Indirect Radioiodination of DARPin G3 Using N-succinimidyl-Para-Iodobenzoate Improves the Contrast of HER2 Molecular Imaging

 ,
,  ,
, 
Abstract

1. Introduction
2. Results
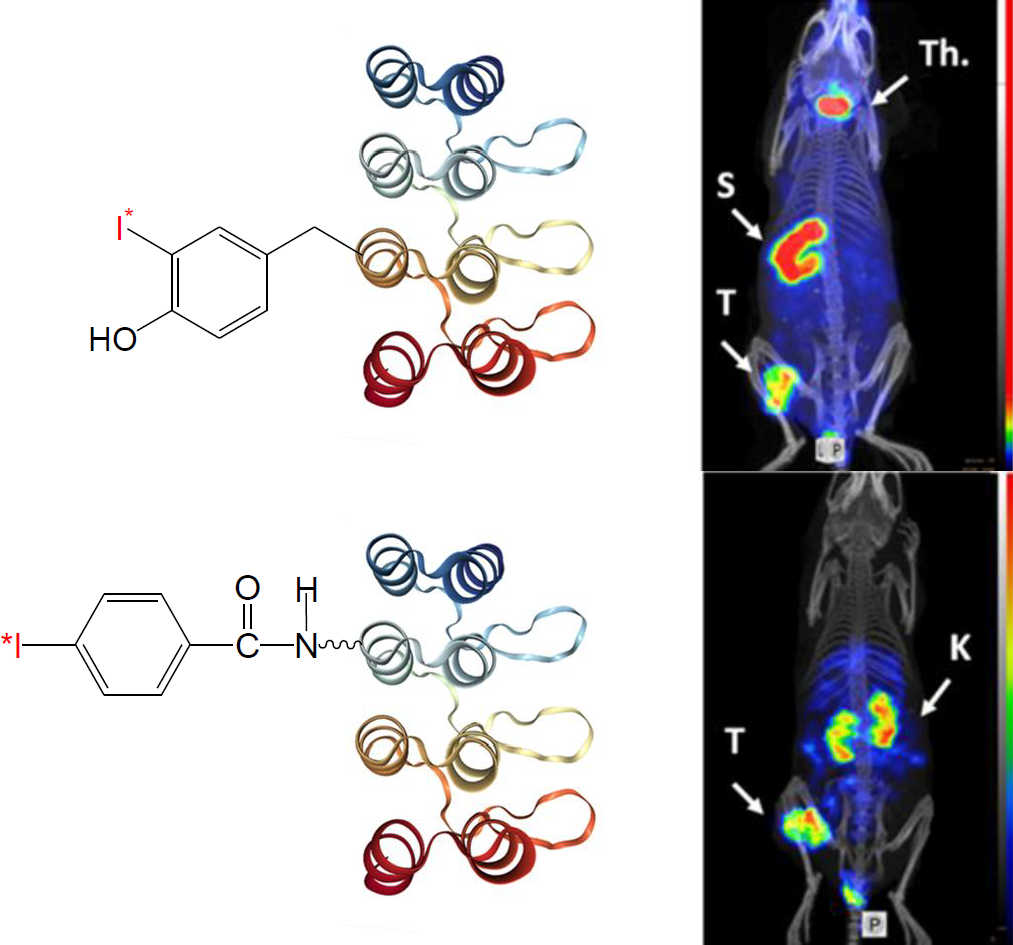
2.1. Radiolabeling and Stability
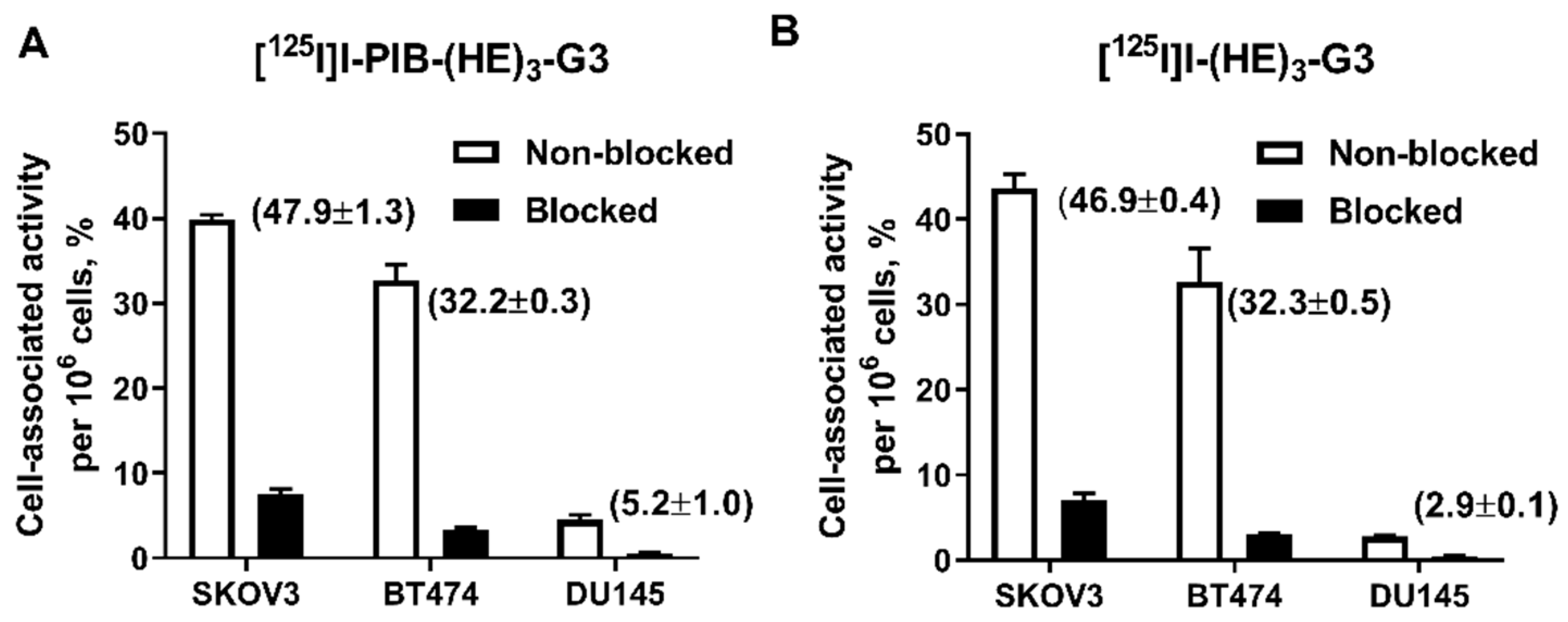
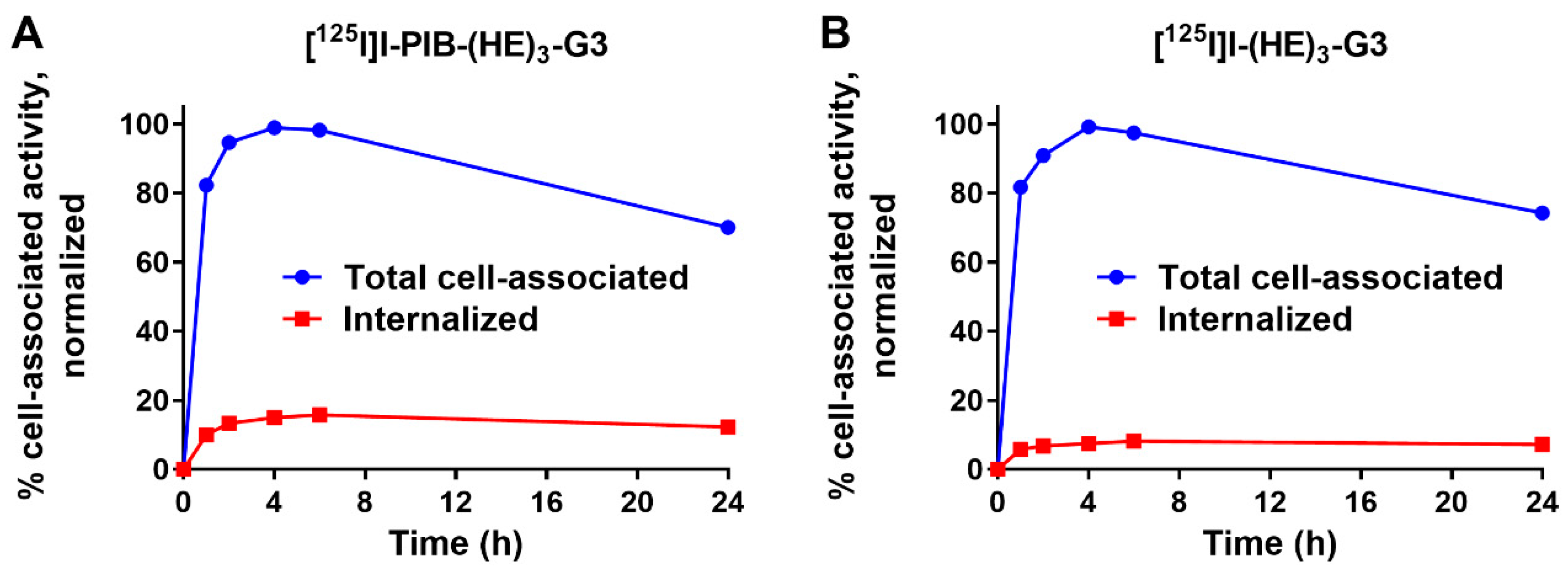
2.2. In Vitro Characterization of Radiolabeled DARPins
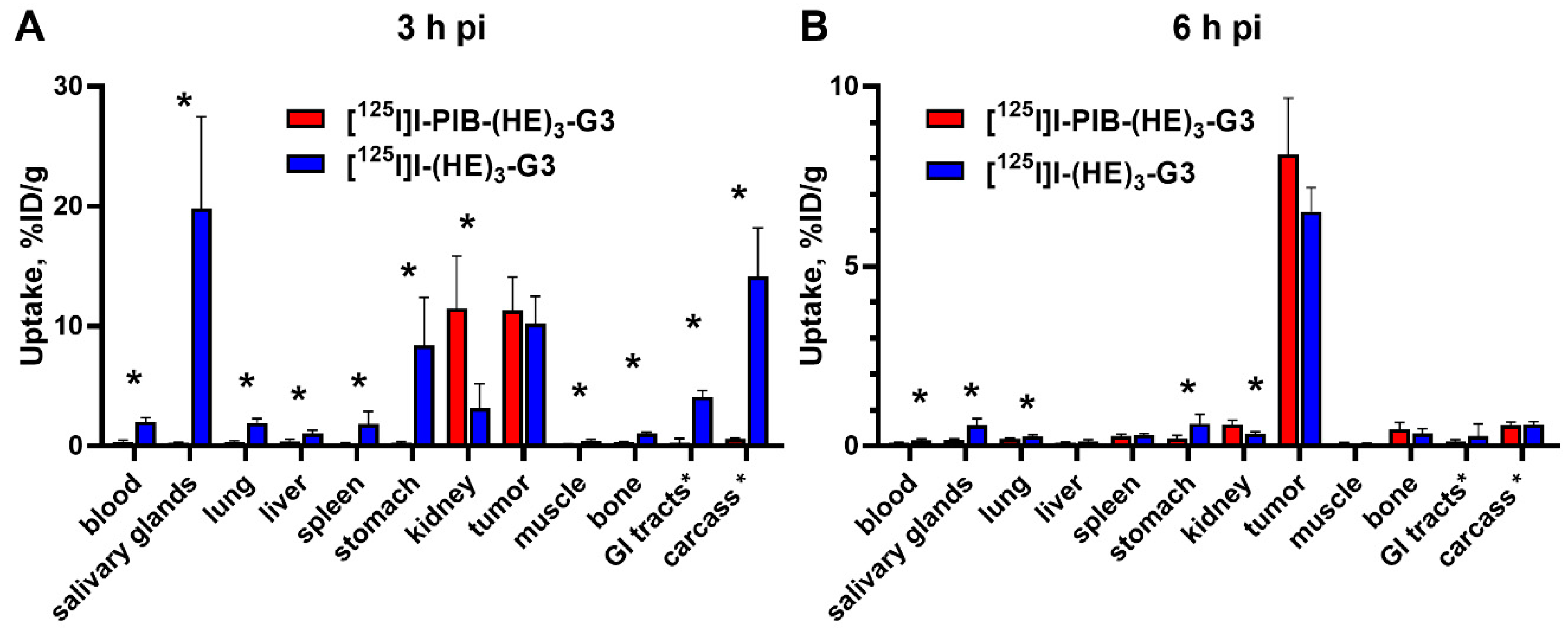
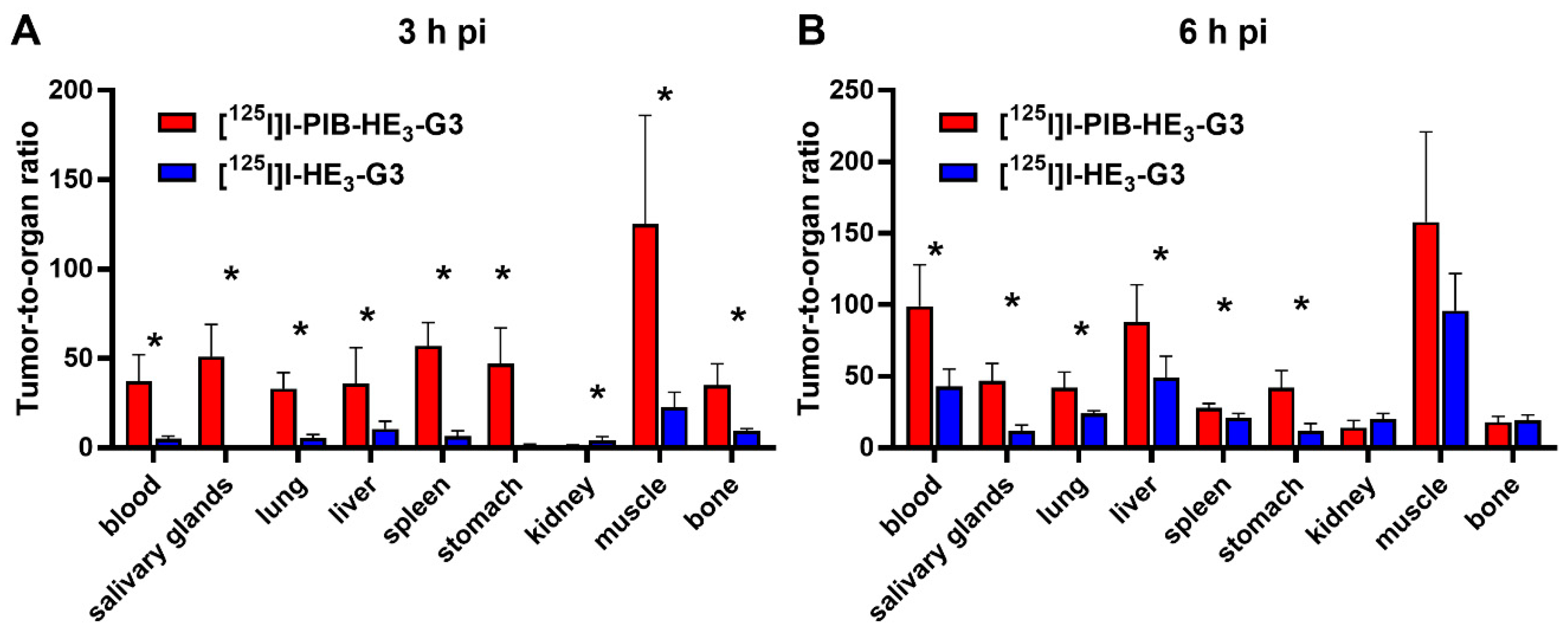
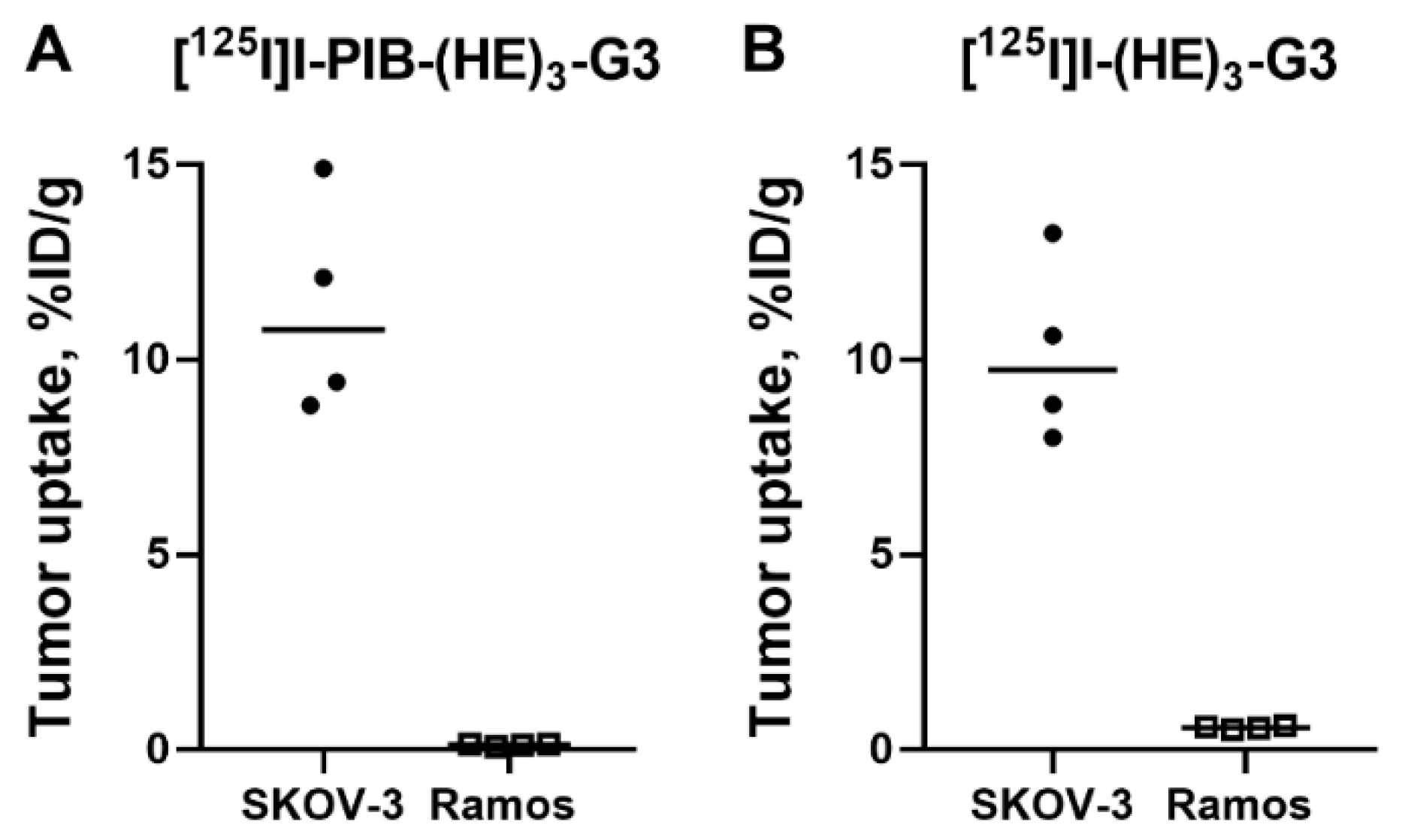
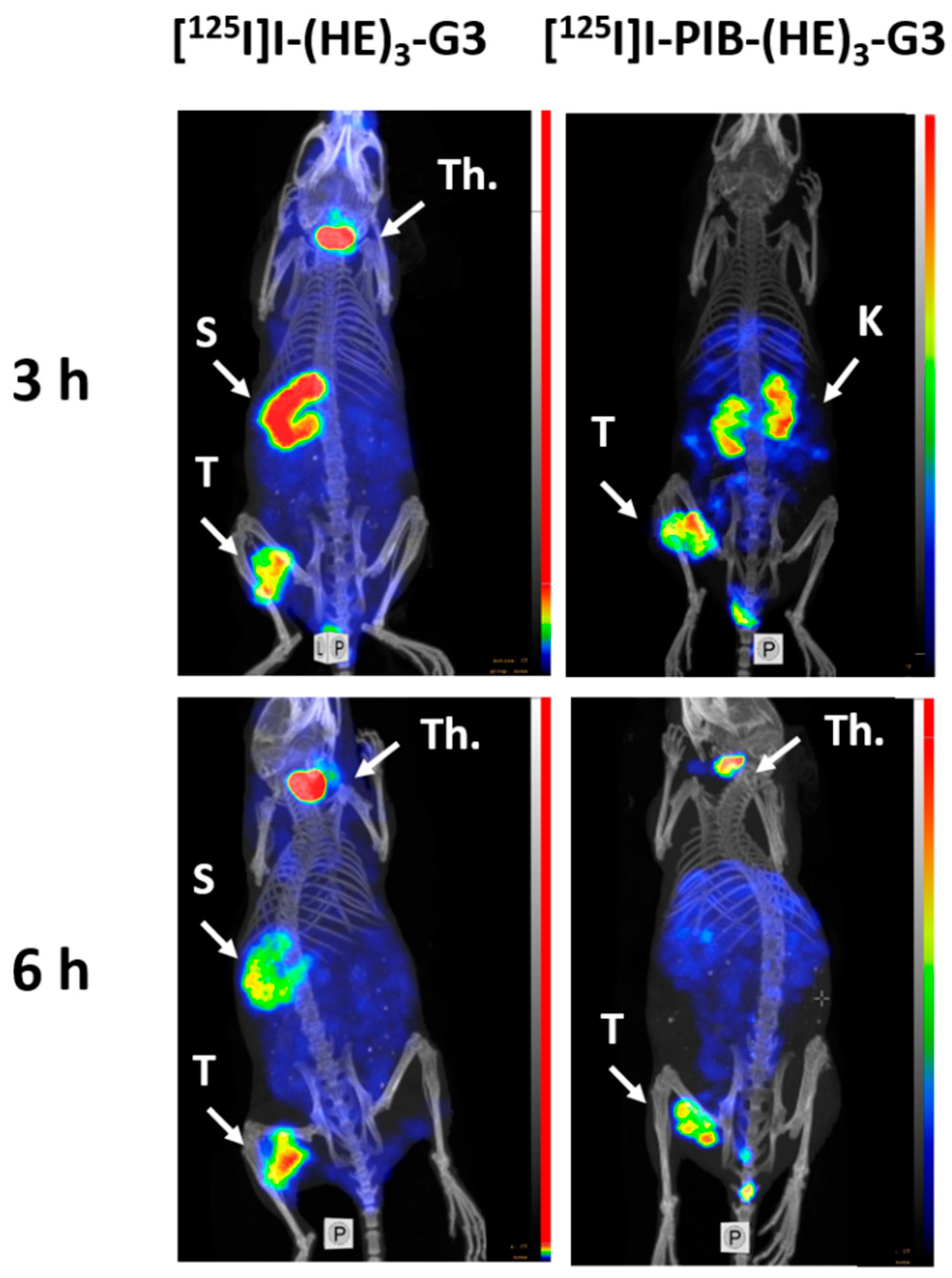
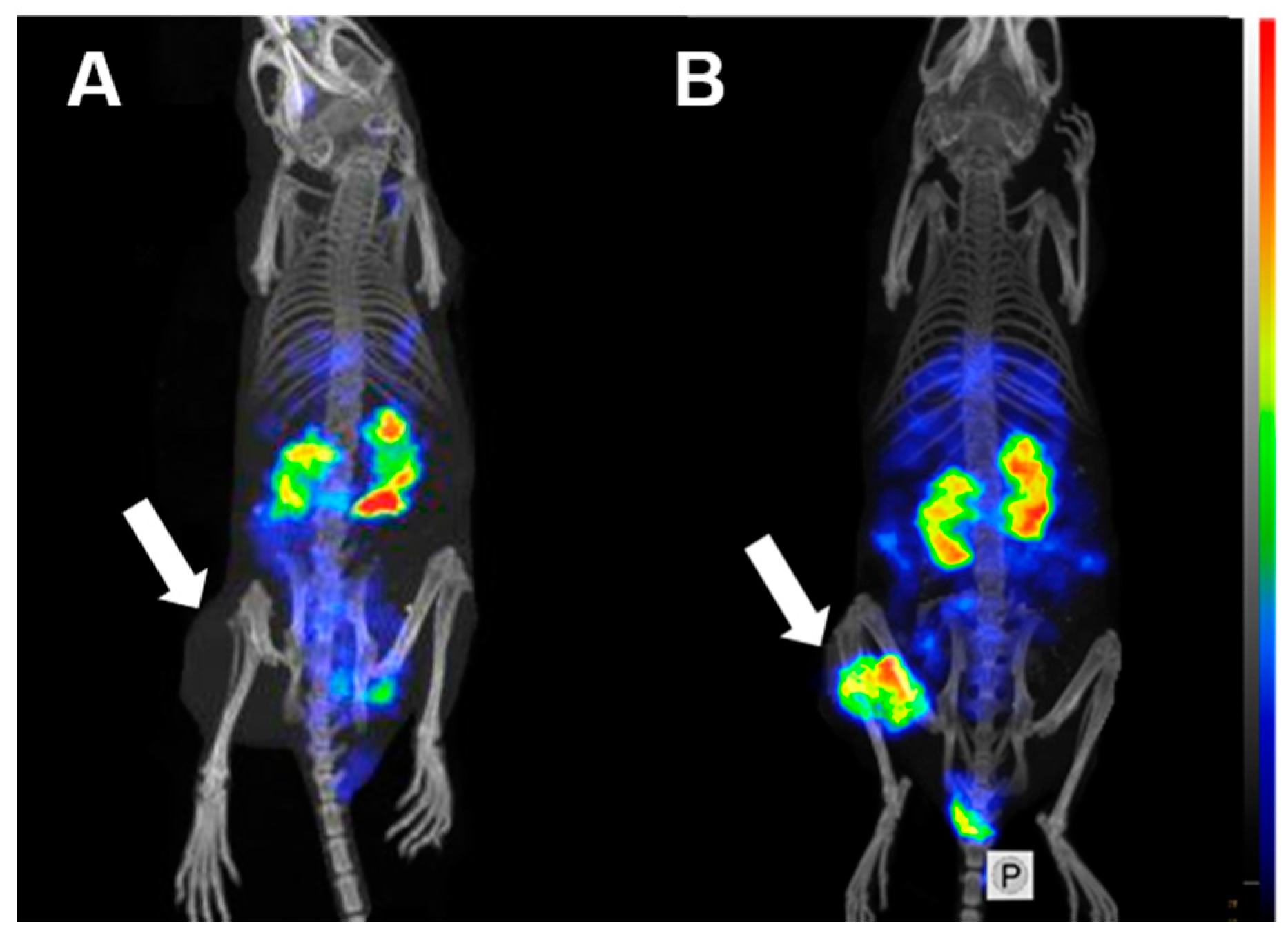
2.3. Animal Studies
3. Discussion
4. Materials and Methods
4.1. Direct Radioiodination
4.2. Indirect Radioiodination
4.3. Label Stability
4.4. In Vitro Characterization
4.5. Biodistribution Studies
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ADAPT | Albumin-Binding Domain Derived Affinity Protein |
| BSA | Bovine Serum Albumin |
| CT | Computed Tomography |
| DARPin | Designed Ankyrin Repeat Protein |
| HER2 | Human Epidermal Growth Factor Receptor 2 |
| iTLC | Instant Thin Layer Chromatography |
| MIP | Maximum Intensity Projections |
| PBS | Phosphate Buffered Saline |
| PET | Positron Emission Tomography |
| PIB | Para-Iodobenzoate |
| RGB | Red, Green and Blue Color Scale |
| SPECT | Single Photon Emission Computed Tomography |
References
- Tolmachev, V.; Stone-Elander, S.; Orlova, A. Radiolabelled Receptor-Tyrosine-Kinase Targeting Drugs For Patient Stratification and Monitoring of Therapy Response: Prospects and Pitfalls. Lancet Oncol. 2010, 10, 992–1000. [Google Scholar] [CrossRef]
- Krasniqi, A.; D’Huyvetter, M.; Devoogdt, N.; Frejd, F.Y.; Sörensen, J.; Orlova, A.; Keyaerts, M.; Tolmachev, V. Same-Day Imaging Using Small Proteins: Clinical Experience and Translational Prospects in Oncology. J. Nucl. Med. 2018, 59, 885–891. [Google Scholar] [CrossRef] [PubMed]
- Nygren, P.A.; Uhlén, M. Scaffolds for Engineering Novel Binding Sites in Proteins. Curr. Opin. Struct. Biol. 1997, 4, 463–469. [Google Scholar] [CrossRef]
- Skerra, A. Alternative Non-Antibody Scaffolds for Molecular Recognition. Curr. Opin. Biotechnol. 2007, 4, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Tolmachev, V.; Orlova, A. Influence of Labelling Methods on Biodistribution and Imaging Properties of Radiolabelled Peptides for Visualisation of Molecular Therapeutic Targets. Curr. Med. Chem. 2010, 24, 2636–2655. [Google Scholar] [CrossRef]
- Plückthun, A. Designed Ankyrin Repeat Proteins (DARPins): Binding Proteins for Research, Diagnostics, and Therapy. Annu. Rev. Pharmacol. Toxicol. 2015, 55, 489–511. [Google Scholar] [CrossRef] [PubMed]
- Boersma, Y.L. Advances in the Application of Designed Ankyrin Repeat Proteins (DARPins) as Research Tools and Protein Therapeutics. Methods Mol. Biol. 2018, 1798, 307–327. [Google Scholar] [CrossRef]
- Goldstein, R.; Sosabowski, J.; Livanos, M.; Leyton, J.; Vigor, K.; Bhavsar, G.; Nagy-Davidescu, G.; Rashid, M.; Miranda, E.; Yeung, J.; et al. Development of the Designed Ankyrin Repeat Protein (DARPin) G3 for HER2 Molecular Imaging. Eur. J. Nucl. Med. Mol. Imaging 2015, 42, 288–301. [Google Scholar] [CrossRef]
- Vorobyeva, A.; Bragina, O.; Altai, M.; Mitran, B.; Orlova, A.; Shulga, A.; Proshkina, G.; Chernov, V.; Tolmachev, V.; Deyev, S. Comparative Evaluation of Radioiodine and Technetium-Labeled DARPin 9_29 for Radionuclide Molecular Imaging of HER2 Expression in Malignant Tumors. Contrast Media Mol. Imaging 2018, 2018, 6930425. [Google Scholar] [CrossRef]
- Deyev, S.; Vorobyeva, A.; Schulga, A.; Proshkina, G.; Güler, R.; Löfblom, J.; Mitran, B.; Garousi, J.; Altai, M.; Buijs, J.; et al. Comparative Evaluation of Two DARPin Variants: Effect of Affinity, Size, and Label on Tumor Targeting Properties. Mol. Pharm. 2019, 3, 995–1008. [Google Scholar] [CrossRef]
- Vorobyeva, A.; Schulga, A.; Konovalova, E.; Güler, R.; Mitran, B.; Garousi, J.; Rinne, S.S.; Löfblom, J.; Orlova, A.; Deyev, S.; et al. Comparison of Tumor-Targeting Properties of Directly and Indirectly Radioiodinated Designed Ankyrin Repeat Protein (DARPin) G3 Variants for Molecular Imaging of HER2. Int. J. Oncol. 2019, 4, 1209–1220. [Google Scholar] [CrossRef] [PubMed]
- Vorobyeva, A.; Schulga, A.; Konovalova, E.; Güler, R.; Löfblom, J.; Sandström, M.; Garousi, J.; Chernov, V.; Bragina, O.; Orlova, A.; et al. Optimal Composition and Position of Histidine-Containing Tags Improves Biodistribution of 99mTc-Labeled DARPins. Sci. Rep. 2019, in press. [Google Scholar]
- Yarden, Y.; Sliwkowski, M.X. Untangling the ErbB Signalling Network. Nat. Rev. Mol. Cell. Biol. 2001, 2, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Roskoski, R., Jr. The ErbB/HER Family of Protein-Tyrosine Kinases and Cancer. Pharmacol. Res. 2014, 79, 34–74. [Google Scholar] [CrossRef] [PubMed]
- Slamon, D.J.; Clark, G.M.; Wong, S.G.; Levin, W.J.; Ullrich, A.; McGuire, W.L. Human Breast Cancer: Correlation of Relapse and Survival with Amplification of the HER-2/neu Oncogene. Science 1987, 235, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Kunz, P.L.; Mojtahed, A.; Fisher, G.A.; Ford, J.M.; Chang, D.T.; Balise, R.R.; Bangs, C.D.; Cherry, A.M.; Pai, R.K. HER2 Expression in Gastric and Gastroesophageal Junction Adenocarcinoma in a US Population: Clinicopathologic Analysis with Proposed Approach to HER2 Assessment. Appl. Immunohistochem. Mol. Morphol. 2012, 1, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Van Cutsem, E.; Bang, Y.J.; Feng-Yi, F.; Xu, J.M.; Lee, K.W.; Jiao, S.C.; Chong, J.L.; López-Sanchez, R.I.; Price, T.; Gladkov, O.; et al. HER2 Screening Data from ToGA: Targeting HER2 in Gastric and Gastroesophageal Junction Cancer. Gastric Cancer 2015, 18, 476–484. [Google Scholar] [CrossRef] [PubMed]
- Gianni, L.; Pienkowski, T.; Im, Y.H.; Roman, L.; Tseng, L.M.; Liu, M.C.; Lluch, A.; Staroslawska, E.; de la Haba-Rodriguez, J.; Im, S.A.; et al. Efficacy and Safety of Neoadjuvant Pertuzumab and Trastuzumab in Women with Locally Advanced, Inflammatory, or Early HER2-Positive Breast Cancer (NeoSphere): A Randomised Multicentre, Open-Label, Phase 2 Trial. Lancet Oncol. 2012, 13, 25–32. [Google Scholar] [CrossRef]
- Tafe, L.J.; Janjigian, Y.Y.; Zaidinski, M.; Hedvat, C.V.; Hameed, M.R.; Tang, L.H.; Hicks, J.B.; Shah, M.A.; Barbashina, V. Human Epidermal Growth Factor Receptor 2 Testing in Gastroesophageal Cancer: Correlation Between Immunohistochemistry and Fluorescence In Situ Hybridization. Arch. Pathol. Lab. Med. 2011, 11, 1460–1465. [Google Scholar] [CrossRef]
- Wolff, A.C.; Hammond, M.E.; Hicks, D.G.; Dowsett, M.; McShane, L.M.; Allison, K.H.; Allred, D.C.; Bartlett, J.M.; Bilous, M.; Fitzgibbons, P.; et al. Recommendations for Human Epidermal Growth Factor Receptor 2 Testing in Breast Cancer: American Society of Clinical Oncology/College of American Pathologists clinical practice guideline update. J. Clin. Oncol. 2013, 31, 3997–4013. [Google Scholar] [CrossRef]
- Bartley, A.N.; Washington, M.K.; Colasacco, C.; Ventura, C.B.; Ismaila, N.; Benson, A.B., 3rd; Carrato, A.; Gulley, M.L.; Jain, D.; Kakar, S.; et al. HER2 Testing and Clinical Decision Making in Gastroesophageal Adenocarcinoma: Guideline From the College of American Pathologists, American Society for Clinical Pathology, and the American Society of Clinical Oncology. J. Clin. Oncol. 2017, 4, 446–464. [Google Scholar] [CrossRef] [PubMed]
- Henry, K.E.; Ulaner, G.A.; Lewis, J.S. Human Epidermal Growth Factor Receptor 2-Targeted PET/Single- Photon Emission Computed Tomography Imaging of Breast Cancer: Noninvasive Measurement of a Biomarker Integral to Tumor Treatment and Prognosis. PET Clin. 2017, 3, 269–288. [Google Scholar] [CrossRef] [PubMed]
- Henry, K.E.; Ulaner, G.A.; Lewis, J.S. Clinical Potential of Human Epidermal Growth Factor Receptor 2 and Human Epidermal Growth Factor Receptor 3 Imaging in Breast Cancer. PET Clin. 2018, 3, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Steiner, D.; Forrer, P.; Plückthun, A. Efficient Selection of DARPins with Sub-Nanomolar Affinities Using SRP Phage Display. J. Mol. Biol. 2008, 5, 1211–1227. [Google Scholar] [CrossRef] [PubMed]
- Zahnd, C.; Wyler, E.; Schwenk, J.M.; Steiner, D.; Lawrence, M.C.; McKern, N.M.; Pecorari, F.; Ward, C.W.; Joos, T.O.; Plückthun, A. A Designed Ankyrin Repeat Protein Evolved to Picomolar Affinity to Her2. J. Mol. Biol. 2007, 4, 1015–1028. [Google Scholar] [CrossRef]
- Tolmachev, V.; Mume, E.; Sjöberg, S.; Frejd, F.Y.; Orlova, A. Influence of Valency and Labelling Chemistry on In Vivo Targeting Using Radioiodinated HER2-Binding Affibody Molecules. Eur. J. Nucl. Med. Mol. Imaging 2009, 4, 692–701. [Google Scholar] [CrossRef] [PubMed]
- Lindbo, S.; Garousi, J.; Mitran, B.; Altai, M.; Buijs, J.; Orlova, A.; Hober, S.; Tolmachev, V. Radionuclide Tumor Targeting Using ADAPT Scaffold Proteins: Aspects of Label Positioning and Residualizing Properties of the Label. J. Nucl. Med. 2018, 1, 93–99. [Google Scholar] [CrossRef]
- Wilbur, D.S.; Hadley, S.W.; Hylarides, M.D.; Abrams, P.G.; Beaumier, P.A.; Morgan, A.C.; Reno, J.M.; Fritzberg, A.R. Development of a Stable Radioiodinating Reagent to Label Monoclonal Antibodies for Radiotherapy of Cancer. J. Nucl. Med. 1989, 2, 216–226. [Google Scholar]
- Orlova, A.; Höglund, J.; Lubberink, M.; Lebeda, O.; Gedda, L.; Lundqvist, H.; Tolmachev, V.; Sundin, A. Comparative Biodistribution of the Radiohalogenated (Br, I and At) Antibody A33. Implications for In Vivo Dosimetry. Cancer Biother. Radiopharm. 2002, 4, 385–396. [Google Scholar] [CrossRef]
- Orlova, A.; Wållberg, H.; Stone-Elander, S.; Tolmachev, V. On the Selection of a Tracer for PET Imaging of HER2-Expressing Tumors: Direct Comparison of a 124I-Labeled Affibody Molecule and Trastuzumab in a Murine Xenograft Model. J. Nucl. Med. 2009, 3, 417–425. [Google Scholar] [CrossRef]
- Tolmachev, V.; Tran, T.A.; Rosik, D.; Sjöberg, A.; Abrahmsén, L.; Orlova, A. Tumor Targeting Using Affibody Molecules. Interplay of Affnity, Target Expression Level, and Binding Site Composition. J. Nucl. Med. 2012, 53, 953–960. [Google Scholar] [CrossRef] [PubMed]
- McLarty, K.; Cornelissen, B.; Scollard, D.A.; Done, S.J.; Chun, K.; Reilly, R.M. Associations Between the Uptake of 111In-DTPA-Trastuzumab, HER2 Density and Response to Trastuzumab (Herceptin) in Athymic Mice Bearing Subcutaneous Human Tumour Xenografts. Eur. J. Nucl. Med. Mol. Imaging 2009, 1, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Malmberg, J.; Tolmachev, V.; Orlova, A. Imaging Agents for In Vivo Molecular Profiling of Disseminated Prostate Cancer: Cellular Processing of [(111)In]-Labeled CHX-A″DTPA-Trastuzumab and Anti-HER2 ABY-025 Affibody in Prostate Cancer Cell Lines. Exp. Ther. Med. 2011, 2, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Barta, P.; Malmberg, J.; Melicharova, L.; Strandgård, J.; Orlova, A.; Tolmachev, V.; Laznicek, M.; Andersson, K. Protein Interactions with HER-Family Receptors Can Have Different Characteristics Depending on the Hosting Cell Line. Int. J. Oncol. 2012, 5, 1677–1682. [Google Scholar] [CrossRef]
- Orlova, A.; Bruskin, A.; Sjöström, A.; Lundqvist, H.; Gedda, L.; Tolmachev, V. Cellular Processing of (125)I- and (111)In-Labeled Epidermal Growth Factor (EGF) Bound to Cultured A431 Tumor Cells. Nucl. Med. Biol. 2000, 8, 827–835. [Google Scholar] [CrossRef]
- Garousi, J.; Lindbo, S.; Borin, J.; von Witting, E.; Vorobyeva, A.; Oroujeni, M.; Mitran, B.; Orlova, A.; Buijs, J.; Tolmachev, V.; et al. Comparative Evaluation of Dimeric and Monomeric Forms of ADAPT Scaffold Protein for Targeting of HER2-Expressing Tumours. Eur. J. Pharm. Biopharm. 2019, 134, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Dijkers, E.C.; Kosterink, J.G.; Rademaker, A.P.; Perk, L.R.; van Dongen, G.A.; Bart, J.; de Jong, J.R.; de Vries, E.G.; Lub-de Hooge, M.N. Development and Characterization of Clinical-Grade 89Zr-Trastuzumab for HER2/neu ImmunoPET Imaging. J. Nucl. Med. 2009, 6, 974–981. [Google Scholar] [CrossRef] [PubMed]
- Vegt, E.; de Jong, M.; Wetzels, J.F.; Masereeuw, R.; Melis, M.; Oyen, W.J.; Gotthardt, M.; Boerman, O.C. Renal Toxicity of Radiolabeled Peptides and Antibody Fragments: Mechanisms, Impact on Radionuclide Therapy, and Strategies for Prevention. J. Nucl. Med. 2010, 7, 1049–1058. [Google Scholar] [CrossRef]
- Eckelman, W.C.; Kilbourn, M.R.; Mathis, C.A. Specific to Nonspecific Binding in Radiopharmaceutical Studies: It’s Not So Simple As It Seems! Nucl. Med. Biol. 2009, 3, 235–237. [Google Scholar] [CrossRef]
- Weigelt, B.; Peterse, J.L.; van’t Veer, L.J. Breast Cancer Metastasis: Markers and Models. Nat. Rev. Cancer 2005, 8, 591–602. [Google Scholar] [CrossRef]
- Riihimäki, M.; Hemminki, A.; Sundquist, K.; Sundquist, J.; Hemminki, K. Metastatic Spread in Patients with Gastric Cancer. Oncotarget 2016, 32, 52307–52316. [Google Scholar] [CrossRef] [PubMed]
- Pruszynski, M.; Koumarianou, E.; Vaidyanathan, G.; Revets, H.; Devoogdt, N.; Lahoutte, T.; Zalutsky, M.R. Targeting Breast Carcinoma with Radioiodinated anti-HER2 Nanobody. Nucl. Med. Biol. 2013, 40, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Pruszynski, M.; Koumarianou, E.; Vaidyanathan, G.; Revets, H.; Devoogdt, N.; Lahoutte, T.; Lyerly, H.K.; Zalutsky, M.R. Improved tumor targeting of anti-HER2 nanobody through N-succinimidyl 4-guanidinomethyl-3-iodobenzoate radiolabeling. J. Nucl. Med. 2014, 55, 650–656. [Google Scholar] [CrossRef] [PubMed]
- Tolmachev, V.; Orlova, A.; Andersson, K. Methods for Radiolabelling of Monoclonal Antibodies. Methods Mol. Biol. 2014, 1060, 309–330. [Google Scholar] [CrossRef] [PubMed]
- Wållberg, H.; Orlova, A. Slow Internalization of Anti-HER2 Synthetic Affibody Monomer 111In-DOTA-ZHER2:342-pep2: Implications for Development of Labeled Tracers. Cancer Biother. Radiopharm. 2008, 23, 435–442. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test Solution | Normalized DARPin-Associated Activity, % | |
|---|---|---|
| [125I]I-PIB-(HE)3-G3 | [125I]I-(HE)3-G3 | |
| PBS (control) | 100 ± 0 | 99 ± 1 |
| 5000-fold KI | 99 ± 0 | 100 ± 0 |
| KD1(pM) | Weight (%) | KD2(nM) | Weight (%) | |
|---|---|---|---|---|
| [125I]I-PIB-(HE)3-G3 | 54 ± 3 | 65 | 3.39 ± 0.02 | 35 |
| [125I]I-(HE)3-G3 | 81 ± 4 | 60 | 2.76 ± 0.08 | 40 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vorobyeva, A.; Schulga, A.; Rinne, S.S.; Günther, T.; Orlova, A.; Deyev, S.; Tolmachev, V. Indirect Radioiodination of DARPin G3 Using N-succinimidyl-Para-Iodobenzoate Improves the Contrast of HER2 Molecular Imaging. Int. J. Mol. Sci. 2019, 20, 3047. https://doi.org/10.3390/ijms20123047
Vorobyeva A, Schulga A, Rinne SS, Günther T, Orlova A, Deyev S, Tolmachev V. Indirect Radioiodination of DARPin G3 Using N-succinimidyl-Para-Iodobenzoate Improves the Contrast of HER2 Molecular Imaging. International Journal of Molecular Sciences. 2019; 20(12):3047. https://doi.org/10.3390/ijms20123047
Chicago/Turabian StyleVorobyeva, Anzhelika, Alexey Schulga, Sara S. Rinne, Tyran Günther, Anna Orlova, Sergey Deyev, and Vladimir Tolmachev. 2019. "Indirect Radioiodination of DARPin G3 Using N-succinimidyl-Para-Iodobenzoate Improves the Contrast of HER2 Molecular Imaging" International Journal of Molecular Sciences 20, no. 12: 3047. https://doi.org/10.3390/ijms20123047
APA StyleVorobyeva, A., Schulga, A., Rinne, S. S., Günther, T., Orlova, A., Deyev, S., & Tolmachev, V. (2019). Indirect Radioiodination of DARPin G3 Using N-succinimidyl-Para-Iodobenzoate Improves the Contrast of HER2 Molecular Imaging. International Journal of Molecular Sciences, 20(12), 3047. https://doi.org/10.3390/ijms20123047