Regulatory Mechanism of ABA and ABI3 on Vegetative Development in the Moss Physcomitrella patens

Abstract
1. Introduction
2. Results
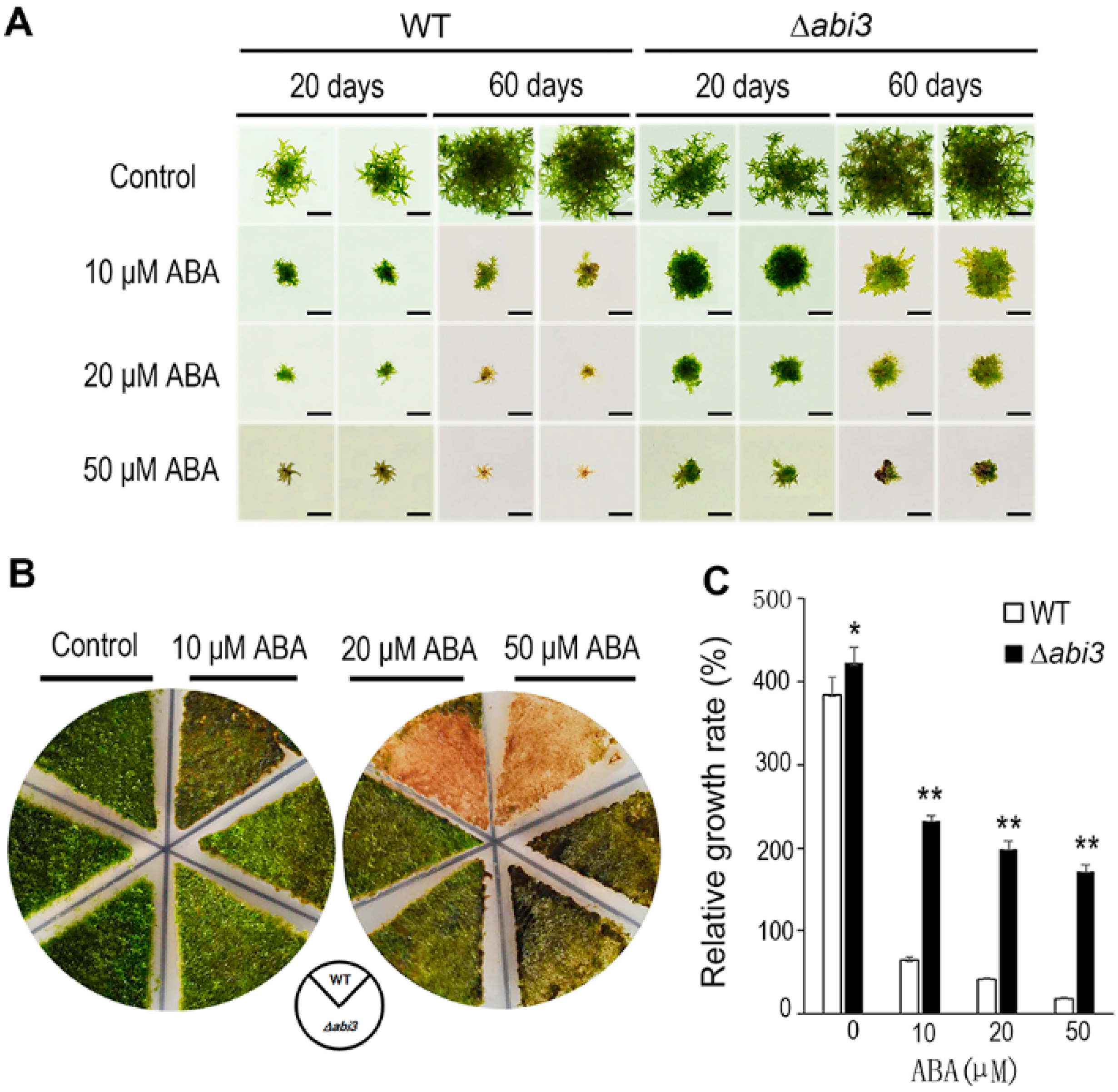
2.1. ∆abi3 Plants Were Insensitivity to ABA Treatment When Compared with WT Plants
2.2. Caulonema Formation Was Accelerated and Chloronema Branching Was Reduced in ∆abi3 Plants, However, ABA Inhibited This Phenotype
2.3. Gametophyte Growth Was Faster and Auxin Concentration Was Higher in ∆abi3 Plants Than Those of WT Plants
2.4. IAA Induced Caulonemata Formation and Inhibited the Formation of Chloronema Branches in WT Plants
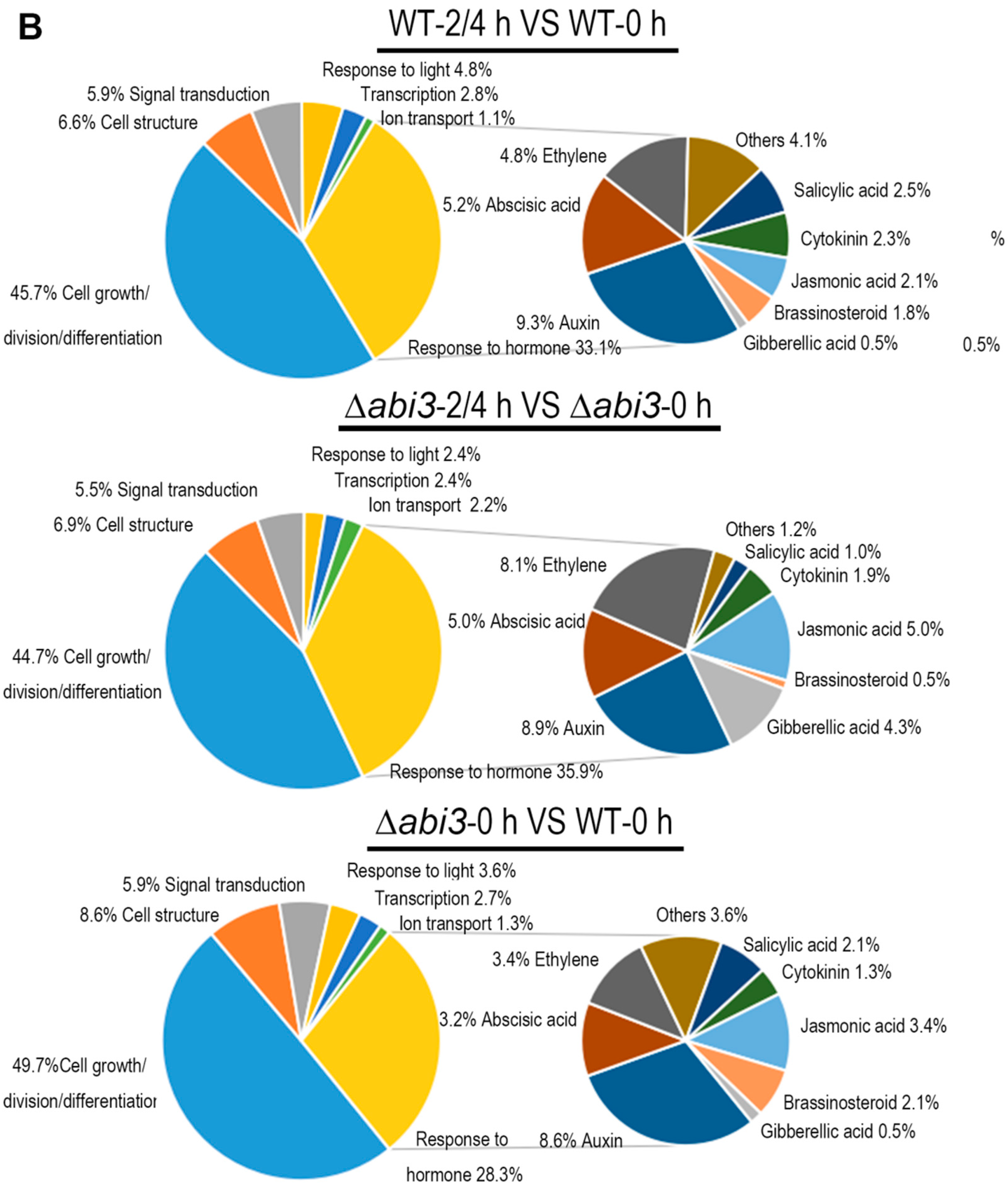
2.5. Role of ABA and ABI3 in Global Gene Expression
2.6. Role of ABA and ABI3 in Growth and Development Gene Expression
2.7. Role of ABA and ABI3 in Auxin-Related Gene Expression
2.8. qRT-PCR Analysis of Auxin-Related Genes
3. Discussion
3.1. ABA and ABI3 Regulate Vegetative Development in P. patens
3.2. Regulatory Mechanism of ABA and ABI3 on Vegetative Development
4. Materials and Methods
4.1. Plant Materials
4.2. Stereoscopic Microscope
4.3. Measurement of Auxin Concentration by Enzyme-Linked Immunosorbent Assay (ELISA)
4.4. RNA Isolation and Sequencing
4.5. RNA-Seq Data Preprocessing
4.6. DEGs and Functional Enrichment Analyses
4.7. Quantitative Real-Time Reverse Transcription PCR (qRT-PCR) Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Medina-Puche, L.; Blanco-Portales, R.; Molina-Hidalgo, F.J.; Cumplido-Laso, G.; García-Caparrósa, N.; Moyano-Cañete, E.; Caballero-Repullo, J.L.; Muñoz-Blanco, J. Extensive transcriptomic studies on the roles played by abscisic acid and auxins in the development and ripening of strawberry fruits. Funct. Integr. Genom. 2016, 16, 671–692. [Google Scholar] [CrossRef] [PubMed]
- Sharp, R.E.; LeNoble, M.E.; Else, M.A.; Thorne, E.T.; Gherardi, F. Endogenous ABA maintains shoot growth in tomato independently of effects on plant water balance: Evidence for an interaction with ethylene. J. Exp. Bot. 2000, 51, 1575–1584. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lin, L.; Ye, T.; Lu, Y.; Xi, C.; Yan, W. The inhibitory effect of ABA on floral transition is mediated by ABI5 in Arabidopsis. J. Exp. Bot. 2013, 64, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Kai, S.; Qian, C.; Wu, Y.; Liu, R.; Zhang, H.; Wang, S.; Tang, S.; Yang, W.; Xie, Q. ABSCISIC ACID-INSENSITIVE negatively regulates flowering through directly promoting Arabidopsis FLOWERING LOCUS C transcription. J. Exp. Bot. 2016, 67, 195–205. [Google Scholar] [CrossRef]
- Riboni, M.; Conti, L. GIGANTEA Enables Drought Escape Response via Abscisic Acid-Dependent Activation of the Florigens and Suppressor of Overexpression of Constans1. Plant Physiol. 2013, 162, 1706–1719. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, Y.; Takahashi, K.; Inoue, S.; Kinoshita, T. Abscisic Acid Suppresses Hypocotyl Elongation by Dephosphorylating Plasma Membrane H+-ATPase in Arabidopsis thaliana. Plant Cell Physiol. 2014, 55, 845–853. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.; Zhao, Y.; Gao, J.; Xiang, C.; Zhu, J.K. The ABA receptor PYL9 together with PYL8 plays an important role in regulating lateral root growth. Sci. Rep. 2016, 6, 27177. [Google Scholar] [CrossRef] [PubMed]
- Signora, L.; De Smet, I.; Foyer, C.H.; Zhang, H.M. ABA plays a central role in mediating them regulatory effects of nitrate on root branching in Arabidopsis. Plant J. 2001, 28, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Kao, C.; Cocciolone, S.; Mccarty, D.R. Maize VP1 complements Arabidopsis abi3 and confers a novel ABA/auxin interaction in roots. Plant J. 2001, 28, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Brocard-Gifford, I.; Lynch, T.J.; Garcia, M.E.; Malhotra, B.; Finkelstein, R.R. The Arabidopsis thaliana ABSCISIC ACID-INSENSITIVE8 Locus Encodes a Novel Protein Mediating Abscisic Acid and Sugar Responses Essential for Growth. Plant Cell 2004, 16, 406–421. [Google Scholar] [CrossRef] [PubMed]
- Antoni, R.; Gonzalez-Guzman, M.; Rodriguez, L.; Peirats-Llobet, M.; Pizzio, G.A.; Fernandez, M.A.; Winne, N.D.; Jaeger, G.D.; Dietrich, D.; Bennett, M.J.; et al. PYRABACTIN RESISTANCE1-LIKE8 Plays an Important Role for the Regulation of Abscisic Acid Signaling in Root. Plant Physiol. 2013, 161, 931–941. [Google Scholar] [CrossRef] [PubMed]
- Finkelstein, R.; Reeves, W.; Ariizumi, T.; Steber, C. Molecular aspects of seed dormancy. Annu. Rev. Plant Biol. 2008, 59, 387–415. [Google Scholar] [CrossRef] [PubMed]
- Rohde, A.; De Rycke, R.; Beeckman, T.; Engler, G.; Van Montagu, M.; Boerjan, W. ABI3 affects plastid differentiation in dark-grown Arabidopsis seedlings. Plant Cell 2000, 12, 35–52. [Google Scholar] [CrossRef] [PubMed]
- Rohde, A.; Prinsen, E.; De Rycke, R.; Engler, G.; Van Montagu, M.; Boerjan, W. PtABI3 impinges on the growth and differentiation of embryonic leaves during bud set in poplar. Plant Cell 2002, 14, 1885–1901. [Google Scholar] [CrossRef] [PubMed]
- Horstman, A.; Li, M.; Heidmann, I.; Weemen, M.; Chen, B.; Muiño, J.M.; Angenent, G.C.; Boutilier, K. The BABY BOOM Transcription Factor Activates the LEC1-ABI3-FUS3-LEC2 Network to Induce Somatic Embryogenesis. Plant Physiol. 2017, 175, 848–857. [Google Scholar] [CrossRef] [PubMed]
- Brady, S.M.; Sarkar, S.F.; Bonetta, D.; Mccourt, P. The ABSCISIC ACID INSENSITIVE 3 (ABI3) gene is modulated by farnesylation and is involved in auxin signaling and lateral root development in Arabidopsis. Plant J. 2003, 34, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, H.; Zhao, Y.; Feng, Z.; Li, Q.; Yang, H.Q.; Luan, S.; Li, J.; He, Z.-H. Auxin controls seed Dormancy through stimulation of abscisic acid signaling by inducing ARF-mediated ABI3 activation in Arabidopsis. Proc. Natl. Acad. Sci. USA 2013, 110, 15485–15490. [Google Scholar] [CrossRef] [PubMed]
- Rensing, S.A.; Lang, D.; Zimmer, A.D.; Terry, A.; Salamov, A.; Shapiro, H.; Nishiyama, T.; Perroud, P.; Lindquist, E.A.; Kamisugi, Y.; et al. The Physcomitrella Genome Reveals Evolutionary Insights into the Conquest of Land by Plants. Science 2008, 319, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Marella, H.H.; Sakata, Y.; Quatrano, R.S. Characterization and functional analysis of ABISCISIC ACID INSENSITIVE3-like genes from Physcomitrella patens. Plant J. 2006, 46, 1032–1044. [Google Scholar] [CrossRef] [PubMed]
- Khandelwal, A.; Cho, S.H.; Marella, H.; Sakata, Y.; Perroud, P.F.; Pan, A.; Quatrano, R.S. Role of ABA and ABI3 in Desiccation Tolerance. Science 2010, 327, 546. [Google Scholar] [CrossRef] [PubMed]
- Tan, T.; Sun, Y.; Peng, X.; Wu, G.; Bao, F.; He, Y.; Zhou, H.; Lin, H. ABSCISIC ACID INSENSITIVE3 Is Involved in Cold Response and Freezing Tolerance Regulation in Physcomitrella patens. Front. Plant Sci. 2017, 8, 1599. [Google Scholar] [CrossRef] [PubMed]
- Goode, J.A.; Stead, A.D.; Duckett, J.G. Redifferentiation of moss Protonemata: An experimental and immunofluorescence study of brood cell formation. Can. J. Bot. 1993, 71, 1510–1519. [Google Scholar] [CrossRef]
- Thelander, M.; Olsson, T.; Ronne, H. Effect of the energy supply on filamentous growth and development in physcomitrella patens. J. Exp. Bot. 2005, 56, 653–662. [Google Scholar] [CrossRef] [PubMed]
- Fujita, T.; Sakaguchi, H.; Hiwatashi, Y.; Wagstaff, S.J.; Ito, M.; Deguchi, H.; Sato, T.; Hasebe, M. Convergent evolution of shoots in land plants: Lack of auxin polar transport in moss shoots. Evol. Dev. 2008, 10, 176–186. [Google Scholar] [CrossRef] [PubMed]
- Bevan, M.; Bancroft, I.; Bent, E.; Love, K.; Goodman, H.; Dean, C.; Bergkamp, R.; Dirkse, W.; van Staveren, M.; Stiekema, W.; et al. Analysis of 1.9 Mb of contiguous sequence from chromosome 4 of Arabidopsis thaliana. Nature 1998, 391, 485–488. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Kamiya, N.; Morinaka, Y.; Matsuoka, M.; Sazuka, T. Auxin Biosynthesi by the YUCCA Genes in Rice. Plant Physiol. 2007, 143, 1362–1371. [Google Scholar] [CrossRef] [PubMed]
- Higgins, C.F. ABC TRANSPORTERS: From Microorganisms to Man. Annu. Rev. Cell Biol. 1992, 8, 67–113. [Google Scholar] [CrossRef] [PubMed]
- Marchant, A.; Kargul, J.; May, S.T.; Muller, P.; Delbarre, A.; Perrot-Rechenmann, C.; Bennett, M.J. AUX1 regulates root gravitropism in Arabidopsis by facilitating auxin uptake within root apical tissues. Embo J. 1999, 18, 2066–2073. [Google Scholar] [CrossRef] [PubMed]
- Paponov, I.A.; Teale, W.; Trebar, M.; Blilou, I.; Palme, K. The PIN auxin efflux facilitators: Evolutionary and functional perspectives. Trends Plant Sci. 2005, 10, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Hammes, U.Z.; Taylor, C.G.; Schachtman, D.P.; Nielsen, E. High-Affinity Auxin Transport by the AUX1 Influx Carrier Protein. Curr. Biol. 2006, 16, 1123–1127. [Google Scholar] [CrossRef] [PubMed]
- Ballas, N.; Wong, L.M.; Ke, M.; Theologis, A. Two auxin-responsive domains interact positively to induce expression of the early indoleacetic acid-inducible gene PS-IAA4/5. Proc. Natl. Acad. Sci. USA 1995, 92, 3483–3487. [Google Scholar] [CrossRef] [PubMed]
- Ulmasov, T.; Hagen, G.; Guilfoyle, T.J. ARF1, a Transcription Factor That Binds to Auxin Response Elements. Science 1997, 276, 1865–1868. [Google Scholar] [CrossRef] [PubMed]
- Hagen, G.; Guilfoyle, T. Auxin-responsive gene expression: Genes, promoters and regulatory factors. Plant Mol. Biol. 2002, 49, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Barrero, J.M.; Piqueras, P.; González-Guzmán, M.; Serrano, R.; Rodríguez, P.L.; Ponce, M.R.; Micol, J.L. A mutational analysis of the ABA1 gene of Arabidopsis thaliana highlights the involvement of ABA in vegetative development. J. Exp. Bot. 2005, 56, 2071–2083. [Google Scholar] [CrossRef] [PubMed]
- Hauser, F.; Waadt, R.; Schroeder, J. Evolution of Abscisic Acid Synthesis and Signaling Mechanisms. Curr. Biol. 2011, 21, R346–R355. [Google Scholar] [CrossRef] [PubMed]
- Fujii, H.; Zhu, J.K. Arabidopsis mutant deficient in 3 abscisic acid-activated protein kinases reveals critical roles in growth, reproduction, and stress. Proc. Natl. Acad. Sci. USA 2009, 106, 8380–8385. [Google Scholar] [CrossRef] [PubMed]
- Gonzalezguzman, M.; Pizzio, G.A.; Antoni, R.; Verasirera, F.; Merilo, E.; Bassel, G.W.; Fernández, M.A.; Holdsworth, M.J. Arabidopsis PYR/PYL/RCAR Receptors Play a Major Role in Quantitative Regulation of Stomatal Aperture and Transcriptional Response to Abscisic Acid. Plant Cell 2012, 24, 2483–2496. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, K.; Suzuki, N.; Kuwamura, M.; Nishikawa, Y.; Nakatani, M.; Ohtawa, H.; Takezawa, D.; Seki, M.; Tanaka, M.; Taji, T.; et al. Group A PP2Cs evolved in land plants as key regulators of intrinsic desiccation tolerance. Nat. Commun. 2013, 4, 2219. [Google Scholar] [CrossRef] [PubMed]
- Johri, M.M.; Desai, S. Auxin regulation of caulonema formation in moss protonema. Nat. New Biol. 1973, 245, 223–224. [Google Scholar] [CrossRef] [PubMed]
- Ashton, N.W.; Grimsley, N.H.; Cove, D.J. Analysis of gametophytic development in the moss, Physcomitrella patens, using auxin and cytokinin resistant mutants. Planta 1979, 14, 427–435. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Duan, Y.; Hua, D.; Fan, G.; Wang, L.; Liu, Y.; Chen, Z.; Han, L.; Qu, Li.; Gong, Z. DEXH Box RNA Helicase-Mediated Mitochondrial Reactive Oxygen Species Production in Arabidopsis Mediates Crosstalk between Abscisic Acid and Auxin Signaling. Plant Cell 2012, 24, 1815–1833. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Li, C.; Wu, Z.; Jia, Y.; Wang, H.; Sun, S.; Mao, C.; Wang, X. Abscisic Acid Regulates Auxin Homeostasis in Rice Root Tips to Promote Root Hair Elongation. Front. Plant Sci. 2017, 8, 1121. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Hua, D.; He, J.; Duan, Y.; Chen, Z.; Hong, X.; Gong, Z. Auxin Response Factor2 (ARF2) and Its Regulated Homeodomain Gene HB33 Mediate Abscisic Acid Response in Arabidopsis. PLoS Genet. 2011, 7, e1002172. [Google Scholar] [CrossRef] [PubMed]
- Belin, C.; Megies, C.; Hauserová, E.; Lopez-Molina, L. Abscisic acid represses growth of the Arabidopsis embryonic axis after germination by enhancing auxin signaling. Plant Cell 2009, 21, 2253–2268. [Google Scholar] [CrossRef] [PubMed]
- Fukaki, H.; Tameda, S.; Masuda, H.; Tasaka, M. Lateral root formation is blocked by a gain-of-function mutation in the SOLITARY-ROOT/IAA14 gene of Arabidopsis. Plant J. 2002, 29, 153–168. [Google Scholar] [CrossRef] [PubMed]
- Tiryaki, I.; Staswick, P.E. An Arabidopsis mutant defective in jasmonate response is allelic to the auxin-signaling mutant axr1. Plant Physiol. 2002, 130, 887–894. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, M.A.; Liu, J.; Enders, T.A.; Bartel, B.; Strader, L.C. A gain-of-function mutation in IAA16 confers reduced responses to auxin and abscisic acid and impedes plant growth and fertility. Plant Mol. Biol. 2012, 79, 359–373. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.L.; Li, Y.Q.; Wu, Y.R.; Liu, Y.C.; Xie, Q. The sHSP22 heat-shock protein requires the ABI1 protein phosphatase to modulate polar auxin transport and downstream responses. Plant Physiol. 2018, 179, 2406–2425. [Google Scholar] [CrossRef] [PubMed]
- Shkolnik-Inbar, D.; Bar-Zvi, D. ABI4 Mediates Abscisic Acid and Cytokinin Inhibition of Lateral Root Formation by Reducing Polar Auxin Transport in Arabidopsis. Plant Cell 2010, 22, 3560–3573. [Google Scholar] [CrossRef] [PubMed]
- Yuan, T.T.; Xu, H.H.; Zhang, K.X.; Guo, T.T.; Lu, Y.T. Glucose inhibits root meristem growth via ABA INSENSITIVE 5, which represses PIN1 accumulation and auxin activity in Arabidopsis. Plant Cell Environ. 2014, 37, 1338–1350. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Symbol | WT-2 h vs. WT-0 h | ∆abi3-2 h vs. ∆abi3-0 h | ∆abi3-0 h vs. WT-0 h ∆abi3-2 h vs. WT-2 h ∆abi3-4 h vs. WT-4 h |
|---|---|---|---|---|
| WT-4 h vs. WT-0 h | ∆abi3-4 h vs. ∆abi3-0 h | |||
| Pp3c11_11790 | YUC6 | U U U | ||
| Pp3c1_11500 | YUC6 | U U U | ||
| Pp3c13_21970 | YUC8 | D D | D D | U U U |
| Pp3c12_5490 | AUX1 | D D | D D | D D D |
| Pp3c15_640 | AUX1 | D D | D D | D D D |
| Pp3c19_12670 | AUX1 | D D D | ||
| Pp3c22_23050 | AUX1 | D D | D D D | |
| Pp3c3_9870 | AUX1 | D D | D D D | |
| Pp3c9_620 | AUX1 | D D | D D | D D D |
| Pp3c6_27190 | AUX1 | D D | D D | |
| Pp3c9_3590 | AUX1 | U U | U U | D D D |
| Pp3c14_8850 | PIN4 | D D | D D | D D D |
| Pp3c23_10200 | PIN7 | D D | D D | D D D |
| Pp3c24_2970 | PIN4 | D D | D D | D D D |
| Pp3c17_16450 | ABCB19 | U U U | ||
| Pp3c6_7260 | ABCB19 | D D | D D | U U U |
| Pp3c12_11230 | ABCB4 | D D | D D | U U U |
| Pp3c24_6610 | IAA7 | D D | U U U | |
| Pp3c8_14720 | IAA7 | U U U | ||
| Pp3c4_13010 | ARF16 | D D | D D | U U U |
| Pp3c5_9420 | ARF2 | D D | D D | U U U |
| Pp3c1_14480 | ARF6 | D D | D D | U U U |
| Pp3c2_25890 | ARF6 | D D | U U U | |
| Pp3c4_12970 | ARF16 | D D | U U U | |
| Pp3c6_21370 | ARF7 | D D | ||
| Pp3c6_26890 | ARF16 | D D | ||
| Pp3c9_17560 | ARF3 | D D | ||
| Pp3c9_21330 | ARF6 | D D | D D | |
| Pp3c13_4720 | ARF6 | D D | D D | |
| Pp3c14_16990 | ARF6 | D D | D D | |
| Pp3c15_21880 | ARF3 | D D | ||
| Pp3c15_9710 | ARF6 | D D | D D |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, M.; Li, Q.; Chen, Z.; Lv, Q.; Bao, F.; Wang, X.; He, Y. Regulatory Mechanism of ABA and ABI3 on Vegetative Development in the Moss Physcomitrella patens. Int. J. Mol. Sci. 2018, 19, 2728. https://doi.org/10.3390/ijms19092728
Zhao M, Li Q, Chen Z, Lv Q, Bao F, Wang X, He Y. Regulatory Mechanism of ABA and ABI3 on Vegetative Development in the Moss Physcomitrella patens. International Journal of Molecular Sciences. 2018; 19(9):2728. https://doi.org/10.3390/ijms19092728
Chicago/Turabian StyleZhao, Mengkai, Qilong Li, Zhenhua Chen, Qiang Lv, Fang Bao, Xiaoqin Wang, and Yikun He. 2018. "Regulatory Mechanism of ABA and ABI3 on Vegetative Development in the Moss Physcomitrella patens" International Journal of Molecular Sciences 19, no. 9: 2728. https://doi.org/10.3390/ijms19092728
APA StyleZhao, M., Li, Q., Chen, Z., Lv, Q., Bao, F., Wang, X., & He, Y. (2018). Regulatory Mechanism of ABA and ABI3 on Vegetative Development in the Moss Physcomitrella patens. International Journal of Molecular Sciences, 19(9), 2728. https://doi.org/10.3390/ijms19092728