Characterization of the First Bacterial and Thermostable GDP-Mannose 3,5-Epimerase


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
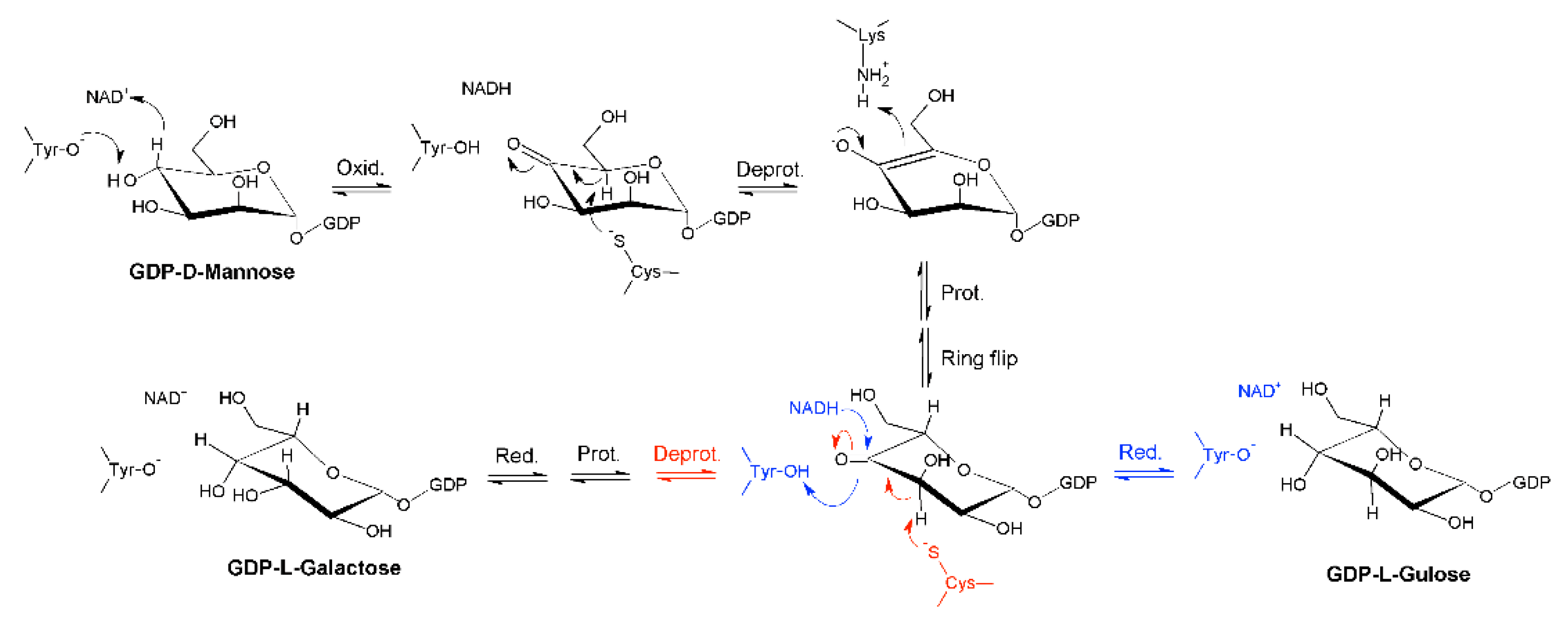
:1. Introduction
2. Results
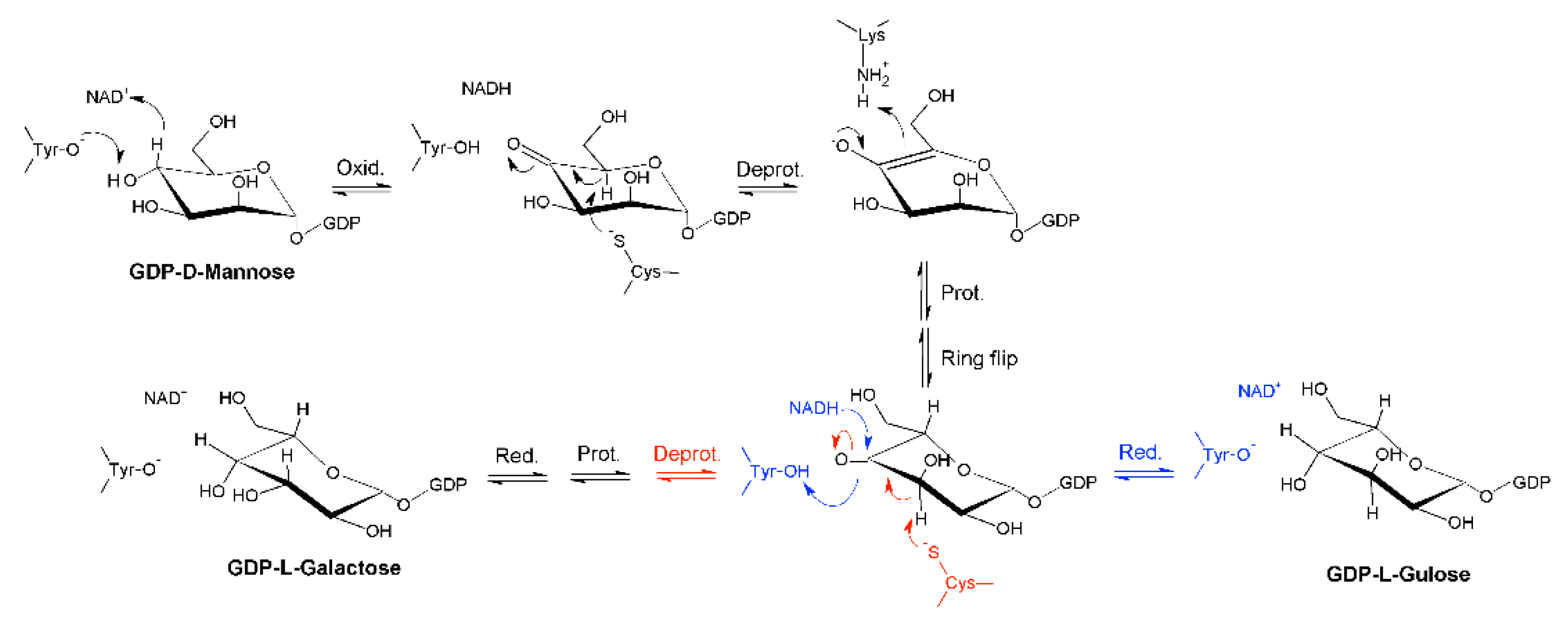
2.1. Identification of a Bacterial GM35E Homologue
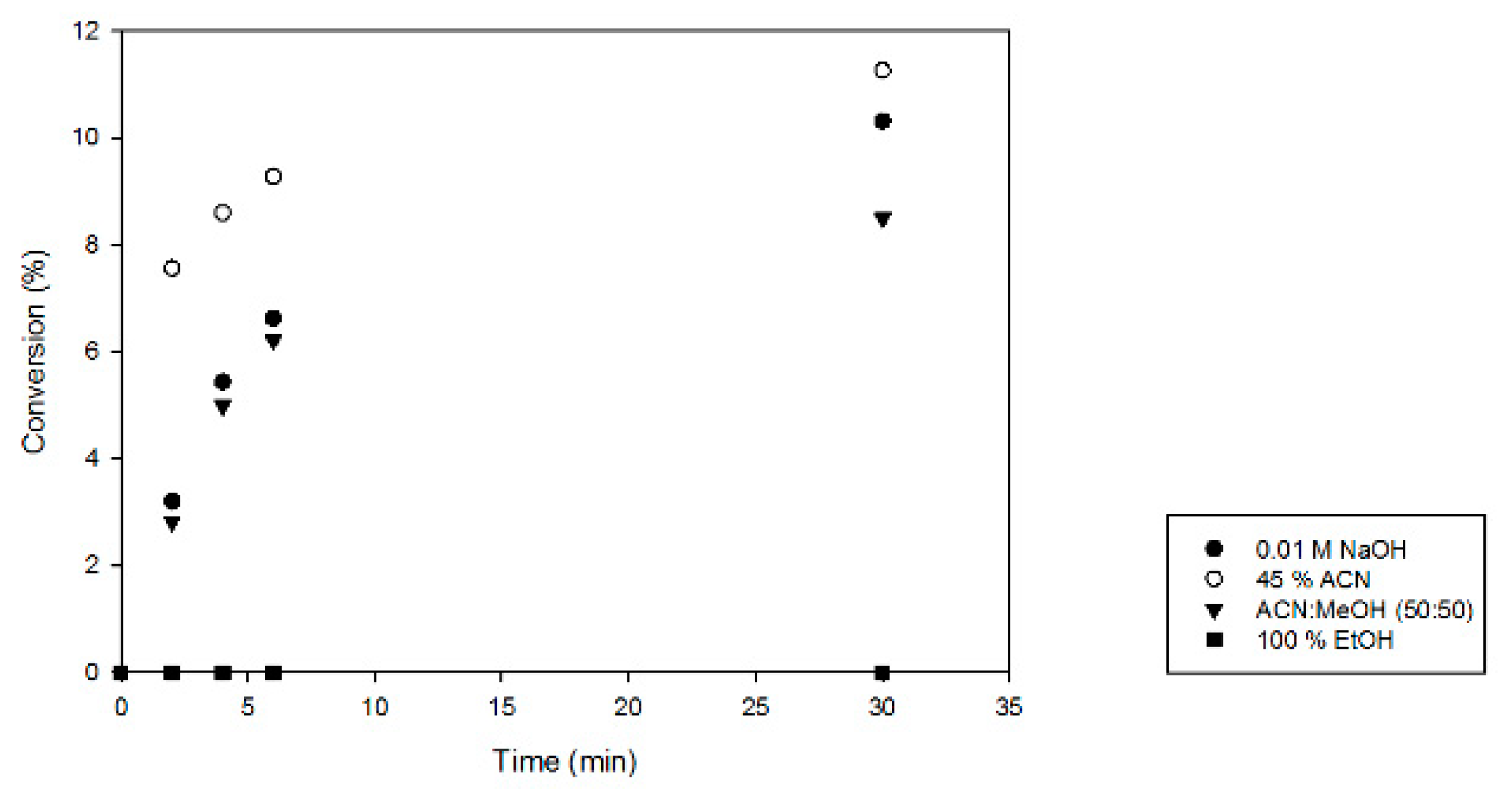
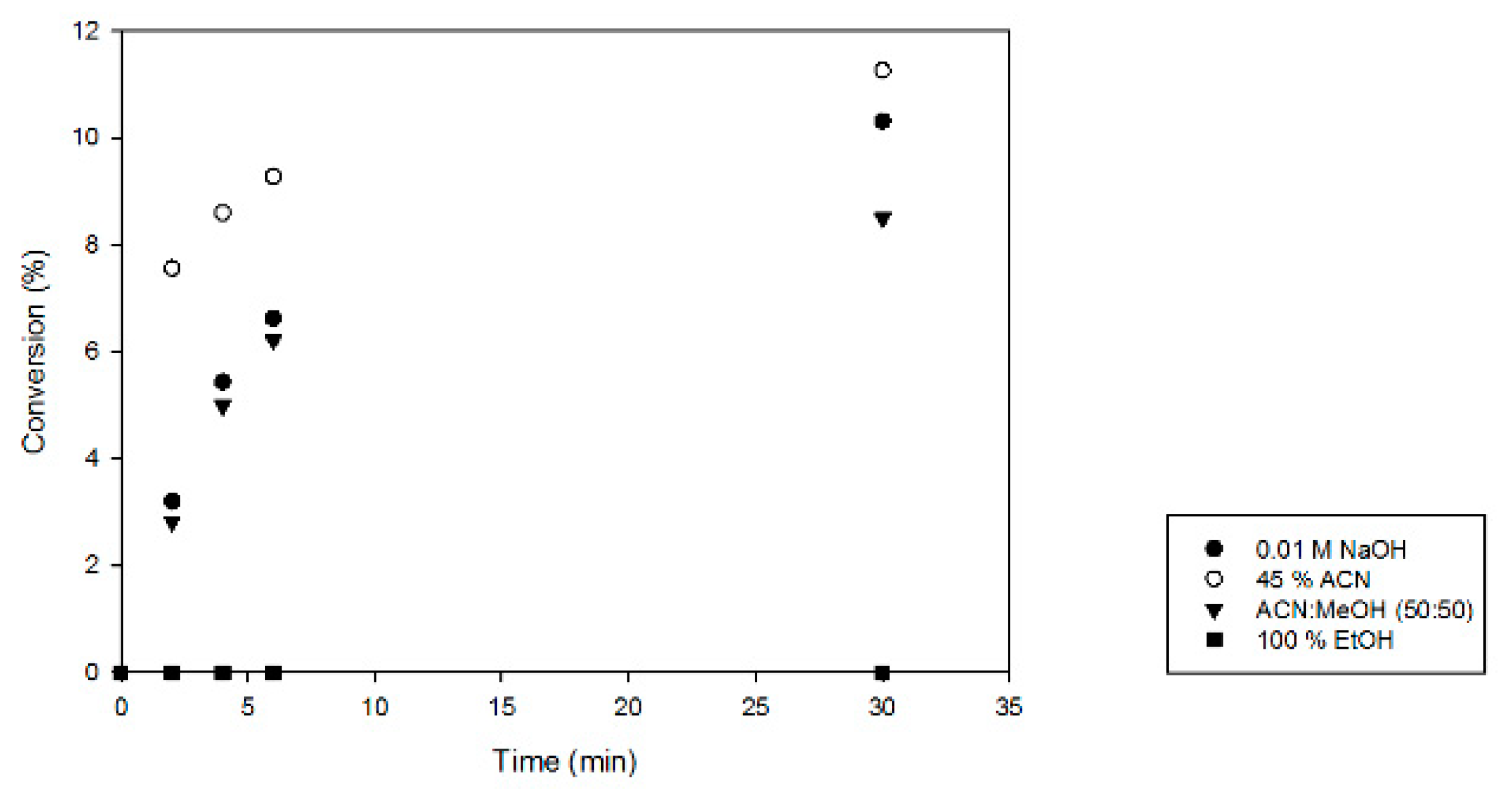
2.2. Optimization of Activity Testing
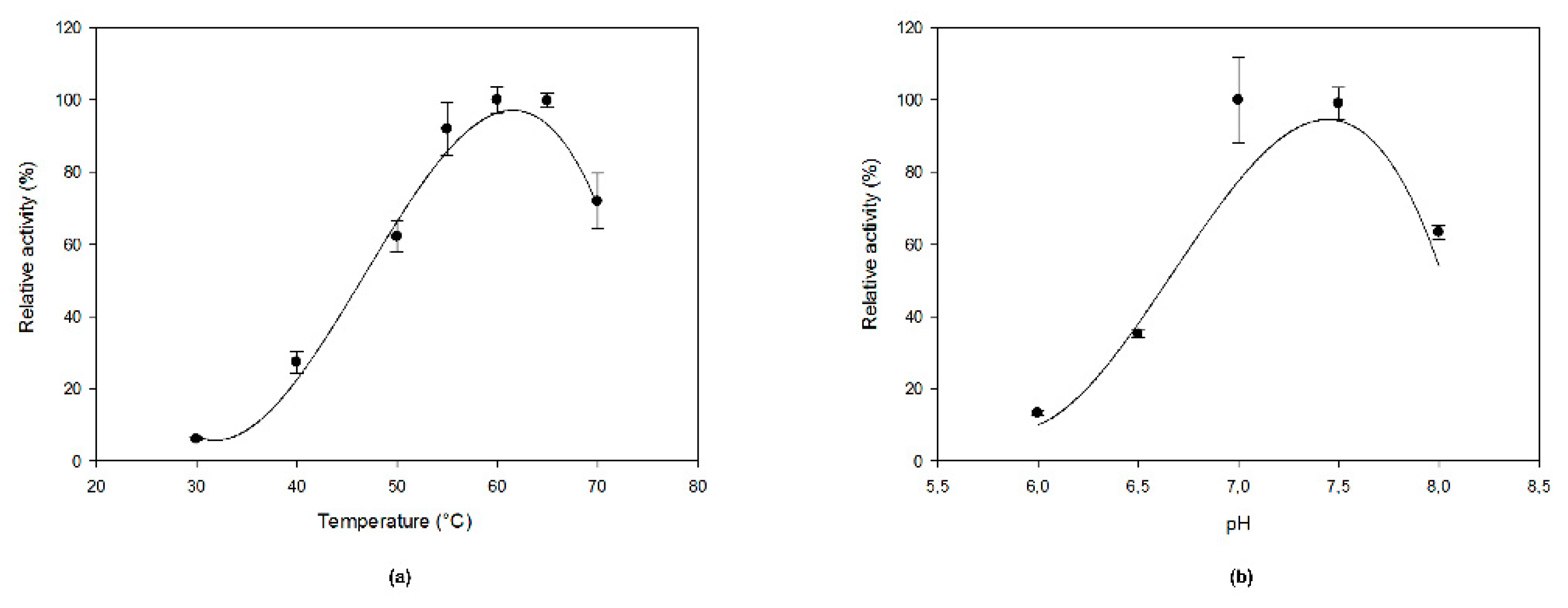
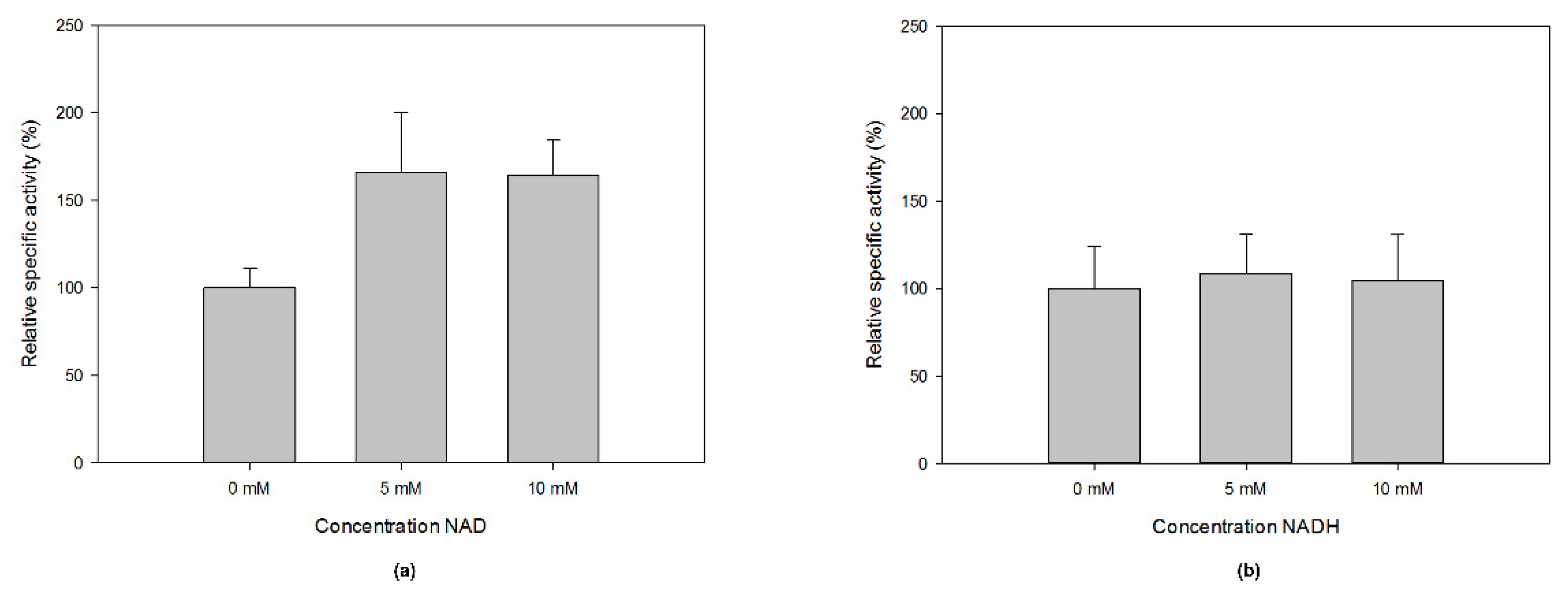
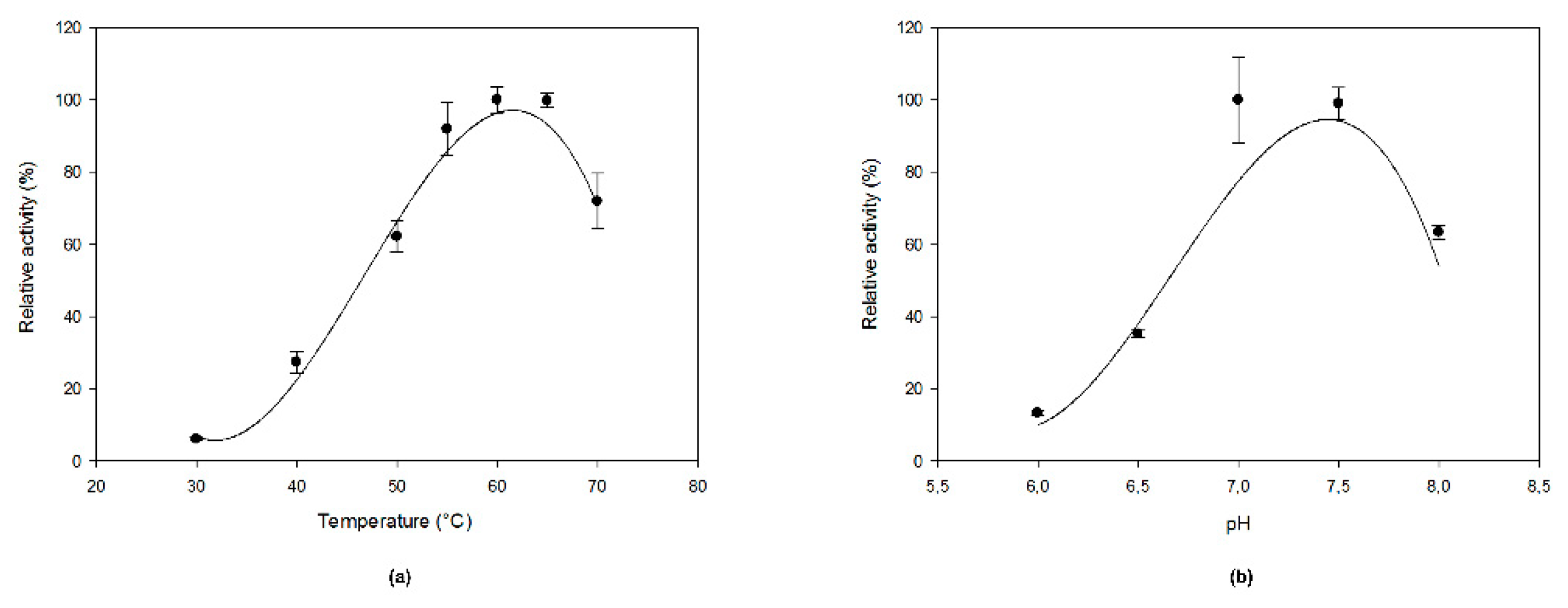
2.3. Optimal Temperature, pH, And Kinetic Properties
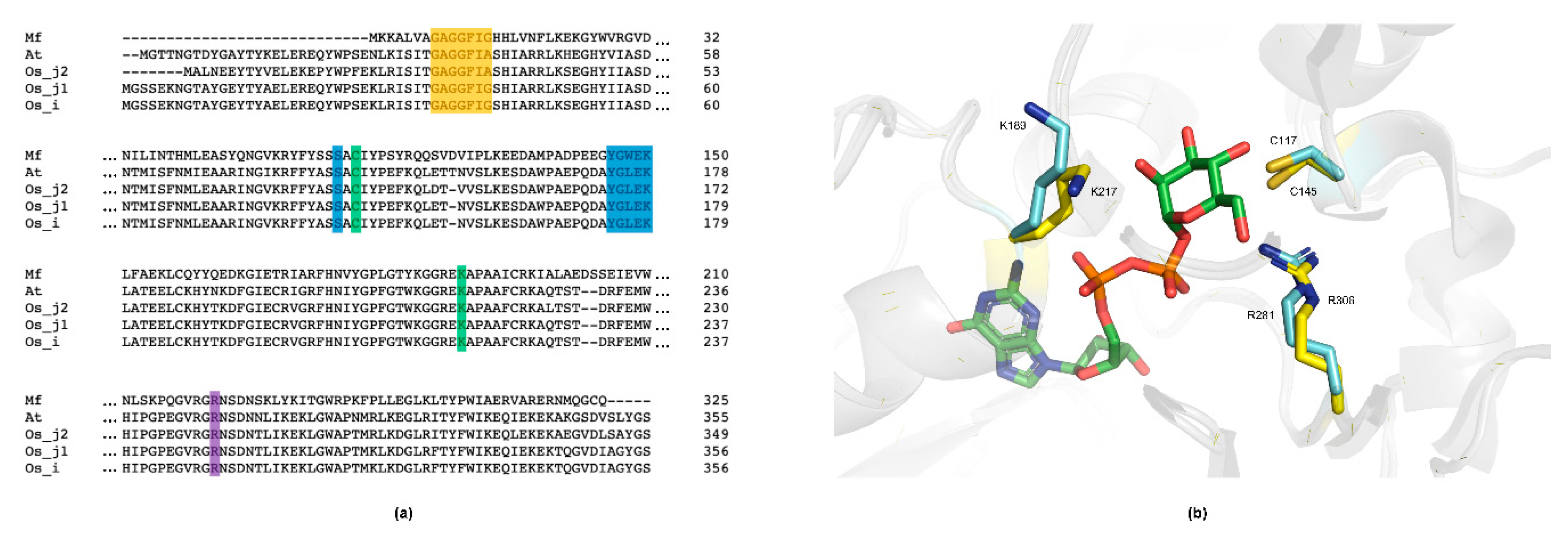
2.4. Sequence and Structure Analysis of MfGM35E
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Gene Cloning And Transformation
4.3. Enzyme Production And Purification
4.4. Optimization of Product Analysis and Enzyme Inactivation
4.5. Activity Testing and Enzyme Characterization
4.6. Thermal Shift Assay
4.7. Sequence Analysis
4.8. Homology Modelling
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| GM35E | GDP-mannose 3,5-epimerase |
| Gal4E | UDP-glucose 4-epimerase |
| CEP | carbohydrate epimerase |
| GDP-Man | GDP-d-mannose |
| GDP-l-Gal | GDP-l-galactose |
| GDP-l-Gul | GDP-l-gulose |
| l-AA | l-ascorbic acid |
| LPS | lipopolysaccharide |
| SDR | short-chain dehydrogenase/reductase |
| ACN | acetonitrile |
| EtOH | ethanol |
| MeOH | methanol |
| LB | lysogeny broth |
| IPTG | isopropyl β-d-thiogalactopyranoside |
| PMSF | phenylmethane sulfonyl fluoride |
| DSF | differential scanning fluorimetry |
| FRET | fluorescence resonance energy transfer |
References
- Hélaine, V.; Mahdi, R.; Sudhir Babu, G.V.; de Berardinis, V.; Wohlgemuth, R.; Lemaire, M.; Guérard-Hélaine, C. Straightforward synthesis of terminally phosphorylated l-sugars via multienzymatic cascade reactions. Adv. Synth. Catal. 2015, 357, 1703–1708. [Google Scholar] [CrossRef]
- Xia, T.Y.; Li, Y.B.; Yin, Z.J.; Meng, X.B.; Li, S.C.; Li, Z.J. Synthesis of l-glucose and l-galactose derivatives from d-sugars. Chinese Chem. Lett. 2014, 25, 1220–1224. [Google Scholar] [CrossRef]
- Mulloy, B.; Forster, M.J. Conformation and dynamics of heparin and heparan sulfate. Glycobiology 2000, 10, 1147–1156. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Schmaltz, R.M.; Bozeman, T.C.; Paul, R.; Rishel, M.J.; Tsosie, K.S.; Hecht, S.M. Selective tumor cell targeting by the disaccharide moiety of bleomycin. J. Am. Chem. Soc. 2013, 135, 2883–2886. [Google Scholar] [CrossRef] [PubMed]
- Mathé, C.; Gosselin, G. l-Nucleoside enantiomers as antivirals drugs: A mini-review. Antiviral Res. 2006, 71, 276–281. [Google Scholar] [CrossRef]
- Gumina, G.; Song, G.Y.; Chu, C.K. l-Nucleosides as chemotherapeutic agents. FEMS Microbiol. Lett. 2001, 202, 9–15. [Google Scholar]
- Beerens, K.; Desmet, T.; Soetaert, W. Enzymes for the biocatalytic production of rare sugars. J. Ind. Microbiol. Biotechnol. 2012, 39, 823–834. [Google Scholar] [CrossRef]
- Ahmed, Z. Production of natural and rare pentoses using microorganisms and their enzymes. Electron. J. Biotechnol. 2001, 4, 103–111. [Google Scholar] [CrossRef]
- D’Alonzo, D.; Guaragna, A.; Palumbo, G. Recent advances in monosaccharide synthesis: A journey into l-hexose world. Curr. Org. Chem. 2009, 13, 71–98. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, T.; Jiang, B.; Mu, W. Enzymatic approaches to rare sugar production. Biotechnol. Adv. 2017, 35, 267–274. [Google Scholar] [CrossRef]
- Frihed, T.G.; Bols, M.; Pedersen, C.M. Synthesis of l-hexoses. Chem. Rev. 2015, 115, 3615–3676. [Google Scholar] [CrossRef] [PubMed]
- Major, L.L.; Wolucka, B.A.; Naismith, J.H. Structure and function of GDP-mannose-3′,5′-epimerase: An enzyme which performs three chemical reactions at the same active site. J. Am. Chem. Soc. 2005, 127, 18309–18320. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Sánchez, C.; Chen, M.; Edwards, D.J.; Shen, B. The biosynthetic gene cluster for the antitumor drug bleomycin from Streptomyces verticillus ATCC15003 supporting functional interactions between nonribosomal peptide synthetases and a polyketide synthase. Chem. Biol. 2000, 7, 623–642. [Google Scholar] [CrossRef]
- Schroeder, B.R.; Ghare, M.I.; Bhattacharya, C.; Paul, R.; Yu, Z.; Zaleski, P.A.; Bozeman, T.C.; Rishel, M.J.; Hecht, S.M. The disaccharide moiety of bleomycin facilitates uptake by cancer cells. J. Am. Chem. Soc. 2014, 136, 13641–13656. [Google Scholar] [CrossRef] [PubMed]
- Woodyer, R.D.; Christ, T.N.; Deweese, K.A. Single-step bioconversion for the preparation of l-gulose and l-galactose. Carbohydr. Res. 2010, 345, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Delattre, C.; Fenoradosoa, T.A.; Michaud, P. Galactans: An overview of their most important sourcing and applications as natural polysaccharides. Brazilian Arch. Biol. Technol. 2011, 54, 1075–1092. [Google Scholar] [CrossRef]
- Gutiérrez, M.; Capson, T.; Guzmán, H.M.; Quiñoá, E.; Riguera, R. l-Galactose as a natural product: Isolation from a marine octocoral of the first α-l-galactosyl saponin. Tetrahedron Lett. 2004, 45, 7833–7836. [Google Scholar] [CrossRef]
- Osbourn, A.; Goss, R.J.M.; Field, R.A. The saponins-polar isoprenoids with important and diverse biological activities. Nat. Prod. Rep. 2011, 28, 1261–1268. [Google Scholar] [CrossRef] [PubMed]
- Goudsmit, E.M.; Neufeld, E.F. Formation of GDP-l-galactose from GDP-d-mannose. Biochem. Biophys. Res. Commun. 1967, 26, 730–735. [Google Scholar] [CrossRef]
- Siow, R.S.; Teoh, S.; Teo, S.S.; Shukor, M.Y.B.A.; Phang, S.M.; Ho, C.L. Molecular cloning and characterization of GDP-mannose-3′,5′-epimerase from Gracilaria changii. J. Appl. Phycol. 2013, 25, 1309–1318. [Google Scholar] [CrossRef]
- Hebda, P.A.; Behrman, E.J.; Barber, G. The guanosine 5′-diphosphate d-mannose:guanosine l-galactose epimerase of Chlorella pyrenoidosa. Arch. Biochmistery Biophys. 1979, 194, 496–502. [Google Scholar] [CrossRef]
- Watanabe, K.; Suzuki, K.; Kitamura, S. Characterization of a GDP-d-mannose 3″,5″-epimerase from rice. Phytochemistry 2005, 67, 338–346. [Google Scholar] [CrossRef] [PubMed]
- Wolucka, B.A.; Van Montagu, M. GDP-mannose 3′,5′-epimerase forms GDP-l-gulose, a putative intermediate for the de novo biosynthesis of vitamin C in plants. J. Biol. Chem. 2003, 278, 47483–47490. [Google Scholar] [CrossRef]
- Bulley, S.; Laing, W. The regulation of ascorbate biosynthesis. Curr. Opin. Plant Biol. 2016, 33, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Van Overtveldt, S.; Verhaeghe, T.; Joosten, H.J.; van den Bergh, T.; Beerens, K.; Desmet, T. A structural classification of carbohydrate epimerases: From mechanistic insights to practical applications. Biotechnol. Adv. 2015, 33, 1814–1828. [Google Scholar] [CrossRef] [PubMed]
- Beerens, K.; Soetaert, W.; Desmet, T. UDP-hexose 4-epimerases: A view on structure, mechanism and substrate specificity. Carbohydr. Res. 2015, 414, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Beerens, K.; Soetaert, W.; Desmet, T. Characterization and mutational analysis of the UDP-Glc (NAc) 4-epimerase from Marinithermus hydrothermalis. Appl. Microbiol. Biotechnol. 2013, 97, 7733–7740. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.T.B.; Tanner, M.E. Mechanism and active site residues of GDP-fucose synthase. J. Am. Chem. Soc. 2008, 130, 17593–17602. [Google Scholar] [CrossRef]
- Giraud, M.-F.; Naismith, J.H. The rhamnose pathway. Curr. Opin. Struct. Biol. 2000, 10, 687–696. [Google Scholar] [CrossRef]
- Watt, G.; Leoff, C.; Harper, A.D.; Bar-Peled, M. A bifunctional 3,5-epimerase/4-keto reductase for nucleotide-rhamnose synthesis in Arabidopsis. Plant Physiol. 2004, 134, 1337–1346. [Google Scholar] [CrossRef]
- Bloom, J.D.; Labthavikul, S.T.; Otey, C.R.; Arnold, F.H. Protein stability promotes evolvability. Proc. Natl. Acad. Sci. USA 2006, 103, 5869–5874. [Google Scholar] [CrossRef] [Green Version]
- Eijsink, V.G.H.; Gaseidnes, S.; Borchert, T.V.; van den Burg, B. Directed evolution of enzyme stability. Biomol. Eng. 2005, 22, 21–30. [Google Scholar] [CrossRef]
- Pol, A.; Barends, T.R.M.; Dietl, A.; Khadem, A.F.; Eygensteyn, J.; Jetten, M.S.M.; Op den Camp, H.J.M. Rare earth metals are essential for methanotrophic life in volcanic mudpots. Environ. Microbiol. 2014, 16, 255–264. [Google Scholar] [CrossRef]
- Gil, A.; Siegel, D.; Permentier, H.; Reijngoud, D.; Dekker, F.; Bischoff, R. Stability of energy metabolites—An often overlooked issue in metabolomics studies: A review. Electrophoresis 2015, 36, 2156–2169. [Google Scholar] [CrossRef]
- Cerdobbel, A.; De Winter, K.; Aerts, D.; Kuipers, R.; Joosten, H.-J.; Soetaert, W.; Desmet, T. Increasing the thermostability of sucrose phosphorylase by a combination of sequence- and structure-based mutagenesis. Protein Eng. Des. Sel. 2011, 24, 829–834. [Google Scholar] [CrossRef] [Green Version]
- Hill, B.L.; Figueroa, C.M.; Diez, M.D.A.; Lunn, J.E.; Iglesias, A.A.; Ballicora, M.A. On the stability of nucleoside diphosphate glucose metabolites: Implications for studies of plant carbohydrate metabolism. J. Exp. Bot. 2017, 68, 3331–3337. [Google Scholar] [CrossRef]
- Dowdle, J.; Ishikawa, T.; Gatzek, S.; Rolinski, S.; Smirnoff, N. Two genes in Arabidopsis thaliana encoding GDP-l-galactose phosphorylase are required for ascorbate biosynthesis and seedling viability. Plant J. 2007, 52, 673–689. [Google Scholar] [CrossRef]
- Rosano, C.; Bisso, A.; Izzo, G.; Tonetti, M.; Sturla, L.; De Flora, A.; Bolognesi, M. Probing the catalytic mechanism of GDP-4-keto-6-deoxy-d-mannose epimerase/reductase by kinetic and crystallographic characterization of site-specific mutants. J. Mol. Biol. 2000, 303, 77–91. [Google Scholar] [CrossRef]
- Somers, W.S.; Stahl, M.L.; Sullivan, F.X. GDP-fucose synthetase from Escherichia coli: Structure of a unique member of the short-chain dehydrogenase/reductase family that catalyzes two distinct reactions at the same active site. Structure 1998, 6, 1601–1612. [Google Scholar] [CrossRef]
- Kavanagh, K.L.; Jörnvall, H.; Persson, B.; Oppermann, U. The SDR superfamily: Functional and structural diversity within a family of metabolic and regulatory enzymes. Cell. Mol. Life Sci. 2008, 65, 3895–3906. [Google Scholar] [CrossRef]
- Beerens, K.; Van Overtveldt, S.; Desmet, T. The “epimerring” highlights the potential of carbohydrate epimerases for rare sugar production. Biocatal. Biotransform. 2017, 35, 230–237. [Google Scholar] [CrossRef]
- Vazquez-Figueroa, E.; Yeh, V.; Broering, J.M.; Chaparro-Riggers, J.F.; Bommarius, A.S. Thermostable variants constructed via the structure-guided consensus method also show increased stability in salts solutions and homogeneous aqueous-organic media. Protein Eng. Des. Sel. 2008, 21, 673–680. [Google Scholar] [CrossRef] [Green Version]
- Doukyu, N.; Ogino, H. Organic solvent-tolerant enzymes. Biochem. Eng. J. 2010, 48, 270–282. [Google Scholar] [CrossRef]
- Serebryakova, L.T.; Zorin, N.A.; Karyakin, A.A. Improvement of hydrogenase enzyme activity by water-miscible organic solvents. Enzyme Microb. Technol. 2009, 44, 329–333. [Google Scholar] [CrossRef]
- Liu, C.; Yang, G.; Wu, L.; Tian, G.; Zhang, Z.; Feng, Y. Switch of substrate specificity of hyperthermophilic acylaminoacyl peptidase by combination of protein and solvent engineering. Protein Cell 2011, 2, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Stepankova, V.; Bidmanova, S.; Koudelakova, T.; Prokop, Z.; Chaloupkova, R.; Damborsky, J. Strategies for stabilization of enzymes in organic solvents. ACS Catal. 2013, 3, 2823–2836. [Google Scholar] [CrossRef]
- Klibanov, A.M. Improving enzymes by using them in organic solvents. Nature 2001, 409, 241–246. [Google Scholar] [CrossRef]
- Zhu, Q.; Chen, Q.; Song, Y.; Huang, H.; Li, J.; Ma, J.; Li, Q.; Ju, J. Deciphering the sugar biosynthetic pathway and tailoring steps of nucleoside antibiotic A201A unveils a GDP-l-galactose mutase. Proc. Natl. Acad. Sci. USA 2017, 114, 4948–4953. [Google Scholar] [CrossRef]
- Sanapala, S.R.; Kulkarni, S.S. Expedient route to access rare deoxy amino l-sugar building blocks for the assembly of bacterial glycoconjugates. J. Am. Chem. Soc. 2016, 138, 4938–4947. [Google Scholar] [CrossRef]
- Allard, S.T.M.; Giraud, M.-F.; Naismith, J.H. Epimerases: Structure, function and mechanism. Cell. Mol. Life Sci. 2001, 58, 1650–1665. [Google Scholar] [CrossRef]
- King, J.D.; Mulrooney, E.F.; Vinogradov, E.; Kneidinger, B.; Mead, K.; Lam, J.S. lfnA from Pseudomonas aeruginosa O12 and wbuX from Escherichia coli O145 encode membrane-associated proteins and are required for expression of 2,6-dideoxy-2-acetamidino-l-galactose in lipopolysaccharide O an. J. Bacteriol. 2008, 190, 1671–1679. [Google Scholar] [CrossRef]
- Mulrooney, E.F.; Poon, K.K.H.; McNally, D.J.; Brisson, J.R.; Lam, J.S. Biosynthesis of UDP-N-acetyl-l-fucosamine, a precursor to the biosynthesis of lipopolysaccharide in Pseudomonas aeruginosa serotype O11. J. Biol. Chem. 2005, 280, 19535–19542. [Google Scholar] [CrossRef]
- Zhang, Y.M.; Rock, C.O. Membrane lipid homeostasis in bacteria. Nat. Rev. Microbiol. 2008, 6, 222–233. [Google Scholar] [CrossRef]
- Sohlenkamp, C.; Geiger, O. Bacterial membrane lipids: Diversity in structures and pathways. FEMS Microbiol. Rev. 2015, 40, 133–159. [Google Scholar] [CrossRef]
- Stadthagen, G.; Sambou, T.; Guerin, M.; Barilone, N.; Boudou, F.; Korduláková, J.; Charles, P.; Alzari, P.M.; Lemassu, A.; Daffé, M.; et al. Genetic basis for the biosynthesis of methylglucose lipopolysaccharides in Mycobacterium tuberculosis. J. Biol. Chem. 2007, 282, 27270–27276. [Google Scholar] [CrossRef]
- Lerouge, I.; Verreth, C.; Michiels, J.; Carlson, R.W.; Datta, A.; Gao, M.Y.; Vanderleyden, J. Three genes encoding for putative methyl- and acetyltransferases map adjacent to the wzm and wzt genes and are essential for O-antigen biosynthesis in Rhizobium etli CE3. Mol. Plant. Microbe. Interact. 2003, 16, 1085–1093. [Google Scholar] [CrossRef]
- Ohashi, H.; Wahl, C.; Ohashi, T.; Elling, L.; Fujiyama, K. Effective synthesis of guanosine 5′-diphospho-b-l-galactose. Adv. Synth. Catal. 2017, 359, 4227–4234. [Google Scholar] [CrossRef]
- Eastwood, H.; Xia, F.; Lo, M.; Zhou, J.; Jordan, J.B.; McCarter, J.; Barnhart, W.W.; Gahm, K.-H. Development of a nucleotide sugar purification method using a mixed mode column & mass spectrometry detection. J. Pharm. Biomed. Anal. 2015, 115, 402–409. [Google Scholar]
- Schmölzer, K.; Gutmann, A.; Diricks, M.; Desmet, T.; Nidetzky, B. Sucrose synthase: A unique glycosyltransferase for biocatalytic glycosylation process development. Biotechnol. Adv. 2015, 34, 88–111. [Google Scholar] [CrossRef]
- Desmet, T.; Soetaert, W.; Bojarová, P.; Kařen, V.; Dijkhuizen, L.; Eastwick-Field, V.; Schiller, A. Enzymatic glycosylation of small molecules: Challenging substrates require tailored catalysts. Chem. A Eur. J. 2012, 18, 10786–10801. [Google Scholar] [CrossRef]
- Palcic, M.M. Glycosyltransferases as biocatalysts. Curr. Opin. Chem. Biol. 2011, 15, 226–233. [Google Scholar] [CrossRef]
- Kren, V.V.; Rezanka, T. Sweet antibiotics-The role of glycosidic residues in antibiotic and antitumor activity and their randomization. FEMS Microbiol. Rev. 2008, 32, 858–889. [Google Scholar] [CrossRef]
- Klapproth, J.; Rapp, E.; Mahour, R.; Schildbach, A.; Reichl, U.; Schierhorn, A.; Pietzsch, M.; Rexer, T.F.T. One pot synthesis of GDP-mannose by a multi-enzyme cascade for enzymatic assembly of lipid-linked oligosaccharides. Biotechnol. Bioeng. 2017, 115, 192–205. [Google Scholar]
- Pfeiffer, M.; Bulfon, D.; Weber, H.; Nidetzky, B. A kinase-independent one-pot multienzyme cascade for an expedient synthesis of guanosine 5′-diphospho-d-mannose. Adv. Synth. Catal. 2016, 358, 3809–3816. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; Thompson, J.D.; Higgins, D.G.; Mcwilliam, H.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 1–6. [Google Scholar] [CrossRef]

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gevaert, O.; Van Overtveldt, S.; Beerens, K.; Desmet, T. Characterization of the First Bacterial and Thermostable GDP-Mannose 3,5-Epimerase. Int. J. Mol. Sci. 2019, 20, 3530. https://doi.org/10.3390/ijms20143530
Gevaert O, Van Overtveldt S, Beerens K, Desmet T. Characterization of the First Bacterial and Thermostable GDP-Mannose 3,5-Epimerase. International Journal of Molecular Sciences. 2019; 20(14):3530. https://doi.org/10.3390/ijms20143530
Chicago/Turabian StyleGevaert, Ophelia, Stevie Van Overtveldt, Koen Beerens, and Tom Desmet. 2019. "Characterization of the First Bacterial and Thermostable GDP-Mannose 3,5-Epimerase" International Journal of Molecular Sciences 20, no. 14: 3530. https://doi.org/10.3390/ijms20143530
APA StyleGevaert, O., Van Overtveldt, S., Beerens, K., & Desmet, T. (2019). Characterization of the First Bacterial and Thermostable GDP-Mannose 3,5-Epimerase. International Journal of Molecular Sciences, 20(14), 3530. https://doi.org/10.3390/ijms20143530