ATP-Binding Cassette Transporter Regulates N,N′-diacetylchitobiose Transportation and Chitinase Production in Trichoderma asperellum T4


Abstract
1. Introduction
2. Results and Discussion
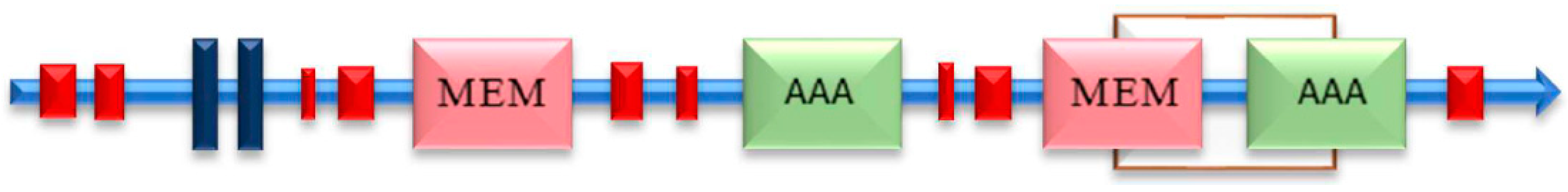
2.1. Characterization of the ABC-B Transport Genes in T. asperellum T4
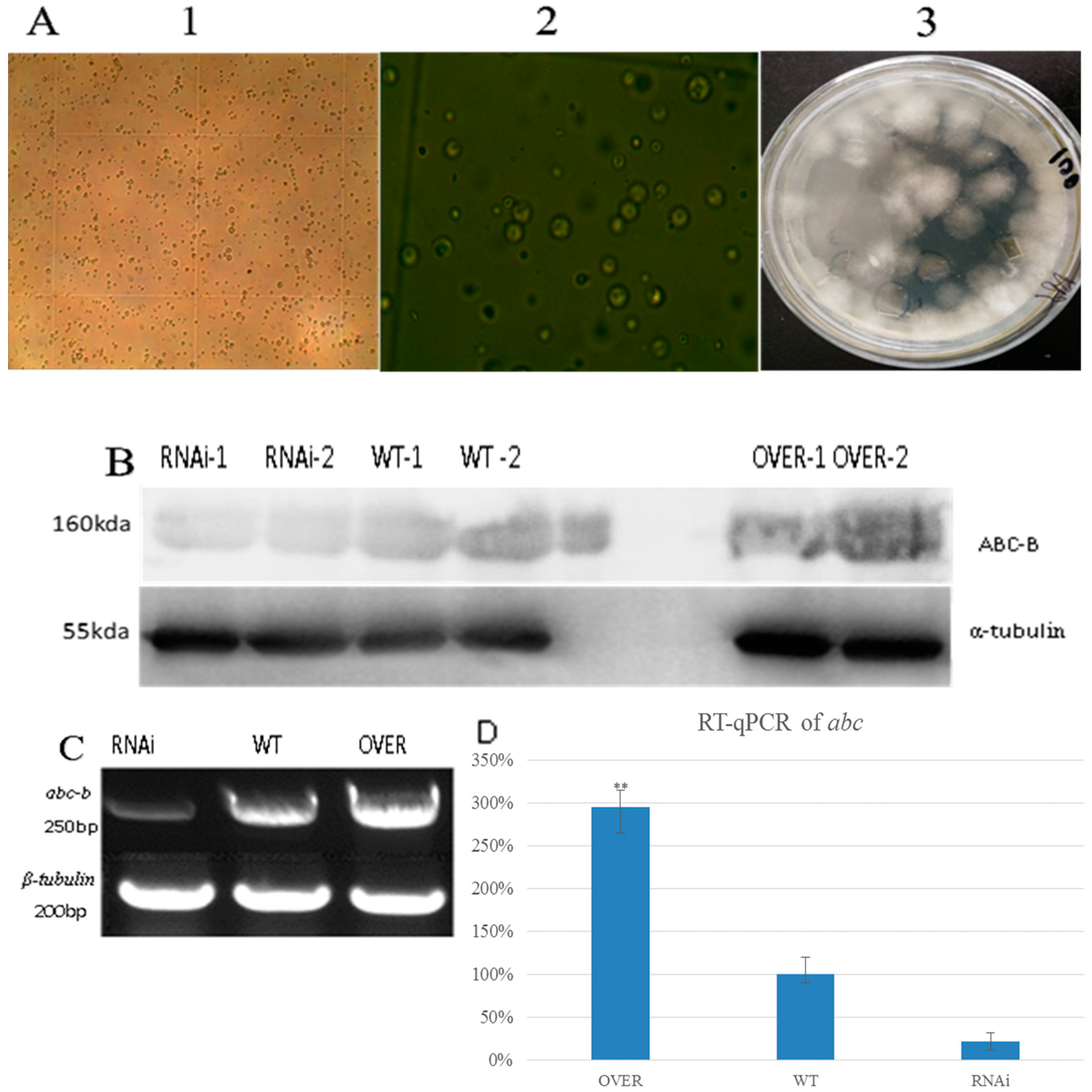
2.2. Growth of Protoplasts and Expression Level of the ABC-B Transporter
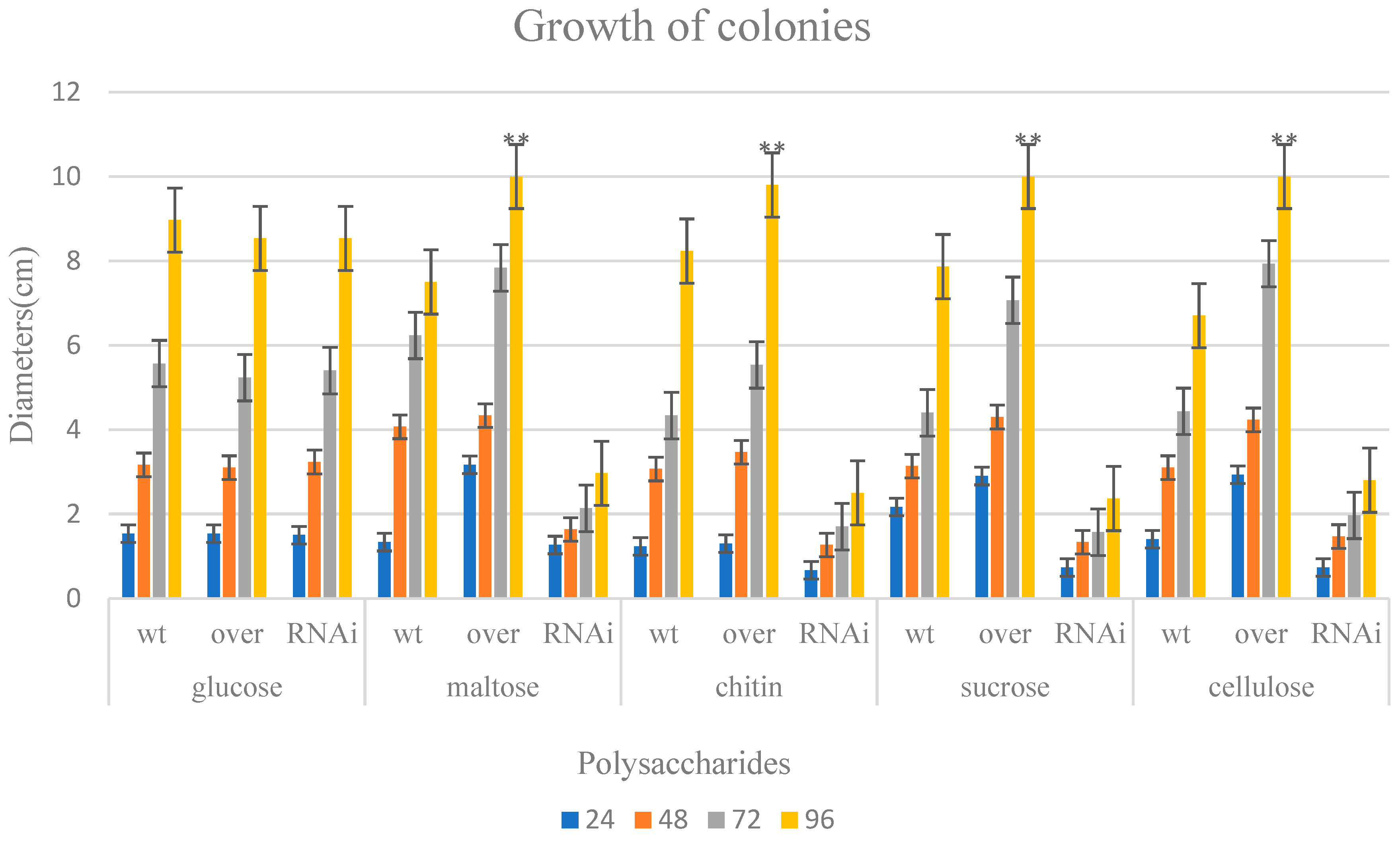
2.3. Growth Conditions Causing a Lack of ABC on Different Polysaccharides
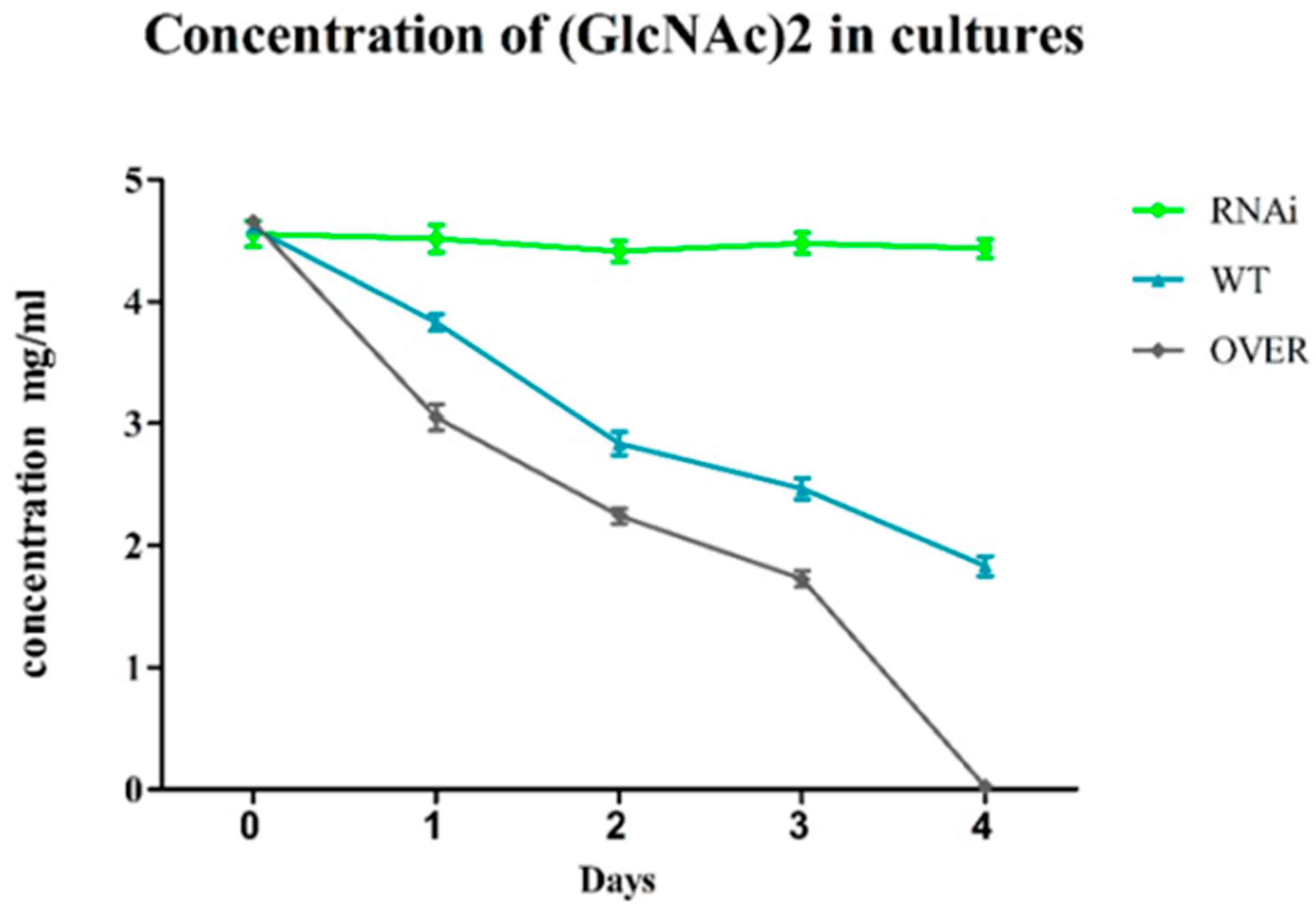
2.4. ABC-B Is Essential for (GlcNAc)2 Transportation
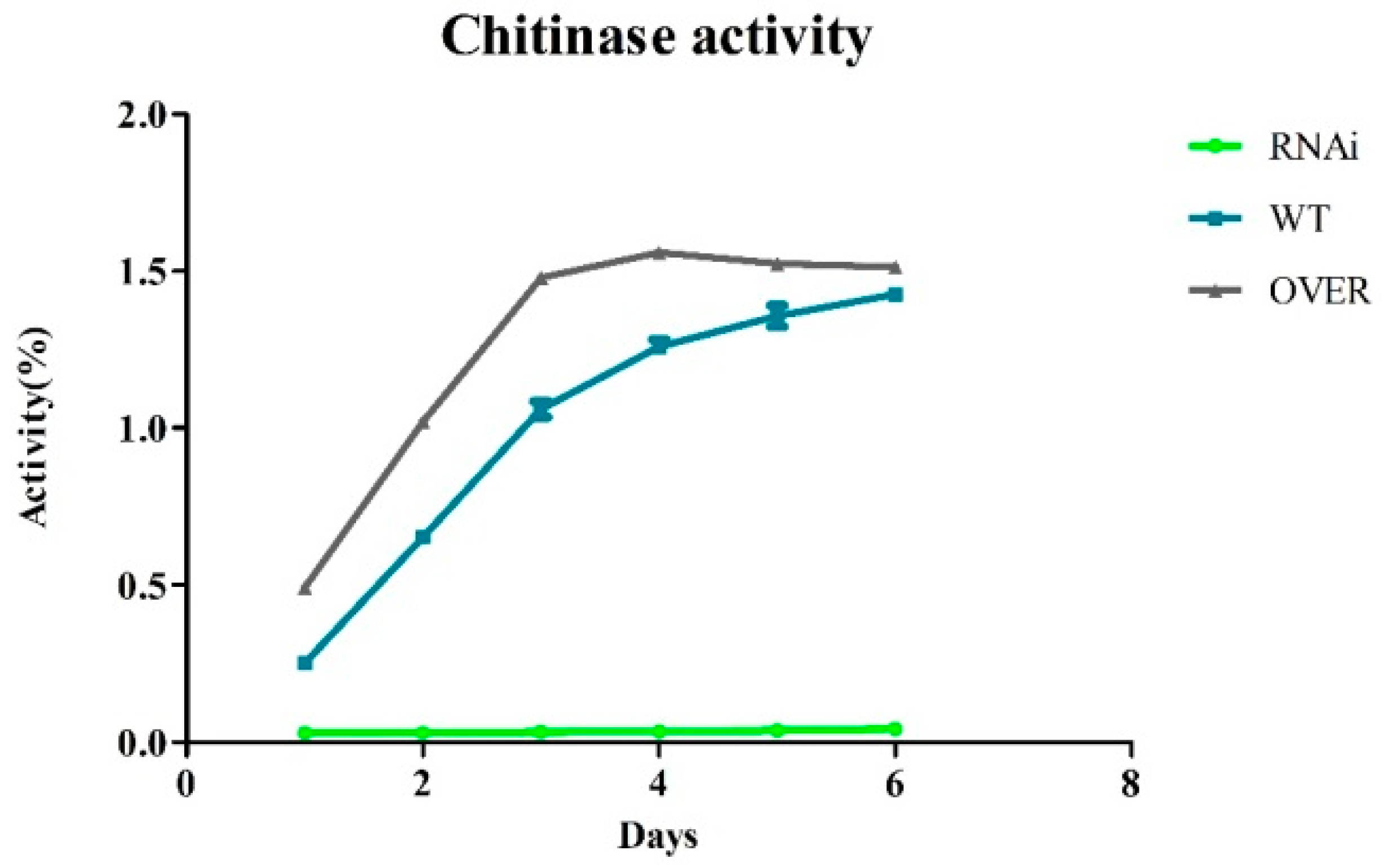
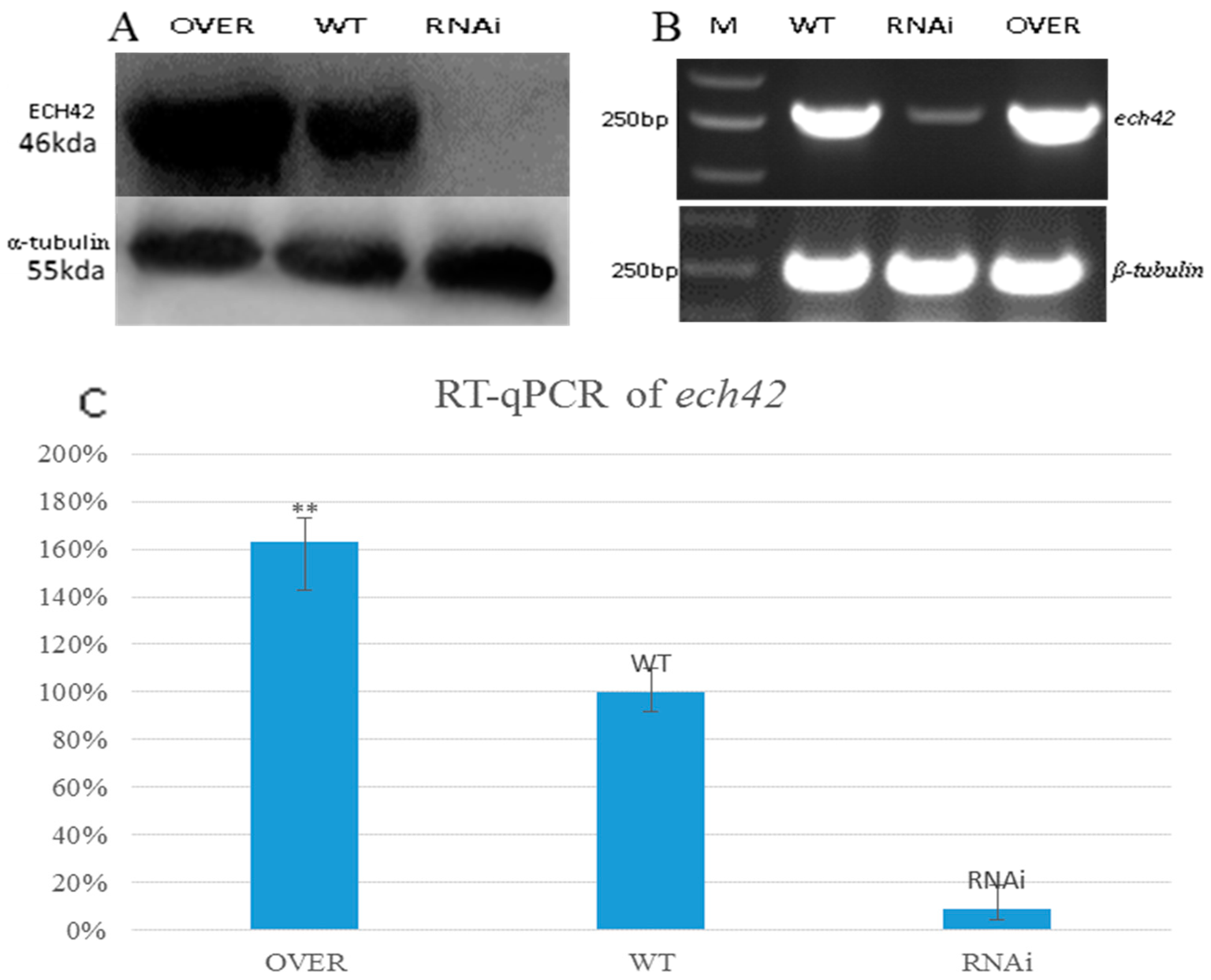
2.5. The ABC-B Transporter Is Essential for Chitinase Induction
3. Materials and Methods
3.1. Bacteria, Fungus, Plasmids, and Culture Conditions
3.2. Extraction of RNA, PCR Amplification, and RT-qPCR
3.3. Plasmid Construction
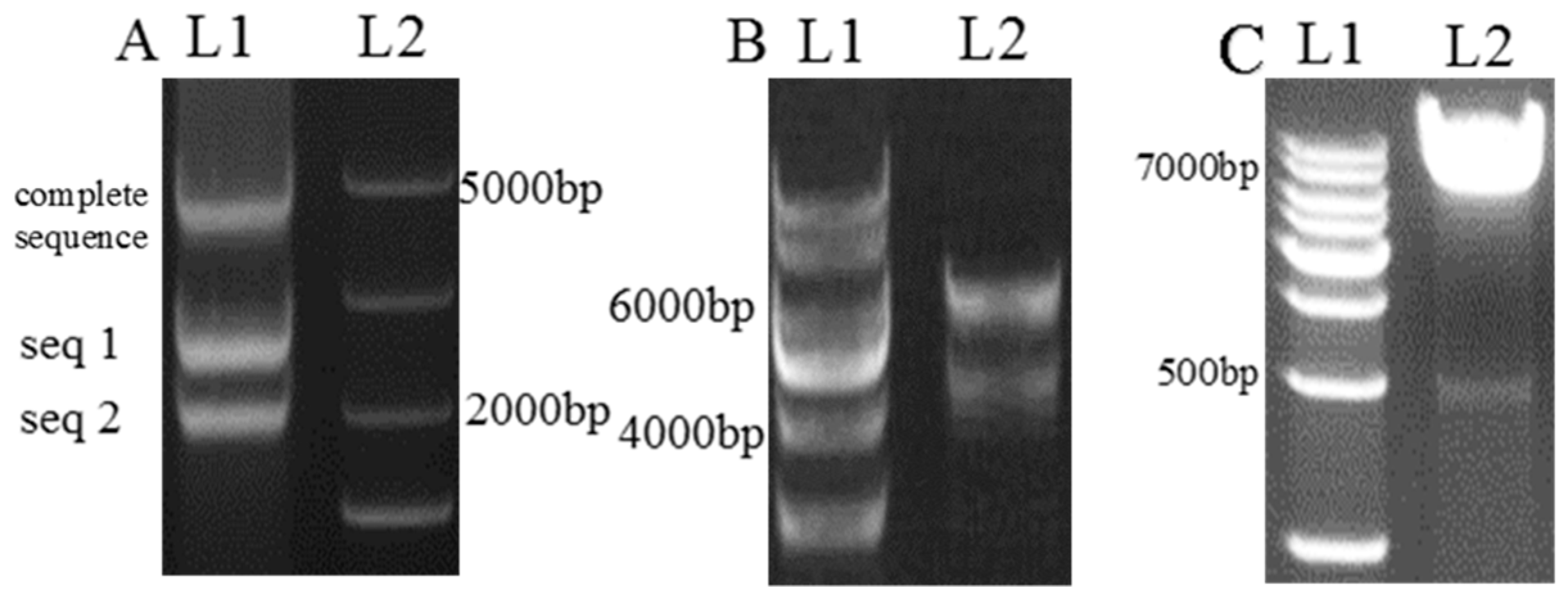
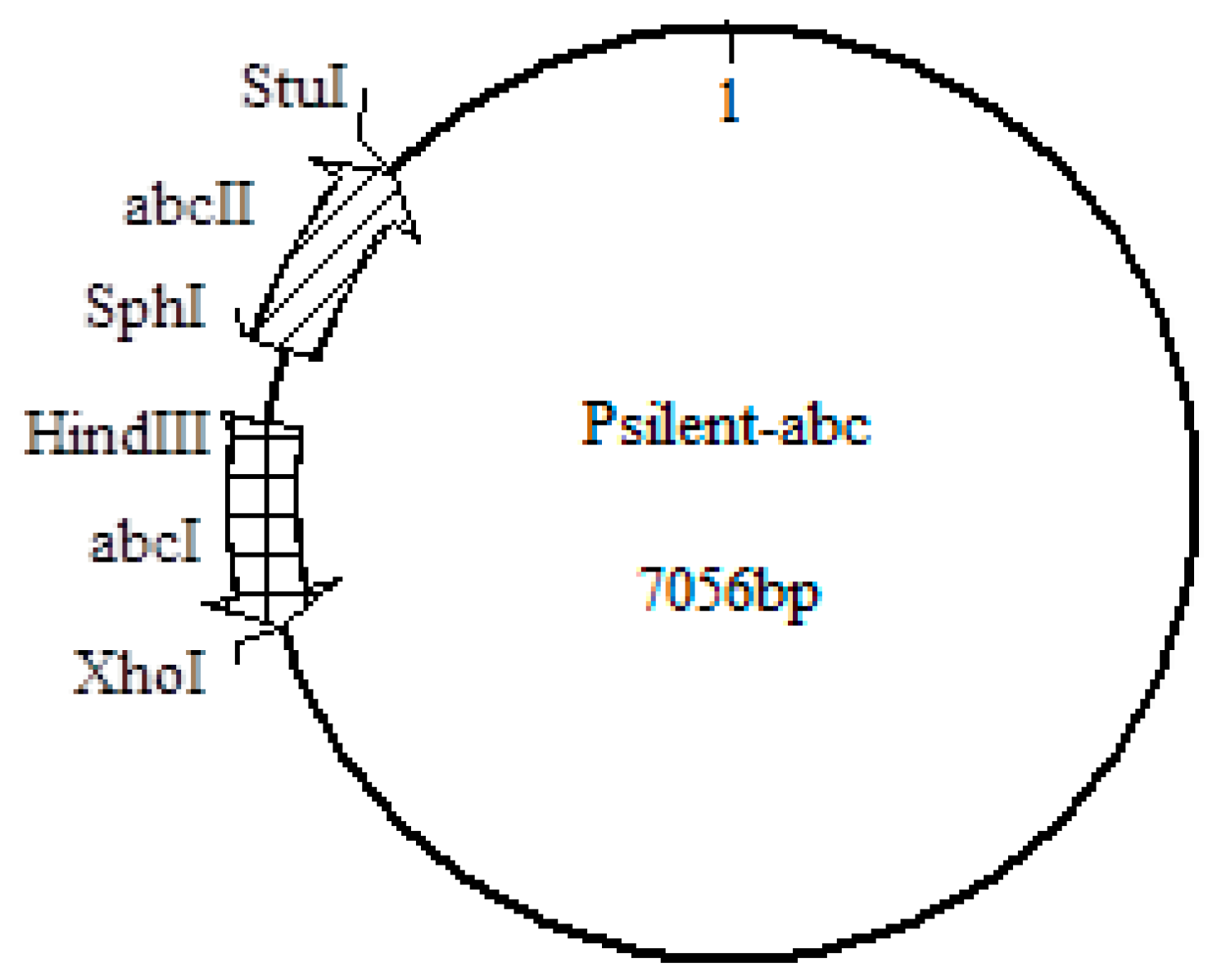
3.3.1. Construction of the Interference Plasmid
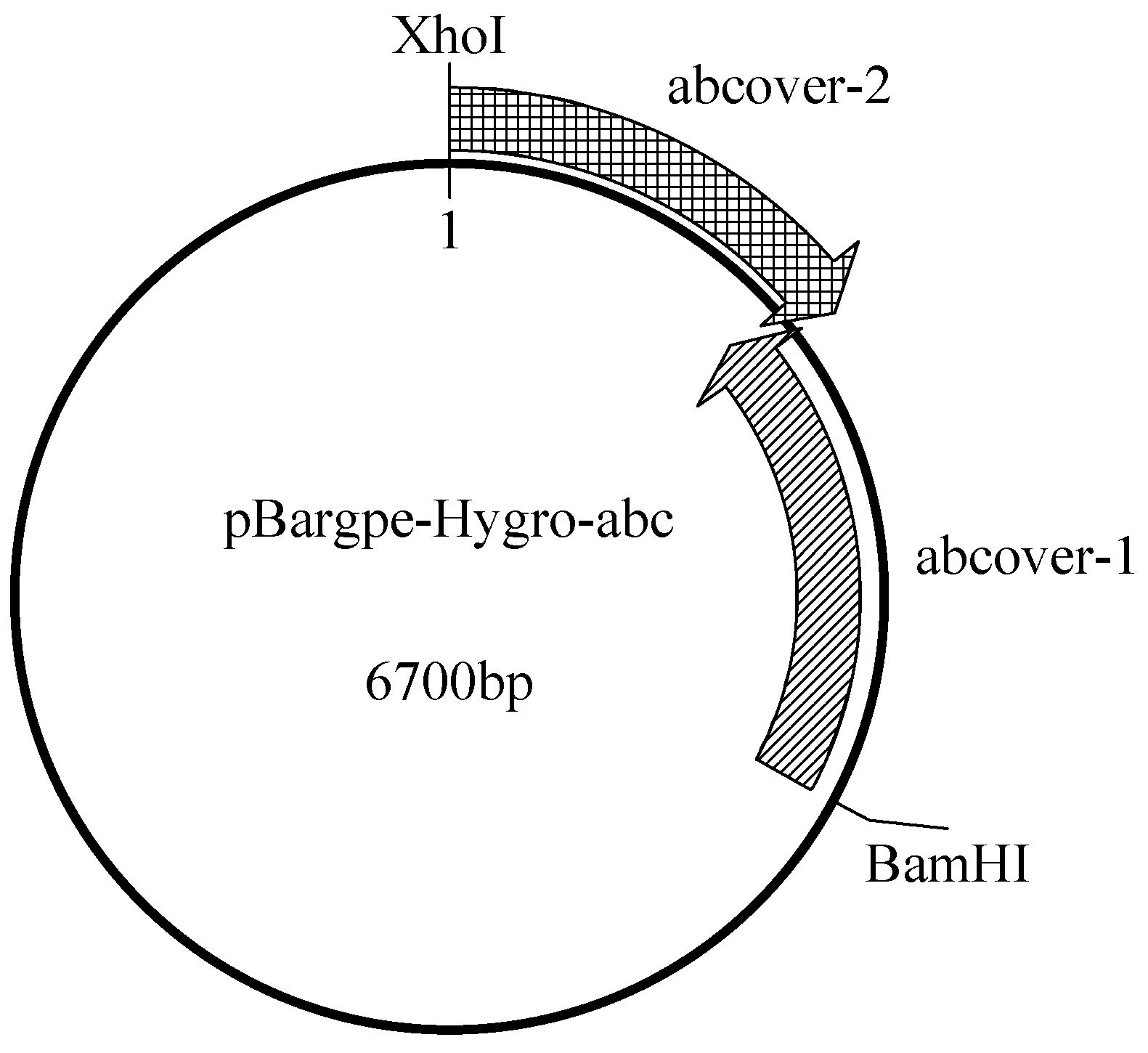
3.3.2. Construction of the Overexpression Plasmid
3.4. Construction of Protoplasts and Transformants
3.5. Detection of the (GlcNAc)2 Concentration
3.6. Western Blot
3.7. Extraction of Fungi Wall Protein
3.8. Chitinase Activity
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Ruocco, M.; Lanzuise, S.; Vinale, F.; Marra, R.; Turrà, D.; Woo, S.L.; Lorito, M. Identification of a New Biocontrol Gene in Trichoderma atroviride: The Role of an ABC Transporter Membrane Pump in the Interaction with Different Plant-Pathogenic Fungi. Mol. Plant-Microbe Interact. 2009, 22, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Wu, S.; Tomasiak, T.M.; Mergel, C.; Winter, M.B.; Stiller, S.B.; Robles-Colmanares, Y.; Stroud, R.M.; Tampe, R.; Craik, C.S.; et al. Subnanometre-resolution electron cryomicroscopy structure of a heterodimeric ABC exporter. Nature 2015, 517, 396–400. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, M. The CydDC ABC transporter of Escherichia coli: New roles for a reductant efflux pump. Biochem. Soc. Trans. 2015, 43, 908–912. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ponte-Sucre, A. Availability and applications of ATP-binding cassette (ABC) transporter blockers. Appl. Microbiol. Biotechnol. 2007, 76, 279–286. [Google Scholar] [CrossRef]
- Dean, M.; Hamon, Y.; Chimini, G. The human ATP-binding cassette (ABC) transporter superfamily. J. Lipid Res. 2001, 42, 1007–1017. [Google Scholar] [CrossRef] [PubMed]
- Dean, M.; Annilo, T. Evolution of the ATP-binding cassette (ABC) transporter superfamily in vertebrates. Ann. Rev. Genom. Hum. Gen. 2005, 6, 123–142. [Google Scholar] [CrossRef] [PubMed]
- Ter Beek, J.; Guskov, A.; Slotboom, D.J. Structural diversity of ABC transporters. J. Cell Biol. 2014, 143, 419–435. [Google Scholar] [CrossRef]
- Wang, T.; Fu, G.; Pan, X.; Wu, J.; Gong, X.; Wang, J.; Shi, Y. Structure of a bacterial energy-coupling factor transporter. Nature 2013, 497, 272–276. [Google Scholar] [CrossRef] [PubMed]
- Rees, D.C.; Johnson, E.; Lewinson, O. ABC transporters: The power to change. Nat. Rev. Mol. Cell Biol. 2009, 10, 218–227. [Google Scholar] [CrossRef]
- Rice, A.J.; Park, A.; Pinkett, H.W. Diversity in ABC transporters: Type I, II and III importers. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 426–437. [Google Scholar] [CrossRef] [PubMed]
- Locher, K.P. Structure and mechanism of ATP-binding cassette transporters. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 239–245. [Google Scholar] [CrossRef]
- Kang, J.; Park, J.; Choi, H.; Burla, B.; Kretzschmar, T.; Lee, Y.; Martinoia, E. Plant ABC transporters. Arab. Book/Am. Soc. Plant Biol. 2011, 9. [Google Scholar] [CrossRef] [PubMed]
- Borghi, L.; Kang, J.; Ko, D.; Lee, Y.; Martinoia, E. The role of ABCG-type ABC transporters in phytohormone transport. Biochem. Soc. Trans. 2015, 43, 924–930. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Choi, Y.; Burla, B.; Kim, Y.Y.; Jeon, B.; Maeshima, M.; Yoo, J.Y.; Martinoia, E.; Lee, Y. The ABC transporter AtABCB14 is a malate importer and modulates stomatal response to CO2. Nat. Cell Biol. 2008, 10, 1217–1223. [Google Scholar] [CrossRef] [PubMed]
- De Marcos Lousa, C.; Van Roermund, C.W.; Postis, V.L.; Dietrich, D.; Kerr, I.D.; Wanders, R.J.; Baldwin, S.A.; Baker, A.; Theodoulou, F.L. Intrinsic acyl-CoA thioesterase activity of a peroxisomal ABC transporter is required for transport and metabolism of fatty acids. Proc. Natl. Acad. Sci. USA 2013, 110, 1279–1284. [Google Scholar] [CrossRef]
- Howell, C.R. Mechanisms employed by Trichoderma species in the biological control of plant diseases: The history and evolution of current concepts. Plant Dis. 2003, 87, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Zhao, Y.; Ran, X.; Guo, L.; Zhao, D.G. Overexpression of a New Chitinase Gene EuCHIT2 Enhances Resistance to Erysiphe cichoracearum DC in Tobacco Plants. Int. J. Mol. Sci. 2017, 18, 2361. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Wang, X.; Liu, S.; Zhang, K.; Cai, Z.; Chen, X.; Zhang, Y.; Liu, J.; Wang, A. The Endochitinase of Clonostachysrosea Expression in Bacillus amyloliquefaciens Enhances the Botrytis cinerea Resistance of Tomato. Int. J. Mol. Sci. 2018, 19, 2221. [Google Scholar] [CrossRef] [PubMed]
- El-Katatny, M.H.; Gudelj, M.; Robra, K.H.; Elnaghy, M.A.; Gübitz, G.M. Characterization of a chitinase and an endo-ß-1,3-glucanase from Trichoderma harzianum Rifai T24 involved in control of the phytopathogen Sclerotium rolfsii. Appl. Microbiol. Biotechnol. 2001, 56, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Langner, T.; Göhre, V. Fungal chitinases: Function, regulation, and potential roles in plant/pathogen interactions. Curr. Genet. 2016, 62, 243–254. [Google Scholar] [CrossRef]
- Jung, W.J.; Park, R.D. Bioproduction of chitooligosaccharides: Present and perspectives. Mar. Drugs 2014, 12, 5328–5356. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Fu, X.; Yang, Q.; Guo, Y.; Liu, Z.; Jiang, Z. Cloning, expression, purification and application of a novel chitinase from a thermophilic marine bacterium Paenibacillus barengoltzii. Food Chem. 2016, 192, 1041–1048. [Google Scholar] [CrossRef] [PubMed]
- Kzhyshkowska, J.; Yin, S.; Liu, T.; Riabov, V.; Mitrofanova, I. Role of chitinase-like proteins in cancer. Biol. Chem. 2016, 397, 231–247. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, A.F.; García-Fraga, B.; López-Seijas, J.; Sieiro, C. Optimizing the expression of a Heterologous chitinase: A study of different promoters. Bioengineered 2017, 8, 428–432. [Google Scholar] [CrossRef] [PubMed]
- Saito, A.; Fujii, T.; Shinya, T.; Shibuya, N.; Ando, A.; Miyashita, K. The msiK gene, encoding the ATP-hydrolysing component of N,N′-diacetylchitobiose ABC transporters, is essential for induction of chitinase production in Streptomyces coelicolor A3(2). Microbiology 2008, 154 Pt 11, 3358–3365. [Google Scholar] [CrossRef]
- Niu, X.; Liu, C.C.; Xiong, Y.J.; Yang, M.M.; Ma, F.; Liu, Z.H.; Yuan, S. The Modes of Action of ChiIII, a Chitinase from Mushroom Coprinopsis cinerea, Shift with Changes in the Length of GlcNAc Oligomers. J. Agric. Food Chem. 2016, 64, 6958–6968. [Google Scholar] [CrossRef]
- Brunner, K.; Montero, M.; Mach, R.L.; Peterbauer, C.K.; Kubicek, C.P. Expression of the ech42 (endochitinase) gene of Trichoderma atroviride under carbon starvation is antagonized via a BrlA-like cis-acting element. FEMS Microbiol. Lett. 2003, 218, 259–264. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Theodoulou, F.L.; Kerr, I.D. ABC transporter research: Going strong 40 years on. Biochem. Soc. Trans. 2015, 43, 1033–1040. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.E.; Saraste, M.; Runswick, M.J.; Gay, N.J. Distantly related sequences in the α- and β-subunits of ATP synthase, myosin, kinases and other ATP-requiring enzymes and a common nucleotide binding fold. EMBO J. 1982, 8, 945–951. [Google Scholar] [CrossRef]
- Yazaki, K. ABC transporters involved in the transport of plant secondary metabolites. FEBS Lett. 2006, 580, 1183–1191. [Google Scholar] [CrossRef]
- Verrier, P.J.; Bird, D.; Burla, B.; Dassa, E.; Forestier, C.; Geisler, M.; Klein, M.; Kolukisaoglu, U.; Lee, Y.; Martinoia, E.; et al. Plant ABC proteins: A unified nomenclature and updated inventory. Trends Plant Sci. 2008, 13, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Scarcelli, J.J.; Colussi, P.A.; Fabre, A.L.; Boles, E.; Orlean, P.; Taron, C.H. Uptake of radiolabeled GlcNAc into Saccharomyces cerevisiae via native hexose transporters and its in vivo incorporation into GPI precursors in cells expressing heterologous GlcNAc kinase. FEMS Yeast Res. 2012, 12, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Robb, M.; Hobbs, J.K.; Woodiga, S.A.; Shapiro-Ward, S.; Suits, M.D.; McGregor, N.; Brumer, H.; Yesilkaya, H.; King, S.J.; Boraston, A.B. Molecular Characterization of N-glycan Degradation and Transport in Streptococcus pneumoniae and Its Contribution to Virulence. PLoS Pathog. 2017, 13, e1006090. [Google Scholar] [CrossRef]
- Le, B.; Yang, S.H. Characterization of a chitinase from Salinivibrio sp. BAO-1801 as an antifungal activity and a biocatalyst for producing chitobiose. J. Basic Microbiol. 2018, 58, 848–856. [Google Scholar] [CrossRef] [PubMed]
- Anthony, P.; Davey, M.R.; Power, J.B.; Washington, C.; Lowe, K.C. Synergistic enhancement of protoplast growth by oxygenated perfluorocarbon and Pluronic F-68. Plant Cell Rep. 1994, 13, 251–255. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Name | Content | Product Size (bp) | Function |
|---|---|---|---|---|
| Overexpression primer | TAABCP1-5 | CGGATCCATGGCAGACCACGGTGAAGAGAAAG | 2451 ABCOVER-1 | ABC upstream sequence |
| TAABCP1-3 | GGCATGCCTTCTGCCCTCAGTATGC | |||
| TAABCP2-5 | CCTCGAGAGCCCTTGTGGTCTCG | 1923 ABCOVER-2 | ABC downstream sequence | |
| TAABCP2-3 | AAGGCCTAGCCCTTGTGGTCTCG | |||
| Interference primer | TAABCI-5 | CAAGCTTCTTCTGCCCTCAGTATGC | 500 ABCI | Crucial section of ABC to interfere |
| TAABCI-3 | CCTCGAGAGCCCTTGTGGTCTCG | |||
| TAABCII-5 | GGCATGCCTTCTGCCCTCAGTATGC | 500 ABCII | ||
| TAABCII-3 | AAGGCCTAGCCCTTGTGGTCTCG | |||
| ABC qPCR primer | TAABCq-5 | CTCAGACCCACTCTCCGAA | 280 | Expression level of ABC |
| TAABCq-3 | GGGTCTGAGAGCGGAAAA | |||
| ECH42 qPCR primer | Ech42q-5 | CAACTGGGGTATCTATGGC | 250 | Expression level of ECH42 |
| Ech42q-3 | TTGCTGCAGAAGGGAAG | |||
| β-tubulin qPCR primer | Tubulin-5 | CACCTTCCTCCATACCCTCACC | 200 | Reference gene |
| Tubulin-3 | TGTCCAACACCACCGCCATC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Cheng, M.; Zhao, S.; Lin, C.; Song, J.; Yang, Q. ATP-Binding Cassette Transporter Regulates N,N′-diacetylchitobiose Transportation and Chitinase Production in Trichoderma asperellum T4. Int. J. Mol. Sci. 2019, 20, 2412. https://doi.org/10.3390/ijms20102412
Liu H, Cheng M, Zhao S, Lin C, Song J, Yang Q. ATP-Binding Cassette Transporter Regulates N,N′-diacetylchitobiose Transportation and Chitinase Production in Trichoderma asperellum T4. International Journal of Molecular Sciences. 2019; 20(10):2412. https://doi.org/10.3390/ijms20102412
Chicago/Turabian StyleLiu, He, Ming Cheng, Shanshan Zhao, Congyu Lin, Jinzhu Song, and Qian Yang. 2019. "ATP-Binding Cassette Transporter Regulates N,N′-diacetylchitobiose Transportation and Chitinase Production in Trichoderma asperellum T4" International Journal of Molecular Sciences 20, no. 10: 2412. https://doi.org/10.3390/ijms20102412
APA StyleLiu, H., Cheng, M., Zhao, S., Lin, C., Song, J., & Yang, Q. (2019). ATP-Binding Cassette Transporter Regulates N,N′-diacetylchitobiose Transportation and Chitinase Production in Trichoderma asperellum T4. International Journal of Molecular Sciences, 20(10), 2412. https://doi.org/10.3390/ijms20102412