Promotion of Cell Death in Cisplatin-Resistant Ovarian Cancer Cells through KDM1B-DCLRE1B Modulation


Abstract
:1. Introduction
2. Results
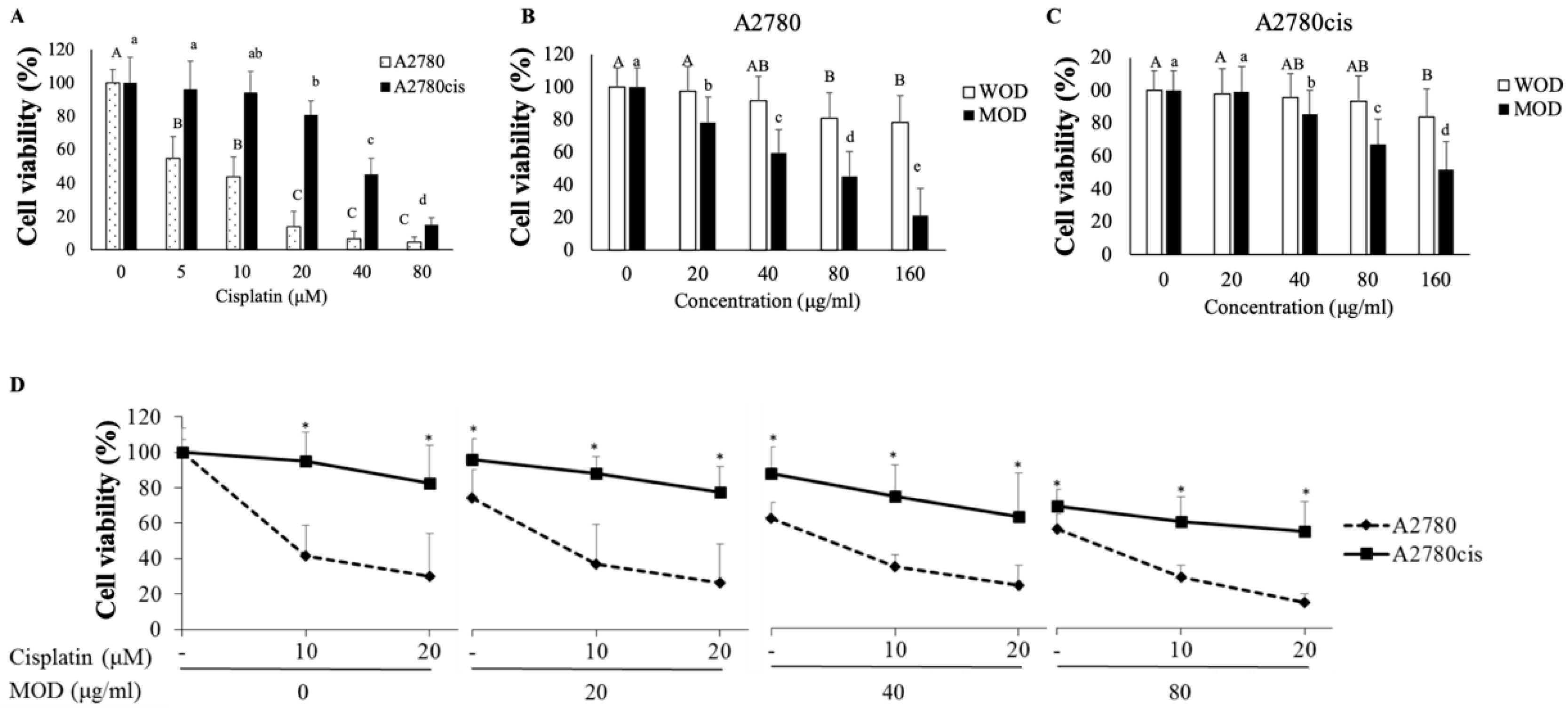
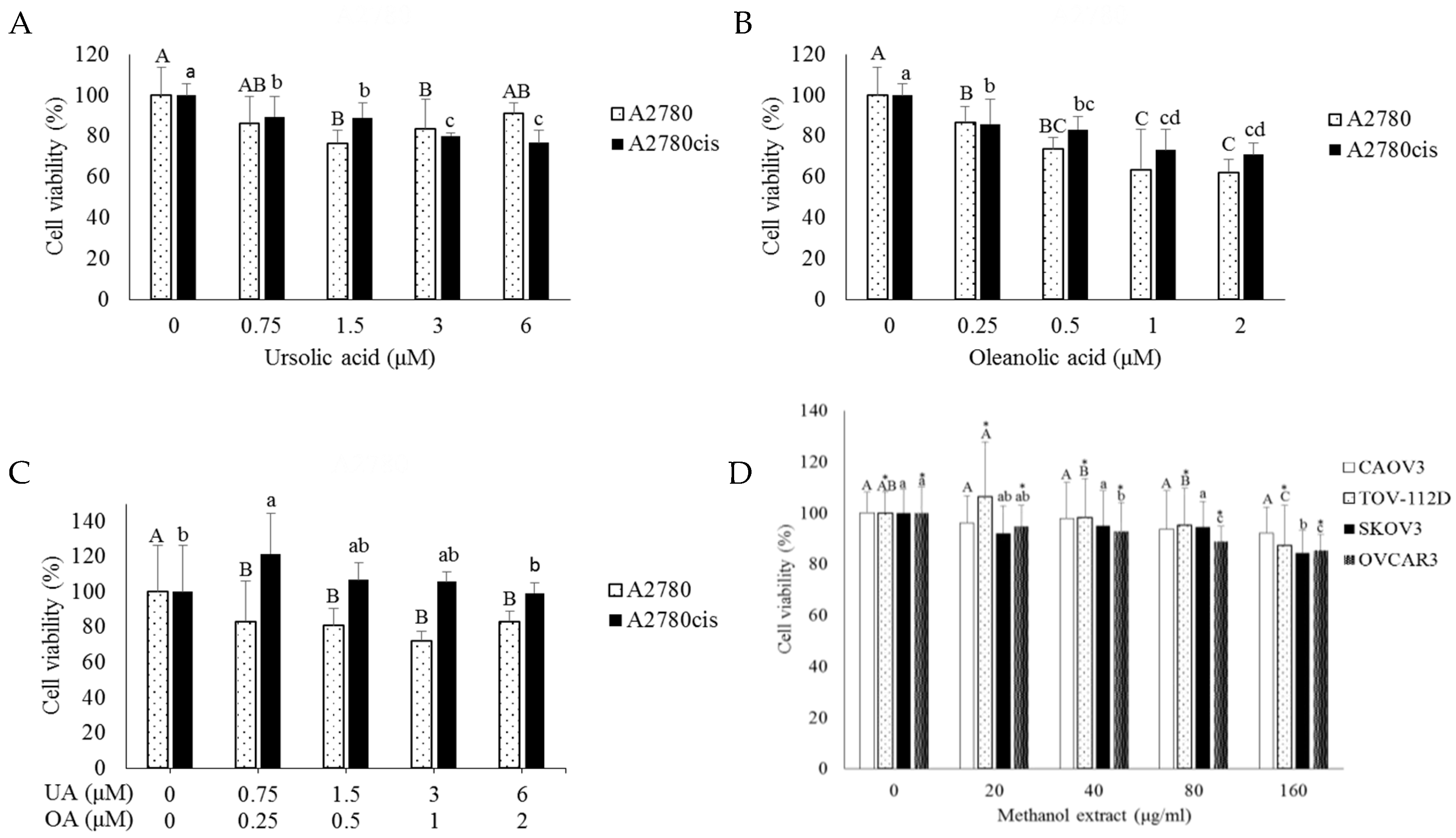
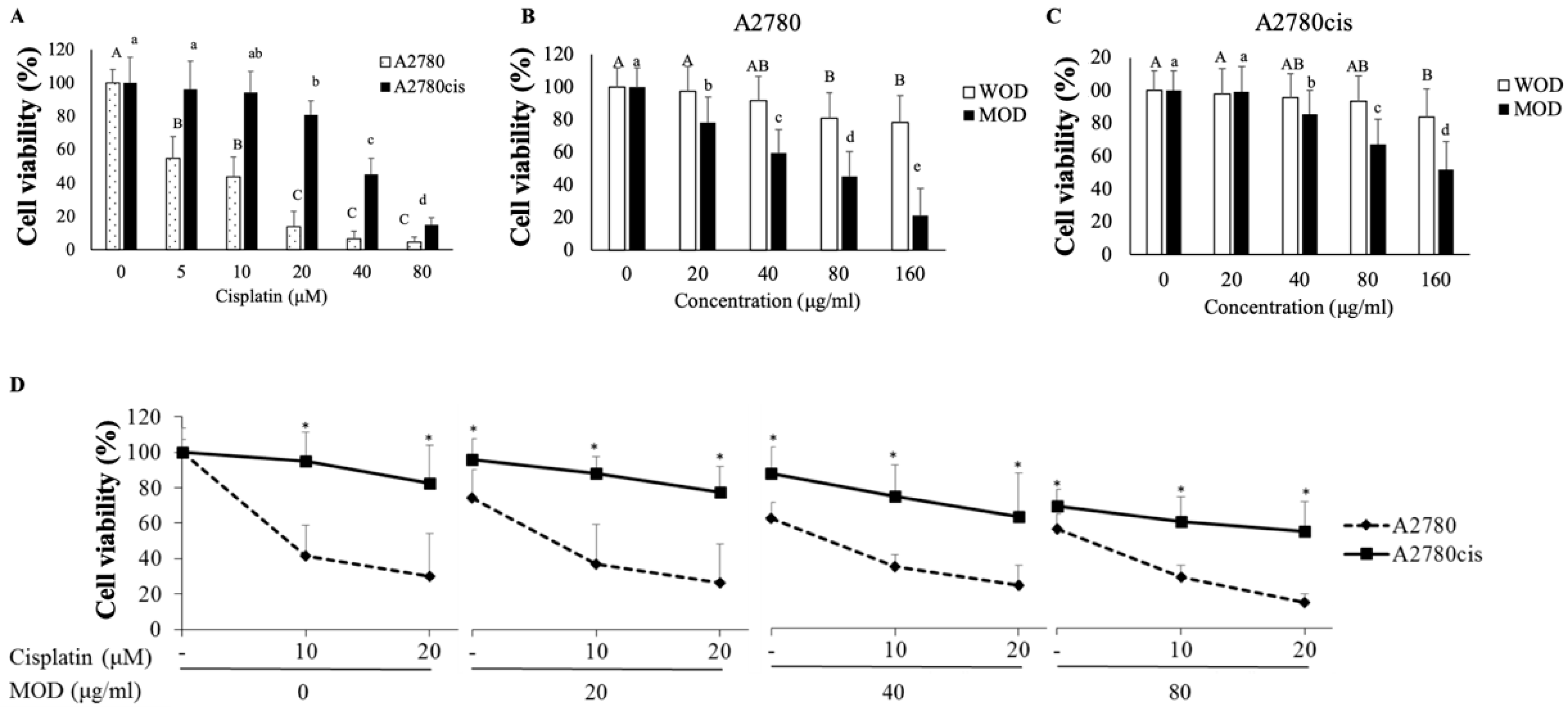
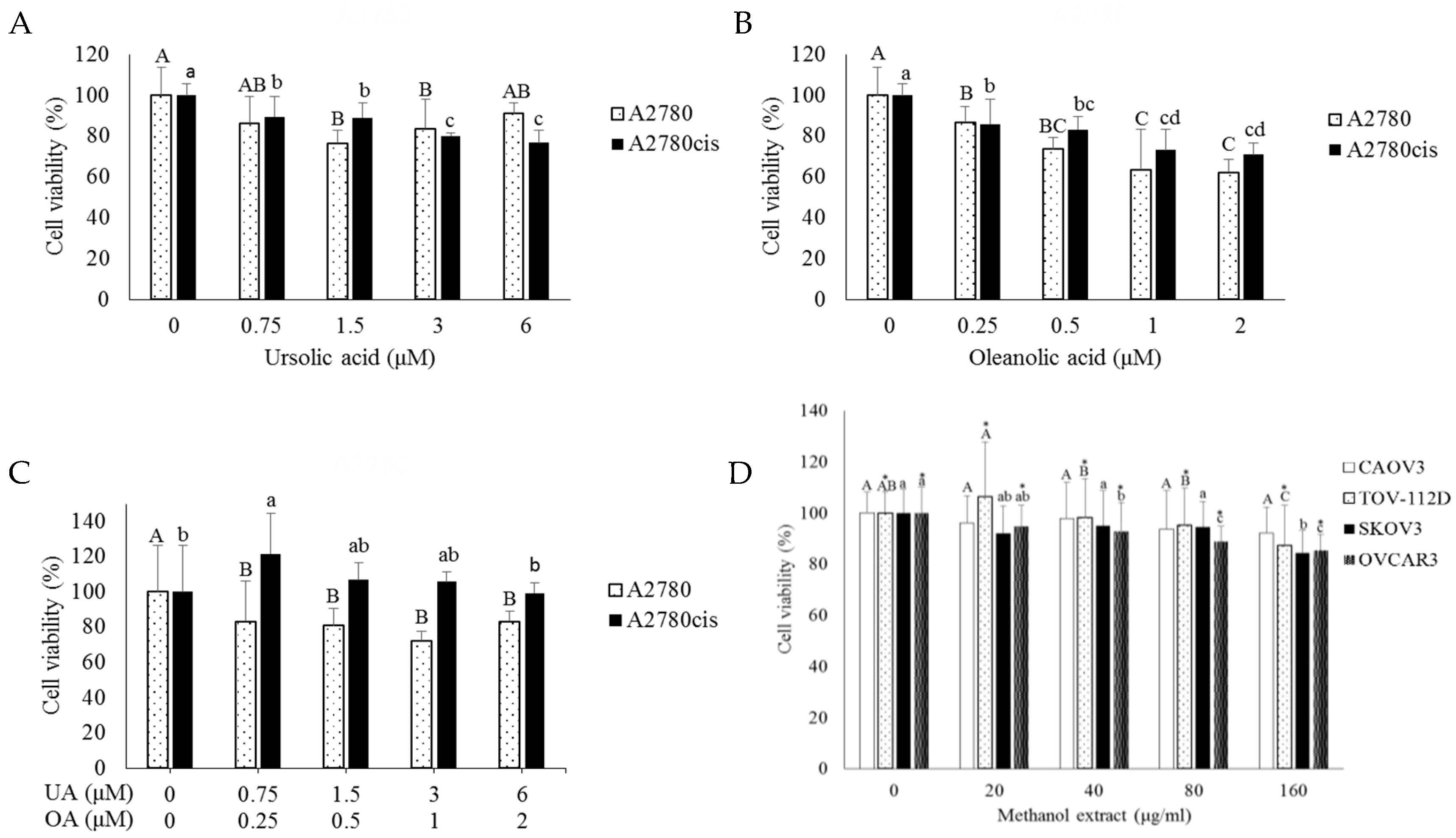
2.1. Combination Treatment with Cisplatin and O. diffusa Extracts Reduces Cell Viability
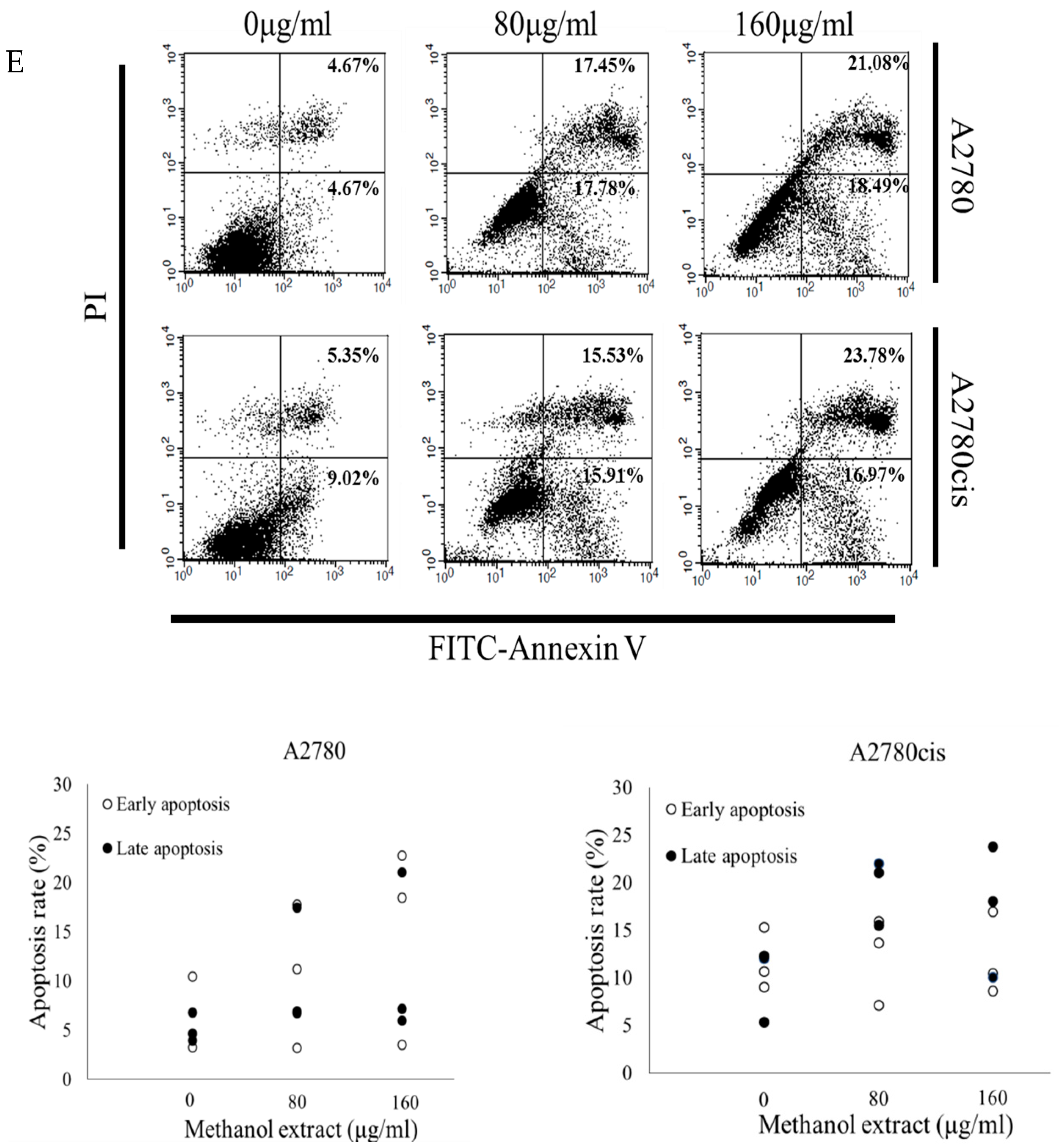
2.2. MOD Treatment Induces Apoptosis in Ovarian Cancer Cells
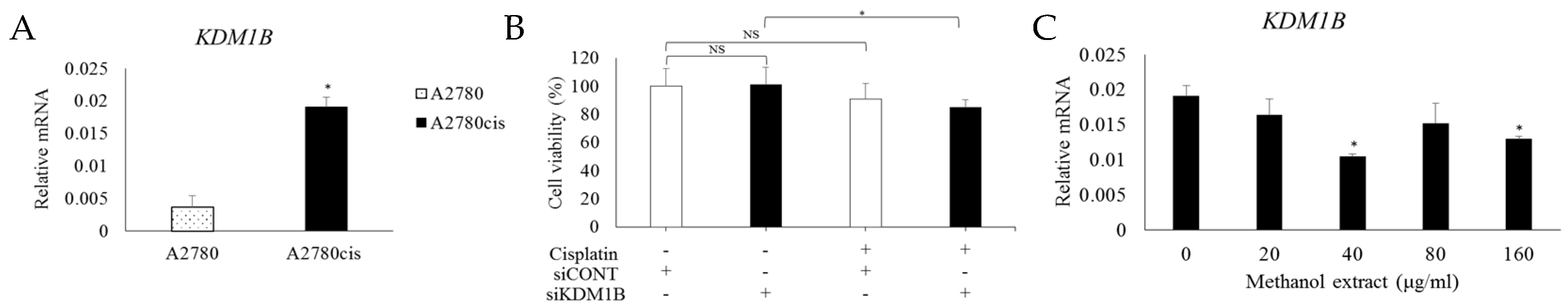
2.3. Downregulation of KDM1B Induces Cisplatin Sensitivity in CRC
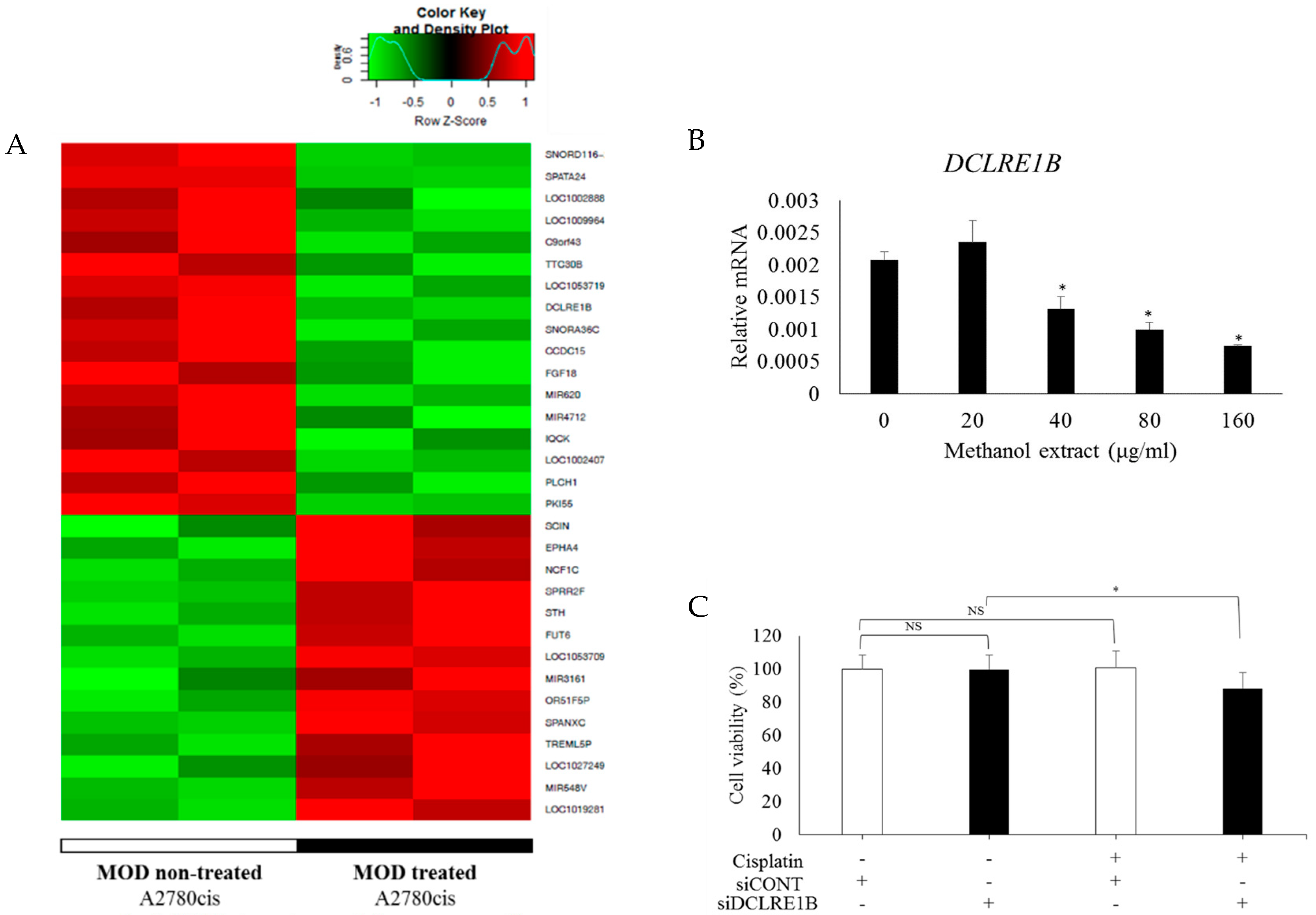
2.4. Downregulation of DCLRE1B is a Factor of CRC Death
2.5. MOD Treatment Induced Cell Death of CRC by Regulating KDM1B and DCLRE1B Expression
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Preparation of O. diffusa Extract
4.3. High-Performance Liquid Chromatography (HPLC)
4.4. MTT Assay for Cell Viability
4.5. Apoptotic Detection by Flow Cytometry
4.6. Microarray and Data Analysis
4.7. siRNA Transfection Assay
4.8. Reverse Transcription and Quantitative RT-PCR
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ACN | Acetonitrile |
| cDNA | Complementary DNA |
| CRC | Cisplatin-resistant ovarian cancer cells |
| CSC | Cisplatin-sensitive ovarian cancer cells |
| DEG | Differentially expressed gene |
| DMSO | Dimethyl sulphoxide |
| DNMTs | DNA methyltransferases |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| HDACs | Histone decaetylases |
| HPLC | High-performance liquid chromatography |
| H3K4me1/2 | Mono- and dimethyl-lysine 4 of histone H3 |
| ICL | Inter-strand crosslinks |
| LSD1/KDM1A | Lysine-specific demethylase 1 |
| MOD | Methanol extract of OD |
| MTT | 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazoliumbromide |
| PBS | Phosphate-buffered saline |
| PI | Propidium iodide |
| OA | Oleanolic acid |
| OD | Oldenlandia diffusa |
| SNM1B/DCLRE1B | DNA cross-link repair 1B |
| UA | Ursolic acid |
| WOD | Water extract of OD |
References
- Tang, W.; Hemm, I.; Bertram, B. Recent development of antitumor agents from Chinese herbal medicines. Part II. High molecular compounds(3). Planta Med. 2003, 69, 193–201. [Google Scholar] [CrossRef]
- Leung, A.Y. Traditional toxicity documentation of Chinese Materia Medica--an overview. Toxicol. Pathol. 2006, 34, 319–326. [Google Scholar] [CrossRef]
- Gu, G.; Barone, I.; Gelsomino, L.; Giordano, C.; Bonofiglio, D.; Statti, G.; Menichini, F.; Catalano, S.; Ando, S. Oldenlandia diffusa extracts exert antiproliferative and apoptotic effects on human breast cancer cells through ERalpha/Sp1-mediated p53 activation. J. Cell Physiol. 2012, 227, 3363–3372. [Google Scholar] [CrossRef]
- Gupta, S.; Zhang, D.; Yi, J.; Shao, J. Anticancer activities of Oldenlandia diffusa. J. Herb. Pharm. 2004, 4, 21–33. [Google Scholar] [CrossRef]
- Ovesna, Z.; Vachalkova, A.; Horvathova, K.; Tothova, D. Pentacyclic triterpenoic acids: New chemoprotective compounds. Minireview. Neoplasma 2004, 51, 327–333. [Google Scholar] [PubMed]
- Sunwoo, Y.Y.; Lee, J.H.; Jung, H.Y.; Jung, Y.J.; Park, M.S.; Chung, Y.A.; Maeng, L.S.; Han, Y.M.; Shin, H.S.; Lee, J.; et al. Oldenlandia diffusa Promotes Antiproliferative and Apoptotic Effects in a Rat Hepatocellular Carcinoma with Liver Cirrhosis. Evid. Based Complement. Altern. Med. 2015, 2015, 501508. [Google Scholar] [CrossRef] [PubMed]
- Pu, F.; Chen, F.; Lin, S.; Chen, S.; Zhang, Z.; Wang, B.; Shao, Z. The synergistic anticancer effect of cisplatin combined with Oldenlandia diffusa in osteosarcoma MG-63 cell line in vitro. Onco Targets Ther. 2016, 9, 255–263. [Google Scholar] [Green Version]
- Chung, T.W.; Choi, H.; Lee, J.M.; Ha, S.H.; Kwak, C.H.; Abekura, F.; Park, J.Y.; Chang, Y.C.; Ha, K.T.; Cho, S.H.; et al. Oldenlandia diffusa suppresses metastatic potential through inhibiting matrix metalloproteinase-9 and intercellular adhesion molecule-1 expression via p38 and ERK1/2 MAPK pathways and induces apoptosis in human breast cancer MCF-7 cells. J. Ethnopharmacol. 2017, 195, 309–317. [Google Scholar] [CrossRef]
- Lu, P.H.; Chen, M.B.; Ji, C.; Li, W.T.; Wei, M.X.; Wu, M.H. Aqueous Oldenlandia diffusa extracts inhibits colorectal cancer cells via activating AMP-activated protein kinase signalings. Oncotarget 2016, 7, 45889–45900. [Google Scholar] [CrossRef] [Green Version]
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, K.; Lin, Y.G.; Roman, L.D.; Sood, A.K. Overcoming platinum resistance in ovarian carcinoma. Expert Opin. Investig. Drugs 2010, 19, 1339–1354. [Google Scholar] [CrossRef] [PubMed]
- Miow, Q.H.; Tan, T.Z.; Ye, J.; Lau, J.A.; Yokomizo, T.; Thiery, J.P.; Mori, S. Epithelial-mesenchymal status renders differential responses to cisplatin in ovarian cancer. Oncogene 2015, 34, 1899–1907. [Google Scholar] [CrossRef] [PubMed]
- Demuth, I.; Digweed, M.; Concannon, P. Human SNM1B is required for normal cellular response to both DNA interstrand crosslink-inducing agents and ionizing radiation. Oncogene 2004, 23, 8611–8618. [Google Scholar] [CrossRef] [Green Version]
- Mason, J.M.; Das, I.; Arlt, M.; Patel, N.; Kraftson, S.; Glover, T.W.; Sekiguchi, J.M. The SNM1B/APOLLO DNA nuclease functions in resolution of replication stress and maintenance of common fragile site stability. Hum. Mol. Genet. 2013, 22, 4901–4913. [Google Scholar] [CrossRef] [Green Version]
- Iacobuzio-Donahue, C.A. Epigenetic changes in cancer. Annu. Rev. Pathol. 2009, 4, 229–249. [Google Scholar] [CrossRef]
- Shi, Y.; Lan, F.; Matson, C.; Mulligan, P.; Whetstine, J.R.; Cole, P.A.; Casero, R.A.; Shi, Y. Histone demethylation mediated by the nuclear amine oxidase homolog LSD1. Cell 2004, 119, 941–953. [Google Scholar] [CrossRef] [PubMed]
- Maiques-Diaz, A.; Somervaille, T.C. LSD1: Biologic roles and therapeutic targeting. Epigenomics 2016, 8, 1103–1116. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Jin, Y.; Cui, M.; Zheng, J. Lysine-Specific Demethylase 1 (LSD1) Inhibitor S2101 Induces Autophagy via the AKT/mTOR Pathway in SKOV3 Ovarian Cancer Cells. Med. Sci. Monit. 2016, 22, 4742–4748. [Google Scholar] [CrossRef] [Green Version]
- Katz, T.A.; Vasilatos, S.N.; Harrington, E.; Oesterreich, S.; Davidson, N.E.; Huang, Y. Inhibition of histone demethylase, LSD2 (KDM1B), attenuates DNA methylation and increases sensitivity to DNMT inhibitor-induced apoptosis in breast cancer cells. Breast Cancer Res. Treat. 2014, 146, 99–108. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, C.; Li, W.; Wu, R.; Guo, Y.; Cheng, D.; Yang, Y.; Androulakis, I.P.; Kong, A.N. Pharmacokinetics and Pharmacodynamics of the Triterpenoid Ursolic Acid in Regulating the Antioxidant, Anti-inflammatory, and Epigenetic Gene Responses in Rat Leukocytes. Mol. Pharm. 2017, 14, 3709–3717. [Google Scholar] [CrossRef]
- Available online: http://dspace.ewha.ac.kr/handle/2015.oak/212239 (accessed on 1 June 2016).
- Choi, B.Y.; Joo, J.C.; Lee, Y.K.; Jang, I.S.; Park, S.J.; Park, Y.J. Anti-cancer effect of Scutellaria baicalensis in combination with cisplatin in human ovarian cancer cell. BMC Complement. Altern. Med. 2017, 17, 277. [Google Scholar] [CrossRef]
- Li, Y.; Li, W.; Deng, W.; Gan, Y.; Wu, K.; Sun, J. Synergistic anti-proliferative and pro-apoptotic activities of 5F and cisplatin in human non-small cell lung cancer NCI-H23 cells. Oncol. Lett. 2017, 14, 5347–5353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dronkert, M.L.; Kanaar, R. Repair of DNA interstrand cross-links. Mutat. Res. 2001, 486, 217–247. [Google Scholar] [CrossRef]
- Johnson, S.W.; Swiggard, P.A.; Handel, L.M.; Brennan, J.M.; Godwin, A.K.; Ozols, R.F.; Hamilton, T.C. Relationship between platinum-DNA adduct formation and removal and cisplatin cytotoxicity in cisplatin-sensitive and -resistant human ovarian cancer cells. Cancer Res. 1994, 54, 5911–5916. [Google Scholar] [PubMed]
- Zhen, W.; Link, C.J., Jr.; O’Connor, P.M.; Reed, E.; Parker, R.; Howell, S.B.; Bohr, V.A. Increased gene-specific repair of cisplatin interstrand cross-links in cisplatin-resistant human ovarian cancer cell lines. Mol. Cell Biol. 1992, 12, 3689–3698. [Google Scholar] [CrossRef]
- Luijsterburg, M.S.; van Attikum, H. Chromatin and the DNA damage response: The cancer connection. Mol. Oncol. 2011, 5, 349–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaufort, C.M.; Helmijr, J.C.A.; Piskorz, A.M.; Hoogstraat, M.; Ruigrok-Ritstier, K.; Besselink, N.; Murtaza, M.; van Ijcken, W.F.J.; Heine, A.A.J.; Smid, M.; et al. Ovarian Cancer Cell Line Panel (OCCP): Clinical Importance of In Vitro Morphological Subtypes. PLoS ONE 2014, 9, e103988. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract | Concentration of Treatment | Ursolic Acid | Oleanolic Acid | Units |
|---|---|---|---|---|
| Water extract | 0.187 | 0.054 | mg/g | |
| 0 | 0 | 0 | ||
| 20 | 0.00374 | 0.00108 | ||
| Water extract | 40 | 0.00748 | 0.00216 | µg/mL |
| 80 | 0.01496 | 0.00432 | ||
| 160 | 0.02992 | 0.00864 | ||
| Methanol extract | 15.905 | 5.365 | mg/g | |
| 0 | 0 | 0 | ||
| 20 | 0.3181 | 0.1073 | ||
| Methanol extract | 40 | 0.6462 | 0.2146 | µg/mL |
| 80 | 1.2724 | 0.4292 | ||
| 160 | 2.5448 | 0.8584 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.K.; Lim, J.; Yoon, S.Y.; Joo, J.C.; Park, S.J.; Park, Y.J. Promotion of Cell Death in Cisplatin-Resistant Ovarian Cancer Cells through KDM1B-DCLRE1B Modulation. Int. J. Mol. Sci. 2019, 20, 2443. https://doi.org/10.3390/ijms20102443
Lee YK, Lim J, Yoon SY, Joo JC, Park SJ, Park YJ. Promotion of Cell Death in Cisplatin-Resistant Ovarian Cancer Cells through KDM1B-DCLRE1B Modulation. International Journal of Molecular Sciences. 2019; 20(10):2443. https://doi.org/10.3390/ijms20102443
Chicago/Turabian StyleLee, Yeon Kyu, Jinyeong Lim, So Young Yoon, Jong Cheon Joo, Soo Jung Park, and Yoon Jung Park. 2019. "Promotion of Cell Death in Cisplatin-Resistant Ovarian Cancer Cells through KDM1B-DCLRE1B Modulation" International Journal of Molecular Sciences 20, no. 10: 2443. https://doi.org/10.3390/ijms20102443
APA StyleLee, Y. K., Lim, J., Yoon, S. Y., Joo, J. C., Park, S. J., & Park, Y. J. (2019). Promotion of Cell Death in Cisplatin-Resistant Ovarian Cancer Cells through KDM1B-DCLRE1B Modulation. International Journal of Molecular Sciences, 20(10), 2443. https://doi.org/10.3390/ijms20102443