The Differential Expression of Mevalonate Pathway Genes in the Gut of the Bark Beetle Dendroctonus rhizophagus (Curculionidae: Scolytinae) Is Unrelated to the de Novo Synthesis of Terpenoid Pheromones

 , and
, and
Abstract
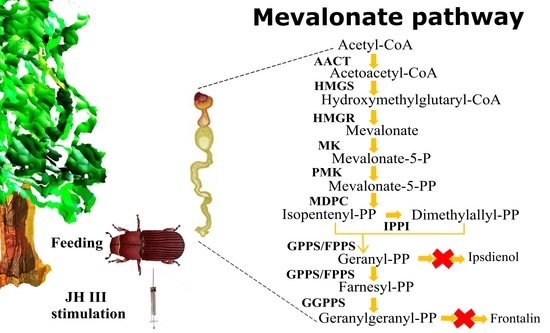
1. Introduction
2. Results
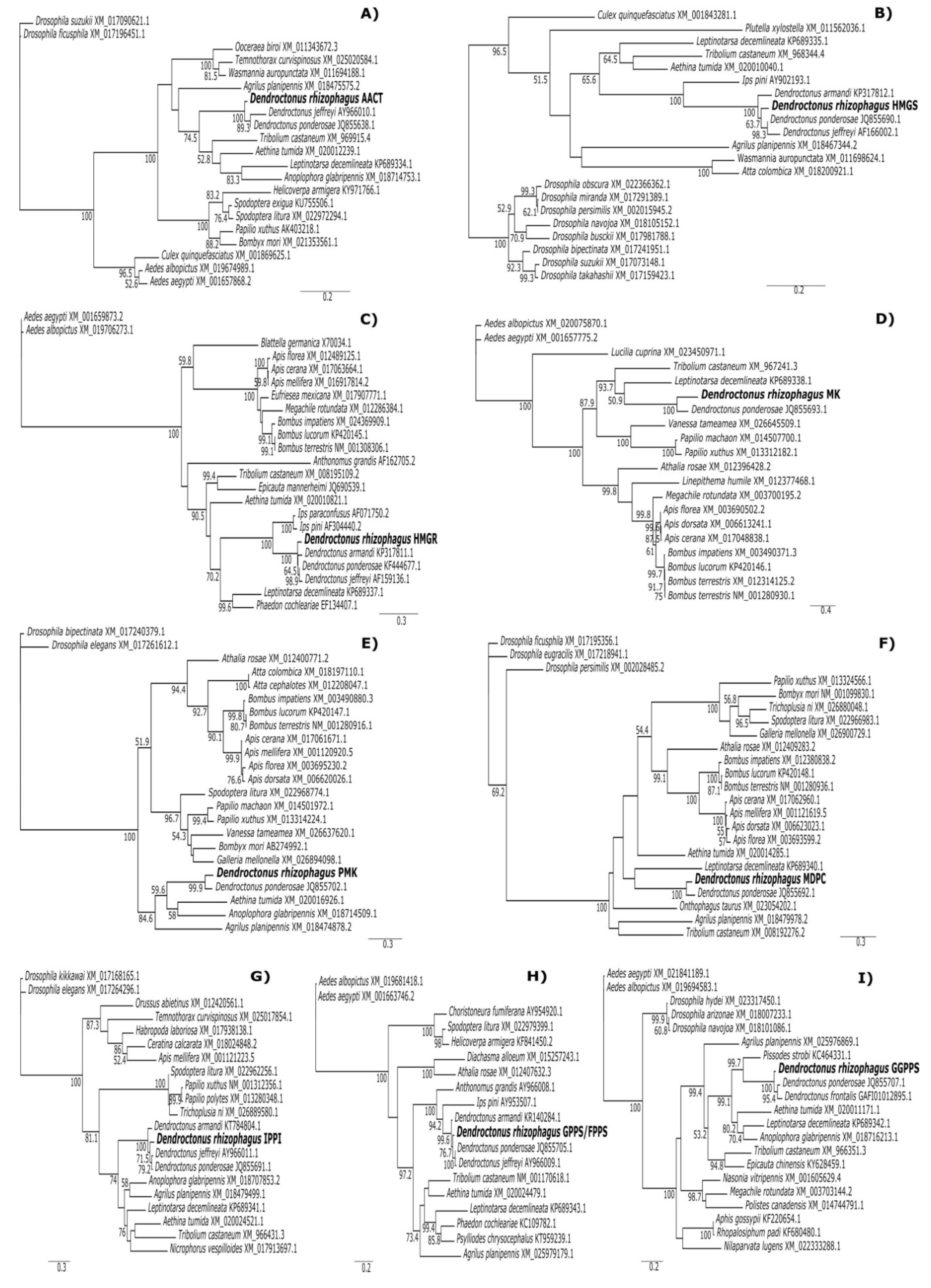
2.1. Identification of MVA Pathway Genes and Phylogenetic Analysis
2.2. Molecular Characterization of MVA Pathway Genes
2.3. Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR)
2.4. Quantification of Volatile Compounds
3. Discussion
4. Materials and Methods
4.1. Insect Collection
4.2. Treatments Aimed at Induction of MVA Pathway Genes
4.3. Total RNA Isolation and cDNA Synthesis
4.4. Amplification and Cloning of MVA Pathway cDNAs
4.5. In Silico Characterization of MVA Pathway Genes
4.6. Phylogenetic Analysis
4.7. RT-qPCR Assays
4.8. Statistical Analysis
4.9. Quantification of Volatile Compounds
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MVA | Mevalonate |
| RT-qPCR | Quantitative real-time Polymerase Chain Reaction |
| GC-MS | Gas Chromatography Mass Spectrometry |
References
- Billings, R.F.; Clarke, S.R.; Espino-Mendoza, V.; Cordón-Cabrera, P.; Meléndez-Figueroa, B.; Ramón-Campos, J.; Baeza, G. Bark beetle outbreaks and fire: A devastating combination for Central America´s pine forest. Unasylva 2004, 217, 15–21. [Google Scholar]
- Raffa, K.F.; Aukema, B.H.; Bentz, B.J.; Carroll, A.L.; Hicke, J.; Turner, M.G.; Romme, W.H. Cross-scale drivers of natural disturbances prone to anthropogenic amplification: The dynamics of bark beetle eruptions. BioScience 2008, 58, 501–517. [Google Scholar] [CrossRef]
- Borden, J.H. Aggregation pheromones. In Bark Beetles in North American Conifers: A System for the Study of Evolutionary Biology; Mitton, J.B., Sturgeon, K.M., Eds.; University of Texas Press: Austin, TX, USA, 1982; pp. 74–139. [Google Scholar]
- Tittiger, C.; Blomquist, G.J. Pheromone biosynthesis in bark beetles. Curr. Opin. Insect Sci. 2017, 24, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Seybold, S.J.; Huber, D.P.W.; Lee, J.C.; Graves, A.D.; Bohlmann, J. Pine monoterpenes and pine bark beetles: A marriage of convenience for defense and chemical communication. Phytochem. Rev. 2006, 5, 143–178. [Google Scholar] [CrossRef]
- Wyatt, T.D. Pheromones and Animal Behavior: Chemical Signals and Signatures, 2nd ed.; University Press: Cambridge, UK, 2014; pp. 65–103. [Google Scholar]
- Hunt, D.W.A.; Borden, J.H. Conversion of verbenols to verbenone by yeasts isolated from Dendroctonus ponderosae (Coleoptera: Scolytidae). J. Chem. Ecol. 1990, 16, 1385–1397. [Google Scholar] [CrossRef] [PubMed]
- Hunt, D.W.A.; Borden, J.H.; Lindgren, B.S.; Gries, G. The role of autoxidation of α-pinene in the production of pheromones of Dendroctonus ponderosae (Coleoptera: Scolytidae). Can. J. For. Res. 1989, 19, 1275–1282. [Google Scholar] [CrossRef]
- Chiu, C.C.; Keeling, C.I.; Bohlmann, J. Monoterpenyl esters in juvenile mountain pine beetle and sex-specific release of the aggregation pheromone trans-verbenol. Proc. Natl. Acad. Sci. USA 2018, 115, 3652–3657. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.C.; Keeling, C.I.; Bohlmann, J. The cytochrome P450 CYP6DE1 catalyzes the conversion of α-pinene into the mountain pine beetle aggregation pheromone trans-verbenol. Sci. Rep. 2019, 9, 1477. [Google Scholar] [CrossRef]
- Sandstrom, P.; Welch, W.H.; Blomquist, G.J.; Tittiger, C. Functional expression of a bark beetle cytochrome P450 that hydroxylates myrcene to ipsdienol. Insect Biochem. Mol. Biol. 2006, 36, 835–845. [Google Scholar] [CrossRef]
- Sandstrom, P.; Ginzel, M.D.; Bearfield, J.C.; Welch, W.H.; Blomquist, G.J.; Tittiger, C. Myrcene hydroxylases do not determine enantiomeric composition of pheromonal ipsdienol in Ips spp. J. Chem. Ecol. 2008, 34, 1584–1592. [Google Scholar] [CrossRef]
- Seybold, S.J.; Quilici, D.R.; Tillman, J.A.; Vanderwel, D.; Wood, D.L.; Blomquist, G.J. De novo biosynthesis of the aggregation pheromone components ipsenol and ipsdienol by the pine bark beetles Ips paraconfusus Lanier and Ips pini (Say)(Coleoptera: Scolytidae). Proc. Natl. Acad. Sci. USA 1995, 92, 8393–8397. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Kim, A.C.; Gorzalski, A.J.; MacLean, M.; Young, S.; Ginzel, M.D.; Blomquist, G.J.; Tittiger, C. Functional characterization of myrcene hydroxylases from two geographically distinct Ips pini populations. Insect Biochem. Mol. Biol. 2013, 43, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Gorzalski, A.; Nguyen, T.T.; Liu, X.; Jeffrey, C.; Blomquist, G.J.; Tittiger, C. exo-Brevicomin biosynthesis in the fat body of the mountain pine beetle, Dendroctonus ponderosae. J. Chem. Ecol. 2014, 40, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Byers, J.A. Chemical ecology of bark beetles. Experientia 1989, 45, 271–283. [Google Scholar] [CrossRef]
- Erbilgin, N.; Mori, S.R.; Sun, J.H.; Stein, J.D.; Owen, D.R.; Merrill, L.D.; Campos Bolaños, R.; Raffa, K.F.; Méndez Montiel, T.; Wood, D.L.; et al. Response to host volatiles by native and introduced populations of Dendroctonus valens (Coleptera: Curculionidae, Scolytinae) in North America and China. J. Chem. Ecol. 2007, 33, 131–146. [Google Scholar] [CrossRef] [PubMed]
- Kirkendall, L.R. The evolution of mating systems in bark and ambrosia beetles (Coleoptera: Scolytidae and Platypodidae). Zool. J. Linn. Soc. 1983, 77, 293–352. [Google Scholar] [CrossRef]
- Miller, D.R.; Borden, J.H. Dose-dependent and species-specific responses of pine bark beetles (Coleoptera: Scolytidae) to monoterpenes in association with pheromones. Can. Entomol. 2000, 132, 183–195. [Google Scholar] [CrossRef]
- Lombard, J.; Moreira, D. Origins and early evolution of the mevalonate pathway of isoprenoid biosynthesis in the three domains of life. Mol. Biol. Evol. 2010, 28, 87–99. [Google Scholar] [CrossRef]
- Matsumi, R.; Atomi, H.; Driessen, A.J.; van der Oost, J. Isoprenoid biosynthesis in Archaea–biochemical and evolutionary implications. Res. Microbiol. 2011, 162, 39–52. [Google Scholar] [CrossRef]
- Yeganeh, B.; Wiechec, E.; Ande, S.R.; Sharma, P.; Moghadam, A.R.; Post, M.; Freed, D.H.; Hashemi, M.; Shojaei, S.; Zeki, A.A.; et al. Targeting the mevalonate cascade as a new therapeutic approach in heart disease, cancer and pulmonary disease. Pharmacol. Ther. 2014, 143, 87–110. [Google Scholar] [CrossRef]
- Blomquist, G.J.; Figueroa-Teran, R.; Aw, M.; Song, M.; Gorzalski, A.; Abbott, N.L.; Chang, E.; Tittiger, C. Pheromone production in bark beetles. Insect Biochem. Mol. Biol. 2010, 40, 699–712. [Google Scholar] [CrossRef]
- Hall, G.M.; Tittiger, C.; Blomquist, G.J.; Andrews, G.L.; Mastick, G.S.; Barkawi, L.S.; Bengoa, C.; Seybold, S.J. Male Jeffrey pine beetle, Dendroctonus jeffreyi, synthesizes the pheromone component frontalin in anterior midgut tissue. Insect Biochem. Mol. Biol. 2002, 32, 1525–1532. [Google Scholar] [CrossRef]
- Hall, G.M.; Tittiger, C.; Andrews, G.L.; Mastick, G.S.; Kuenzli, M.; Luo, X.; Seybold, S.J.; Blomquist, G.J. Midgut tissue of male pine engraver, Ips pini, synthesizes monoterpenoid pheromone component ipsdienol de novo. Naturwissenschaften 2002, 89, 79–83. [Google Scholar] [CrossRef]
- Nardi, J.B.; Young, A.G.; Ujhelyi, E.; Tittiger, C.; Lehane, M.J.; Blomquist, G.J. Specialization of midgut cells for synthesis of male isoprenoid pheromone components in two scolytid beetles, Dendroctonus jeffreyi and Ips pini. Tissue Cell 2002, 34, 221–231. [Google Scholar] [CrossRef]
- Tittiger, C.; O’Keeffe, C.; Bengoa, C.S.; Barkawi, L.S.; Seybold, S.J.; Blomquist, G.J. Isolation and endocrine regulation of an HMG-CoA synthase cDNA from the male Jeffrey pine beetle, Dendroctonus jeffreyi (Coleoptera: Scolytidae). Insect Biochem. Mol. Biol. 2000, 30, 1203–1211. [Google Scholar] [CrossRef]
- Tittiger, C.; Barkawi, L.S.; Bengoa, C.S.; Blomquist, G.J.; Seybold, S.J. Structure and juvenile hormone-mediated regulation of the HMG-CoA reductase gene from the Jeffrey pine beetle, Dendroctonus jeffreyi. Mol. Cell Endocrinol. 2003, 199, 11–21. [Google Scholar] [CrossRef]
- Tillman, J.A.; Holbrook, G.L.; Dallara, P.L.; Schal, C.; Wood, D.L.; Blomquist, G.J. Endocrine regulation of de novo aggregation pheromone biosynthesis in the pine engraver, Ips pini (Say) (Coleoptera: Scolytidae). Insect Biochem. Mol. Biol. 1998, 28, 705–715. [Google Scholar] [CrossRef]
- Bellés, X.; Martín, D.; Piulachs, M.D. The mevalonate pathway and the synthesis of juvenile hormone in insects. Annu. Rev. Entomol. 2005, 50, 181–199. [Google Scholar] [CrossRef]
- Miziorko, H.M. Enzymes of the mevalonate pathway of isoprenoid biosynthesis. Arch. Biochem. Biophys. 2011, 505, 131–143. [Google Scholar] [CrossRef]
- Keeling, C.I.; Blomquist, G.J.; Tittiger, C. Coordinated gene expression for pheromone biosynthesis in the pine engraver beetle, Ips pini (Coleoptera: Scolytidae). Naturwissenschaften 2004, 91, 324–328. [Google Scholar] [CrossRef]
- Frick, S.; Nagel, R.; Schmidt, A.; Bodemann, R.R.; Rahfeld, P.; Pauls, G.; Brandt, W.; Gershenzon, J.; Boland, W.; Burse, A. Metal ions control product specificity of isoprenyl diphosphate synthases in the insect terpenoid pathway. Proc. Natl. Acad. Sci. USA 2013, 110, 4194–4199. [Google Scholar] [CrossRef]
- Sen, S.E.; Brown, D.C.; Sperry, A.E.; Hitchcock, J.R. Prenyltransferase of larval and adult M. sexta corpora allata. Insect Biochem. Mol. Biol. 2007, 37, 29–40. [Google Scholar] [CrossRef]
- Vandermoten, S.; Charloteaux, B.; Santini, S.; Sen, S.E.; Béliveau, C.; Vandenbol, M.; Francis, F.; Brasseur, R.; Cusson, M.; Haubruge, E. Characterization of a novel aphid prenyltransferase displaying dual geranyl/farnesyl diphosphate synthase activity. FEBS Lett. 2008, 582, 1928–1934. [Google Scholar] [CrossRef][Green Version]
- Keeling, C.I.; Chiu, C.C.; Aw, T.; Li, M.; Henderson, H.; Tittiger, C.; Weng, H.B.; Blomquist, G.J.; Bohlmann, J. Frontalin pheromone biosynthesis in the mountain pine beetle, Dendroctonus ponderosae, and the role of isoprenyl diphosphate synthases. Proc. Natl. Acad. Sci. USA 2013, 110, 18838–18843. [Google Scholar] [CrossRef]
- Mendoza, M.G.; Salinas-Moreno, Y.; Olivo-Martínez, A.; Zúñiga, G. Factors influencing the geographical distribution of Dendroctonus rhizophagus (Coleoptera: Curculionidae: Scolytinae) in the Sierra Madre Occidental México. Environ. Entomol. 2011, 40, 549–559. [Google Scholar] [CrossRef]
- Sánchez-Martínez, G.; Wagner, M.R. Host preference and attack pattern of Dendroctonus rhizophagus (Coleoptera: Curculionidae: Scolytinae): A bark beetle specialist on pine regeneration. Environ. Entomol. 2009, 38, 1197–1204. [Google Scholar] [CrossRef]
- Cibrián-Tovar, D.; Méndez-Montiel, J.T.; Campos-Bolaños, R.; Yates III, H.O.; Flores-Lara, J. Insectos Forestales de México/Forests Insects of México COFAN/NAFC; Universidad Autónoma Chapingo: Texcoco, Mexico, 1995. [Google Scholar]
- Cano-Ramírez, C.; Armendáriz-Toledano, F.; Macías-Sámano, J.E.; Sullivan, B.T.; Zúñiga, G. Electrophysiological and behavioral responses of the bark beetle Dendroctonus rhizophagus to volatiles from host pines and conspecifics. J. Chem. Ecol. 2012, 38, 512–524. [Google Scholar] [CrossRef]
- Niño-Domínguez, A.; Sullivan, B.T.; López-Urbina, J.H.; Macías-Sámano, J.E. Pheromone-mediated mate location and discrimination by two syntopic sibling species of Dendroctonus bark beetles in Chiapas, Mexico. J. Chem. Ecol. 2015, 41, 746–756. [Google Scholar] [CrossRef]
- Symonds, M.R.E.; Gitau-Clarke, C.W. The evolution of aggregation pheromone diversity in bark beetles. In Advances in Insect Physiology; Tittiger, C., Blomquist, G.J., Eds.; Academic Press: New York, NY, USA, 2016; Volume 50, pp. 195–234. [Google Scholar]
- Kinzer, G.W.; Fentiman, A.F., Jr.; Page, T.F., Jr.; Foltz, R.L.; Vité, J.P.; Pitman, G.B. Bark beetle attractants: Identification, synthesis and field bioassay of a new compound isolated from Dendroctonus. Nature 1969, 221, 477. [Google Scholar] [CrossRef]
- Pureswaran, D.S.; Gries, R.; Borden, J.H.; Pierce, H.D., Jr. Dynamics of pheromone production and communication in the mountain pine beetle, Dendroctonus ponderosae Hopkins, and the pine engraver, Ips pini (Say) (Coleoptera: Scolytidae). Chemoecology 2000, 10, 153–168. [Google Scholar] [CrossRef]
- Rudinsky, J.A.; Morgan, M.E.; Libbey, L.M.; Putnam, T.B. Antiaggregative-rivalry pheromone of the mountain pine beetle, and a new arrestant of the southern pine beetle. Environ. Entomol. 1974, 3, 90–98. [Google Scholar] [CrossRef]
- Liu, Z.; Xu, B.; Miao, Z.; Sun, J. The pheromone frontalin and its dual function in the invasive bark beetle Dendroctonus valens. Chem. Senses. 2013, 38, 485–495. [Google Scholar] [CrossRef]
- Bearfield, J.C.; Henry, A.G.; Tittiger, C.; Blomquist, G.J.; Ginzel, M.D. Two regulatory mechanisms of monoterpenoid pheromone production in Ips spp. of bark beetles. J. Chem. Ecol. 2009, 35, 689–697. [Google Scholar] [CrossRef]
- Yu, J.; Dai, L.; Zhang, R.; Li, Z.; Pham, T.; Chen, H. Isolation and expression of HMG-CoA synthase and HMG-CoA reductase genes in different development stages, tissues and treatments of the Chinese white pine beetle, Dendroctonus armandi (Curculionidae: Scolytinae). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2015, 187, 62–70. [Google Scholar] [CrossRef]
- Barkawi, L.S.; Francke, W.; Blomquist, G.J.; Seybold, S.J. Frontalin: De novo biosynthesis of an aggregation pheromone component by Dendroctonus spp. bark beetles (Coleoptera: Scolytidae). Insect Biochem. Mol. Biol. 2003, 33, 773–788. [Google Scholar] [CrossRef]
- Fukao, T.; Song, X.Q.; Mitchell, G.A.; Yamaguchi, S.; Sukegawa, K.; Orii, T.; Kondo, N. Enzymes of ketone body utilization in human tissues: Protein and messenger RNA levels of succinyl-coenzyme A (CoA): 3-ketoacid CoA transferase and mitochondrial and cytosolic acetoacetyl-CoA thiolases. Pediatr. Res. 1997, 42, 498–502. [Google Scholar] [CrossRef]
- Fujii, T.; Ito, K.; Katsuma, S.; Nakano, R.; Shimada, T.; Ishikawa, Y. Molecular and functional characterization of an acetyl-CoA acetyltransferase from the adzuki bean borer moth Ostrinia scapulalis (Lepidoptera: Crambidae). Insect Biochem. Mol. Biol. 2010, 40, 74–78. [Google Scholar] [CrossRef]
- Mascaró, C.; Buesa, C.; Ortiz, J.A.; Haro, D.; Hegardt, F.G. Molecular cloning and tissue expression of human mitochondrial 3-hydroxy-3-methylglutaryl-CoA synthase. Arch. Biochem. Biophys. 1995, 317, 385–390. [Google Scholar] [CrossRef]
- Ayté, J.; Gil-Gómez, G.; Haro, D.; Marrero, P.F.; Hegardt, F.G. Rat mitochondrial and cytosolic 3-hydroxy-3-methylglutaryl-CoA synthases are encoded by two different genes. Proc. Natl. Acad. Sci. USA 1990, 87, 3874–3878. [Google Scholar] [CrossRef]
- Martínez-González, J.; Buesa, C.; Piulachs, M.D.; Belles, X.; Hegardt, F.G. 3-Hydroxy-3-methylglutaryl-coenzyme—A synthase from Blattella germanica: Cloning, expression, developmental pattern and tissue expression. Eur. J. Biochem. 1993, 217, 691–699. [Google Scholar] [CrossRef]
- Couillaud, F.; Feyereisen, R. Assay of HMG-CoA synthase in Diploptera punctata corpora allata. Insect Biochem. 1991, 21, 131–135. [Google Scholar] [CrossRef]
- Jingami, H.; Brown, M.S.; Goldstein, J.L.; Anderson, R.G.W.; Luskey, K.L. Partial deletion of membrane-bound domain of 3-hydroxy-3-methylglutaryl coenzyme A reductase eliminates sterol-enhanced degradation and prevents formation of crystalloid endoplasmic reticulum. J. Cell Biol. 1987, 104, 1693–1704. [Google Scholar] [CrossRef]
- Liscum, L.; Finer-Moore, J.; Stroud, R.M.; Luskey, K.L.; Brown, M.S.; Goldstein, J.L. Domain structure of 3-hydroxy-3-methylglutaryl coenzyme A reductase, a glycoprotein of the endoplasmic reticulum. J. Biol. Chem. 1985, 260, 522–530. [Google Scholar]
- Feyereisen, R.; Farnsworth, D.E. Characterization and regulation of HMG-CoA reductase during a cycle of juvenile hormone synthesis. Mol. Cell Endocrinol. 1987, 53, 227–238. [Google Scholar] [CrossRef]
- Barnes, F.J.; Goodfellow, R.D. Mevalonate kinase: Localization and variation in activity during the development of Sarcophaga bullata. J. Insect Physiol. 1971, 17, 1415–1427. [Google Scholar] [CrossRef]
- Hogenboom, S.; Tuyp, J.J.M.; Espeel, M.; Koster, J.; Wanders, R.J.A.; Waterham, H.R. Phosphomevalonate kinase is a cytosolic protein in humans. J. Lipid Res. 2004, 45, 697–705. [Google Scholar] [CrossRef]
- Biardi, L.; Krisans, S.K. Compartmentalization of Cholesterol Biosynthesis Conversion of mevalonate to farnesyl diphosphate occurs in the peroxisomes. J. Biol. Chem. 1996, 271, 1784–1788. [Google Scholar] [CrossRef]
- Olivier, L.M.; Chambliss, K.L.; Gibson, K.M.; Krisans, S.K. Characterization of phosphomevalonate kinase: Chromosomal localization, regulation, and subcellular targeting. J. Lipid Res. 1999, 40, 672–679. [Google Scholar]
- Ericsson, J.; Runquist, M.; Thelin, A.; Andersson, M.; Chojnacki, T.; Dallner, G. Distribution of prenyltransferases in rat tissues. Evidence for a cytosolic all-trans-geranylgeranyl diphosphate synthase. J. Biol. Chem. 1993, 268, 832–838. [Google Scholar]
- Krisans, S.K.; Ericsson, J.; Edwards, P.A.; Keller, G.A. Farnesyl-diphosphate synthase is localized in peroxisomes. J. Biol. Chem. 1994, 269, 14165–14169. [Google Scholar]
- Martín, D.; Piulachs, M.D.; Cunillera, N.; Ferrer, A.; Bellés, X. Mitochondrial targeting of farnesyl diphosphate synthase is a widespread phenomenon in eukaryotes. Biochim. Biophys. Acta. 2007, 1773, 419–426. [Google Scholar] [CrossRef]
- Paton, V.G.; Shackelford, J.E.; Krisans, S.K. Cloning and subcellular localization of hamster and rat isopentenyl diphosphate dimethylallyl diphosphate isomerase. A PTS1 motif targets the enzyme to peroxisomes. J. Biol. Chem. 1997, 272, 18945–18950. [Google Scholar] [CrossRef]
- Bearfield, J.C.; Keeling, C.I.; Young, S.; Blomquist, G.J.; Tittiger, C. Isolation, endocrine regulation and mRNA distribution of the 3-hydroxy-3-methylglutaryl coenzyme A synthase (HMG-S) gene from the pine engraver, Ips pini (Coleoptera: Scolytidae). Insect Mol. Biol. 2006, 15, 187–195. [Google Scholar] [CrossRef]
- Tittiger, C.; Blomquist, G.J.; Ivarsson, P.; Borgeson, C.E.; Seybold, S.J. Juvenile hormone regulation of HMG-R gene expression in the bark beetle Ips paraconfusus (Coleoptera: Scolytidae): Implications for male aggregation pheromone biosynthesis. Cell Mol. Life Sci. 1999, 55, 121–127. [Google Scholar] [CrossRef]
- Goldstein, J.L.; Brown, M.S. Regulation of the mevalonate pathway. Nature. 1990, 343, 425–430. [Google Scholar] [CrossRef]
- Keeling, C.I.; Bearfield, J.C.; Young, S.; Blomquist, G.J.; Tittiger, C. Effects of juvenile hormone on gene expression in the pheromone-producing midgut of the pine engraver beetle, Ips pini. Insect Mol. Biol. 2006, 15, 207–216. [Google Scholar] [CrossRef]
- Aw, T.; Schlauch, K.; Keeling, C.I.; Young, S.; Bearfield, J.C.; Blomquist, G.J.; Tittiger, C. Functional genomics of mountain pine beetle (Dendroctonus ponderosae) midguts and fat bodies. BMC Genom. 2010, 11, 215. [Google Scholar] [CrossRef]
- Burg, J.S.; Espenshade, P.J. Regulation of HMG-CoA reductase in mammals and yeast. Prog. Lipid Res. 2011, 50, 403–410. [Google Scholar] [CrossRef]
- Burg, J.S.; Powell, D.W.; Chai, R.; Hughes, A.L.; Link, A.J.; Espenshade, P.J. Insig regulates HMG-CoA reductase by controlling enzyme phosphorylation in fission yeast. Cell Metab. 2008, 8, 522–531. [Google Scholar] [CrossRef]
- Li, H.; Wang, H.; Hackett, M.; Schooley, D.A. The structure of dolichols isolated from Manduca sexta larvae. Insect Biochem. Mol. Biol. 1995, 25, 1019–1026. [Google Scholar] [CrossRef]
- Lindorfer, M.A.; Sherman, N.E.; Woodfork, K.A.; Fletcher, J.E.; Hunt, D.F.; Garrison, J.C. G protein γ subunits with altered prenylation sequences are properly modified when expressed in Sf9 cells. J. Biol. Chem. 1996, 271, 18582–18587. [Google Scholar] [CrossRef]
- Silberkang, M.; Havel, C.M.; Friend, D.S.; McCarthy, B.J.; Watson, J.A. Isoprene synthesis in isolated embryonic Drosophila cells. I. Sterol-deficient eukaryotic cells. J. Biol. Chem. 1983, 258, 8503–8511. [Google Scholar]
- Gilg, A.B.; Bearfield, J.C.; Tittiger, C.; Welch, W.H.; Blomquist, G.J. Isolation and functional expression of an animal geranyl diphosphate synthase and its role in bark beetle pheromone biosynthesis. Proc. Natl. Acad. Sci. USA 2005, 102, 9760–9765. [Google Scholar] [CrossRef]
- Gilg, A.B.; Tittiger, C.; Blomquist, G.J. Unique animal prenyltransferase with monoterpene synthase activity. Naturwissenschaften 2009, 96, 731–735. [Google Scholar] [CrossRef]
- Zhao, M.; Dai, L.; Fu, D.; Gao, J.; Chen, H. Electrophysiological and behavioral responses of Dendroctonus armandi (Coleoptera: Curculionidae: Scolytinae) to two candidate pheromone components: Frontalin and exo-brevicomin. Chemoecology 2017, 27, 91–99. [Google Scholar] [CrossRef]
- Grégoire, J.C. The greater European spruce beetle. In Dynamics of Forest Insect Populations, 1st ed.; Springer: Boston, MA, USA, 1988; pp. 455–478. [Google Scholar]
- Furniss, M.M. Biology of Dendroctonus punctatus (Coleoptera: Scolytidae). Ann. Entomol. Soc. Am. 1995, 88, 173–182. [Google Scholar] [CrossRef]
- Furniss, R.L.; Carolin, V.M. Western Forest Insects; No. 1339; USDA Forest Service, Miscellaneous Publication: Washington, DC, USA, 1977.
- Wood, S.L. Tribe Tomicini. Genus Dendroctonus Erichson. In The Bark and Ambrosia Beetles of North and Central America (Coleoptera: Scolytidae), a Taxonomic Monograph; Brigham Young University: Provo, UT, USA, 1982; pp. 150–203. [Google Scholar]
- Zúñiga, G. Personal Communication; Instituto Politécnico Nacional: Mexico City, Mexico, 2019. [Google Scholar]
- Zúñiga, G.; Correa, G.M.; Cisneros, R.; Moreno, Y.S. Zonas de sobreposición en las áreas de distribución geográfica de las especies mexicanas de Dendroctonus Erichson (Coleoptera: Scolytidae) y sus implicaciones ecológico-evolutivas. Acta Zool. Mex. 1999, 77, 1–22. [Google Scholar] [CrossRef]
- Byers, J.A. Bark beetle conversion of a plant compound to a sex-specific inhibitor of pheromone attraction. Science 1983, 220, 624–626. [Google Scholar] [CrossRef]
- Renwick, J.A.; Hughes, P.R.; Krull, I.S. Selective production of cis-and trans-verbenol from (-)-and (+)-alpha by a bark beetle. Science 1976, 191, 199–201. [Google Scholar] [CrossRef]
- Francke, W.; Vité, J.P. Oxygenated terpenes in pheromone systems of bark beetles. Z. Angew. Entomol. 1983, 96, 146–156. [Google Scholar] [CrossRef]
- Armendáriz-Toledano, F.; Zúñiga, G. Illustrated Key to Species of Genus Dendroctonus (Coleoptera: Curculionidae) Occurring in Mexico and Central America. J. Insect Sci. 2017, 17, 1–15. [Google Scholar] [CrossRef]
- Cano-Ramírez, C.; López, M.F.; Cesar-Ayala, A.K.; Pineda-Martínez, V.; Sullivan, B.T.; Zúñiga, G. Isolation and expression of cytochrome P450 genes in the antennae and gut of pine beetle Dendroctonus rhizophagus (Curculionidae: Scolytinae) following exposure to host monoterpenes. Gene 2013, 520, 47–63. [Google Scholar] [CrossRef]
- López, M.F.; Cano-Ramírez, C.; Cesar-Ayala, A.K.; Ruiz, E.A.; Zúñiga, G. Diversity and expression of P450 genes from Dendroctonus valens LeConte (Curculionidae: Scolytinae) in response to different kairomones. Insect Biochem. Mol. Biol. 2013, 43, 417–432. [Google Scholar] [CrossRef]
- Torres-Banda, V.; López, M.F.; Obregón-Molina, G.; Zuñiga, G. Identification and Comparison of Genes Associated with Detoxification Process between Two Sibling Bark Beetle Species by Intestine-Specific Transcriptome Profile Analysis. Unpublished work. 2019. [Google Scholar]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Artimo, P.; Jonnalagedda, M.; Arnold, K.; Baratin, D.; Csardi, G.; de Castro, E.; Duvaud, S.; Flegel, V.; Fortier, A.; Gasteiger, E.; et al. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res. 2012, 40, W597–W603. [Google Scholar] [CrossRef]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server. In The Proteomics Protocols Handbook; Walker, J.M., Ed.; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar]
- Emanuelsson, O.; Nielsen, H.; Brunak, S.; von Heijne, G. Predicting subcellular localization of proteins based on their N-terminal amino acid sequence. J. Mol. Biol. 2000, 300, 1005–1016. [Google Scholar] [CrossRef]
- Sonnhammer, E.L.; Von Heijne, G.; Krogh, A. A hidden Markov model for predicting transmembrane helices in protein sequences. In Sixth International Conference on Intelligent Systems for Molecular Biology; Glasgow, J., Littlejohn, T., Major, F., Lathrop, R., Sankoff, D., Sensen, C., Eds.; AAAI Press: Menlo Park, CA, USA, 1998. [Google Scholar]
- Gouet, P.; Robert, X.; Courcelle, E. ESPript/ENDscript: Extracting and rendering sequence and 3D information from atomic structures of proteins. Nucleic Acids Res. 2003, 31, 3320–3323. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Abascal, F.; Zardoya, R.; Posada, D. ProtTest: Selection of best-fit models of protein evolution. Bioinformatics 2005, 21, 2104–2105. [Google Scholar] [CrossRef]
- Lefort, V.; Longueville, J.E.; Gascuel, O. SMS: Smart model selection in PhyML. Mol. Biol. Evol. 2017, 34, 2422–2424. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Dai, L.; Ma, M.; Wang, C.; Shi, Q.; Zhang, R.; Chen, H. Cytochrome P450s from the Chinese white pine beetle, Dendroctonus armandi (Curculionidae: Scolytinae): Expression profiles of different stages and responses to host allelochemicals. Insect Biochem. Mol. Biol. 2015, 65, 35–46. [Google Scholar] [CrossRef]
- Dai, L.; Ma, M.; Gao, G.; Chen, H. Dendroctonus armandi (Curculionidae: Scolytinae) cytochrome P450s display tissue specificity and responses to host terpenoids. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2016, 201, 1–11. [Google Scholar] [CrossRef]
- Obregón-Molina, G.; Cesar-Ayala, A.K.; López, M.F.; Cano-Ramírez, C.; Zúñiga, G. Comparison of orthologous cytochrome P450 genes relative expression patterns in the bark beetles Dendroctonus rhizophagus and Dendroctonus valens (Curculionidae: Scolytinae) during host colonization. Insect Mol. Biol. 2015, 24, 649–661. [Google Scholar] [CrossRef]
- Sullivan, B.T. Electrophysiological and behavioral responses of Dendroctonus frontalis (Coleoptera: Curculionidae) to volatiles isolated from conspecifics. J. Econ. Entomol. 2005, 98, 2067–2078. [Google Scholar] [CrossRef]
- Sirinupong, N.; Suwanmanee, P.; Doolittle, R.F.; Suvachitanont, W. Molecular cloning of a new cDNA and expression of 3-hydroxy-3-methylglutaryl-CoA synthase gene from Hevea brasiliensis. Planta 2005, 221, 502–512. [Google Scholar] [CrossRef]
- Andrade-Pavón, D.; Cuevas-Hernández, R.I.; Trujillo-Ferrara, J.G.; Hernández-Rodríguez, C.; Ibarra, J.A.; Villa-Tanaca, L. Recombinant 3-hydroxy 3-methyl glutaryl-CoA reductase from Candida glabrata (Rec-CgHMGR) obtained by heterologous expression, as a novel therapeutic target model for testing synthetic drugs. Appl. Biochem. Biotechnol. 2017, 182, 1478–1490. [Google Scholar] [CrossRef]
- Potter, D.; Miziorko, H.M. Identification of catalytic residues in human mevalonate kinase. J. Biol. Chem. 1997, 272, 25449–25454. [Google Scholar] [CrossRef]
- Schafer, B.L.; Bishop, R.W.; Kratunis, V.J.; Kalinowski, S.S.; Mosley, S.T.; Gibson, K.M.; Tanaka, R.D. Molecular cloning of human mevalonate kinase and identification of a missense mutation in the genetic disease mevalonic aciduria. J. Biol. Chem. 1992, 267, 13229–13238. [Google Scholar]
- Yang, D.; Shipman, L.W.; Roessner, C.A.; Scott, A.I.; Sacchettini, J.C. Structure of the Methanococcus jannaschii mevalonate kinase, a member of the GHMP kinase superfamily. J. Biol. Chem. 2002, 277, 9462–9467. [Google Scholar] [CrossRef]
- Herdendorf, T.J.; Miziorko, H.M. Phosphomevalonate kinase: Functional investigation of the recombinant human enzyme. Biochemistry 2006, 45, 3235–3242. [Google Scholar] [CrossRef]
- Smit, A.; Mushegian, A. Biosynthesis of isoprenoids via mevalonate in Archaea: The lost pathway. Genome Res. 2000, 10, 1468–1484. [Google Scholar] [CrossRef]
- Byres, E.; Alphey, M.S.; Smith, T.K.; Hunter, W.N. Crystal structures of Trypanosoma brucei and Staphylococcus aureus mevalonate diphosphate decarboxylase inform on the determinants of specificity and reactivity. J. Mol. Biol. 2007, 371, 540–553. [Google Scholar] [CrossRef]
- Krepkiy, D.; Miziorko, H.M. Identification of active site residues in mevalonate diphosphate decarboxylase: Implications for a family of phosphotransferases. Protein Sci. 2004, 13, 1875–1881. [Google Scholar] [CrossRef]
- Chen, R.; Harada, Y.; Bamba, T.; Nakazawa, Y.; Gyokusen, K. Overexpression of an isopentenyl diphosphate isomerase gene to enhance trans-polyisoprene production in Eucommia ulmoides Oliver. BMC Biotechnol. 2012, 12, 78. [Google Scholar] [CrossRef]
- Diaz, M.E.; Mayoral, J.G.; Priestap, H.; Nouzova, M.; Rivera-Perez, C.; Noriega, F.G. Characterization of an isopentenyl diphosphate isomerase involved in the juvenile hormone pathway in Aedes aegypti. Insect Biochem. Mol. Biol. 2012, 42, 751–757. [Google Scholar] [CrossRef]
- Chen, A.; Kroon, P.A.; Poulter, C.D. Isoprenyl diphosphate synthases: Protein sequence comparisons, a phylogenetic tree, and predictions of secondary structure. Protein Sci. 1994, 3, 600–607. [Google Scholar] [CrossRef]
- Thabet, I.; Guirimand, G.; Guihur, A.; Lanoue, A.; Courdavault, V.; Papon, N.; Bouzid, S.; Giglioli-Guivarc’h, N.; Simkin, A.J.; Clastre, M. Characterization and subcellular localization of geranylgeranyl diphosphate synthase from Catharanthus roseus. Mol. Biol. Rep. 2012, 39, 3235–3243. [Google Scholar] [CrossRef]
- Zhu, X.; Suzuki, K.; Saito, T.; Okada, K.; Tanaka, K.; Nakagawa, T. Geranylgeranyl pyrophosphate synthase encoded by the newly isolated gene GGPS6 from Arabidopsis thaliana is localized in mitochondria. Plant Mol. Biol. 1997, 35, 331–341. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Species | Blast NCBI GenBank No. | % Similarity | Blast UniProt UniProt No. | % Similarity |
|---|---|---|---|---|---|
| AACT | Dendroctonus ponderosae | JQ855638.1 | 93 | I1VJ17 | 97.6 |
| Dendroctonus jeffreyi | AY966010.1 | 92 | Q56CY6 | 91.5 | |
| Leptinotarsa decemlineata | KP689334.1 | 72 | A0A0H4IRH6 | 81.1 | |
| Tribolium castaneum | XM_969915.4 | 72 | D6WAN7 | 79.9 | |
| HMGS | Dendroctonus ponderosae | JQ855690.1 | 95 | I1VJ69 | 96.7 |
| Dendroctonus jeffreyi | AF166002.1 | 94 | Q9NDA8 | 93.7 | |
| Dendroctonus armandi | KP317812.1 | 92 | A0A0F7LEZ1 | 94.5 | |
| Ips pini | AY902193.1 | 72 | Q5EE42 | 75.9 | |
| Leptinotarsa decemlineata | KP689335.1 | 71 | A0A0H4J592 | 67.6 | |
| Tribolium castaneum | XM_968344.4 | 71 | D6WTE1 | 65.9 | |
| HMGR | Dendroctonus ponderosae | KF444677.1 | 96 | U5ZZ40 | 96.8 |
| Dendroctonus jeffreyi | AF159136.1 | 95 | Q9N6G8 | 96.3 | |
| Dendroctonus armandi | KP317811.1 | 93 | A0A0F7LG51 | 95.4 | |
| Ips paraconfusus | AF071750.2 | 75 | Q9XY99 | 78.5 | |
| Ips pini | AF304440.1 | 74 | Q95WT1 | 78.1 | |
| Leptinotarsa decemlineata | KP689337.1 | 72 | A0A0H4ISG3 | 66.6 | |
| Tribolium castaneum | XM_008195109.2 | 69 | A0A139WIA9 | 68.3 | |
| MK | Dendroctonus ponderosae | JQ855693.1 | 83 | I1VJ72 | 69.8 |
| Leptinotarsa decemlineata | KP689338.1 | - | A0A0H4IUW0 | 41.8 | |
| PMK | Dendroctonus ponderosae | JQ855702.1 | 91 | I1VJ81 | 89.1 |
| Leptinotarsa decemlineata | KP689339.1 | 68 | A0A0H4IRI1 | 59.2 | |
| MDPC | Dendroctonus ponderosae | JQ855692.1 | 93 | I1VJ71 | 92 |
| Leptinotarsa decemlineata | KP689340.1 | 68 | A0A0H4J596 | 61.6 | |
| Tribolium castaneum | XM_008192276.2 | 66 | D6WE42 | 64.4 | |
| IPPI | Dendroctonus ponderosae | JQ855691.1 | 94 | I1VJ70 | 92.9 |
| Dendroctonus jeffreyi | AY966011.1 | 94 | Q56CY5 | 92.9 | |
| Dendroctonus armandi | KT784804.1 | 86 | A0A0N9H5S7 | 89.6 | |
| Leptinotarsa decemlineata | KP689341.1 | 71 | A0A0H4IVW7 | 70.1 | |
| Tribolium castanem | XM_966431.3 | 65 | D6WP58 | 76.6 | |
| GPPS/FPPS | Dendroctonus ponderosae | JQ855705.1 | 95 | I1VJ84 | 96.3 |
| Dendroctonus jeffreyi | AY966009.1 | 94 | Q56CY7 | 94.9 | |
| Dendroctonus armandi | KR140284.1 | 93 | A0A0N9EI79 | 96 | |
| Ips pini | AY953507.1 | 72 | Q58GE9 | 72.1 | |
| Tribolium castanem | NM_001170618.1 | 70 | D6WSE5 | 70.4 | |
| Leptinotarsa decemlineata | KP689343.1 | 66 | A0A0H4IUW5 | 55.5 | |
| GGPPS | Dendroctonus ponderosae | JQ855707.1 | 94 | I1VJ86 | 95.3 |
| Dendroctonus frontalis | GAFI01012895.1 | 93 | T1DSZ4 | 94.6 | |
| Leptinotarsa decemlineata | KP689342.1 | 72 | A0A0H4ISG7 | 75.7 | |
| Tribolium castaneum | XM_966351.3 | 71 | D6WD18 | 68.3 |
| Protein | ORF 1 (bp) Accession No. | Aa 2 | M.W. 3 (kDa) | pI 3 | Subcellular Localization Predictions | Secondary Structure Elements | |||
|---|---|---|---|---|---|---|---|---|---|
| TargetP | ProtComp 9.0 | TMH 5 | α-H 6 | β-S 6 | |||||
| AACT | 1239 MK387135 | 412 | 43.2 | 8.64 | 0.458 mTP 4, 0.036 SP 4, 0.441 Other | Mitochondrial | No TMH | 11 | 16 |
| HMGS | 1374 MK387139 | 457 | 50.7 | 6.04 | 0.017 mTP, 0.643 SP, 0.648 Other | Cytoplasmic | No TMH | 15 | 17 |
| HMGR | 2550 MK387137 | 849 | 93.4 | 8.07 | 0.124 mTP, 0.471 SP, 0.398 Other | Membrane bound Endoplasmic Reticulum | 5 TMH | 16 | 11 |
| MK | 1308 MK387142 | 435 | 48.3 | 6.17 | 0.497 mTP, 0.046 SP, 0.579 Other | Cytoplasmic | No TMH | 11 | 13 |
| PMK | 558 MK387143 | 185 | 21.3 | 6.89 | 0.092 mTP, 0.077 SP, 0.881 Other | Peroxisomal | No TMH | 8 | 5 |
| MDPC | 1161 MK387140 | 386 | 42.9 | 7.01 | 0.058 mTP, 0.275 SP, 0.748 Other | Extracellular | No TMH | 11 | 17 |
| IPPI | 759 MK387141 | 252 | 28.9 | 6.31 | 0.348 mTP, 0.133 SP, 0.404 Other | Cytoplasmic | No TMH | 8 | 9 |
| GPPS/FPPS | 1290 MK387138 | 429 | 49.5 | 8.7 | 0.778 mTP, 0.063 SP, 0.223 Other | Cytoplasmic/Mitochondrial | No TMH | 16 | 2 |
| GGPPS | 894 MK387136 | 297 | 34.3 | 6.33 | 0.108 mTP, 0.091 SP, 0.855 Other | Cytoplasmic | No TMH | 15 | 2 |
| Compound | Female | Male |
|---|---|---|
| fenchyl alcohol | 0.01–0.04 1 2/12 2 | - 0/12 |
| cis-verbenol | - 0/12 | - 0/12 |
| trans-verbenol | - 0/12 | - 0/12 |
| myrtenal | - 0/12 | 0.007 1/12 |
| myrtenol | - 0/12 | 0.03–0.1 3/12 |
| verbenone | 0.002–0.003 2/12 | 0.002–0.05 3/12 |
| Gene | Species | Male | Female | Technique | Treatment | Reference |
|---|---|---|---|---|---|---|
| AACT | I. pini | U | U* | RT-qPCR | Feeding | [32] |
| I. pini | C | U* | RT-qPCR | JH III | [70] | |
| HMGS | I. pini | U | U* | RT-qPCR | Feeding | [32] |
| I. pini | U | U | RT-qPCR | JH III | [70] | |
| I. pini | U* | U | RT-qPCR | JH III | [67] | |
| D. jeffreyi | U* | C | Northern blot | JH III | [27] | |
| D. armandi | U* | U | RT-qPCR | Feeding | [48] | |
| D. armandi | U* | C | RT-qPCR | JH III | [48] | |
| I. confusus | U | NR | RT-qPCR | Feeding | [47] | |
| I. confusus | U | NR | RT-qPCR | JH III | [47] | |
| HMGR | I. paraconfusus | U* | U | Northern blot | JH III | [68] |
| I. pini | U* | U | Northern blot | JH III | [25] | |
| I. pini | U | U* | RT-qPCR | Feeding | [32] | |
| I. pini | U | U* | RT-qPCR | JH III | [70] | |
| I. confusus | U | NR | RT-qPCR | Feeding | [47] | |
| I. confusus | U | NR | RT-qPCR | JH III | [47] | |
| D. jeffreyi | U* | C | Northern blot | JH III | [28] | |
| D. ponderosae | U* | U | RT-qPCR | Feeding | [71] | |
| D. armandi | U* | U | RT-qPCR | Feeding | [48] | |
| D. armandi | U* | C | RT-qPCR | JH III | [48] | |
| MK | I. pini | U* | C | RT-qPCR | JH III | [70] |
| MDPC | I. pini | U* | U | RT-qPCR | Feeding | [32] |
| I. pini | U | U* | RT-qPCR | JH III | [70] | |
| IPPI | I. pini | U | U* | RT-qPCR | Feeding | [32] |
| I. pini | U | U* | RT-qPCR | JH III | [70] | |
| GPPS/FPPS | GPPS I. pini | U* | D | RT-qPCR | Feeding | [32] |
| GPPS I. pini | U | U* | RT-qPCR | JH III | [70] | |
| FPPS I. pini | U* | D | RT-qPCR | Feeding | [32] | |
| FPPS I. pini | U* | D | RT-qPCR | JH III | [70] | |
| D. ponderosae | U* | C | RT-qPCR | Feeding | [36] | |
| GPPS I. confusus | U | NR | RT-qPCR | Feeding | [47] | |
| GPPS I. confusus | U | NR | RT-qPCR | JH III | [47] | |
| GGPPS | D. ponderosae | U* | C | RT-qPCR | Feeding | [71] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarabia, L.E.; López, M.F.; Obregón-Molina, G.; Cano-Ramírez, C.; Sánchez-Martínez, G.; Zúñiga, G. The Differential Expression of Mevalonate Pathway Genes in the Gut of the Bark Beetle Dendroctonus rhizophagus (Curculionidae: Scolytinae) Is Unrelated to the de Novo Synthesis of Terpenoid Pheromones. Int. J. Mol. Sci. 2019, 20, 4011. https://doi.org/10.3390/ijms20164011
Sarabia LE, López MF, Obregón-Molina G, Cano-Ramírez C, Sánchez-Martínez G, Zúñiga G. The Differential Expression of Mevalonate Pathway Genes in the Gut of the Bark Beetle Dendroctonus rhizophagus (Curculionidae: Scolytinae) Is Unrelated to the de Novo Synthesis of Terpenoid Pheromones. International Journal of Molecular Sciences. 2019; 20(16):4011. https://doi.org/10.3390/ijms20164011
Chicago/Turabian StyleSarabia, Laura Elisa, María Fernanda López, Gabriel Obregón-Molina, Claudia Cano-Ramírez, Guillermo Sánchez-Martínez, and Gerardo Zúñiga. 2019. "The Differential Expression of Mevalonate Pathway Genes in the Gut of the Bark Beetle Dendroctonus rhizophagus (Curculionidae: Scolytinae) Is Unrelated to the de Novo Synthesis of Terpenoid Pheromones" International Journal of Molecular Sciences 20, no. 16: 4011. https://doi.org/10.3390/ijms20164011
APA StyleSarabia, L. E., López, M. F., Obregón-Molina, G., Cano-Ramírez, C., Sánchez-Martínez, G., & Zúñiga, G. (2019). The Differential Expression of Mevalonate Pathway Genes in the Gut of the Bark Beetle Dendroctonus rhizophagus (Curculionidae: Scolytinae) Is Unrelated to the de Novo Synthesis of Terpenoid Pheromones. International Journal of Molecular Sciences, 20(16), 4011. https://doi.org/10.3390/ijms20164011