H.; Morioka, H.; Nishio, R.; Takeuchi, A.; Takeda, A. AMPA-induced extracellular Zn2+ influx into nigral dopaminergic neurons causes movement disorder in rats. Neurotoxicology 2018, 69, 23–28. [Google Scholar] [CrossRef]
- Chang, Y.; Du, C.; Han, L.; Lv, S.; Zhang, J.; Bian, G.; Tang, G.; Liu, Y.; Chen, T.; Liu, J. Enhanced AMPA receptor-mediated excitatory transmission in the rodent rostromedial tegmental nucleus following lesion of the nigrostriatal pathway. Neurochem. Int. 2019, 122, 85–93. [Google Scholar] [CrossRef]
- Ouattara, B.; Hoyer, D.; Grégoire, L.; Morissette, M.; Gasparini, F.; Gomez-Mancilla, B.; Paolo, T.D. Changes of AMPA receptors in MPTP monkeys with levodopa-induced dyskinesias. Neuroscience 2010, 167, 1160–1167. [Google Scholar] [CrossRef]
- Calon, F.; Rajput, A.H.; Hornykiewicz, O.; Bédard, P.J.; Di, P.T. Levodopa-induced motor complications are associated with alterations of glutamate receptors in Parkinson’s disease. Neurobiol. Dis. 2003, 14, 404–416. [Google Scholar] [CrossRef]
- Hadzic, M.; Jack, A.; Wahle, P. Ionotropic glutamate receptors: Which ones, when, and where in the mammalian neocortex. J. Comp. Neurol. 2017, 525, 976–1033. [Google Scholar] [CrossRef]
- Pinheiro, P.; Mulle, C. Kainate receptors. Cell Tissue Res. 2006, 326, 457–482. [Google Scholar] [CrossRef]
- Evans, A.J.; Gurung, S.; Henley, J.M.; Nakamura, Y.; Wilkinson, K.A. Exciting times: New advances towards understanding the regulation and roles of kainate receptors. Neurochem. Res. 2019, 44, 572–584. [Google Scholar] [CrossRef]
- Kieval, J.Z.; Hubert, G.W.; Charara, A.; Paré, J.F.; Smith, Y. Subcellular and subsynaptic localization of presynaptic and postsynaptic kainate receptor subunits in the monkey striatum. J. Neurosci. 2001, 21, 8746–8757. [Google Scholar] [CrossRef]
- Jin, X.T.; Smith, Y. Activation of presynaptic kainate receptors suppresses GABAergic synaptic transmission in the rat globus pallidus. Neuroscience 2007, 149, 338–349. [Google Scholar] [CrossRef][Green Version]
- Lauri, S.E.; Vesikansa, A.; Segerstråle, M.; Collingridge, G.L.; Isaac, J.T.; Taira, T. Functional maturation of CA1 synapses involves activity-dependent loss of tonic kainate receptor-mediated inhibition of glutamate release. Neuron 2006, 50, 415–429. [Google Scholar] [CrossRef]
- Daw, M.I.; Scott, H.L.; Isaac, J.T. Developmental synaptic plasticity at the thalamocortical input to barrel cortex: Mechanisms and roles. Mol. Cell. Neurosci. 2007, 34, 493–502. [Google Scholar] [CrossRef][Green Version]
- Maraschi, A.; Ciammola, A.; Folci, A.; Sassone, F.; Ronzitti, G.; Cappelletti, G.; Silani, V.; Sato, S.; Hattori, N.; Mazzanti, M.; et al. Parkin regulates kainate receptors by interacting with the GluK2 subunit. Nat. Commun. 2014, 5, 5182. [Google Scholar] [CrossRef]
- Masilamoni, G.J.; Smith, Y. Metabotropic glutamate receptors: Targets for neuroprotective therapies in Parkinson disease. Curr. Opin. Pharm. 2018, 38, 72–80. [Google Scholar] [CrossRef]
- Mishina, M.; Suzuki, M.; Ishii, K.; Sakata, M.; Wagatsuma, K.; Ishibashi, K.; Toyohara, J.; Zhang, M.R.; Kimura, K.; Ishiwata, K. Density of metabotropic glutamate receptors subtype 1 in Parkinson’s disease compared to healthy elderly—A ITMM PET study-. J. Neurol. Sci. 2017, 381, 806–807. [Google Scholar] [CrossRef]
- Yamasaki, T.; Fujinaga, M.; Kawamura, K.; Furutsuka, K.; Nengaki, N.; Shimoda, Y.; Shiomi, S.; Takei, M.; Hashimoto, H.; Yui, J.; et al. Dynamic changes in striatal mGluR1 but not mGluR5 during pathological progression of Parkinson’s disease in human alpha-synuclein A53T transgenic rats: A multi-PET imaging study. J. Neurosci. 2016, 36, 375–384. [Google Scholar] [CrossRef]
- Shigemoto, R.; Nomura, S.; Ohishi, H.; Sugihara, H.; Nakanishi, S.; Mizuno, N. Immunohistochemical localization of a metabotropic glutamate receptor, mGluR5, in the rat brain. Neurosci. Lett. 1993, 163, 53–57. [Google Scholar] [CrossRef]
- Romano, C.; Sesma, M.A.; McDonald, C.T.; O’malley, K.; VandenPol, A.N.; Olney, J.W. Distribution of metabotropic glutamate receptor mGluR5 immunoreactivity in rat brain. J. Comp. Neurol. 1995, 355, 455–469. [Google Scholar] [CrossRef]
- Kang, Y.; Henchcliffe, C.; Verma, A.; Vallabhajosula, S.; He, B.; Kothari, P.J.; Pryor, K.O.; Mozley, P.D. 18F-FPEB PET/CT Shows mGluR5 Upregulation in Parkinson’s Disease. J. Neuroimaging 2019, 29, 97–103. [Google Scholar] [CrossRef]
- García-Montes, J.R.; Solís, O.; Enríquez, T.J.; Ruiz, D.I.; Drucker, C.R.; Moratalla, R. Genetic Knockdown of mGluR5 in Striatal D1R-Containing Neurons Attenuates l-DOPA-Induced Dyskinesia in Aphakia Mice. Mol. Neurobiol. 2019, 56, 4037–4050. [Google Scholar] [CrossRef]
- Crabbé, M.; Vander, P.A.; Weerasekera, A.; Himmelreich, U.; Baekelandt, V.; Van, L.K.; Casteels, C. Altered mGluR5 binding potential and glutamine concentration in the 6-OHDA rat model of acute Parkinson’s disease and levodopa-induced dyskinesia. Neurobiol. Aging 2018, 61, 82–92. [Google Scholar] [CrossRef]
- Bradley, S.; Challiss, R. Defining protein kinase/phosphatase isoenzymic regulation of mGlu5 receptor-stimulated phospholipase C and Ca2+ responses in astrocytes. Br. J. Pharm. 2011, 164, 755–771. [Google Scholar] [CrossRef]
- Sarantis, K.; Tsiamaki, E.; Kouvaros, S.; Papatheodoropoulos, C.; Angelatou, F. Adenosine A2A receptors permit mGluR5-evoked tyrosine phosphorylation of NR 2B (Tyr1472) in rat hippocampus: A possible key mechanism in NMDA receptor modulation. J. Neurochem. 2015, 135, 714–726. [Google Scholar] [CrossRef]
- Ohishi, H.; Ogawa-Meguro, R.; Shigemoto, R.; Kaneko, T.; Nakanishi, S.; Mizuno, N. Immunohistochemical localization of metabotropic glutamate receptors, mGluR2 and mGluR3, in rat cerebellar cortex. Neuron 1994, 13, 55–66. [Google Scholar] [CrossRef]
- Tamaru, Y.; Nomura, S.; Mizuno, N.; Shigemoto, R. Distribution of metabotropic glutamate receptor mGluR3 in the mouse CNS: Differential location relative to pre-and postsynaptic sites. Neuroscience 2001, 106, 481–503. [Google Scholar] [CrossRef]
- Johnson, K.A.; Mateo, Y.; Lovinger, D.M. Metabotropic glutamate receptor 2 inhibits thalamically-driven glutamate and dopamine release in the dorsal striatum. Neuropharmacology 2017, 117, 114–123. [Google Scholar] [CrossRef]
- Jia, Y.J.; Deng, J.H.; Zhang, W.Z.; Sun, Z.L.; Yang, J.; Yu, Y.; Gong, X.L.; Jia, J.; Wang, X.M. The role of group II metabotropic glutamate receptors in the striatum in electroacupuncture treatment of Parkinsonian rats. CNS Neurosci. Ther. 2017, 23, 23–32. [Google Scholar] [CrossRef]
- Lin, C.H.; You, J.R.; Wei, K.C.; Gean, P.W. Stimulating ERK/PI3K/NFκB signaling pathways upon activation of mGluR2/3 restores OGD-induced impairment in glutamate clearance in astrocytes. Eur. J. Neurosci. 2014, 39, 83–96. [Google Scholar] [CrossRef]
- Shi, K.; Liu, X.; Hou, L.; Qiao, D.; Lin, X. Effects of exercise on mGluR-mediated glutamatergic transmission in the striatum of hemiparkinsonian rats. Neurosci. Lett. 2019, 705, 143–150. [Google Scholar] [CrossRef]
- Nakajima, Y.; Iwakabe, H.; Akazawa, C.; Nawa, H.; Shigemoto, R.; Mizuno, N.; Nakanishi, S. Molecular characterization of a novel retinal metabotropic glutamate receptor mGluR6 with a high agonist selectivity for L-2-amino-4-phosphonobutyrate. J. Biol. Chem. 1993, 268, 11868–11873. [Google Scholar]
- Kinoshita, A.; Shigemoto, R.; Ohishi, H.; Vander, P.H.; Mizuno, N. Immunohistochemical localization of metabotropic glutamate receptors, mGluR7a and mGluR7b, in the central nervous system of the adult rat and mouse: A light and electron microscopic study. J. Comp. Neurol. 1998, 393, 332–352. [Google Scholar] [CrossRef]
- Corti, C.; Restituito, S.; Rimland, J.M.; Brabet, I.; Corsi, M.; Pin, J.P.; Ferraguti, F. Cloning and characterization of alternative mRNA forms for the rat metabotropic glutamate receptors mGluR7 and mGluR8. Eur. J. Neurosci. 1998, 10, 3629–3641. [Google Scholar] [CrossRef]
- Ferraguti, F.; Shigemoto, R. Metabotropic glutamate receptors. Cell Tissue Res. 2006, 326, 483–504. [Google Scholar] [CrossRef]
- Gerlai, R.; Roder, J.C.; Hampson, D.R. Altered spatial learning and memory in mice lacking the mGluR4 subtype of metabotropic glutamate receptor. Behav. Neurosci. 1998, 112, 525. [Google Scholar] [CrossRef]
- Avdeeva, N.V.; Sidorova, S.A.; Gudyrev, O.S.; Ol’ga, A.O.; Golubev, I.V. Mechanism of neuroprotective effect of mGluR4 agonists. Res. Results Pharmacol. 2019, 5, 43. [Google Scholar] [CrossRef][Green Version]
- Betts, M.J.; O’neill, M.J.; Duty, S. Allosteric modulation of the group III mGlu4 receptor provides functional neuroprotection in the 6-hydroxydopamine rat model of Parkinson’s disease. Br. J. Pharm. 2012, 166, 2317–2330. [Google Scholar] [CrossRef]
- Charvin, D.; Di, P.T.; Bezard, E.; Halldin, C.; Duvey, G.; Gregoire, L.; Takano, A.; Pioli, E.; Medori, R.; Conquet, F. A Novel mglur4 compound alleviates motor symptoms in primate models of parkinson’s disease. J. Neurol. Sci. 2017, 381, 97. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, L.; Chen, Y.; Shi, H.; Huang, X.; Wang, Y.; Wei, Y.; Xue, W.; Han, J. Effect and mechanism of mGluR6 on the biological function of rat embryonic neural stem cells. Biosci. Biotechnol. Biochem. 2019, 83, 1027–1034. [Google Scholar] [CrossRef]
- Zhang, J.; Zhao, J.; Chen, Y.; Shi, H.; Huang, X.; Wang, Y.; Wang, Y.; Wei, Y.; Xue, W.; Han, J. Effect of mGluR7 on proliferation of human embryonic neural stem cells. Medicine 2019, 98, e14683. [Google Scholar] [CrossRef]
- Feng, S.; Xiao, J.; Han, F.; Chen, L.; Gao, W.; Mao, G.; Huang, H. Neurorestorative clinical application standards for the culture and quality control of neural progenitor/precursor cells (version 2017). J. Neurorestoratol. 2018, 6, 65–68. [Google Scholar] [CrossRef]
- Johnson, K.A.; Conn, P.J.; Niswender, C.M. Glutamate receptors as therapeutic targets for Parkinson’s disease. CNS Neurol. Disord. Drug Targets 2009, 8, 475–491. [Google Scholar] [CrossRef]
- Bubser, M.; Zadow, B.; Kronthaler, U.O.; Felsheim, U.; Rückert, N.G.; Schmidt, W.J. Behavioural pharmacology of the non-competitive NMDA antagonists dextrorphan and ADCI: Relations between locomotor stimulation, anticataleptic potential and forebrain dopamine metabolism. Naunyn-Schmiedebergs Arch Pharm. 1997, 355, 767–773. [Google Scholar] [CrossRef]
- Melo-Thomas, L.; Gil-Martínez, A.L.; Cuenca, L.; Estrada, C.; Gonzalez-Cuello, A.; Schwarting, R.K.; Herrero, M.T. Electrical stimulation or MK-801 in the inferior colliculus improve motor deficits in MPTP-treated mice. Neurotoxicology 2018, 65, 38–43. [Google Scholar] [CrossRef]
- Konieczny, J.; Ossowska, K.; Schulze, G.; Coper, H.; Wolfarth, S. L-701,324, a selective antagonist at the glycine site of the NMDA receptor, counteracts haloperidol-induced muscle rigidity in rats. Psychopharmacology 1999, 143, 235–243. [Google Scholar] [CrossRef]
- Bartlett, M.J.; Joseph, R.M.; LePoidevin, L.M.; Parent, K.L.; Laude, N.D.; Lazarus, L.B.; Heien, M.L.; Estevez, M.; Sherman, S.J.; Falk, T. Long-term effect of sub-anesthetic ketamine in reducing L-DOPA-induced dyskinesias in a preclinical model. Neurosci. Lett. 2016, 612, 121–125. [Google Scholar] [CrossRef]
- Nash, J.E.; Fox, S.H.; Henry, B.; Hill, M.P.; Peggs, D.; McGuire, S.; Maneuf, Y.; Hille, C.; Brotchie, J.M.; Crossman, A.R. Anti-Parkinsonian actions of ifenprodil in the MPTP-lesioned marmoset model of Parkinson’s disease. Exp. Neurol. 2000, 165, 136–142. [Google Scholar] [CrossRef]
- Bortolanza, M.; Bariotto-Dos-Santos, K.D.; Dos-Santos-Pereira, M.; da-Silva, C.A.; Del-Bel, E. Antidyskinetic Effect of 7-Nitroindazole and Sodium Nitroprusside Associated with Amantadine in a Rat Model of Parkinson’s Disease. Neurotox. Res. 2016, 30, 88–100. [Google Scholar] [CrossRef]
- Flores, A.J.; Bartlett, M.J.; Root, B.K.; Parent, K.L.; Heien, M.L.; Porreca, F.; Polt, R.; Sherman, S.J.; Falk, T. The Combination of the Opioid Glycopeptide MMP-2200 and a NMDA Receptor Antagonist Reduced L-Dopa-Induced Dyskinesia and MMP-2200 by Itself Reduced Dopamine Receptor 2-Like Agonist-Induced Dyskinesia. Neuropharmacology 2018, 141, 260–271. [Google Scholar] [CrossRef]
- Blanchet, P.J.; Konitsiotis, S.; Whittemore, E.R.; Zhou, Z.L.; Woodward, R.M.; Chase, T.N. Differing effects ofN-methyl-D-aspartate receptor subtype selective antagonists on dyskinesias in levodopa-treated 1-methyl-4-phenyl-tetrahydropyridine monkeys. J. Pharm. Exp. Ther. 1999, 290, 1034–1040. [Google Scholar]
- Starr, M. Anti-Parkinsonian actions of glutamate antagonists-alone and with L-DOPA: A review of evidence and suggestions for possible mechanisms. J. Neural. Transm. Parkinson’s Dis. Dement. Sect. 1995, 10, 141–185. [Google Scholar] [CrossRef]
- Jin, D.H.; Jung, Y.W.; Ko, B.H.; Moon, I.S. Immunoblot Analyses on the Differential Djstri~ ution of NR2A and NR2B Subunits in the Adult Rat Brain. Mol. Cells 1997, 7, 749–754. [Google Scholar]
- Schito, A.M.; Pizzuti, A.; Di, M.E.; Schenone, A.; Ratti, A.; Defferrari, R.; Bellone, E.; Mancardi, G.; Ajmar, F.; Mandich, P. mRNA distribution in adult human brain of GRIN2B, a N-methyl-D-aspartate (NMDA) receptor subunit. Neurosci. Lett. 1997, 239, 49–53. [Google Scholar] [CrossRef]
- Steece-Collier, K.; Chambers, L.K.; Jaw-Tsai, S.S.; Menniti, F.S.; Greenamyre, J.T. Anti-Parkinsonian actions of CP-101,606, an antagonist of NR2B subunit-containing N-methyl-d-aspartate receptors. Exp. Neurol. 2000, 163, 239–243. [Google Scholar] [CrossRef]
- Igarashi, M.; Habata, T.; Akita, H.; Noda, K.; Ogata, M.; Saji, M. The NR2B Antagonist, Ifenprodil, Corrects the L-Dopa-Induced Deficit of Bilateral Movement and Reduces C-Fos Expression in the Subthalamic Nucleus of Hemiparkinsonian Rats. Neurosci. Res. 2015, 96, 45–53. [Google Scholar] [CrossRef]
- Michel, A.; Downey, P.; Van, D.X.; De, W.C.; Schwarting, R.; Scheller, D. Behavioural Assessment of the A2a/NR2B Combination in the Unilateral 6-OHDA-Lesioned Rat Model: A New Method to Examine the Therapeutic Potential of Non-Dopaminergic Drugs. PLoS ONE 2015, 10, e0135949. [Google Scholar] [CrossRef]
- Michel, A.; Nicolas, J.M.; Rose, S.; Jackson, M.; Colman, P.; Briône, W.; Sciberras, D.; Muglia, P.; Scheller, D.K.; Citron, M.; et al. Anti-Parkinsonian Effects of the “Radiprodil and Tozadenant” Combination in MPTP-Treated Marmosets. PLoS ONE 2017, 12, e0182887. [Google Scholar] [CrossRef]
- Hartrampf, F.W.; Barber, D.M.; Gottschling, K.; Leippe, P.; Hollmann, M.; Trauner, D. Development of a photoswitchable antagonist of NMDA receptors. Tetrahedron 2017, 73, 4905–4912. [Google Scholar] [CrossRef]
- Klockgether, T.; Turski, L.; Honor, T.; Zhang, Z.; Gash, D.M.; Kurlan, R.; Greenamyre, T. The AMPA receptor antagonist NBQX has anti-Parkinsonian effects in monoamine-depleted rats and MPTP-treated monkeys. Ann. Neurol. 1991, 30, 717–723. [Google Scholar] [CrossRef]
- Löschmann, P.; Kunow, M.; Wachtel, H. Synergism of NBQX with dopamine agonists in the 6-OHDA rat model of Parkinson’s disease. J. Neural Transm. 1992, 38, 55–64. [Google Scholar]
- Wachtel, H.; Kunow, M.; Löschmann, P.A. NBQX (6-nitro-sulfamoyl-benzo-quinoxaline-dione) and CPP (3-carboxy-piperazin-propyl phosphonic acid) potentiate dopamine agonist induced rotations in substantia nigra lesioned rats. Neurosci. Lett. 1992, 142, 179–182. [Google Scholar] [CrossRef]
- Brotchie, J.M. Nondopaminergic mechanisms in levodopa-induced dyskinesia. Mov. Disord. 2005, 20, 919–931. [Google Scholar] [CrossRef]
- Paul, D.; Allakonda, L.; Sahu, A.; Surendran, S.; Satheeshkumar, N. Pharmacokinetics and brain uptake study of novel AMPA receptor antagonist perampanel in SD rats using a validated UHPLC-QTOF-MS method. J. Pharm. Biomed. Anal. 2018, 149, 234–241. [Google Scholar] [CrossRef]
- Lattanzi, S.; Grillo, E.; Brigo, F.; Silvestrini, M. Efficacy and safety of perampanel in Parkinson’s disease. A systematic review with meta-analysis. J. Neurol. 2018, 265, 733–740. [Google Scholar] [CrossRef]
- Löscher, W.; Lehmann, H.; Behl, B.; Seemann, D.; Teschendorf, H.J.; Hofmann, H.P.; Lubisch, W.; Höger, T.; Lemaire, H.G.; Groß, G. A new pyrrolyl-quinoxalinedione series of non-NMDA glutamate receptor antagonists: Pharmacological characterization and comparison with NBQX and valproate in the kindling model of epilepsy. Eur. J. Neurosci. 1999, 11, 250–262. [Google Scholar] [CrossRef]
- Bortolotto, Z.A.; Clarke, V.R.; Delany, C.M.; Parry, M.C.; Smolders, I.; Vignes, M.; Ho, K.H.; Miu, P.; Brinton, B.T.; Fantaske, R.; et al. Kainate receptors are involved in synaptic plasticity. Nature 1999, 402, 297. [Google Scholar] [CrossRef]
- O’Neill, M.J.; Bond, A.; Ornstein, P.L.; Ward, M.A.; Hicks, C.A.; Hoo, K.; Bleakman, D.; Lodge, D. Decahydroisoquinolines: Novel competitive AMPA/kainate antagonists with neuroprotective effects in global cerebral ischaemia. Neuropharmacology 1998, 37, 1211–1222. [Google Scholar] [CrossRef]
- Pourmirbabaei, S.; Dolatshahi, M.; Rahmani, F. Pathophysiological clues to therapeutic applications of glutamate mGlu5 receptor antagonists in levodopa-induced dyskinesia. Eur. J. Pharm. 2019, 855, 149–159. [Google Scholar] [CrossRef]
- Battaglia, G.; Busceti, C.L.; Molinaro, G.; Biagioni, F.; Storto, M.; Fornai, F.; Nicoletti, F.; Bruno, V. Endogenous activation of mGlu5 metabotropic glutamate receptors contributes to the development of nigro-striatal damage induced by 1-methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine in mice. J. Neurosci. 2004, 24, 828–835. [Google Scholar] [CrossRef]
- Huang, Y.; Shu, H.; Li, L.; Zhen, T.; Zhao, J.; Zhou, X.; Luo, W. L-DOPA-Induced Motor Impairment and Overexpression of Corticostriatal Synaptic Components Are Improved by the mGluR5 Antagonist MPEP in 6-OHDA-Lesioned Rats. ASN Neuro 2018, 10, 11. [Google Scholar] [CrossRef]
- Johnston, T.H.; Fox, S.H.; McIldowie, M.J.; Piggott, M.J.; Brotchie, J.M. Reduction of L-DOPA-induced dyskinesia by the selective metabotropic glutamate receptor 5 antagonist 3-[(2-methyl-1, 3-thiazol-4-yl) ethynyl] pyridine in the 1-methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine-lesioned macaque model of Parkinson’s disease. J. Pharm. Exp. Ther. 2010, 333, 865–873. [Google Scholar] [CrossRef]
- Masilamoni, G.J.; Bogenpohl, J.W.; Alagille, D.; Delevich, K.; Tamagnan, G.; Votaw, J.R.; Wichmann, T.; Smith, Y. Metabotropic glutamate receptor 5 antagonist protects dopaminergic and noradrenergic neurons from degeneration in MPTP-treated monkeys. Brain 2011, 134, 2057–2073. [Google Scholar] [CrossRef]
- Turle-Lorenzo, N.; Breysse, N.; Baunez, C.; Amalric, M. Functional interaction between mGlu 5 and NMDA receptors in a rat model of Parkinson’s disease. Psychopharmacology 2005, 179, 117–127. [Google Scholar] [CrossRef]
- Grégoire, L.; Morin, N.; Ouattara, B.; Gasparini, F.; Bilbe, G.; Johns, D.; Vranesic, I.; Sahasranaman, S.; Mancilla, B.G.; Paolo, T.D. The acute anti-Parkinsonian and antidyskinetic effect of AFQ056, a novel metabotropic glutamate receptor type 5 antagonist, in l-Dopa-treated parkinsonian monkeys. Parkinsonism Relat. Disord. 2011, 17, 270–276. [Google Scholar] [CrossRef]
- Bezard, E.; Pioli, E.Y.; Li, Q.; Girard, F.; Mutel, V.; Keywood, C.; Tison, F.; Rascol, O.; Poli, S.M. The mGluR5 negative allosteric modulator dipraglurant reduces dyskinesia in the MPTP macaque model. Mov. Disord. 2014, 29, 1074–1079. [Google Scholar] [CrossRef]
- Tison, F.; Keywood, C.; Wakefield, M.; Durif, F.; Corvol, J.C.; Eggert, K.; Lew, M.; Isaacson, S.; Bezard, E.; Poli, S.M.; et al. A phase 2a trial of the novel mglur5-negative allosteric modulator dipraglurant for levodopa-induced dyskinesia in Parkinson’s disease. Mov. Disord. 2016, 31, 1373–1380. [Google Scholar] [CrossRef]
- Dekundy, A.; Pietraszek, M.; Schaefer, D.; Cenci, M.A.; Danysz, W. Effects of group I metabotropic glutamate receptors blockade in experimental models of Parkinson’s disease. Brain Res. Bull. 2006, 69, 318–326. [Google Scholar] [CrossRef]
- Masilamoni, G.J.; Smith, Y. Neuroprotective Properties of Glutamate Metabotropic Glutamate Receptors in Parkinson’s Disease and Other Brain Disorders. In mGLU Receptors; Humana Press: Cham, Switzerland, 2017; pp. 103–127. [Google Scholar]
- Woolley, M.L.; Pemberton, D.J.; Bate, S.; Corti, C.; Jones, D.N.C. The mGlu2 but not the mGlu3 receptor mediates the actions of the mGluR2/3 agonist, LY379268, in mouse models predictive of antipsychotic activity. Psychopharmacology 2008, 196, 431–440. [Google Scholar] [CrossRef]
- Wang, L.; Kitai, S.T.; Xiang, Z. Modulation of excitatory synaptic transmission by endogenous glutamate acting on presynaptic group II mGluRs in rat substantia nigra compacta. J. Neurosci. Res. 2005, 82, 778–787. [Google Scholar] [CrossRef]
- Vernon, A.C.; Palmer, S.; Datla, K.P.; Zbarsky, V.; Croucher, M.J.; Dexter, D.T. Neuroprotective effects of metabotropic glutamate receptor ligands in a 6-hydroxydopamine rodent model of Parkinson’s disease. Eur. J. Neurosci. 2005, 22, 1799–1806. [Google Scholar] [CrossRef]
- Picconi, B.; Pisani, A.; Centonze, D.; Battaglia, G.; Storto, M.; Nicoletti, F.; Bernardi, G.; Calabresi, P. Striatal metabotropic glutamate receptor function following experimental parkinsonism and chronic levodopa treatment. Brain 2002, 125, 2635–2645. [Google Scholar] [CrossRef]
- Battaglia, G.; Busceti, C.L.; Molinaro, G.; Biagioni, F.; Traficante, A.; Nicoletti, F.; Bruno, V. Pharmacological activation of mGlu4 metabotropic glutamate receptors reduces nigrostriatal degeneration in mice treated with 1-methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine. J. Neurosci. 2006, 26, 7222–7229. [Google Scholar] [CrossRef]
- Iderberg, H.; Maslava, N.; Thompson, A.D.; Bubser, M.; Niswender, C.M.; Hopkins, C.R.; Lindsley, C.W.; Conn, P.J.; Jones, C.K.; Cenci, M.A. Pharmacological stimulation of metabotropic glutamate receptor type 4 in a rat model of Parkinson’s disease and L-DOPA-induced dyskinesia: Comparison between a positive allosteric modulator and an orthosteric agonist. Neuropharmacology 2015, 95, 121–129. [Google Scholar] [CrossRef]
- Panarese, J.D.; Engers, D.W.; Wu, Y.J.; Bronson, J.J.; Macor, J.E.; Chun, A. Discovery of VU2957 (valiglurax): An mGlu4 positive allosteric modulator evaluated as a preclinical candidate for the treatment of Parkinson’s disease. ACS Med. Chem. Lett. 2018, 10, 255–260. [Google Scholar] [CrossRef]
- Lopez, S.; Turle-Lorenzo, N.; Acher, F.; De, L.E.; Mele, A.; Amalric, M. Targeting group III metabotropic glutamate receptors produces complex behavioral effects in rodent models of Parkinson’s disease. J. Neurosci. 2007, 27, 6701–6711. [Google Scholar] [CrossRef]
- Stoof, J.; Booij, J.; Drukarch, B. Amantadine as N-methyl-D-aspartic acid receptor antagonist: New possibilities for therapeutic applications? Clin. Neurol. Neurosurg. 1992, 94, 4–6. [Google Scholar] [CrossRef]
- Uitti, R.J.; Rajput, A.H.; Ahlskog, J.E.; Offord, K.P.; Schroeder, D.R.; Ho, M.M.; Prasad, M.; Rajput, A.; Basran, P. Amantadine treatment is an independent predictor of improved survival in Parkinson’s disease. Neurology 1996, 46, 1551–1556. [Google Scholar] [CrossRef]
- Fox, S.H.; Metman, L.V.; Nutt, J.G.; Brodsky, M.; Factor, S.A.; Lang, A.E.; Pope, L.E.; Knowles, N.; Siffert, J. Trial of dextromethorphan/quinidine to treat levodopa-induced dyskinesia in Parkinson’s disease. Mov. Disord. 2017, 32, 893–903. [Google Scholar] [CrossRef]
- Majláth, Z.; Vécsei, L. NMDA antagonists as Parkinson’s disease therapy: Disseminating the evidence. Neurodegen. Dis. Manag. 2014, 4, 23–30. [Google Scholar] [CrossRef]
- Herring, W.J.; Assaid, C.; Budd, K.; Vargo, R.; Mazenko, R.S.; Lines, C.; Ellenbogen, A.D.; Verhagen, M.L. A phase Ib randomized controlled study to evaluate the effectiveness of a single-dose of the NR2B selective N-methyl-D-aspartate antagonist MK-0657 on levodopa-induced dyskinesias and motor symptoms in patients with Parkinson disease. Clin. Neuropharmacol. 2017, 40, 255–260. [Google Scholar] [CrossRef]
- Eggert, K.; Squillacote, D.; Barone, P.; Dodel, R.; Katzenschlager, R.; Emre, M.; Lees, A.J.; Rascol, O.; Poewe, W.; Tolosa, E.; et al. Safety and efficacy of perampanel in advanced Parkinson’s disease: A randomized, placebo-controlled study. Mov. Disord. 2010, 25, 896–905. [Google Scholar] [CrossRef]
- Lees, A.; Fahn, S.; Eggert, K.M.; Jankovic, J.; Lang, A.; Micheli, F.; Mouradian, M.M.; Oertel, W.H.; Olanow, C.W.; Poewe, W.; et al. Perampanel, an AMPA antagonist, found to have no benefit in reducing “off” time in Parkinson’s disease. Mov. Disord. 2012, 27, 284–288. [Google Scholar] [CrossRef]
- Rascol, O.; Barone, P.; Behari, M.; Emre, M.; Giladi, N.; Olanow, C.W.; Ruzicka, E.; Bibbiani, F.; Squillacote, D.; Patten, A.; et al. Perampanel in Parkinson disease fluctuations: A double-blind randomized trial with placebo and entacapone. Clin. Neuropharmacol. 2012, 35, 15–20. [Google Scholar] [CrossRef]
- Stocchi, F.; Rascol, O.; Destee, A.; Hattori, N.; Hauser, R.A.; Lang, A.E.; Poewe, W.; Stacy, M.; Tolosa, E.; Gao, H.; et al. AFQ056 in Parkinson patients with levodopa-induced dyskinesia: 13-week, randomized, dose-finding study. Mov. Disord. 2013, 28, 1838–1846. [Google Scholar] [CrossRef]
- Trenkwalder, C.; Stocchi, F.; Poewe, W.; Dronamraju, N.; Kenney, C.; Shah, A.; Raison, F.; Graf, A. Mavoglurant in Parkinson’s patients with l-Dopa-induced dyskinesias: Two randomized phase 2 studies. Mov. Disord. 2016, 31, 1054–1058. [Google Scholar] [CrossRef]
- Olanow, C.W. Can we achieve neuroprotection with currently available anti-parkinsonian interventions? Neurology 2009, 72, S59–S64. [Google Scholar] [CrossRef]
- Schapira, A.H.; Obeso, J. Timing of treatment initiation in Parkinson’s disease: A need for reappraisal? Ann. Neurol. 2006, 59, 559–562. [Google Scholar] [CrossRef]
- Kieburtz, K.; Olanow, C.W. Translational experimental therapeutics: The translation of laboratory-based discovery into disease-related therapy. Mt. Sinai J. Med. 2007, 74, 7–14. [Google Scholar] [CrossRef]
- Nuzzo, T.; Punzo, D.; Devoto, P.; Rosini, E.; Paciotti, S.; Sacchi, S.; Li, Q.; Thiolat, M.L.; Véga, C.; Carella, M.; et al. The levels of the NMDA receptor co-agonist D-serine are reduced in the substantia nigra of MPTP-lesioned macaques and in the cerebrospinal fluid of Parkinson’s disease patients. Sci. Rep. 2019, 9, 8898. [Google Scholar] [CrossRef]
- Vanle, B.; Olcott, W.; Jimenez, J.; Bashmi, L.; Danovitch, I.; IsHak, W.W. NMDA antagonists for treating the non-motor symptoms in Parkinson’s disease. Transl. Psychiatr. 2018, 8, 117. [Google Scholar] [CrossRef]
- Bennouar, K.E.; Uberti, M.A.; Melon, C.; Bacolod, M.D.; Jimenez, H.N.; Cajina, M.; Goff, L.K.; Doller, D.; Gubellini, P. Synergy between L-DOPA and a novel positive allosteric modulator of metabotropic glutamate receptor 4: Implications for Parkinson’s disease treatment and dyskinesia. Neuropharmacology 2013, 66, 158–169. [Google Scholar] [CrossRef]
- Le Poul, E.; Bolea, C.; Girard, F.; Poli, S.; Charvin, D.; Campo, B.; Bortoli, J.; Bessif, A.; Luo, B.; Koser, A.J.; et al. A potent and selective metabotropic glutamate receptor 4 positive allosteric modulator improves movement in rodent models of Parkinson’s disease. J. Pharm. Exp. Ther. 2012, 343, 167–177. [Google Scholar] [CrossRef]
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).