Three Melanin Pathway Genes, TH, yellow, and aaNAT, Regulate Pigmentation in the Twin-Spotted Assassin Bug, Platymeris biguttatus (Linnaeus)




Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
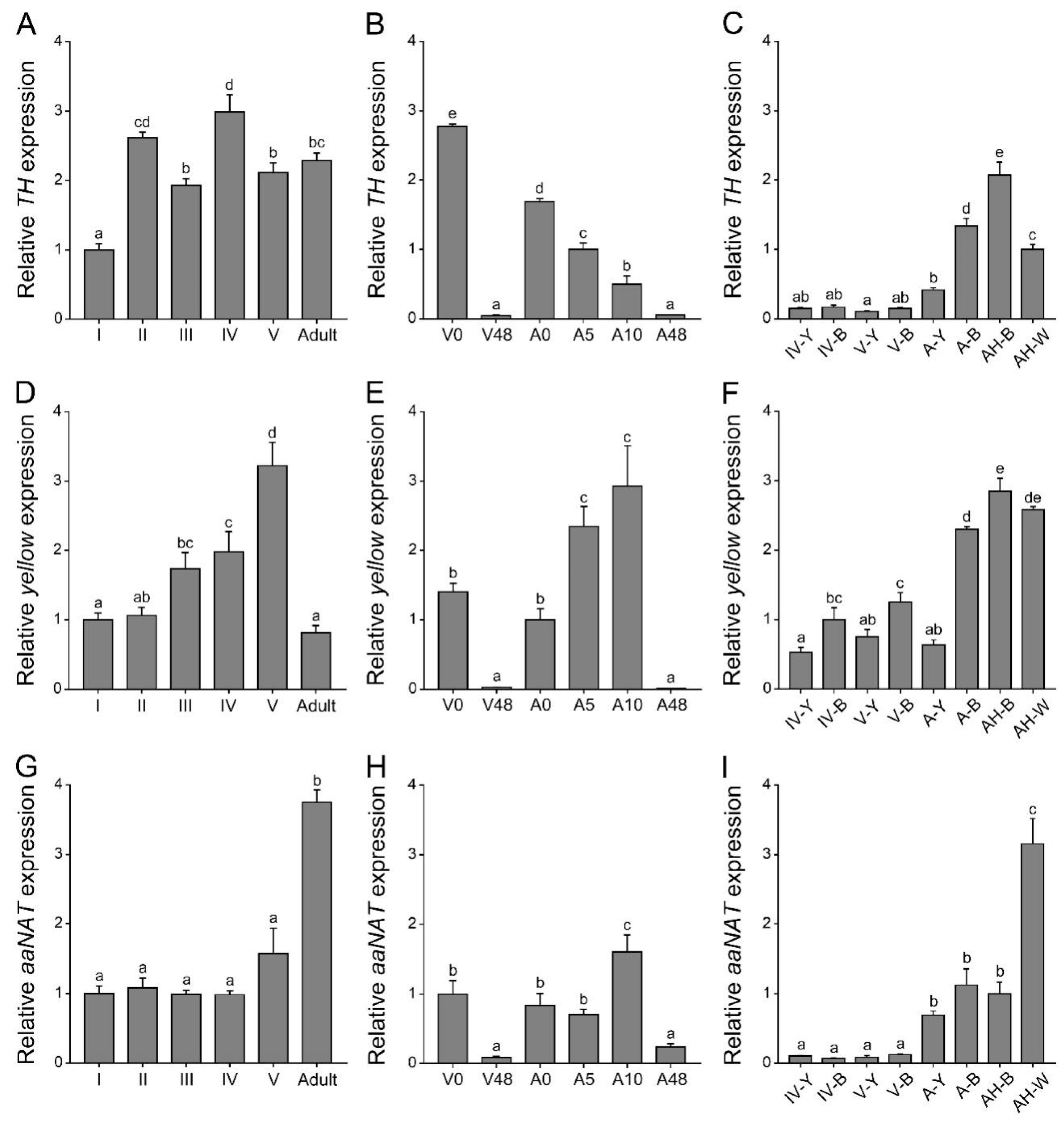
2.1. Expression Patterns of Three Melanin Pathway Genes (TH, yellow, and aaNAT)
2.2. Functional Analyses of TH and yellow
2.3. Functional Analyses of aaNAT
3. Discussion
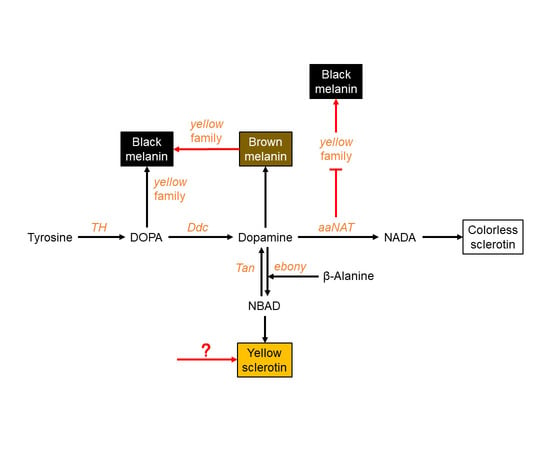
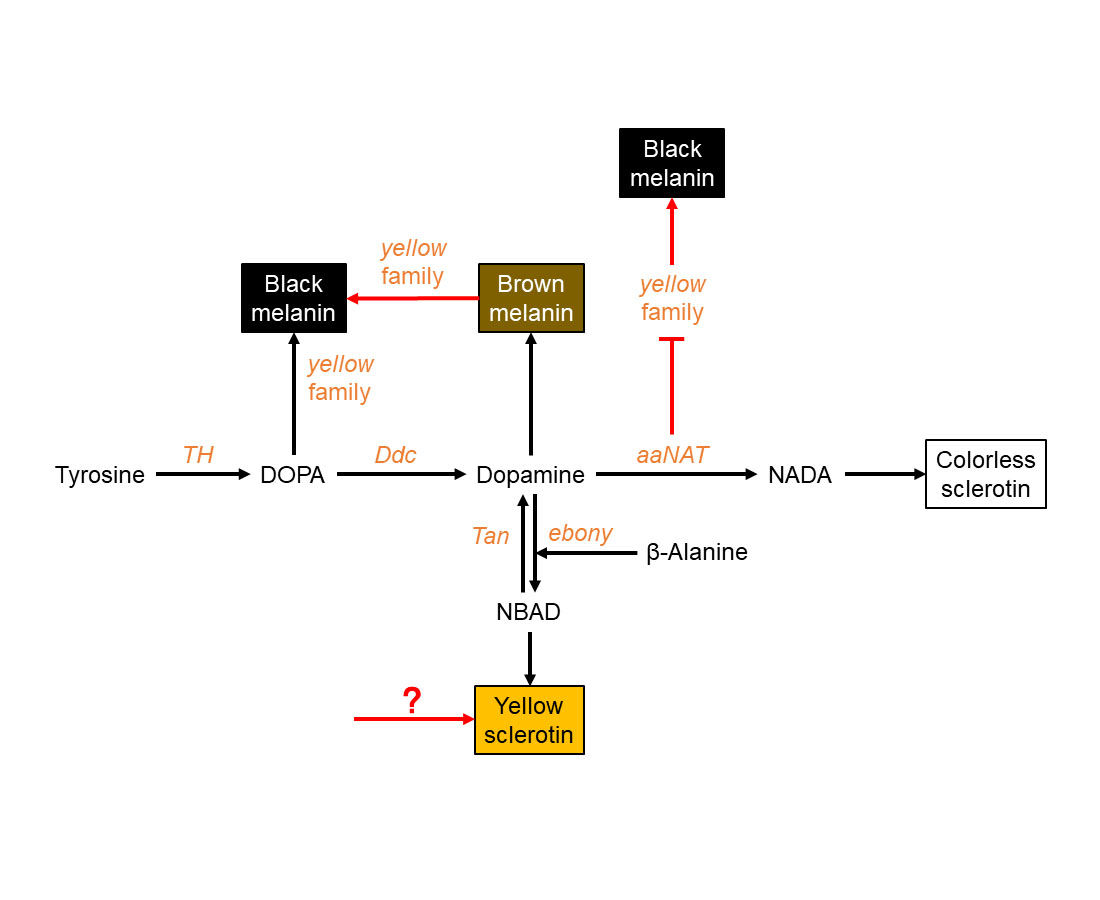
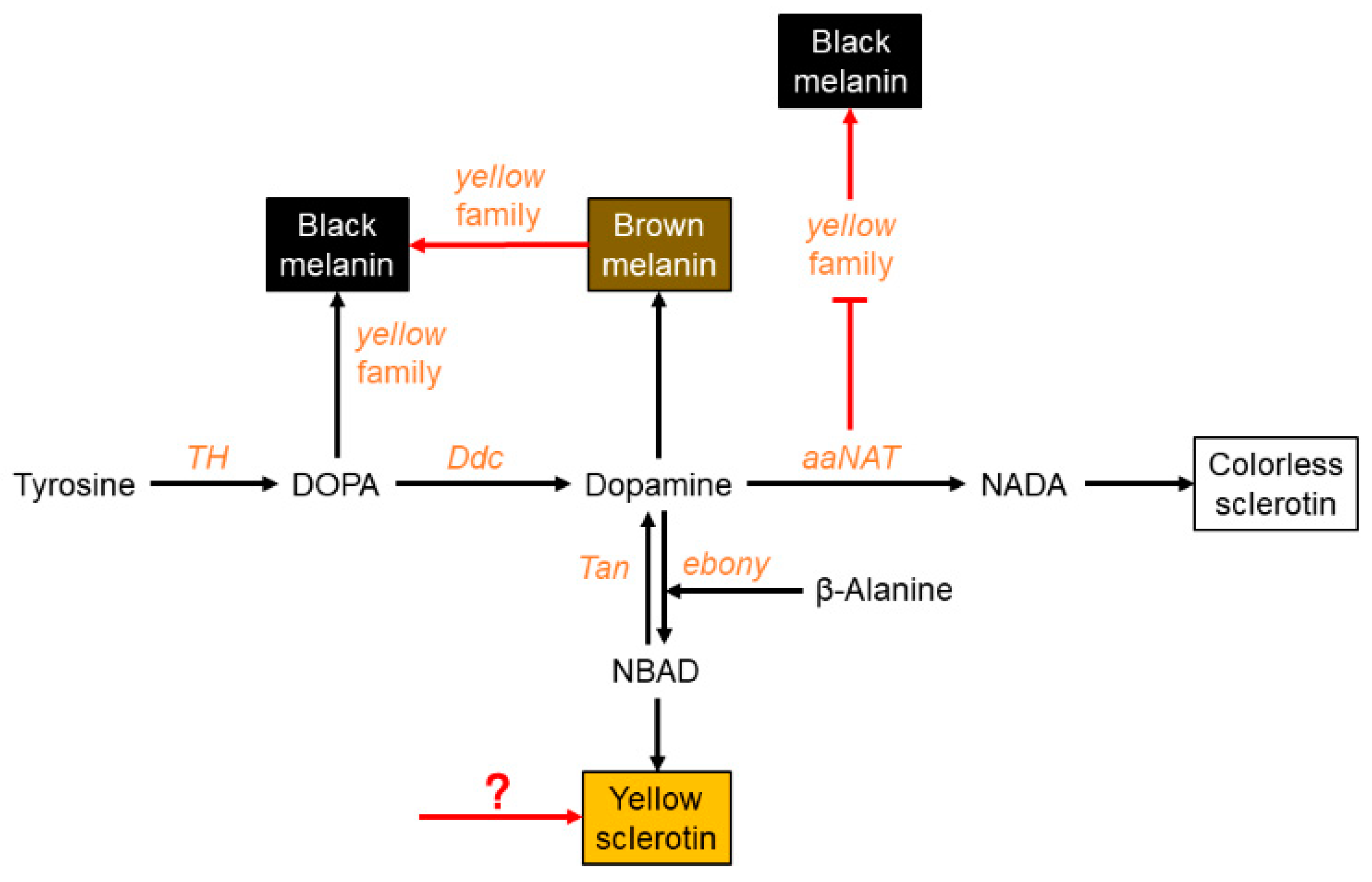
3.1. Black Pigment Is Solely Generated by the Melanin Pathway
3.2. Brown Pigment Could Be Further Converted to Black Melanin
3.3. aaNAT Suppresses the Formation of Black Melanin to Generate Colorless Sclerotin
3.4. A Modified Melanin Pathway Suitable for P. biguttatus
4. Materials and Methods
4.1. Insects
4.2. Total RNA Extraction and cDNA Synthesis
4.3. RNA Interference
4.4. Quantitative Real-Time PCR
4.5. Image Processing
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| aaNAT | arylalkylamine-N-acetyltransferase |
| bp(s) | base pair(s) |
| Cq | quantification cycle |
| Ddc | DOPA decarboxylase |
| DOPA | dihydroxyphenylalanine |
| dsRNA | double-stranded RNA |
| EF1α | elongation factor 1 alpha |
| GFP | green fluorescent protein |
| NADA | N-acetyl dopamine |
| NBAD | N-β-alanyl dopamine |
| NTC | no template controls |
| qPCR | quantitative real-time PCR |
| RNAi | RNA interference |
| TH | tyrosine hydroxylase |
References
- True, J.R. Insect melanism: The molecules matter. Trends Ecol. Evol. 2003, 18, 640–647. [Google Scholar] [CrossRef]
- Wittkopp, P.J.; Beldade, P. Development and evolution of insect pigmentation: Genetic mechanisms and the potential consequences of pleiotropy. Semin. Cell Dev. Biol. 2009, 20, 65–71. [Google Scholar] [CrossRef]
- Nijhout, H.F. The Development and Evolution of Butterfly Wing Patterns; Smithsonian Institution Press: Washington, DC, USA, 1991; p. 297. [Google Scholar]
- Maranda, B.; Hodgetts, R. A characterization of dopamine acetyltransferase in Drosophila melanogaster. Insect Biochem. 1977, 7, 33–43. [Google Scholar] [CrossRef]
- Wittkopp, P.J.; Carroll, S.B.; Kopp, A. Evolution in black and white: Genetic control of pigment patterns in Drosophila. Trends Genet. 2003, 19, 495–504. [Google Scholar] [CrossRef]
- Wittkopp, P.J.; Williams, B.L.; Selegue, J.E.; Carroll, S.B. Drosophila pigmentation evolution: Divergent genotypes underlying convergent phenotypes. Proc. Natl. Acad. Sci. USA 2003, 100, 1808–1813. [Google Scholar] [CrossRef]
- Wright, T.R. The genetics of biogenic amine metabolism, sclerotization, and melanization in Drosophila melanogaster. Adv. Genet. 1987, 24, 127–222. [Google Scholar] [CrossRef]
- Wittkopp, P.J.; True, J.R.; Carroll, S.B. Reciprocal functions of the Drosophila yellow and ebony proteins in the development and evolution of pigment patterns. Development 2002, 129, 1849–1858. [Google Scholar]
- Wittkopp, P.J.; Vaccaro, K.; Carroll, S.B. Evolution of yellow gene regulation and pigmentation in Drosophila. Curr. Biol. 2002, 12, 1547–1556. [Google Scholar] [CrossRef]
- True, J.R.; Yeh, S.D.; Hovemann, B.T.; Kemme, T.; Meinertzhagen, I.A.; Edwards, T.N.; Liou, S.R.; Han, Q.; Li, J. Drosophila tan encodes a novel hydrolase required in pigmentation and vision. PLoS Genet. 2005, 1, e63. [Google Scholar] [CrossRef]
- Jeong, S.; Rebeiz, M.; Andolfatto, P.; Werner, T.; True, J.; Carroll, S.B. The evolution of gene regulation underlies a morphological difference between two Drosophila sister species. Cell 2008, 132, 783–793. [Google Scholar] [CrossRef]
- Wittkopp, P.J.; Stewart, E.E.; Arnold, L.L.; Neidert, A.H.; Haerum, B.K.; Thompson, E.M.; Akhras, S.; Smith-Winberry, G.; Shefner, L. Intraspecific polymorphism to interspecific divergence: Genetics of pigmentation in Drosophila. Science 2009, 326, 540–544. [Google Scholar] [CrossRef]
- Arakane, Y.; Lomakin, J.; Beeman, R.W.; Muthukrishnan, S.; Gehrke, S.H.; Kanost, M.R.; Kramer, K.J. Molecular and functional analyses of amino acid decarboxylases involved in cuticle tanning in Tribolium castaneum. J. Biol. Chem. 2009, 284, 16584–16594. [Google Scholar] [CrossRef]
- Arakane, Y.; Dittmer, N.T.; Tomoyasu, Y.; Kramer, K.J.; Muthukrishnan, S.; Beeman, R.W.; Kanost, M.R. Identification, mRNA expression and functional analysis of several yellow family genes in Tribolium castaneum. Insect Biochem. Mol. Biol. 2010, 40, 259–266. [Google Scholar] [CrossRef]
- Tomoyasu, Y.; Arakane, Y.; Kramer, K.J.; Denell, R.E. Repeated co-options of exoskeleton formation during wing-to-elytron evolution in beetles. Curr. Biol. 2009, 19, 2057–2065. [Google Scholar] [CrossRef]
- Gorman, M.J.; Arakane, Y. Tyrosine hydroxylase is required for cuticle sclerotization and pigmentation in Tribolium castaneum. Insect Biochem. Mol. Biol. 2010, 40, 267–273. [Google Scholar] [CrossRef]
- Noh, M.Y.; Kramer, K.J.; Muthukrishnan, S.; Beeman, R.W.; Kanost, M.R.; Arakane, Y. Loss of function of the yellow-e gene causes dehydration-induced mortality of adult Tribolium castaneum. Dev. Biol. 2015, 399, 315–324. [Google Scholar] [CrossRef]
- Koch, P.B.; Behnecke, B. The molecular basis of melanism and mimicry in a swallowtail butterfly. Curr. Biol. 2000, 10, 591–594. [Google Scholar] [CrossRef]
- Futahashi, R.; Fujiwara, H. Melanin-synthesis enzymes coregulate stage-specific larval cuticular markings in the swallowtail butterfly, Papilio xuthus. Dev. Genes Evol. 2005, 215, 519–529. [Google Scholar] [CrossRef]
- Niu, B.L.; Shen, W.F.; Liu, Y.; Weng, H.B.; He, L.H.; Mu, J.J.; Wu, Z.L.; Jiang, P.; Tao, Y.Z.; Meng, Z.Q. Cloning and RNAi-mediated functional characterization of MaLac2 of the pine sawyer, Monochamus alternatus. Insect Mol. Biol. 2008, 17, 303–312. [Google Scholar] [CrossRef]
- Futahashi, R.; Banno, Y.; Fujiwara, H. Caterpillar color patterns are determined by a two-phase melanin gene prepatterning process: New evidence from tan and laccase2. Evol. Dev. 2010, 12, 157–167. [Google Scholar] [CrossRef]
- Elias-Neto, M.; Soares, M.P.; Simoes, Z.L.; Hartfelder, K.; Bitondi, M.M. Developmental characterization, function and regulation of a Laccase2 encoding gene in the honey bee, Apis mellifera (Hymenoptera, Apinae). Insect Biochem. Mol. Biol. 2010, 40, 241–251. [Google Scholar] [CrossRef]
- Ferguson, L.C.; Maroja, L.; Jiggins, C.D. Convergent, modular expression of ebony and tan in the mimetic wing patterns of Heliconius butterflies. Dev. Genes Evol. 2011, 221, 297–308. [Google Scholar] [CrossRef]
- Mehere, P.; Han, Q.; Christensen, B.M.; Li, J. Identification and characterization of two arylalkylamine N-acetyltransferases in the yellow fever mosquito, Aedes aegypti. Insect Biochem. Mol. Biol. 2011, 41, 707–714. [Google Scholar] [CrossRef]
- Dai, F.Y.; Qiao, L.; Tong, X.L.; Cao, C.; Chen, P.; Chen, J.; Lu, C.; Xiang, Z.H. Mutations of an arylalkylamine N-acetyltransferase, Bm-iAANAT, are responsible for silkworm melanism mutant. J. Biol. Chem. 2010, 285, 19553–19560. [Google Scholar] [CrossRef]
- Zhan, S.; Guo, Q.; Li, M.; Li, M.; Li, J.; Miao, X.; Huang, Y. Disruption of an N-acetyltransferase gene in the silkworm reveals a novel role in pigmentation. Development 2010, 137, 4083–4090. [Google Scholar] [CrossRef]
- Osanai-Futahashi, M.; Ohde, T.; Hirata, J.; Uchino, K.; Futahashi, R.; Tamura, T.; Niimi, T.; Sezutsu, H. A visible dominant marker for insect transgenesis. Nat. Commun. 2012, 3, 1295. [Google Scholar] [CrossRef]
- Noh, M.Y.; Koo, B.; Kramer, K.J.; Muthukrishnan, S.; Arakane, Y. Arylalkylamine N-acetyltransferase 1 gene (TcAANAT1) is required for cuticle morphology and pigmentation of the adult red flour beetle, Tribolium castaneum. Insect Biochem. Mol. Biol. 2016, 79, 119–129. [Google Scholar] [CrossRef]
- Liu, J.; Lemonds, T.R.; Popadic, A. The genetic control of aposematic black pigmentation in hemimetabolous insects: Insights from Oncopeltus fasciatus. Evol. Dev. 2014, 16, 270–277. [Google Scholar] [CrossRef]
- Liu, J.; Lemonds, T.R.; Marden, J.H.; Popadic, A. A pathway analysis of melanin patterning in a Hemimetabolous insect. Genetics 2016, 203, 403–413. [Google Scholar] [CrossRef]
- Lemonds, T.R.; Liu, J.; Popadić, A. The contribution of the melanin pathway to overall body pigmentation during ontogenesis of Periplaneta americana. Insect Sci. 2016, 23, 513–519. [Google Scholar] [CrossRef]
- Singh, I.K.; Singh, S.; Mogilicherla, K.; Shukla, J.N.; Palli, S.R. Comparative analysis of double-stranded RNA degradation and processing in insects. Sci. Rep. 2017, 7, 17059. [Google Scholar] [CrossRef]
- Li, Z.; Zeng, B.; Ling, L.; Xu, J.; You, L.; Aslam, A.F.; Tan, A.; Huang, Y. Enhancement of larval RNAi efficiency by over-expressing Argonaute2 in Bombyx mori. Int. J. Biol. Sci. 2015, 11, 176. [Google Scholar] [CrossRef]
- Angelini, D.R.; Kaufman, T.C. Functional analyses in the milkweed bug Oncopeltus fasciatus (Hemiptera) support a role for Wnt signaling in body segmentation but not appendage development. Dev. Biol. 2005, 283, 409–423. [Google Scholar] [CrossRef]
- Moussian, B. Recent advances in understanding mechanisms of insect cuticle differentiation. Insect Biochem. Mol. Biol. 2010, 40, 363–375. [Google Scholar] [CrossRef]
- Kronforst, M.R.; Barsh, G.S.; Kopp, A.; Mallet, J.; Monteiro, A.; Mullen, S.P.; Protas, M.; Rosenblum, E.B.; Schneider, C.J.; Hoekstra, H.E. Unraveling the thread of nature’s tapestry: The genetics of diversity and convergence in animal pigmentation. Pigment. Cell Melanoma Res. 2012, 25, 411–433. [Google Scholar] [CrossRef]
- Walter, M.F.; Black, B.C.; Afshar, G.; Kermabon, A.Y.; Wright, T.R.; Biessmann, H. Temporal and spatial expression of the yellow gene in correlation with cuticle formation and DOPA decarboxylase activity in Drosophila development. Dev. Biol. 1991, 147, 32–45. [Google Scholar] [CrossRef]
- Johnson, J.; Li, J.; Christensen, B. Cloning and characterization of a dopachrome conversion enzyme from the yellow fever mosquito, Aedes aegypti. Insect Biochem. Mol. Biol. 2001, 31, 1125–1135. [Google Scholar] [CrossRef]
- Han, Q.; Fang, J.M.; Ding, H.Z.; Johnson, J.K.; Christensen, B.M.; Li, J.Y. Identification of Drosophila melanogaster yellow-f and yellow-f2 proteins as dopachrome-conversion enzymes. Biochem. J. 2002, 368, 333–340. [Google Scholar] [CrossRef]
- Futahashi, R.; Fujiwara, H. Regulation of 20-hydroxyecdysone on the larval pigmentation and the expression of melanin synthesis enzymes and yellow gene of the swallowtail butterfly, Papilio xuthus. Insect Biochem. Mol. Biol. 2007, 37, 855–864. [Google Scholar] [CrossRef]
- Futahashi, R.; Sato, J.; Meng, Y.; Okamoto, S.; Daimon, T.; Yamamoto, K.; Suetsugu, Y.; Narukawa, J.; Takahashi, H.; Banno, Y.; et al. Yellow and ebony are the responsible genes for the larval color mutants of the silkworm Bombyx mori. Genetics 2008, 180, 1995–2005. [Google Scholar] [CrossRef]
- Ito, K.; Katsuma, S.; Yamamoto, K.; Kadono-Okuda, K.; Mita, K.; Shimada, T. Yellow-e determines the color pattern of larval head and tail spots of the silkworm Bombyx mori. J. Biol. Chem. 2010, 285, 5624–5629. [Google Scholar] [CrossRef]
- Hintermann, E.; Jenö, P.; Meyer, U.A. Isolation and characterization of an arylalkylamine N-acetyltransferase from Drosophila melanogaster. FEBS Lett. 1995, 375, 148–150. [Google Scholar] [CrossRef]
- Brodbeck, D.; Amherd, R.; Callaerts, P.; Hintermann, E.; Meyer, U.A.; Affolter, M. Molecular and biochemical characterization of the aaNAT1 (Dat) locus in Drosophila melanogaster: Differential expression of two gene products. DNA Cell Biol. 1998, 17, 621–633. [Google Scholar] [CrossRef]
- Walter, M.F.; Zeineh, L.L.; Black, B.C.; McIvor, W.E.; Wright, T.R.; Biessmann, H. Catecholamine metabolism and in vitro induction of premature cuticle melanization in wild type and pigmentation mutants of Drosophila melanogaster. Arch. Insect Biochem. Physiol. 1996, 31, 219–233. [Google Scholar] [CrossRef]
- Andersen, S.O. Insect cuticular sclerotization: A review. Insect Biochem. Mol. Biol. 2010, 40, 166–178. [Google Scholar] [CrossRef]
- Gibert, J.M.; Peronnet, F.; Schlotterer, C. Phenotypic plasticity in Drosophila pigmentation caused by temperature sensitivity of a chromatin regulator network. PLoS Genet. 2007, 3, e30. [Google Scholar] [CrossRef]
- Fujiwara, H.; Ishikawa, H. Molecular mechanism of introduction of the hidden break into the 28S rRNA of insects: Implication based on structural studies. Nucleic Acids Res. 1986, 14, 6393–6401. [Google Scholar] [CrossRef]
- Chesebro, J.; Hrycaj, S.; Mahfooz, N.; Popadić, A. Diverging functions of Scr between embryonic and post-embryonic development in a hemimetabolous insect, Oncopeltus fasciatus. Dev. Biol. 2009, 329, 142–151. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Vandesompele, J. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]
- Bustin, S.A.; Beaulieu, J.F.; Huggett, J.; Jaggi, R.; Kibenge, F.S.; Olsvik, P.A.; Toegel, S. MIQE précis: Practical implementation of minimum standard guidelines for fluorescence-based quantitative real-time PCR experiments. BMC Mol. Biol. 2010, 11, 74. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Li, H.; Du, J.; Zhang, J.; Shen, J.; Cai, W. Three Melanin Pathway Genes, TH, yellow, and aaNAT, Regulate Pigmentation in the Twin-Spotted Assassin Bug, Platymeris biguttatus (Linnaeus). Int. J. Mol. Sci. 2019, 20, 2728. https://doi.org/10.3390/ijms20112728
Zhang Y, Li H, Du J, Zhang J, Shen J, Cai W. Three Melanin Pathway Genes, TH, yellow, and aaNAT, Regulate Pigmentation in the Twin-Spotted Assassin Bug, Platymeris biguttatus (Linnaeus). International Journal of Molecular Sciences. 2019; 20(11):2728. https://doi.org/10.3390/ijms20112728
Chicago/Turabian StyleZhang, Yinqiao, Hu Li, Juan Du, Junzheng Zhang, Jie Shen, and Wanzhi Cai. 2019. "Three Melanin Pathway Genes, TH, yellow, and aaNAT, Regulate Pigmentation in the Twin-Spotted Assassin Bug, Platymeris biguttatus (Linnaeus)" International Journal of Molecular Sciences 20, no. 11: 2728. https://doi.org/10.3390/ijms20112728
APA StyleZhang, Y., Li, H., Du, J., Zhang, J., Shen, J., & Cai, W. (2019). Three Melanin Pathway Genes, TH, yellow, and aaNAT, Regulate Pigmentation in the Twin-Spotted Assassin Bug, Platymeris biguttatus (Linnaeus). International Journal of Molecular Sciences, 20(11), 2728. https://doi.org/10.3390/ijms20112728