Reduced Expression of CbUFO Is Associated with the Phenotype of a Flower-Defective Cosmos bipinnatus

Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
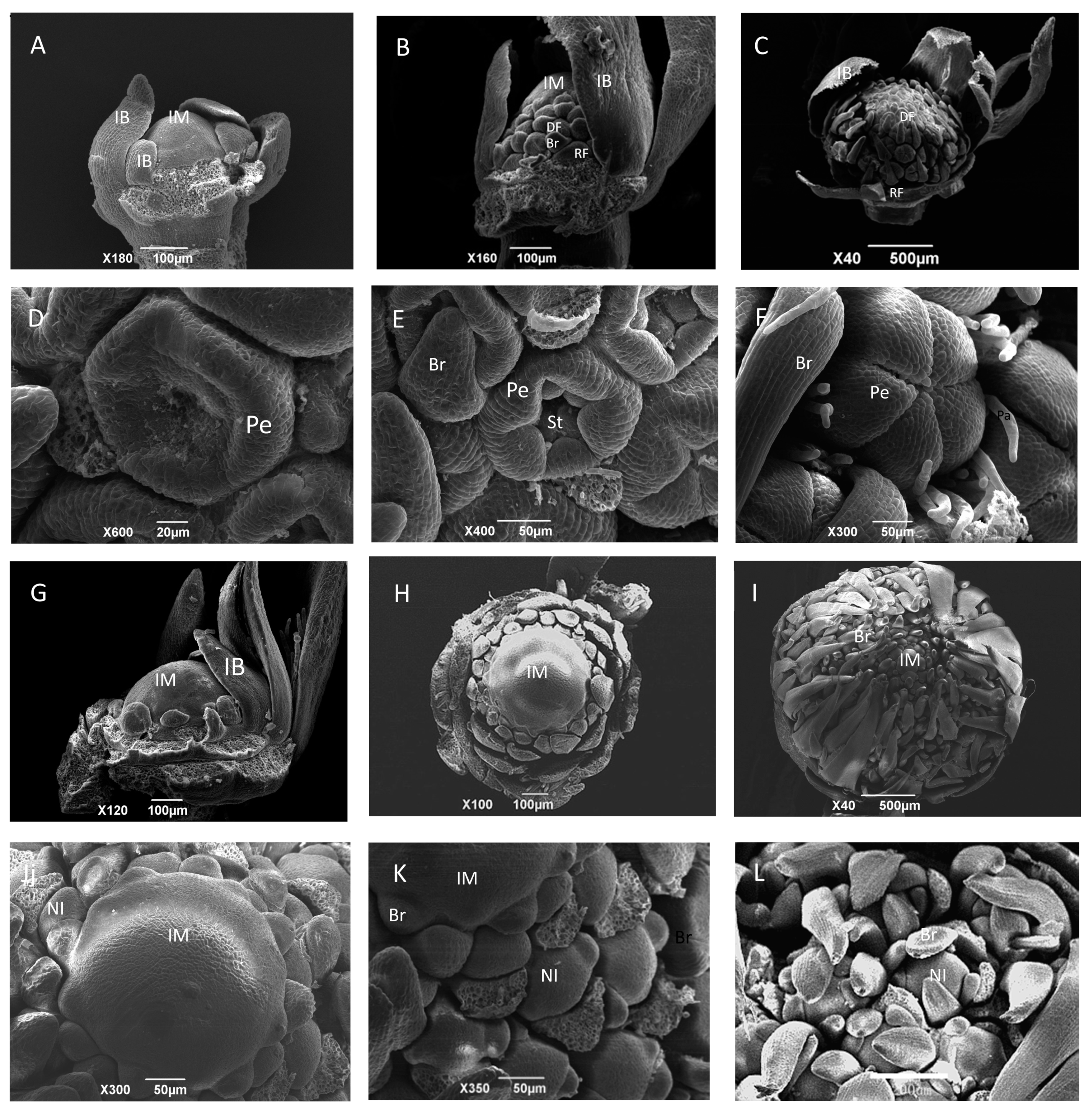
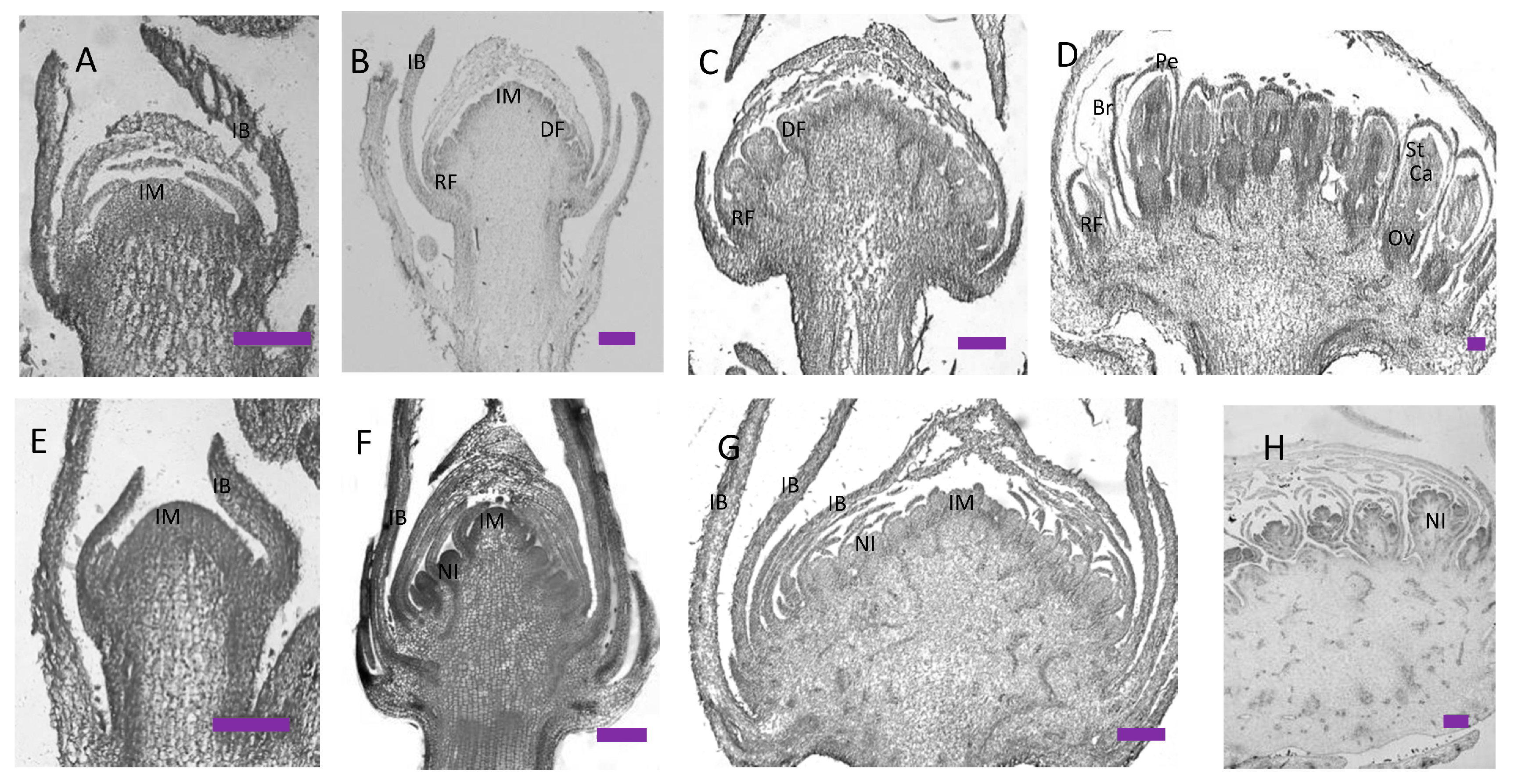
2.1. Morphological and Ontogenetic Characteristics of Wild-Type and gh Cosmos
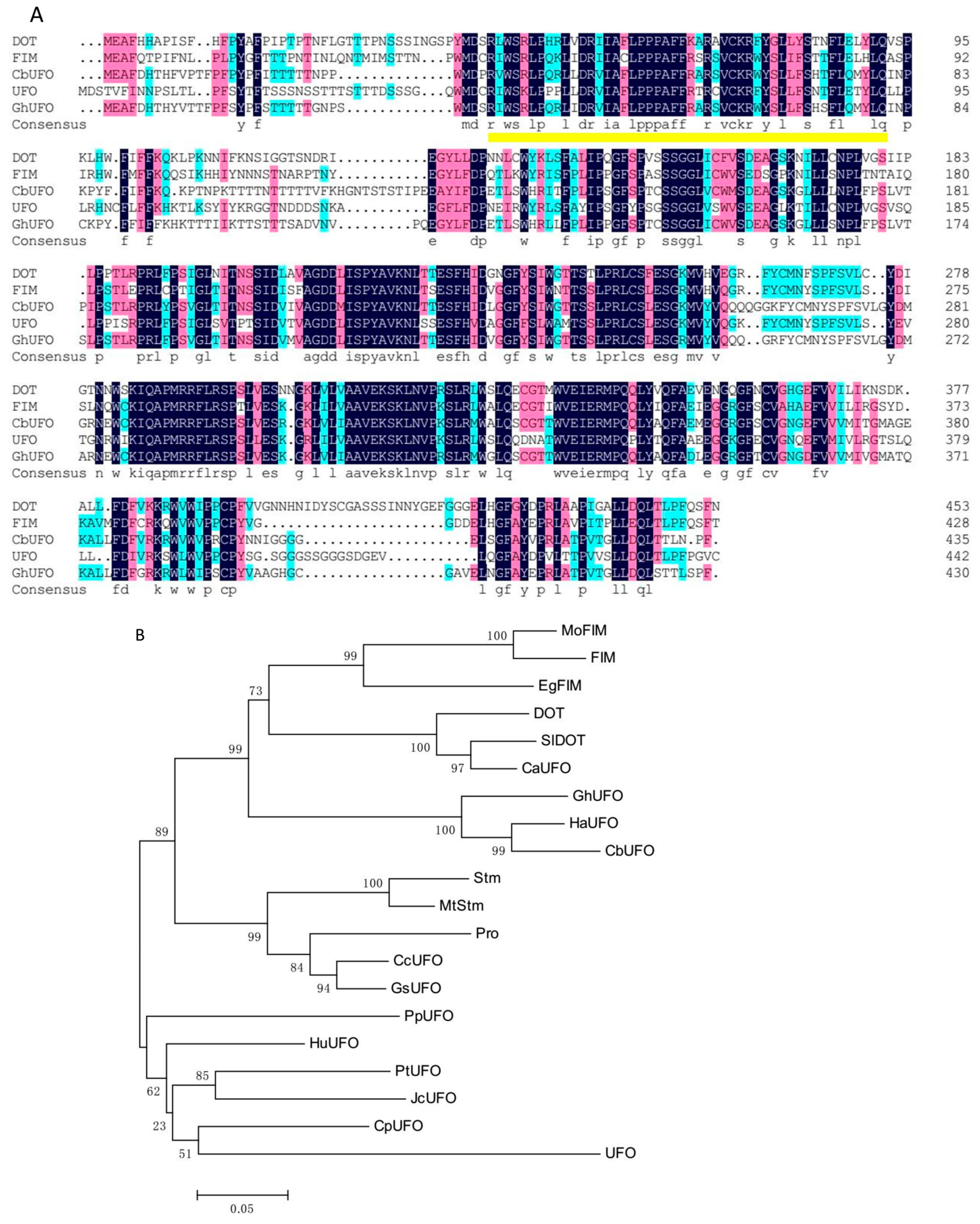
2.2. Sequence Analysis of LFY and UFO Orthologs in Wild-Type and gh Plants
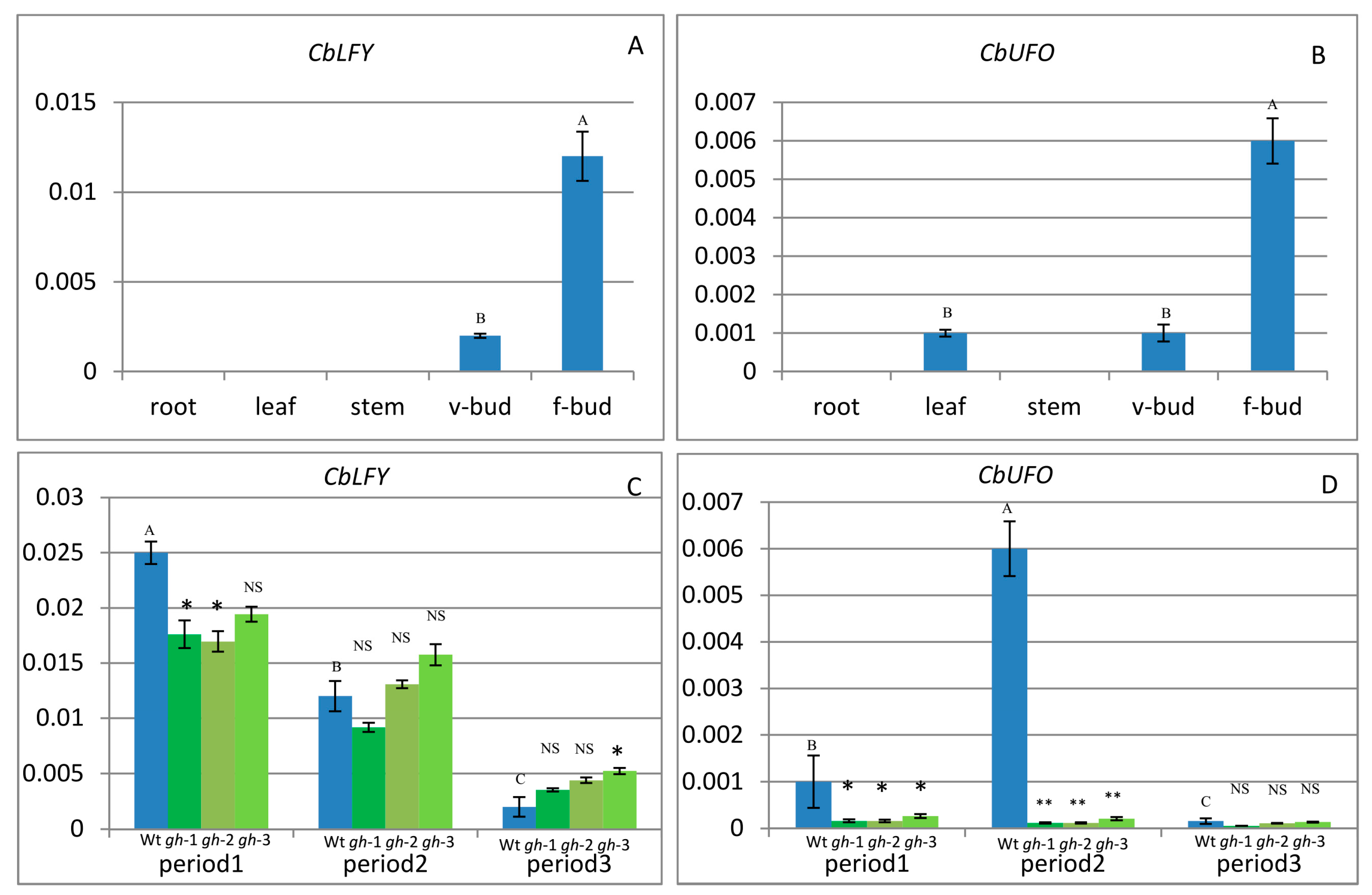
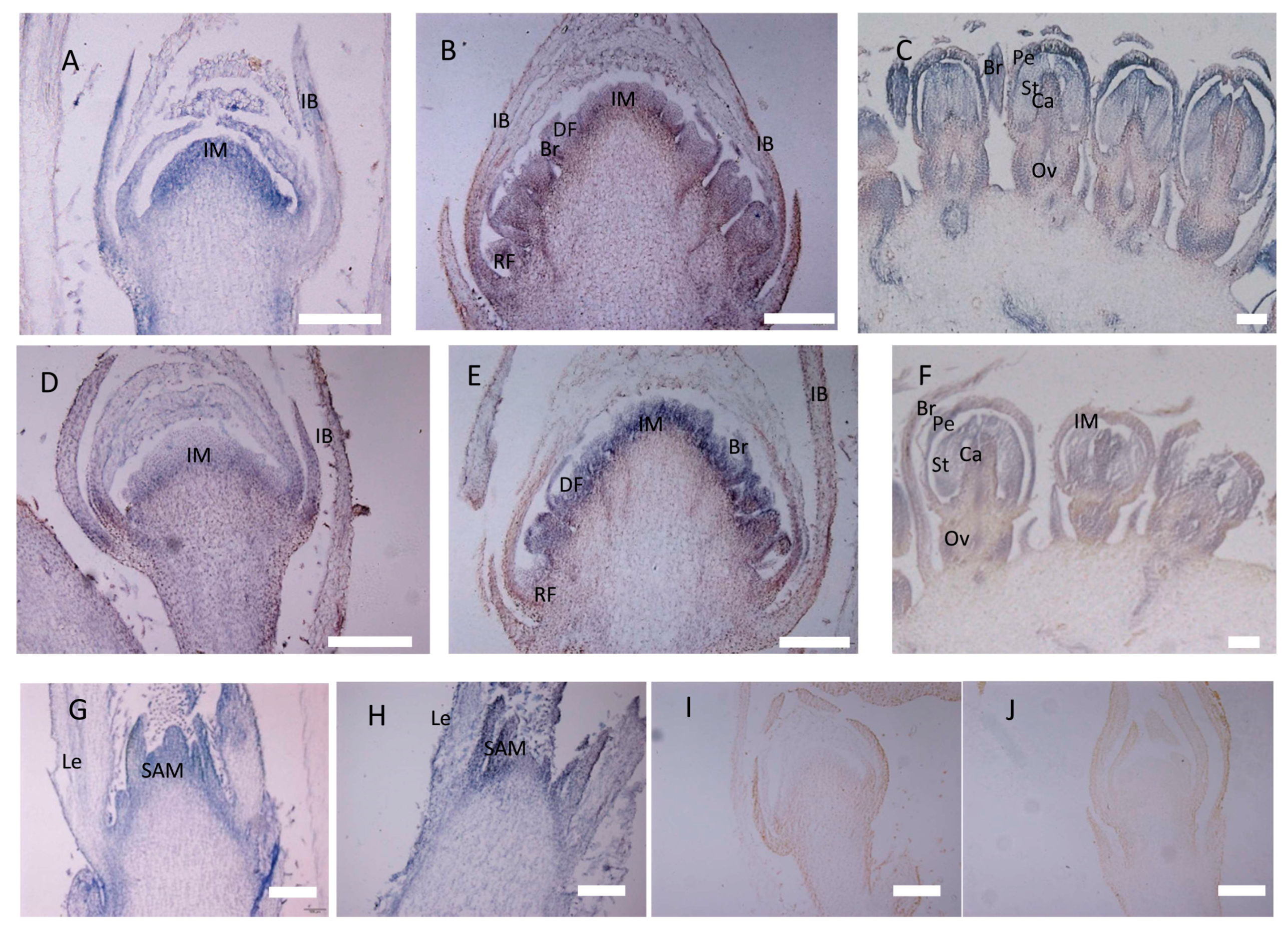
2.3. Expression of CbLFY and CbUFO in Wild-Type Cosmos
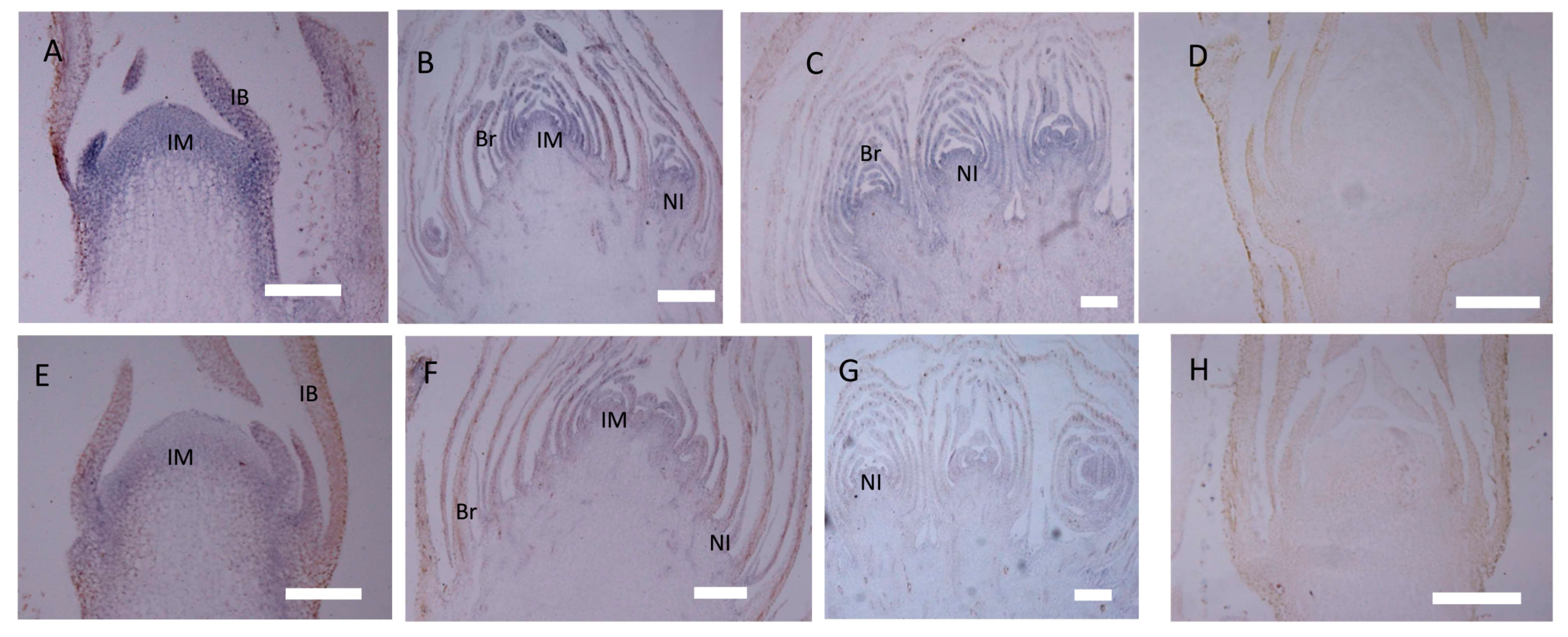
2.4. Expression of CbLFY and CbUFO in gh Cosmos
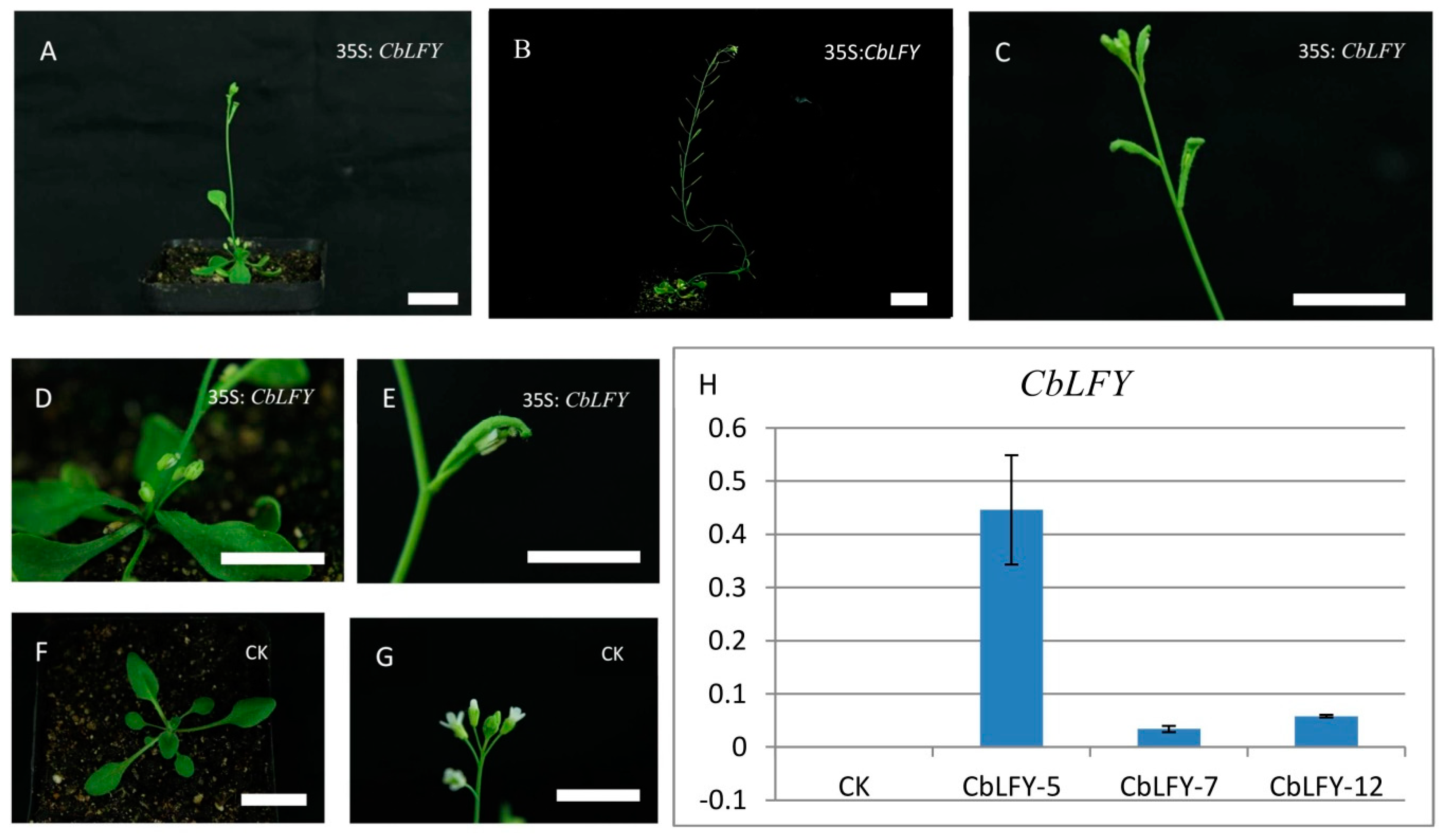
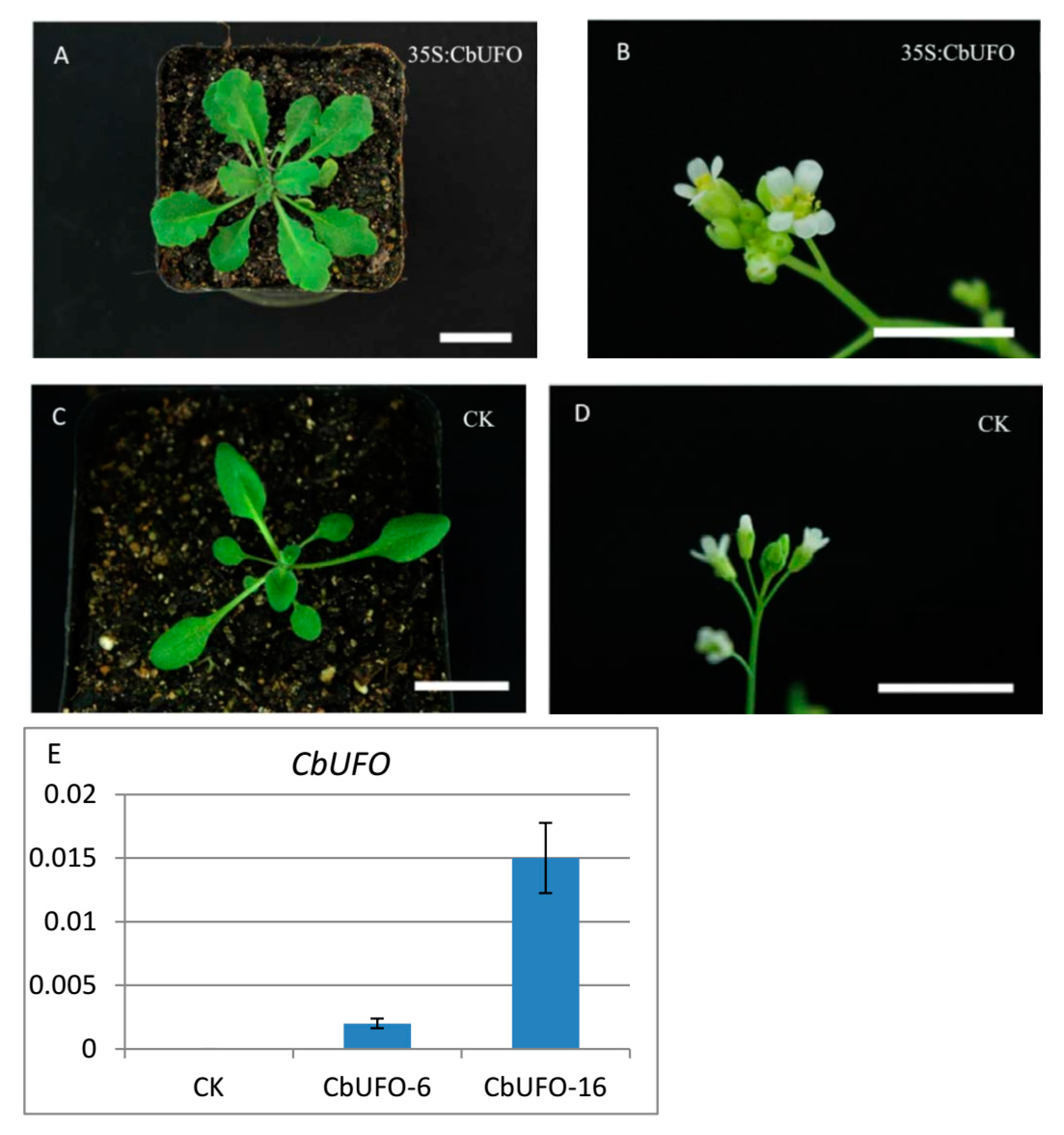
2.5. Transgenic Analysis
3. Discussion
3.1. Potential Causes for the gh Mutation
3.2. Functional Evolution of LFY- and UFO-like Genes in Asteraceae
4. Methods and Materials
4.1. Plant Material
4.2. Stereomicroscope Observation
4.3. Histological Sections
4.4. Scanning Electron Microscopy
4.5. Nucleic Acid Isolation
4.6. Cloning of CbLFY and CbUFO
4.7. Phylogenetic Analysis
4.8. The qPCR Analysis
4.9. In Situ Hybridization
4.10. Transgenic Analysis
4.11. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| UFO | unusual floral organs |
| LFY | leafy |
| TFL1 | terminal flower 1 |
| AP1 | apetala1 |
| DOT | double middle |
| IM | inflorescence meristem |
| FM | floral meristem |
| FUM | floral unit meristem |
| FMI | floral meristem identity |
| RT-PCR | reverse transcription PCR |
| qPCR | real-time quantitative RT-PCR |
References
- Jack, T. Molecular and genetic mechanisms of floral control. Plant Cell 2004, 16, S1–S17. [Google Scholar] [CrossRef]
- Benlloch, R.; Berbel, A.; Serrano-Mislata, A.; Madueno, F. Floral initiation and inflorescence architecture: A comparative view. Ann. Bot. 2007, 100, 659–676. [Google Scholar] [CrossRef]
- McKim, S.; Hay, A. Patterning and evolution of floral structures - marking time. Curr. Opin. Genet. Dev. 2010, 20, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Wellmer, F.; Riechmann, J.L. Gene networks controlling the initiation of flower development. Trends Genet. 2010, 26, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Castel, R.; Kusters, E.; Koes, R. Inflorescence development in petunia: Through the maze of botanical terminology. J. Exp. Bot. 2010, 61, 2235–2246. [Google Scholar] [CrossRef]
- Teo, Z.W.; Song, S.; Wang, Y.Q.; Liu, J.; Yu, H. New insights into the regulation of inflorescence architecture. Trends Plant Sci. 2014, 19, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Prusinkiewicz, P.; Erasmus, Y.; Lane, B.; Harder, L.D.; Coen, E. Evolution and Development of Inflorescence Architectures. Science 2007, 316, 1452–1456. [Google Scholar] [CrossRef]
- Liljegren, S.J.; Gustafson-Brown, C.; Pinyopich, A.; Ditta, G.S.; Yanofsky, M.F. Interactions among APETALA1, LEAFY, and TERMINAL FLOWER1 Specify Meristem Fate. Plant Cell 1999, 11, 1007–1018. [Google Scholar] [CrossRef] [PubMed]
- Chandler, J.W.; Werr, W. Arabidopsis floral phytomer development: Auxin response relative to biphasic modes of organ initiation. J. Exp. Bot. 2014, 65, 3097–3110. [Google Scholar] [CrossRef]
- Weigel, D.; Alvarez, J.; Smyth, D.R.; Yanofsky, M.F.; Meyerowitz, E.M. LEAFY Controls Floral Meristem Identity in Arabidopsis. Plant Cell 1992, 69, 843–859. [Google Scholar] [CrossRef]
- Zhang, S.; Sandal, N.; Polowick, P.L.; Stiller, J.; Stougaard, J.; Fobert, P.R. Proliferating Floral Organs (Pfo), a Lotus japonicus gene required for specifying floral meristem determinacy and organ identity, encodes an F-box protein. Plant J. 2003, 33, 607–619. [Google Scholar] [CrossRef] [PubMed]
- Allen, K.D.; Sussex, I.M. Falsiflora and anantha control early stages of floral meristem development in tomato (Lycopersicon esculentum Mill.). Planta 1996, 200, 254–264. [Google Scholar] [CrossRef]
- Souer, E.; Rebocho, A.B.; Bliek, M.; Kusters, E.; de Bruin, R.A.; Koes, R. Patterning of inflorescences and flowers by the F-Box protein DOUBLE TOP and the LEAFY homolog ABERRANT LEAF AND FLOWER of petunia. Plant Cell 2008, 20, 2033–2048. [Google Scholar] [CrossRef]
- Chae, E.; Tan, Q.K.; Hill, T.A.; Irish, V.F. An Arabidopsis F-box protein acts as a transcriptional co-factor to regulate floral development. Development 2008, 135, 1235–1245. [Google Scholar] [CrossRef]
- Levin, J.Z.; Meyerowitz, E.M. UFO: An Arabidopsis gene involved in both floral meristem and floral organ development. Plant Cell 1995, 7, 529–548. [Google Scholar] [CrossRef]
- Wilkinson, M.D.; Haughn, G.W. UNUSUAL FLORAL ORGANS Controls Meristem Identity and Organ Primordia Fate in Arabidopsis. Plant Cell 1995, 7, 1485–1499. [Google Scholar] [CrossRef]
- Broholm, S.K.; Teeri, T.H.; Elomaa, P. Molecular Control of Inflorescence Development in Asteraceae; Advances in Botanical Research; Academic Press: Oxford, UK, 2014; Volume 72, pp. 297–333. [Google Scholar]
- Zhao, Y.; Zhang, T.; Broholm, S.K.; Tahtiharju, S.; Mouhu, K.; Albert, V.A.; Teeri, T.H.; Elomaa, P. Evolutionary Co-Option of Floral Meristem Identity Genes for Patterning of the Flower-Like Asteraceae Inflorescence. Plant Physiol. 2016, 172, 284–296. [Google Scholar] [PubMed]
- Ibanez, M.S.; Mercado, M.I.; Coll Araoz, M.V.; Zannier, M.L.; Grau, A.; Ponessa, G.I. Flower structure and developmental stages of the capitulum of Smallanthus sonchifolius (Asteraceae): Reproductive implications. J. Plant Res. 2017, 130, 327–337. [Google Scholar] [CrossRef]
- Classen-Bockhoff, R.; Bull-Herenu, K. Towards an ontogenetic understanding of inflorescence diversity. Ann. Bot. 2013, 112, 1523–1542. [Google Scholar] [CrossRef] [PubMed]
- Pozner, R.; Zanotti, C.; Johnson, L.A. Evolutionary origin of the Asteraceae capitulum: Insights from Calyceraceae. Am. J. Bot. 2012, 99, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kanen, C.; Gill, N.; Kingm, G.; Bowersj, E.; Berges, H.; Gouzy, J.; Bachlava, E.; Langladen, B.; Lai, Z.; Stewart, M. Progress towards a reference genome for sunflower. Botanique 2011, 89, 429–437. [Google Scholar] [CrossRef]
- Wang, Y.; Ma, Y.; Fu, J.; Qi, S.; Ma, H.; Dai, S. Isolation and functional analysis of the ClM8 - FRUITFULL -like MADS-box gene from Chrysanthemum lavandulifolium. Sci. Hortic. 2013, 161, 125–133. [Google Scholar] [CrossRef]
- Risseeuw, E.; Venglat, P.; Xiang, D.; Komendant, K.; Daskalchuk, T.; Babic, V.; Crosby, W.; Datla, R. An activated form of UFO alters leaf development and produces ectopic floral and inflorescence meristems. PLoS ONE 2013, 8, e83807. [Google Scholar] [CrossRef] [PubMed]
- Coen, E.S.; Romero, J.M.; Doyle, S.; Elliott, R.; Murphy, G.; Carpenter, R. Floricaula: A homeotic gene required for flower development in antirrhinum majus. Cell 1990, 63, 1311–1322. [Google Scholar] [CrossRef]
- Souer, E.; van der Krol, A.; Kloos, D.; Spelt, C.; Bliek, M.; Mol, J.; Koes, R. Genetic control of branching pattern and floral identity during Petunia inflorescence development. Development 1998, 125, 733–742. [Google Scholar]
- Chuck, G.; Whipple, C.; Jackson, D.; Hake, S. The maize SBP-box transcription factor encoded by tasselsheath4 regulates bract development and the establishment of meristem boundaries. Development 2010, 137, 1243–1250. [Google Scholar] [CrossRef]
- Dinneny, J.R.; Yadegari, R.; Fischer, R.L.; Yanofsky, M.F.; Weigel, D. The role of JAGGED in shaping lateral organs. Development 2004, 131, 1101–1110. [Google Scholar] [CrossRef]
- Hepworth, S.R.; Klenz, J.E.; Haughn, G.W. UFO in the Arabidopsis inflorescence apex is required for floral-meristem identity and bract suppression. Planta 2006, 223, 769–778. [Google Scholar] [CrossRef]
- Yamaguchi, A.; Wu, M.F.; Yang, L.; Wu, G.; Poethig, R.S.; Wagner, D. The microRNA regulated SBP-box transcription factor SPL3 is a direct upstream activator of LEAFY, FRUITFULL, and APETALA1. Dev. Cell 2009, 17, 268–278. [Google Scholar] [CrossRef]
- Jungeun, L.; Mijin, O.; Hanna, P.; Ilha, L. SOC1 translocated to the nucleus by interaction with AGL24 directly regulates leafy. Plant J. 2010, 55, 832–843. [Google Scholar]
- Prenner, G.; Vergara-Silva, F.; Rudall, P.J. The key role of morphology in modelling inflorescence architecture. Trends Plant Sci. 2009, 14, 302–309. [Google Scholar] [CrossRef]
- Zhineng, L.; Guofeng, L.; Jiaqi, Z.; Junwei, Z.; Manzhu, B. Extraction of high-quality tissue-specific RNA from London plane trees (Platanus acerifolia), permitting the construction of a female inflorescence cDNA library. Funct. Plant Biol. 2008, 35, 159–165. [Google Scholar]
- Wang, Z.J.; Huang, J.Q.; Huang, Y.J.; Chen, F.F.; Zheng, B.S. Cloning and Characterization of a Homologue of the FLORICAULA/LEAFY Gene in Hickory (Carya cathayensis Sarg). Plant Mol. Biol. Report. 2011, 30, 794–805. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- O’neill, J.W.; Bier, E. Double-label in situ hybridization using biotin and digoxigenin-tagged RNA probes. BioTechniques 1994, 17, 870–874. [Google Scholar]
- Ma, Y.P.; Fang, X.H.; Chen, F.; Dai, S.L. DFL, a FLORICAULA/LEAFY homologue gene from Dendranthema lavandulifolium is expressed both in the vegetative and reproductive tissues. Plant Cell Rep. 2008, 27, 647–654. [Google Scholar] [CrossRef]
- Braissant, O.; Wahli, W. A simplified in situ hybridization protocol using non-radioactively labeled probes to detect abundant and rare mRNAs on tissue sections. Int. J. Food Contamination 1998, 2, 1–8. [Google Scholar]
- Neta, R.; David-Schwartz, R.; Peretz, Y.; Sela, I.; Rabinowitch, H.D.; Flaishman, M.; Kamenetsky, R. Flower development in garlic: The ups and downs of gaLFY expression. Planta 2011, 233, 1063–1072. [Google Scholar] [CrossRef]
- Clough, B.; Atilola, F.A.; Black, J.; Pasvol, G. Plasmodium falciparum: The importance of IgM in the rosetting of parasite-infected erythrocytes. Exp. Parasitol. 1998, 89, 129–132. [Google Scholar] [CrossRef]










© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, F.; Lan, W.; Zhou, Q.; Liu, B.; Chen, F.; Zhang, S.; Bao, M.; Liu, G. Reduced Expression of CbUFO Is Associated with the Phenotype of a Flower-Defective Cosmos bipinnatus. Int. J. Mol. Sci. 2019, 20, 2503. https://doi.org/10.3390/ijms20102503
Li F, Lan W, Zhou Q, Liu B, Chen F, Zhang S, Bao M, Liu G. Reduced Expression of CbUFO Is Associated with the Phenotype of a Flower-Defective Cosmos bipinnatus. International Journal of Molecular Sciences. 2019; 20(10):2503. https://doi.org/10.3390/ijms20102503
Chicago/Turabian StyleLi, Fei, Wu Lan, Qin Zhou, Baojun Liu, Feng Chen, Sisi Zhang, Manzhu Bao, and Guofeng Liu. 2019. "Reduced Expression of CbUFO Is Associated with the Phenotype of a Flower-Defective Cosmos bipinnatus" International Journal of Molecular Sciences 20, no. 10: 2503. https://doi.org/10.3390/ijms20102503
APA StyleLi, F., Lan, W., Zhou, Q., Liu, B., Chen, F., Zhang, S., Bao, M., & Liu, G. (2019). Reduced Expression of CbUFO Is Associated with the Phenotype of a Flower-Defective Cosmos bipinnatus. International Journal of Molecular Sciences, 20(10), 2503. https://doi.org/10.3390/ijms20102503