Cathepsin L-like Cysteine Proteinase Genes Are Associated with the Development and Pathogenicity of Pine Wood Nematode, Bursaphelenchus xylophilus


Abstract
1. Introduction
2. Results
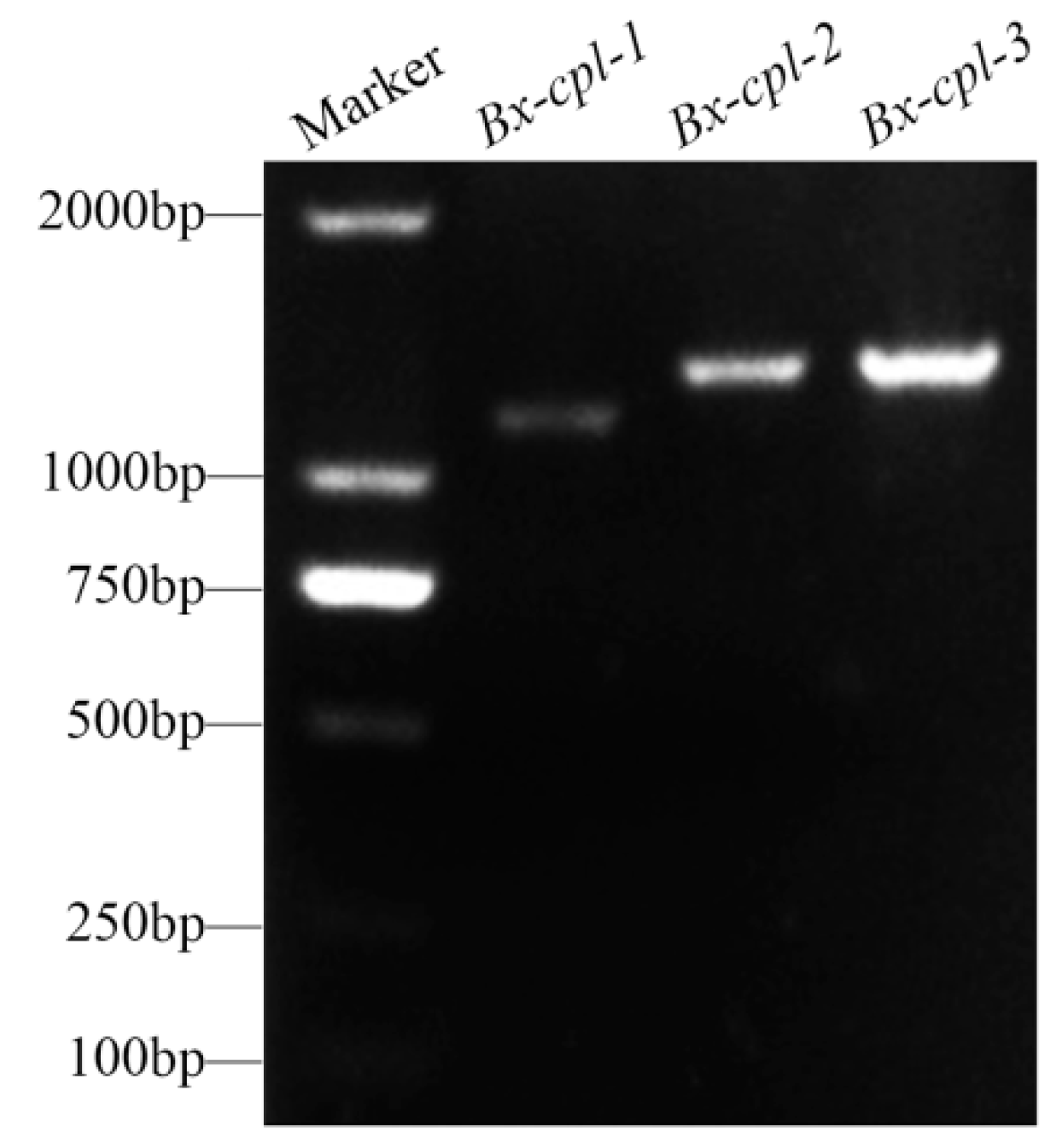
2.1. Cloning and Sequence Analysis of Three Cathepsin L-Like Cysteine Proteinase Genes from B. xylophilus
2.2. Localization of Bx-cpl in B. xylophilus
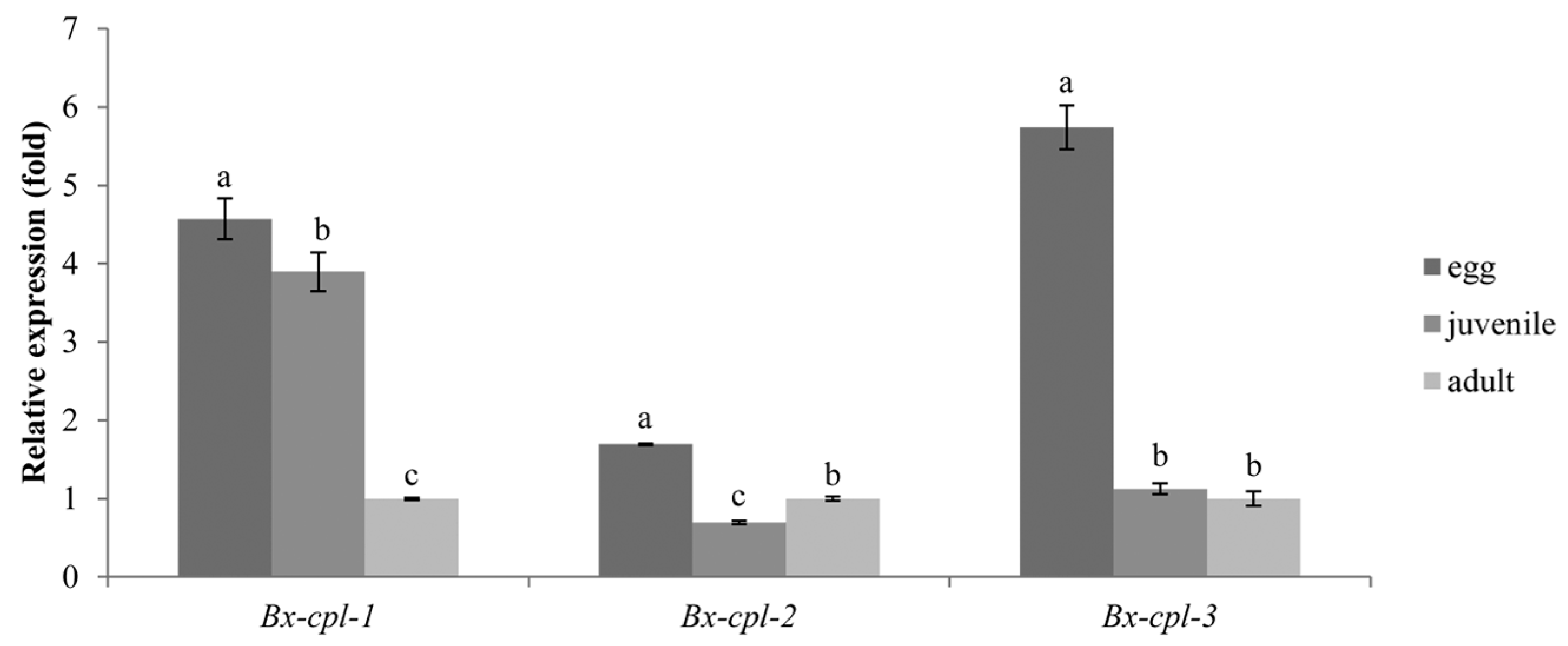
2.3. Expression of Bx-cpl at PWN Developmental Stages
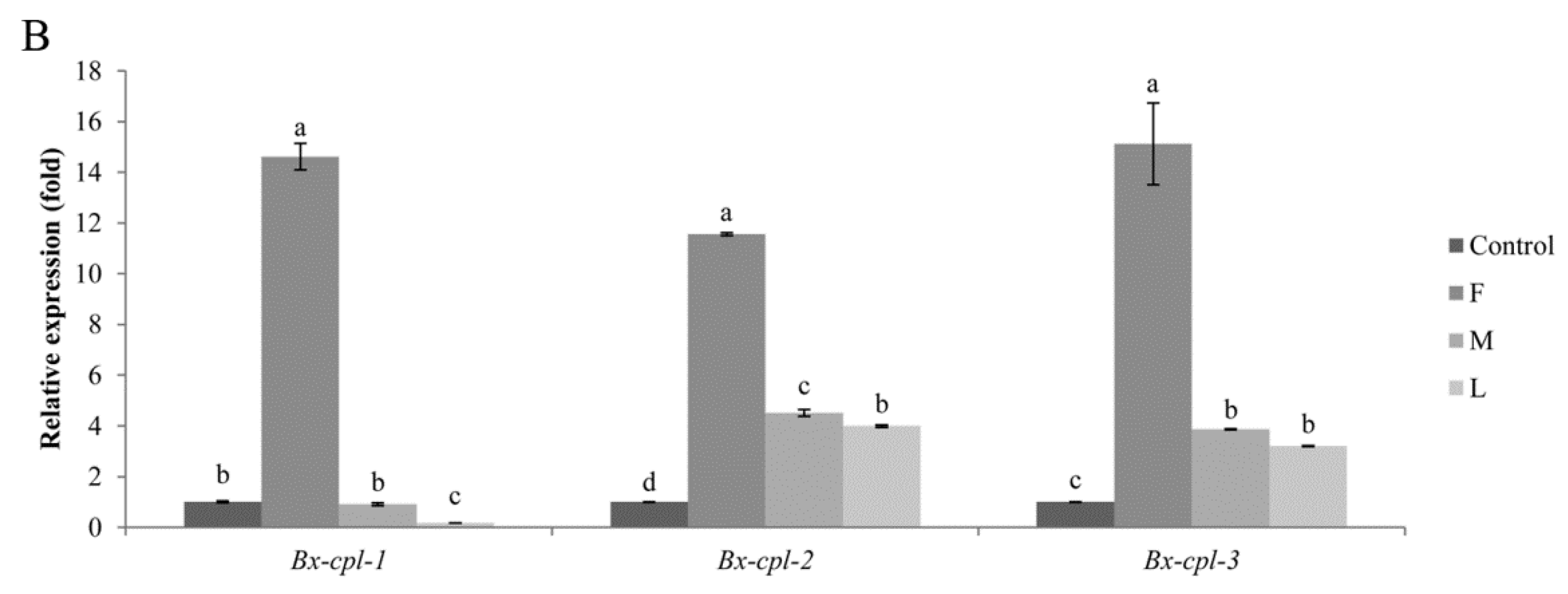
2.4. Expression of Bx-cpl at PWD Development Stages
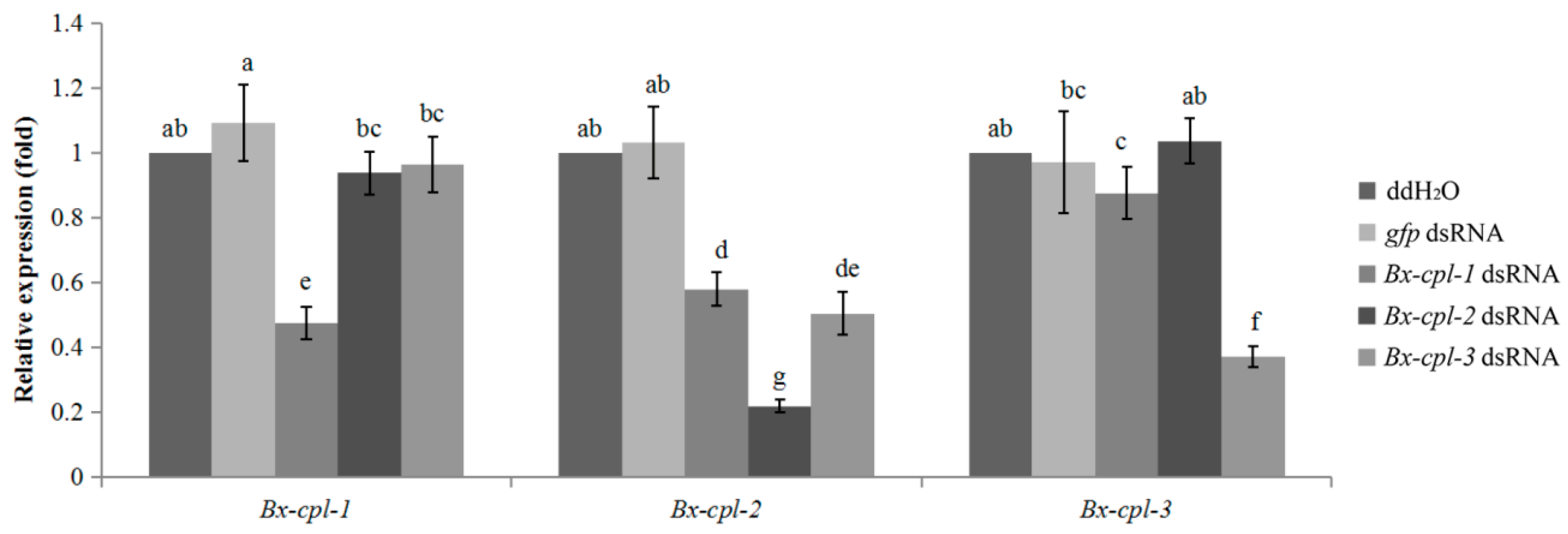
2.5. Detection of RNAi Efficiency
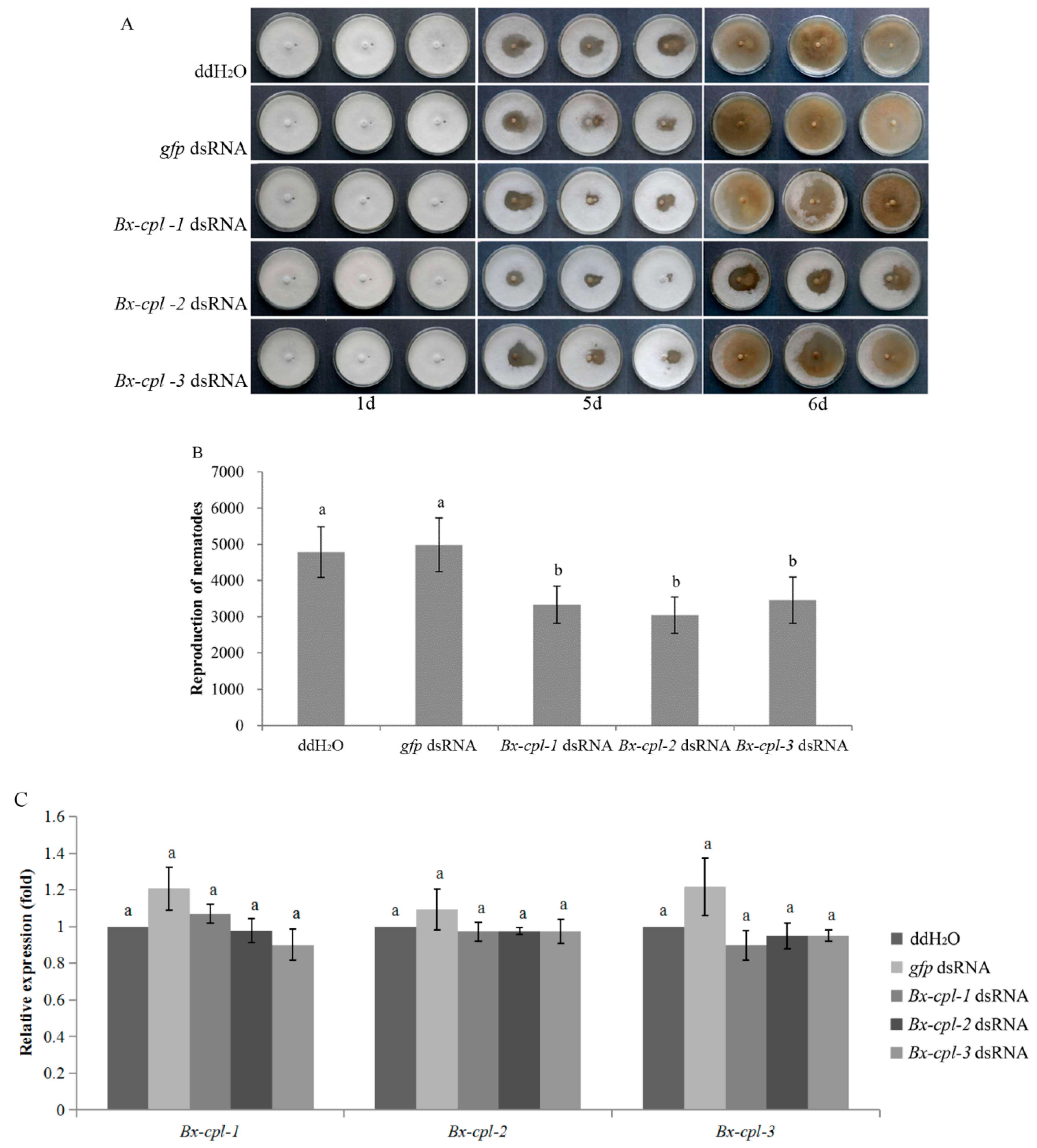
2.6. Feeding and Reproduction of B. xylophilus after RNAi
2.7. Pathogenicity of B. xylophilus after RNAi
3. Discussion
4. Materials and Methods
4.1. Nematode Culture and Collection
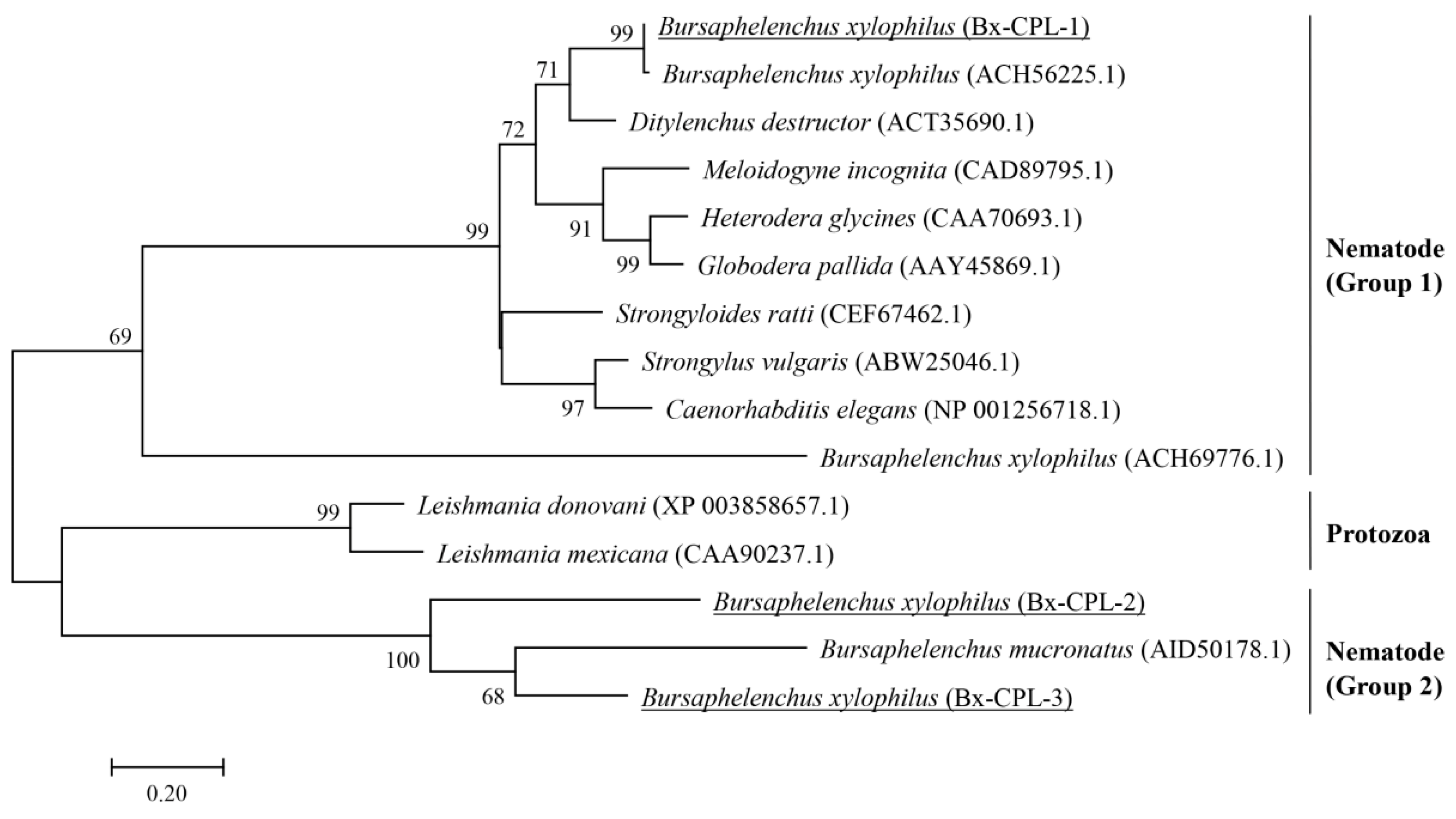
4.2. RNA Extraction, PCR Amplification of Bx-cpls, and Phylogenetic Analysis
4.3. In Situ Hybridization (ISH)
4.4. Synthesis of Bx-cpl dsRNA and Interference
4.5. The qPCR and Expression Analysis of Bx-cpls
4.6. Analysis of Reproduction and Pathogenicity of B. xylophilus after RNAi
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shinya, R.; Morisaka, H.; Takeuchi, Y.; Futai, K.; Ueda, M. Making headway in understanding pine wilt disease: What do we perceive in the postgenomic era? J. Biosci. Bioeng. 2013, 116, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Dwinell, L.D. First report of pinewood nematode (Bursaphelenchus xylophilus) in Mexico. Plant Dis. 1993, 77, 846A. [Google Scholar] [CrossRef]
- Mamiya, Y. History of pine wilt disease in Japan. J. Nematol. 1988, 20, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Liang, J.; Yan, D.H.; Zhang, X.Y. Research advances of pine wood nematode disease in China. World For. Res. 2010, 23, 59–63. [Google Scholar] [CrossRef]
- Yi, C.K.; Byun, B.H.; Park, J.D.; Yang, S.I.; Chang, K.H. First finding of the pine wood nematode, Bursaphelenchus xylophilus (Steiner et Buhrer) Nickle and its insect vector in Korea. Res Rep For Res Inst Seoul. 1989, 3, 141–149. [Google Scholar]
- Mota, M.M.; Braasch, H.; Bravo, M.A.; Penas, A.C.; Burgermeister, W.; Metge, K.; Sousa, E. First report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1999, 1, 727–734. [Google Scholar] [CrossRef]
- Abelleira, A.; Picoaga, A.; Mansilla, J.P.; Aguin, O. Detection of Bursaphelenchus xylophilus, causal agent of pine wilt disease on Pinus pinaster in northwestern Spain. Plant Dis. 2011, 95, 776. [Google Scholar] [CrossRef]
- Khan, F.A.; Gbadegesin, R.A. On the occurrence of nematode induced pine wilt disease in Nigeria. Pak. J. Nematol. 1991, 57, 162–164. [Google Scholar] [CrossRef]
- Kikuchi, T.; Aikawa, T.; Kosaka, H.; Pritchard, L.; Ogura, N.; Jones, J.T. Expressed sequence tag (EST) analysis of the pine wood nematode Bursaphelenchus xylophilus and B. mucronatus. Mol. Biochem. Parasitol. 2007, 155, 9–17. [Google Scholar] [CrossRef]
- Kikuchi, T.; Cotton, J.A.; Dalzell, J.J.; Hasegawa, K.; Kanzaki, N.; McVeigh, P.; Takanashi, T.; Tsai, I.J.; Assefa, S.A.; Cock, P.J.; et al. Genomic insights into the origin of parasitism in the emerging plant pathogen Bursaphelenchus xylophilus. PLoS Pathog. 2011, 7, e1002219. [Google Scholar] [CrossRef]
- Shinya, R.; Morisaka, H.; Kikuchi, T.; Takeuchi, Y.; Ueda, M.; Futai, K. Secretome Analysis of the pine wood nematode Bursaphelenchus xylophilus reveals the tangled roots of parasitism and its potential for molecular mimicry. PLoS ONE 2013, 8, e67377. [Google Scholar] [CrossRef] [PubMed]
- Tsai, I.J.; Tanaka, R.; Kanzaki, N.; Akiba, M.; Yokoi, T.; Espada, M.; Jones, J.T.; Kikuchi, T. Transcriptional and morphological changes in the transition from mycetophagous to phytophagous phase in the plant-parasitic nematode Bursaphelenchus xylophilus. Mol. Plant Pathol. 2016, 17, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, J.; Anjo, S.; Fonseca, L.; Egas, C.; Manadas, B.; Abrantes, I. Bursaphelenchus xylophilus and B. mucronatus secretomes: A comparative proteomic analysis. Sci. Rep. 2016, 6, 39007. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, T.; Jones, J.T.; Aikawa, T.; Kosaka, H.; Ogura, N. A family of glycosyl hydrolase family 45 cellulases from the pine wood nematode Bursaphelenchus xylophilus. FEBS Lett. 2004, 572, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Fan, Y.; Zheng, H.; Du, F.; Zhang, K.Q.; Huang, X.; Wang, L.; Zhang, M.; Niu, Q. Isolation and characterization of a novel endoglucanase from a Bursaphelenchus xylophilus metagenomic library. PLoS ONE 2013, 8, e82437. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, T.; Shibuya, H.; Aikawa, T.; Jones, J.T. Cloning and characterization of pectate lyases expressed in the esophageal gland of the pine wood nematode Bursaphelenchus xylophilus. Mol. Plant Microbe Interact. 2006, 19, 280–287. [Google Scholar] [CrossRef]
- Lee, D.W.; Kang, J.S.; Jung, C.S.; Han, H.R.; Moon, Y.S.; Park, S.J.; Lee, S.H.; Koh, Y.H. Identification and biochemical analysis of a novel pectate lyase 3 gene in Bursaphelenchus xylophilus. J. Asia-Pac. Entomol. 2013, 16, 335–342. [Google Scholar] [CrossRef]
- Kikuchi, T.; Li, H.M.; Karim, N.; Kennedy, M.W.; Moens, M.; Jones, J.T. Identification of putative expansin-like genes from the pine wood nematode, Bursaphelenchus xylophilus, and evolution of the expansin gene family within the nematoda. Nematology 2009, 11, 355–364. [Google Scholar] [CrossRef]
- Kim, Y.H.; Kim, A.Y.; Choi, B.H.; Han, H.R.; Koh, Y.H. ExpansinB3 as a marker for detecting pine wood nematode-infected pine trees. J. Asia-Pac. Entomol. 2017, 20, 1228–1233. [Google Scholar] [CrossRef]
- Lin, S.F.; Jian, H.; Zhao, H.J.; Yang, D.; Liu, Q. Cloning and characterization of a venom allergen-like protein gene cluster from the pinewood nematode Bursaphelenchus xylophilus. Exp. Parasitol. 2011, 127, 440–447. [Google Scholar] [CrossRef]
- Xu, X.L.; Wu, X.Q.; Ye, J.R.; Huang, L. Molecular characterization and functional analysis of three pathogenesis-related cytochrome P450 genes from Bursaphelenchus xylophilus (Tylenchida, Aphelenchoidoidea). Int. J. Mol. Sci. 2015, 16, 5216–5234. [Google Scholar] [CrossRef]
- Rhoads, M.L.; Fetterer, R.H. Extracellular matrix: A tool for defining the extracorporeal function of parasite proteases. Parasitol. Today 1997, 13, 119–122. [Google Scholar] [CrossRef]
- Malagón, D.; Benítez, R.; Kašný, M.; Adroher, F.J. Peptidases in parasitic nematodes. A review. In Parasites: Ecology, Diseases and Management; Erzinger, G.S., Ed.; Nova Science Publishers Inc.: New York, NY, USA, 2013; pp. 61–102. ISBN 978-1-62257-692-0. [Google Scholar]
- Sajid, M.; McKerrow, J.H. Cysteine proteases of parasitic organisms. Mol. Biochem. Parasitol. 2002, 120, 1–21. [Google Scholar] [CrossRef]
- Britton, C.; Murray, L. A cathepsin L protease essential for Caenorhabditis elegans embryogenesis is functionally conserved in parasitic nematodes. Mol. Biochem. Parasitol. 2002, 122, 21–33. [Google Scholar] [CrossRef]
- Urwin, P.E.; Lilley, C.J.; McPherson, M.J.; Atkinson, H.J. Characterization of two cDNAs encoding cysteine proteinases from the soybean cyst nematode Heterodera glycines. Parasitology 1997, 114, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Neveu, C.; Abad, P.; Castagnone-Sereno, P. Molecular cloning and characterization of an intestinal cathepsin L protease from the plant-parasitic nematode Meloidogyne incognita. Physiol. Mol. Plant Pathol. 2003, 63, 159–165. [Google Scholar] [CrossRef]
- Shingles, J.; Lilley, C.J.; Atkinson, H.J.; Urwin, P.E. Meloidogyne incognita: Molecular and biochemical characterisation of a cathepsin L cysteine proteinase and the effect on parasitism following RNAi. Exp. Parasitol. 2007, 115, 114–120. [Google Scholar] [CrossRef]
- Neveu, C.; Jaubert, S.; Abad, P.; Castagnone-Sereno, P. A set of genes differentially expressed between avirulent and virulent Meloidogyne incognita near-isogenic lines encode secreted proteins. Mol. Plant Microbe Interact. 2003, 16, 1077. [Google Scholar] [CrossRef]
- Dutta, T.K.; Papolu, P.K.; Banakar, P.; Choudhary, D.; Sirohi, A.; Rao, U. Tomato transgenic plants expressing hairpin construct of a nematode protease gene conferred enhanced resistance to root-knot nematodes. Front. Microbiol. 2015, 6, 260. [Google Scholar] [CrossRef]
- Duarte, A.; Maleita, C.; Tiago, I.; Curtis, R.; Abrantes, I. Molecular characterization of putative parasitism genes in the plant-parasitic nematode Meloidogyne hispanica. J. Helminthol. 2016, 90, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Urwin, P.E.; Lilley, C.J.; Atkinson, H.J. Ingestion of double-stranded RNA by preparasitic juvenile cyst nematodes leads to RNA interference. Mol. Plant Microbe Interact. 2002, 15, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Dalton, J.P.; Neill, S.O.; Stack, C.; Collins, P.; Walshe, A.; Sekiya, M.; Doyle, S.; Mulcahy, G.; Hoyle, D.; Khaznadji, E.; et al. Fasciola hepatica cathepsin L-like proteases: Biology, function, and potential in the development of first generation liver fluke vaccines. Int. J. Parasitol. 2003, 33, 1173–1181. [Google Scholar] [CrossRef]
- Corvo, I.; Cancela, M.; Cappetta, M.; Pi-Denis, N.; Tort, J.F.; Roche, L. The major cathepsin L secreted by the invasive juvenile Fasciola hepatica prefers proline in the S2 subsite and can cleave collagen. Mol. Biochem. Parasitol. 2009, 167, 41. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.F.; Peng, D.L.; Sun, J.H.; Huang, W.K.; Peng, H.; Long, H.B. Cloning and sequence analysis of a new cathepsin L-like cysteine proteinase gene from Ditylenchus destructor. Chin. J. Biotechnol. 2011, 27, 60–68. [Google Scholar] [CrossRef]
- Pan, Y.Y.; Huang, L.; Wu, X.Q. Bioinformatic and expression analysis of a cathepsin gene Bmcath1 in Bursaphelechus mucronatus. J. Nanjing For. Univ. 2015, 39, 12–16. [Google Scholar] [CrossRef]
- Hashmi, S.; Britton, C.; Liu, J.; Guiliano, D.B.; Oksov, Y.; Lustigman, S. Cathepsin L is essential for embryogenesis and development of Caenorhabditis elegans. J. Biol. Chem. 2002, 277, 3477. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.N.; Wu, X.Q.; Ye, J.R.; Xue, Q. Identification of autophagy in the pine wood nematode Bursaphelenchus xylophilus and the molecular characterization and functional analysis of two novel autophagy-related genes, BxATG1 and BxATG8. Int. J. Mol. Sci. 2016, 17, 279. [Google Scholar] [CrossRef] [PubMed]
- Rhoads, M.L.; Fetterer, R.H. Developmentally regulated secretion of cathepsin L-like cysteine proteases by Haemonchus contortus. J. Parasitol. 1995, 81, 505–512. [Google Scholar] [CrossRef]
- Qiu, X.W.; Wu, X.Q.; Huang, L.; Tian, M.Q.; Ye, J.R. Specifically expressed genes of the nematode Bursaphelenchus xylophilus involved with early interactions with pine trees. PLoS ONE 2013, 8, e78063. [Google Scholar] [CrossRef]
- Kang, J.S.; Lee, H.; Moon, I.S.; Lee, Y.; Koh, Y.H.; Je, Y.H.; Lim, K.J.; Lee, S.H. Construction and characterization of subtractive stage-specific expressed sequence tag (EST) libraries of the pinewood nematode Bursaphelenchus xylophilus. Genomics 2009, 94, 70–77. [Google Scholar] [CrossRef]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Rosso, M.N.; Dubrana, M.P.; Cimbolini, N.; Jaubert, S.; Abad, P. Application of RNA interference to root-knot nematode genes encoding esophageal gland proteins. Mol. Plant Microbe Interact. 2005, 18, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xie, H.; Xu, C.L.; Li, D.L.; Zhang, C. RNAi effect of cathepsin B gene on reproduction of Radopholus similis. Sci. Agric. Sin. 2010, 43, 1608–1616. [Google Scholar] [CrossRef]
- Li, X.D.; Zhuo, K.; Luo, M.; Sun, L.; Liao, J. Molecular cloning and characterization of a calreticulin cDNA from the pinewood nematode Bursaphelenchus xylophilus. Exp. Parasitol. 2011, 128, 121–126. [Google Scholar] [CrossRef]
- Wang, X.R.; Cheng, X.; Li, Y.D.; Zhang, J.A.; Zhang, Z.F.; Wu, H.R. Cloning arginine kinase gene and its RNAi in Bursaphelenchus xylophilus causing pine wilt disease. Eur. J. Plant Pathol. 2012, 134, 521–532. [Google Scholar] [CrossRef]
- Cardoso, J.M.S.; Fonseca, L.; Gomes, P.; Egas, C.; Abrantes, I. Molecular characterization and functional analysis of a calponin gene from the pinewood nematode. Forest Pathol. 2015, 45, 467–473. [Google Scholar] [CrossRef]
- Koiwa, H.; Shade, R.E.; Zhu-Salzman, K.; D’Urzo, M.P.; Murdock, L.L.; Bressan, R.A.; Hasegawa, P.M. A plant defensive cystatin (soya cystatin) targets cathepsin L-like digestive cysteine proteinases (DvCALs) in the larval midgut of western corn rootworm (Diabrotica virgifera). FEBS Lett. 2000, 471, 67–70. [Google Scholar] [CrossRef]
- Lustigman, S.; Zhang, J.; Liu, J.; Oksov, Y.; Hashmi, S. RNA interference targeting cathepsin L and Z-like cysteine proteases of Onchocerca volvulus confirmed their essential function during L3 molting. Mol. Biochem. Parasitol. 2004, 138, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Shinya, R.; Takeuchi, Y.; Futai, K. A technique for separating the developmental stages of the propagative form of the pine wood nematode, Bursaphelenchus xylophilus. Nematology 2009, 11, 305–307. [Google Scholar] [CrossRef]
- Ding, X.L.; Ye, J.R.; Wu, X.Q.; Huang, L.; Zhu, L.H.; Lin, S.X. Deep sequencing analyses of pine wood nematode Bursaphelenchus xylophilus microRNAs reveal distinct miRNA expression patterns during the pathological process of pine wilt disease. Gene. 2015, 555, 346–356. [Google Scholar] [CrossRef]
- He, L.X.; Wu, X.Q.; Xue, Q.; Qiu, X.W. Effects of endobacterium (Stenotrophomonas maltophilia) on pathogenesis-related gene expression of pine wood nematode (Bursaphelenchus xylophilus) and pine wilt disease. Int. J. Mol. Sci. 2016, 17, 778. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Regina, W.; Corinna, W.; Alexandra, F.; Jarutat, T.; Astrid, H.; Tobias, B.; William, D.; Barbara, R. A Method for High Quality Digoxigenin-Labeled RNA Probes for in Situ Hybridization. Available online: http://www.ebiotrade.com/custom/upload/140506/1.pdf (accessed on 20 June 2018).
- De Boer, J.M.; Yan, Y.; Smant, G.; Davis, E.L.; Baum, T.J. In-situ hybridization to messenger RNA in Heterodera glycines. J. Nematol. 1998, 30, 309–312. [Google Scholar] [PubMed]
- Yu, L.Z.; Wu, X.Q.; Ye, J.R.; Zhang, S.N.; Wang, C. NOS-like-mediated nitric oxide is involved in Pinus thunbergii response to the invasion of Bursaphelenchus xylophilus. Plant Cell Rep. 2012, 31, 1813–1821. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Infection Rates (%) | Disease Severity Index (DSI) | ||||||
|---|---|---|---|---|---|---|---|---|
| 5th Day | 8th Day | 20th Day | 35th Day | 5th Day | 8th Day | 20th Day | 35th Day | |
| ddH2O | 25 | 50 | 75 | 100 | 6.25 | 25 | 31.25 | 93.75 |
| gfp dsRNA | 50 | 100 | 100 | 100 | 12.5 | 37.5 | 68.75 | 100 |
| Bx-cpl-1 dsRNA | 0 | 25 | 75 | 100 | 0 | 6.25 | 18.75 | 62.5 |
| Bx-cpl-2 dsRNA | 0 | 0 | 50 | 100 | 0 | 0 | 12.5 | 50 |
| Bx-cpl-3 dsRNA | 0 | 25 | 50 | 100 | 0 | 6.25 | 12.5 | 56.25 |
| Name of Primers | Sequence (5′–3′) |
|---|---|
| cDNA Cloning of Three Cathepsin L-like Cysteine Proteinase Genes | |
| 3′ RACE (rapid amplification of cDNA ends) outer primer | TACCGTCGTTCCACTAGTGATTT |
| 3′ RACE inner primer | CGCGGATCCTCCACTAGTGATTTCACTATAGG |
| GSP (gene specific primer) 1-1 | GCAATGGTGGACTTATGGAC |
| GSP2-1 | AATCCAAGAGCCCCGTTATC |
| GSP3-1 | GCACCTACCGAAGCCGATACTA |
| GSP3-2 | CCACTCCAAGACTACCAAGG |
| 5′RACE outer primer | CATGGCTACATGCTGACAGCCTA |
| 5′RACE inner primer | CGCGGATCCACAGCCTACTGATGATCAGTCGATG |
| GSP1-2 | CTTGACGATCCAGTAGTCGC |
| GSP1-3 | CTCGCCATTTGGTCGCATTT |
| GSP2-2 | GGTTCTATCGCCGACATTCT |
| GSP3-3 | AACCAAAGTGTAGCCCCAAT |
| GSP3-4 | TGACCAAAGCGTTGCGAAGT |
| M13F(−47) | CGCCAGGGTTTTCCCAGTCACGAC |
| M13R(−48) | AGCGGATAACAATTTCACACAGGA |
| Preparation of Template DNA for ISH | |
| I-Bx-cpl-1-F | CCTTTCGCTGAATACCGTCGTCTTA |
| I-Bx-cpl-1-R | TGATGACTCAAGCCAGCGGATAACT |
| I-Bx-cpl-1-T7-F | TAATACGACTCACTATAGGGCCTTTCGCTGAATACCGTCGTCTTA |
| I-Bx-cpl-1-T7-R | TAATACGACTCACTATAGGGTGATGACTCAAGCCAGCGGATAACT |
| I-Bx-cpl-2-F | GCTGTGGATGTTGCTACGCTTTTGC |
| I-Bx-cpl-2-R | GCTTCTCCGTAGTCCTCTCCCCATT |
| I-Bx-cpl-2-T7-F | TAATACGACTCACTATAGGGGCTGTGGATGTTGCTACGCTTTTGC |
| I-Bx-cpl-2-T7-R | TAATACGACTCACTATAGGGGCTTCTCCGTAGTCCTCTCCCCATT |
| I-Bx-cpl-3-F | ACAGCAGTGCCAAGCCCGCTCAAAT |
| I-Bx-cpl-3-R | GTGCTCGGGCATTGATGATTCCTCC |
| I-Bx-cpl-3-T7-F | TAATACGACTCACTATAGGGACAGCAGTGCCAAGCCCGCTCAAAT |
| I-Bx-cpl-3-T7-R | TAATACGACTCACTATAGGGGTGCTCGGGCATTGATGATTCCTCC |
| Preparation of Template DNA for dsRNA | |
| Bx-cpl-1-T7-F | GCCAGTCGTCATCACAAA |
| Bx-cpl-1-R | TGTTCCTCATCGGCTTCT |
| Bx-cpl-1-F | TAATACGACTCACTATAGGGGCCAGTCGTCATCACAAA |
| Bx-cpl-1-T7-R | TAATACGACTCACTATAGGGTGTTCCTCATCGGCTTCT |
| Bx-cpl-2-T7-F | TAATACGACTCACTATAGGGACTAGATCCCAGCGCCACT |
| Bx-cpl-2-R | AGCCAACAGTCACGACAGC |
| Bx-cpl-2-F | ACTAGATCCCAGCGCCACT |
| Bx-cpl-2-T7-R | TAATACGACTCACTATAGGGAGCCAACAGTCACGACAGC |
| Bx-cpl-3-T7-F | TAATACGACTCACTATAGGGAGAGCTTCACAGCAGTGCCAAG |
| Bx-cpl-3-R | GTTGAACCTGGTAACTATAGTC |
| Bx-cpl-3-F | GCTTCACAGCAGTGCCAAG |
| Bx-cpl-3-T7-R | TAATACGACTCACTATAGGGAGAGTTGAACCTGGTAACTATAGTC |
| gfp-T7-F | TAATACGACTCACTATAGGGAGACCATGGCCAACACTTGT |
| gfp-R | AGATAATCCCAGCAGCAGTT |
| gfp-F | AGACCATGGCCAACACTTGT |
| gfp-T7-R | TAATACGACTCACTATAGGGAGATAATCCCAGCAGCAGTT |
| Real Time PCR | |
| q-Bx-cpl-1-F | CCAGAAGCCGATGAGGAACA |
| q-Bx-cpl-1-R | CCAGTTTTGTAGAGTTGGAAGC |
| q-Bx-cpl-2-F | AGTCATCGCTGTAATCTGC |
| q-Bx-cpl-2-R | TTGTTGGTGCCATAAGTG |
| q-Bx-cpl-3-F | CTATAACGGAGTCACCTCCAT |
| q-Bx-cpl-3-R | TGCTCTTCACTGAGATCCAGT |
| Actin-F | GCAACACGGAGTTCGTTGTAGA |
| Actin-R | GTATCGTCACCAACTGGGATGA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, Q.; Wu, X.-Q.; Zhang, W.-J.; Deng, L.-N.; Wu, M.-M. Cathepsin L-like Cysteine Proteinase Genes Are Associated with the Development and Pathogenicity of Pine Wood Nematode, Bursaphelenchus xylophilus. Int. J. Mol. Sci. 2019, 20, 215. https://doi.org/10.3390/ijms20010215
Xue Q, Wu X-Q, Zhang W-J, Deng L-N, Wu M-M. Cathepsin L-like Cysteine Proteinase Genes Are Associated with the Development and Pathogenicity of Pine Wood Nematode, Bursaphelenchus xylophilus. International Journal of Molecular Sciences. 2019; 20(1):215. https://doi.org/10.3390/ijms20010215
Chicago/Turabian StyleXue, Qi, Xiao-Qin Wu, Wan-Jun Zhang, Li-Na Deng, and Miao-Miao Wu. 2019. "Cathepsin L-like Cysteine Proteinase Genes Are Associated with the Development and Pathogenicity of Pine Wood Nematode, Bursaphelenchus xylophilus" International Journal of Molecular Sciences 20, no. 1: 215. https://doi.org/10.3390/ijms20010215
APA StyleXue, Q., Wu, X.-Q., Zhang, W.-J., Deng, L.-N., & Wu, M.-M. (2019). Cathepsin L-like Cysteine Proteinase Genes Are Associated with the Development and Pathogenicity of Pine Wood Nematode, Bursaphelenchus xylophilus. International Journal of Molecular Sciences, 20(1), 215. https://doi.org/10.3390/ijms20010215