The Endochitinase of Clonostachysrosea Expression in Bacillus amyloliquefaciens Enhances the Botrytis cinerea Resistance of Tomato

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Construction of pHT43-ech42
2.2. Transformation of ech42 Gene to B. amyloliquefaciens
2.3. Detection the Growth Curves of B. amyloliquefaciens and Recombinant B. amyloliquefaciens Strains
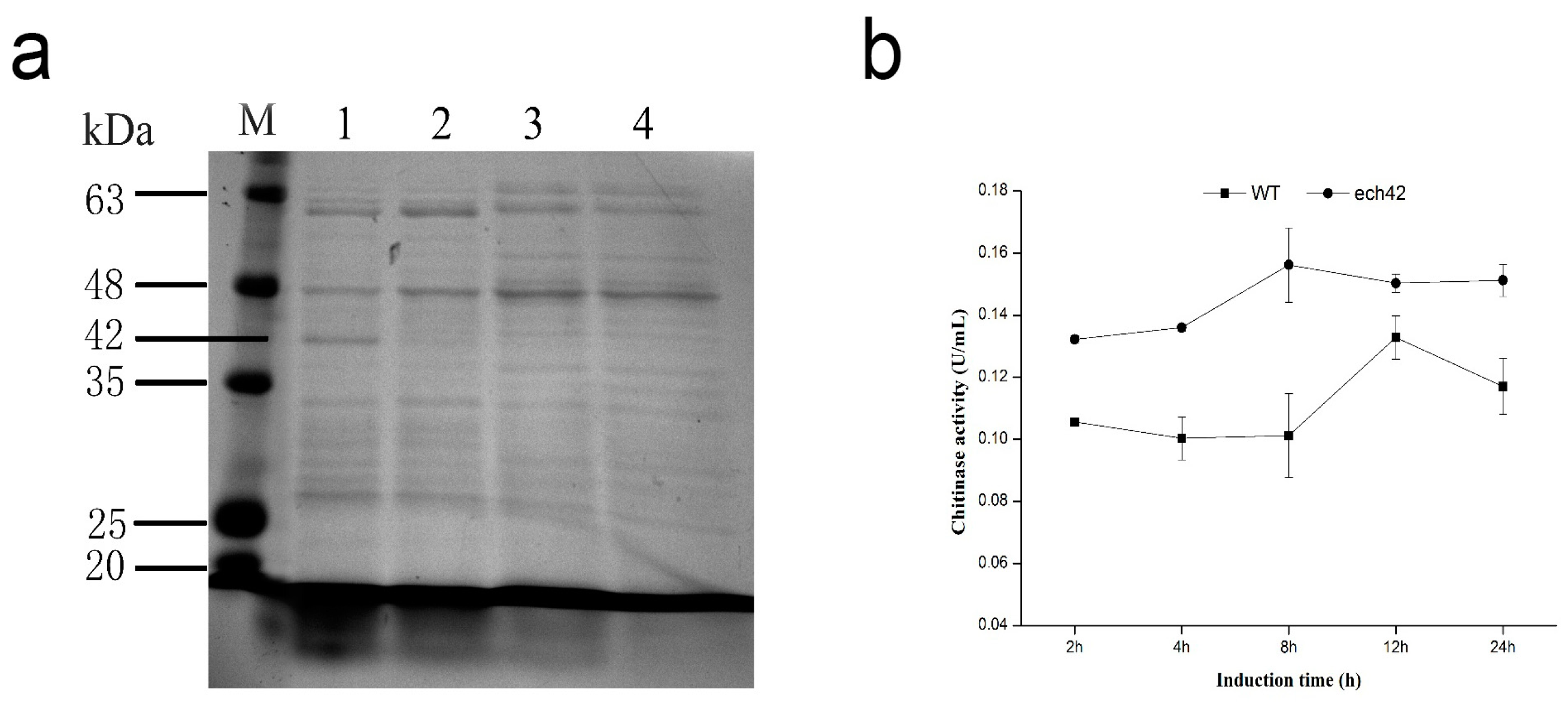
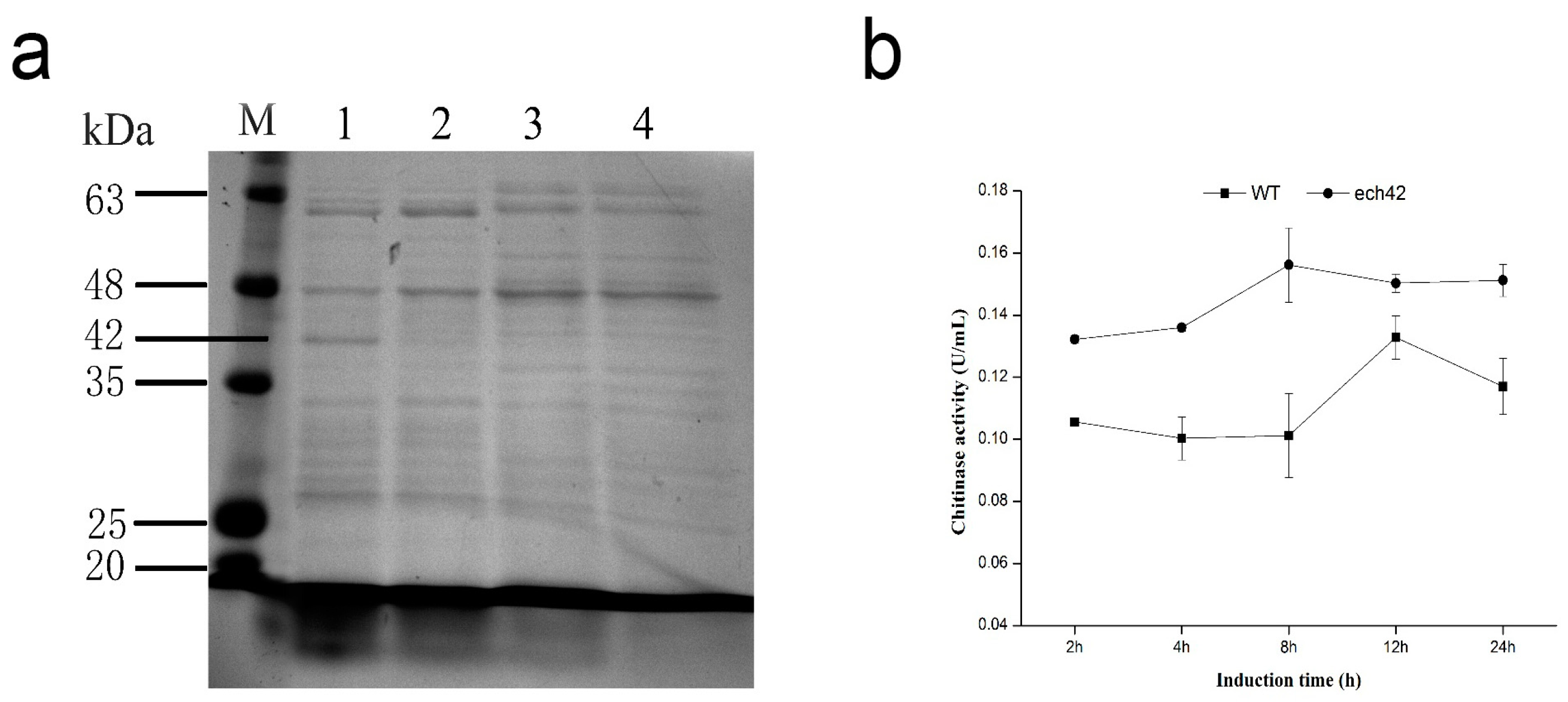
2.4. Detection of Recombinant ech42 Protein by SDS-PAGE
2.5. Chitinase Activity of B. amyloliquefaciens and B. amyloliquefaciens-ech42
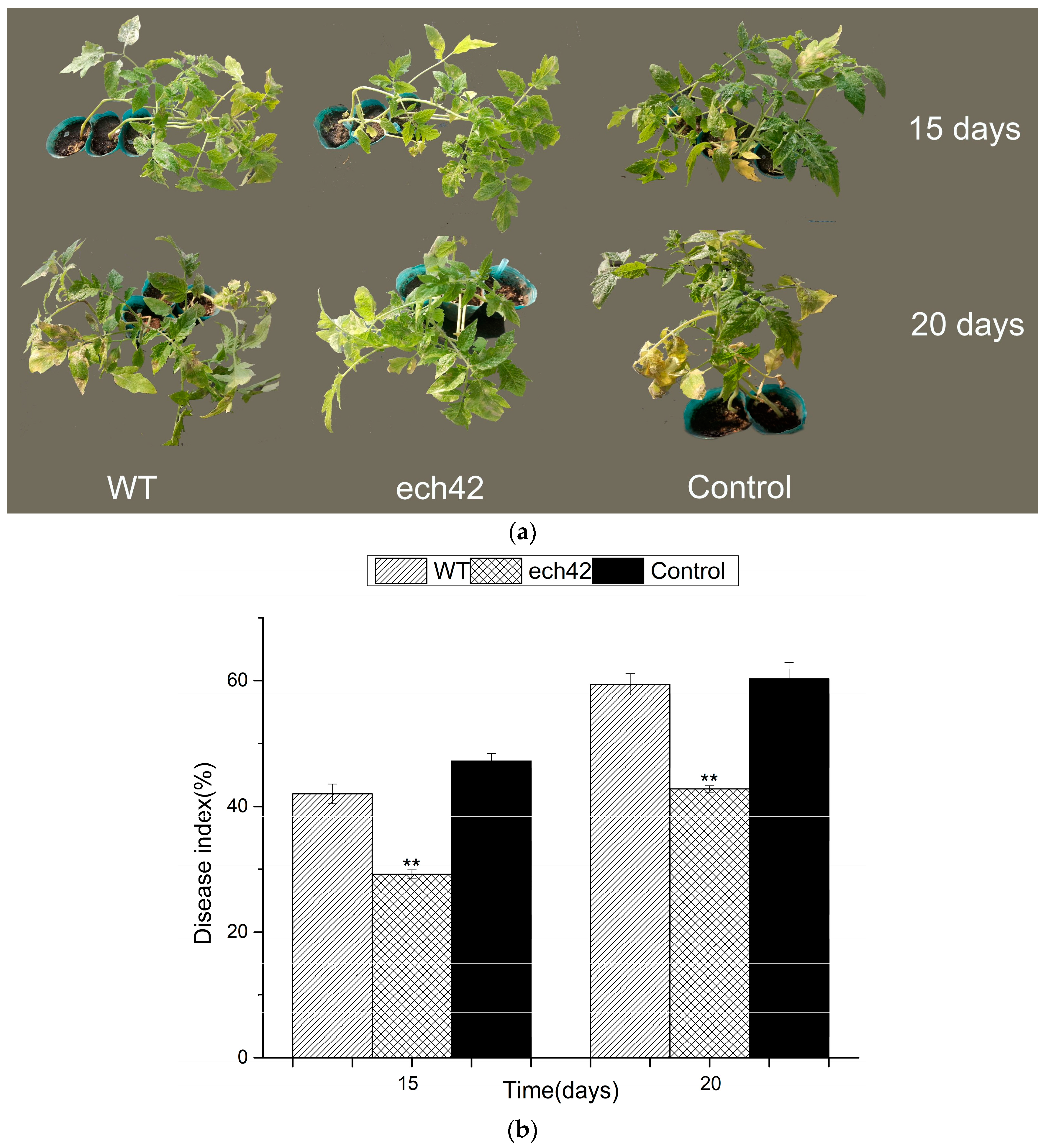
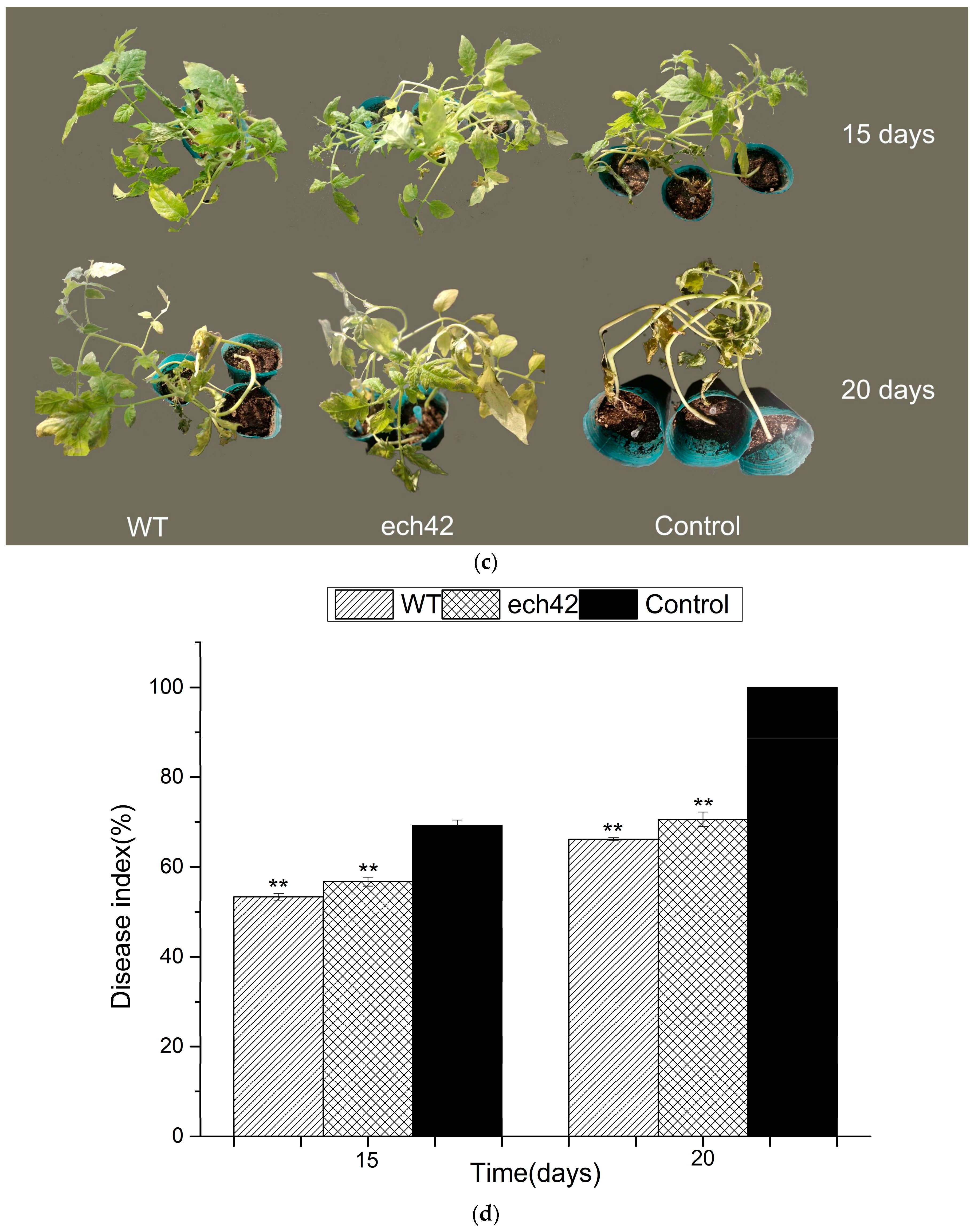
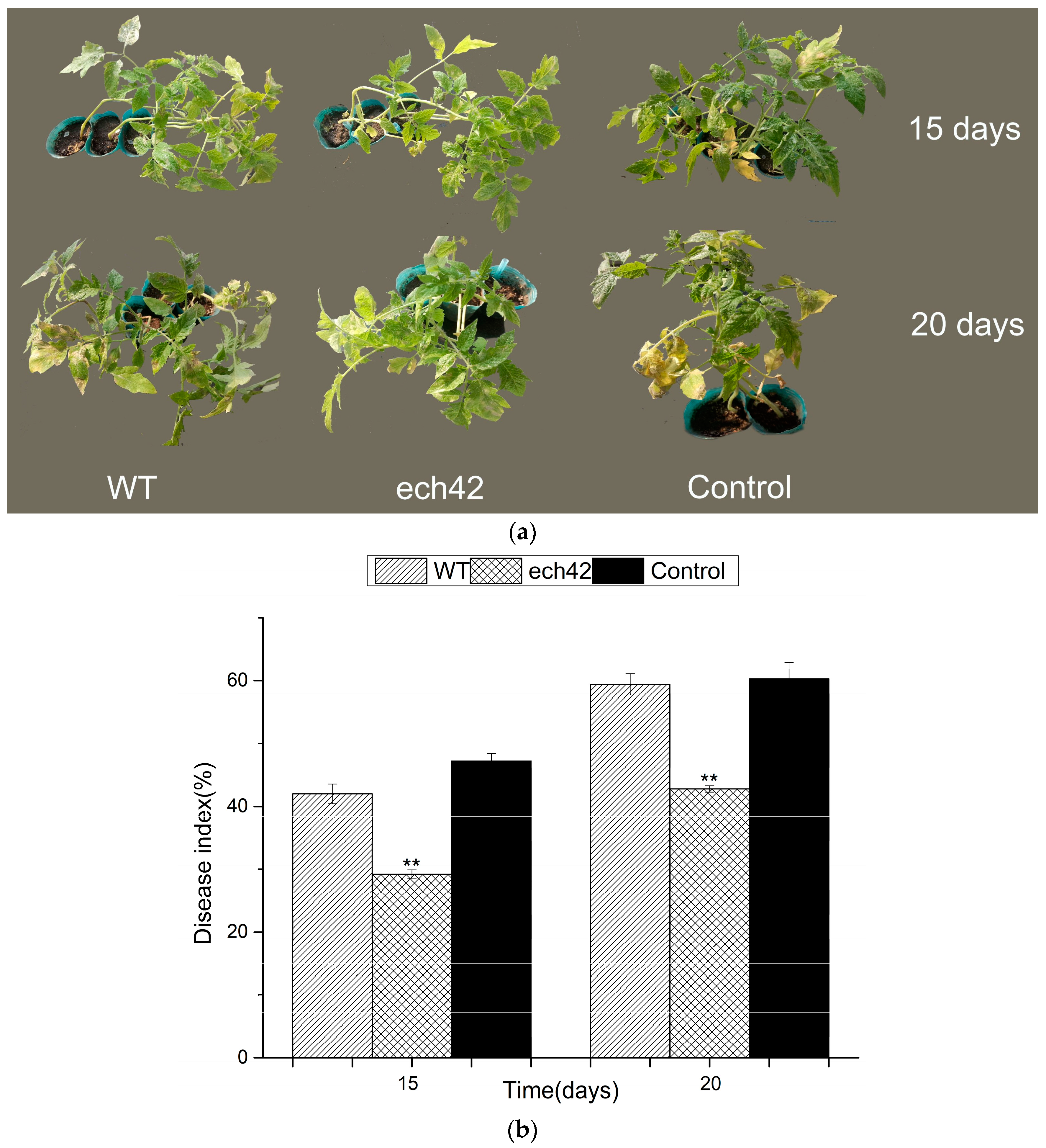
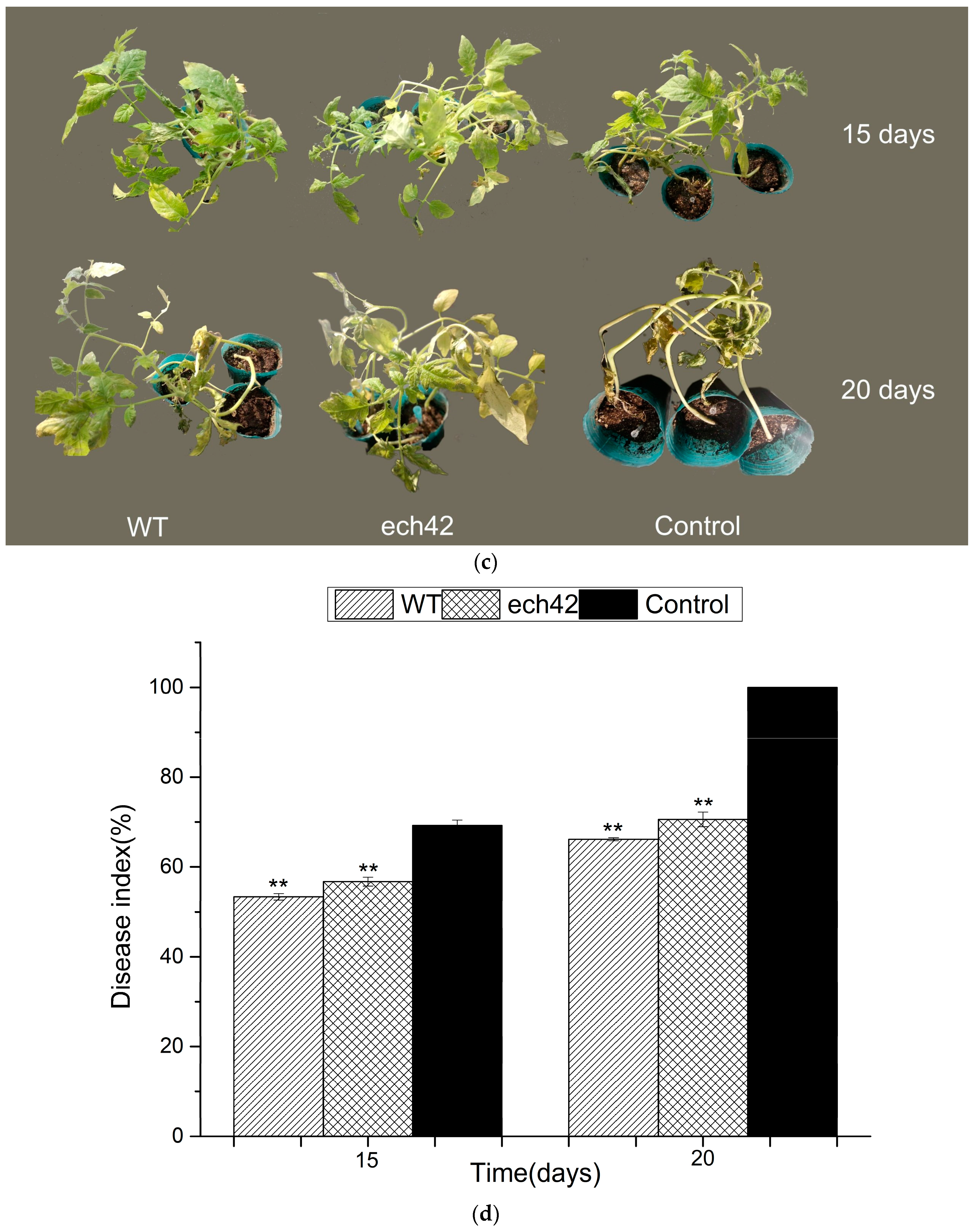
2.6. Biocontrol Efficiency of B. amyloliquefaciens-ech42 vs. Botrytiscinerea in Greenhouse
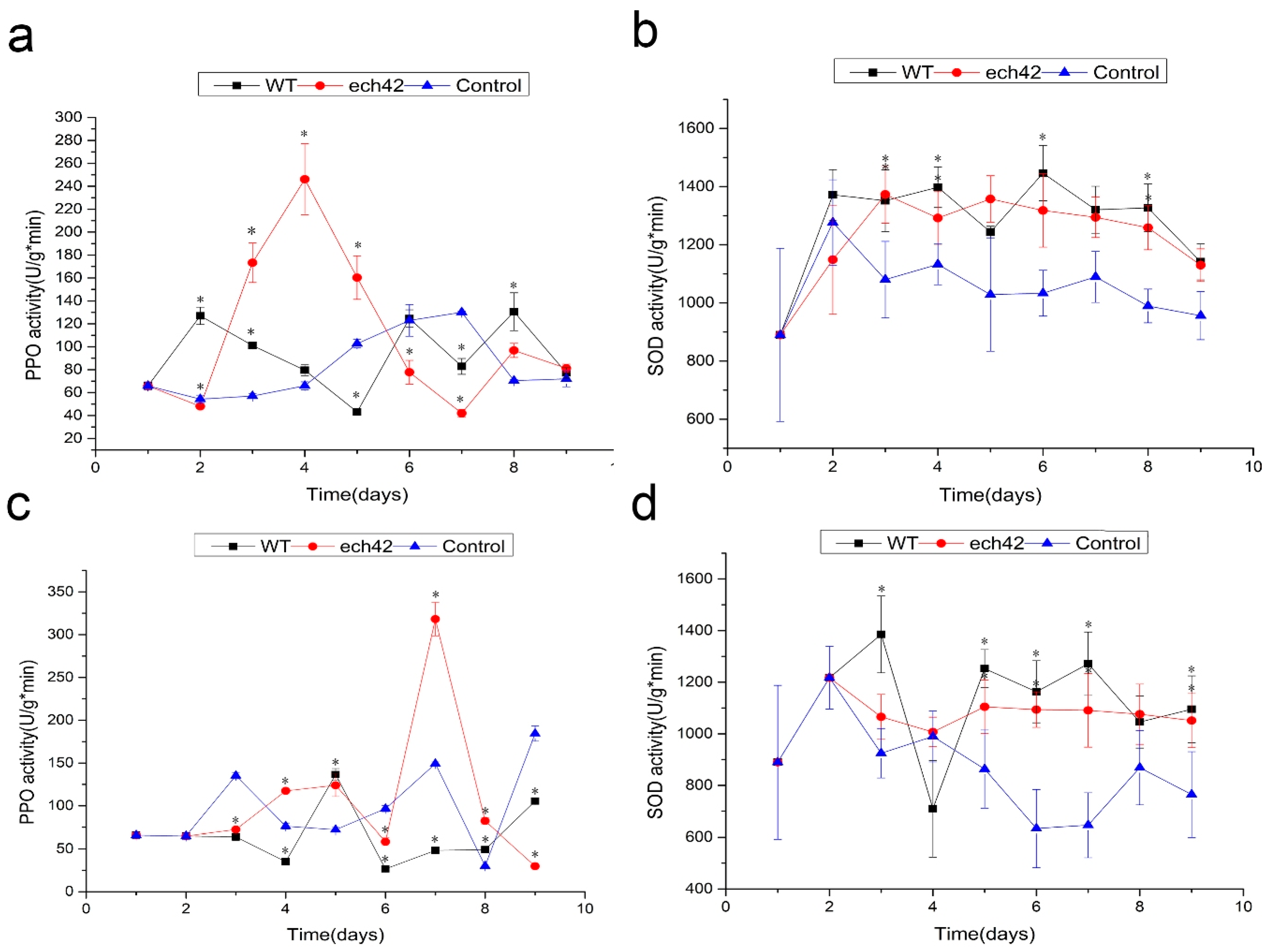
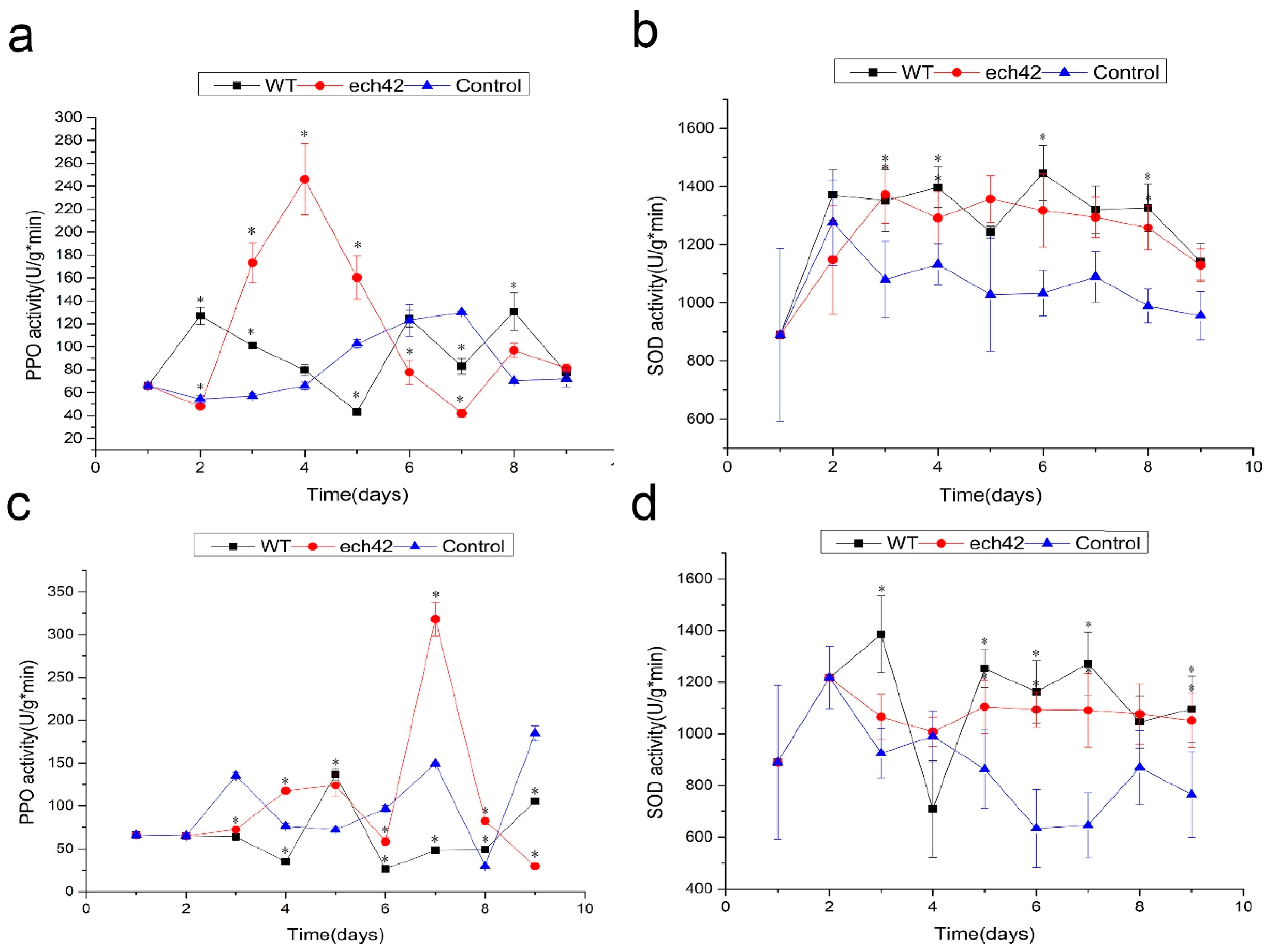
2.7. Changes in Defense-Related Enzyme Activity
3. Discussion
4. Materials and Methods
4.1. Strains and Culture Conditions
4.2. Construction of ech42 Plasmid
4.3. Transformation of ech42 Gene to B. amyloliquefaciens
4.4. Detection the Growth Curves of B. amyloliquefaciens and the Recombinant Strain
4.5. SDS-PAGE Assay
4.6. Determination of Chitinase Activity
4.7. Tomato Plant Growth Experiment
4.8. Defense-Related Enzyme (PPO and SOD) Activity Assays
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Hayyan, M.; Hashim, M.A.; AlNashef, I.M. Superoxide ion: Generation and chemical implications. Chem. Rev. 2016, 116, 3029–3085. [Google Scholar] [CrossRef] [PubMed]
- Krauss, U.; Martinez, A.E.; Ten, H.M.; Arroyo, C. Two-step liquid/solid state scaled-up production of Clonostachys rosea. Mycol. Res. 2002, 106, 1449–1454. [Google Scholar] [CrossRef]
- Xue, A.G. Biological Control of Pathogens Causing Root Rot Complex in Field Pea Using Clonostachys rosea Strain ACM941. Phytopathology 2003, 93, 329–331. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, G.C. The soil microfungi of white cedar forests in ontario. Can. J. Bot. 2011, 48, 333–339. [Google Scholar] [CrossRef]
- Bol, J.F.; Linthorst, H.J.M.; Cornelissen, B.J.C. Plant Pathogenesis-related proteins induced by virus infection. Annu. Rev. Phytopathol. 1990, 28, 113–138. [Google Scholar] [CrossRef]
- Keinath, A.P.; Fravel, D.R.; Papavizas, G.C. Potential of Gliocladiumroseum for biocontrol of Verticilliumdahliae. Phytopathology 1991, 81, 239–243. [Google Scholar] [CrossRef]
- Keyser, C.A.; Jensen, B.; Meyling, N.V. Dual effects of Metarhizium spp. and Clonostachysrosea against an insect and a seed‑borne pathogen in wheat. Pest Manag. Sci. 2016, 72, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Orakc, G.E.; Yamac, M.; Amoreso, M.J.; Cuozzo, S.A. Selection of antagonistic actinomycete isolates as biocontrol agents against root-rot fungi. Fresenius Environ. Bull. 2010, 19, 417–424. [Google Scholar]
- Schöneberg, A.; Musa, T.; Voegele, R.T.; Vogelgsang, S. The potential of antagonistic fungi for control of Fusariumgraminearum and Fusariumcrookwellense varies depending on the experimental approach. J. Appl. Microbiol. 2015, 118, 1165–1179. [Google Scholar] [CrossRef] [PubMed]
- Manuel, D.; Robert, S.G. Antibiotics that inhibit fungal cell wall development. Annu. Rev. Microbiol. 1994, 48, 471–497. [Google Scholar]
- Wenteish, C.; Chinshuh, C.; Sanlang, W. An Antifungal Chitinase Produced by Bacillus cereus with Shrimp and Crab Shell Powder as a Carbon Source. Curr. Microbiol. 2003, 47, 102–108. [Google Scholar]
- Baek, J.M.; Howell, C.R.; Kenerley, C.M. The role of an extracellular chitinase from Trichoderma virens Gv29-8 in the biocontrol of Rhizoctonia solani. Curr. Genet. 1999, 35, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Limón, M.C.; Chacón, M.R.; Mejías, R.; Delgado-Jarana, J.; Rincón, A.M.; Codón, A.C.; Benítez, T. Increased antifungal and chitinase specific activities of Trichoderma harzianum CECT 2413 by addition of a cellulose binding domain. Appl. Microbiol. Biotechnol. 2004, 64, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Viterbo, A.; Haran, S.; Friesem, D.; Ramot, O.; Chet, I. Antifungal activity of a novel endochitinase gene (chit36) from Trichoderma harzianum Rifai TM. FEMS Microbiol. Lett. 2001, 200, 169–174. [Google Scholar] [PubMed]
- Wu, Q.; Bai, L.; Liu, W.; Li, Y.; Lu, C.; Li, Y.; Fu, K.; Yu, C.; Chen, J. Construction of a Streptomyces lydicus A01 transformant with a chit42 gene from Trichoderma harzianum P1 and evaluation of its biocontrol activity against Botrytis cinerea. J. Microbiol. 2013, 51, 166–173. [Google Scholar] [PubMed]
- Siti, N.A.; Nisa, R.M.; Lisdar, I.S. Potential of Chitinolytic Bacillus amyloliquefaciens SAHA 12.07 and Serratia marcescens KAHN 15.12 as Biocontrol Agents of Ganoderma boninense. Res. J. Microbiol. 2015, 10, 452–465. [Google Scholar]
- Horikoshi, K.; Iida, S. Effect of lytic enzyme from Bacillus circulans and chitinase from Streptomyces sp. on Aspergillus oryzae. Nature 1959, 183, 186–187. [Google Scholar] [CrossRef] [PubMed]
- David, B. From natural products to clinically useful antifungals. Biochim. Biophys. Acta 2002, 1587, 224–233. [Google Scholar]
- Chen, X.H.; Koumoutsi, A.; Scholz, R.; Junge, H. Comparative analysis of the complete genome sequence of the plant growth-promoting bacterium Bacillus amyloliquefaciens FZB42. Nat. Biotechnol. 2007, 25, 1007–1014. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.L.; Shih, I.L.; Liang, T.W.; Wang, C.H. Purification and characterization of two antifungal chitinases extracellularly produced by Bacillus amyloliquefaciens V656 in a shrimp and crab shell powder medium. J. Agric. Food Chem. 2002, 50, 2241–2248. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Chen, Y.; Liu, T.; Hu, X.; Gu, Z. Development of a high-efficient transformation system of Bacillus pumilus strain DX01 to facilitate gene isolation via gfp-tagged insertional mutagenesis and visualize bacterial colonization of rice roots. Folia Microbiol. 2013, 58, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Steffens, J.C. Overexpression of polyphenol oxidase in transgenic tomato plants results in enhanced bacterial disease resistance. Planta 2002, 215, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Gong, C.; Liu, Y.; Liu, S.Y.; Cheng, M.Z.; Zhang, Y.; Wang, R.H.; Wang, A.X. Analysis of clonostachys rosea-induced resistance to grey mould disease and identification of the key proteins induced in tomato fruit. Postharvest Biol. Technol. 2017, 123, 83–93. [Google Scholar] [CrossRef]
- Alamelumangai, M.; Dhanalakshmi, J.; Mathumitha, M.; Renganayaki, R.S.; Muthukumaran, P.; Rajalakshmi, N. Modulation of banana polyphenol oxidase (ppo) activity by naturally occurring bioactive compounds from plant extracts. Int. J. Pharma Sci. Res. 2015, 6, 152–155. [Google Scholar]
- Lu, J.; Xue, H.; Pan, Y.; Kan, S.; Liu, M.; Nechitailo, G.S. Effect of spaceflight duration of subcellular morphologies and defense enzyme activities in earth-grown tomato seedlings propagated from space-flown seeds. Russ. J. Phys. Chem. B 2009, 3, 981–986. [Google Scholar] [CrossRef]
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, Y.; Wang, X.; Liu, S.; Zhang, K.; Cai, Z.; Chen, X.; Zhang, Y.; Liu, J.; Wang, A. The Endochitinase of Clonostachysrosea Expression in Bacillus amyloliquefaciens Enhances the Botrytis cinerea Resistance of Tomato. Int. J. Mol. Sci. 2018, 19, 2221. https://doi.org/10.3390/ijms19082221
Zheng Y, Wang X, Liu S, Zhang K, Cai Z, Chen X, Zhang Y, Liu J, Wang A. The Endochitinase of Clonostachysrosea Expression in Bacillus amyloliquefaciens Enhances the Botrytis cinerea Resistance of Tomato. International Journal of Molecular Sciences. 2018; 19(8):2221. https://doi.org/10.3390/ijms19082221
Chicago/Turabian StyleZheng, Yangyang, Xudong Wang, Siyuan Liu, Kewei Zhang, Zhibo Cai, Xiuling Chen, Yao Zhang, Jiayin Liu, and Aoxue Wang. 2018. "The Endochitinase of Clonostachysrosea Expression in Bacillus amyloliquefaciens Enhances the Botrytis cinerea Resistance of Tomato" International Journal of Molecular Sciences 19, no. 8: 2221. https://doi.org/10.3390/ijms19082221
APA StyleZheng, Y., Wang, X., Liu, S., Zhang, K., Cai, Z., Chen, X., Zhang, Y., Liu, J., & Wang, A. (2018). The Endochitinase of Clonostachysrosea Expression in Bacillus amyloliquefaciens Enhances the Botrytis cinerea Resistance of Tomato. International Journal of Molecular Sciences, 19(8), 2221. https://doi.org/10.3390/ijms19082221