Pathogenomics of Endometriosis Development

{kind=link}
Abstract
1. Introduction
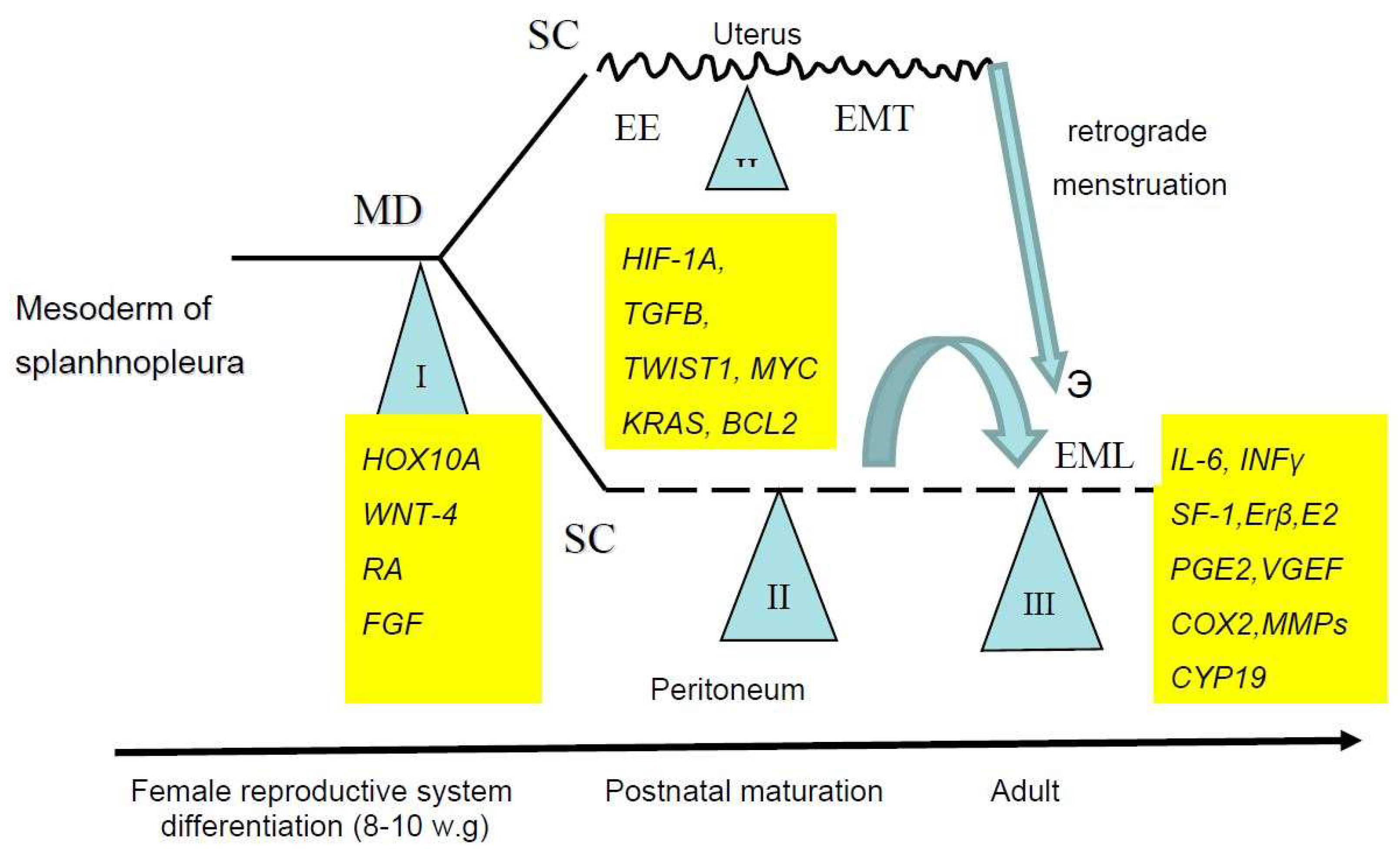
2. Key Stages of Endometriosis Development
2.1. Stem Cells in the Pathogenesis of Endometriosis
2.2. Initial Stages of Endometriosis
2.3. Invasion of Endometriotic SC
3. Discussion
4. Conclusions
Funding
Conflicts of Interest
Abbreviations
| EMDP | Endometriosis development program |
| ESC | Endometrial stem cells |
| SC | Stem cells |
| SP | Sensitive period |
| bmSC | Bone marrow stem cells |
| meSC | Mesenchymal stem cells |
| EMT | Epithelial–mesenchymal transition |
References
- Batt, R.E. A History of Endometriosis, 1st ed.; Springer: London, UK; Dordrecht, The Netherlands; Heidelberg, Germany; New York, NY, USA, 2011; 202p, ISBN 978-0-85729-585-9. [Google Scholar]
- Shubina, A.N.; Egorova, A.A.; Baranov, V.S.; Kiselev, A.V. Recent advances in gene therapy of endometriosis. Recent Pat. DNA Gene Seq. 2013, 7, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Borghese, B.; Zondervan, K.T.; Abrao, M.S.; Chapron, C.; Vaiman, D. Recent insights on the genetics and epigenetics of endometriosis. Clin. Genet. 2017, 91, 254–264. [Google Scholar] [CrossRef] [PubMed]
- Zondervan, K.T.; Rahmioglu, N.; Morris, A.P.; Nyholt, D.R.; Montgomery, G.W.; Becker, C.M.; Missmer, S.A. Beyond endometriosis GWAS: From Genomics to Phenomics to the Patient Europe PMC Funders Group. Semin. Reprod. Med. 2016, 34, 242–254. [Google Scholar] [CrossRef] [PubMed]
- Grimstad, F.W.; Decherney, A. A Review of the Epigenetic Contributions to Endometriosis. Clin. Obstet. Gynecol. 2017, 60, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhao, M. A PubMed-wide study of endometriosis. Genomics 2016, 108, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Akter, S.; Wilshire, G.; Davis, J.W.; Bromfield, J.; Crowder, S.; Pelch, K.; Meng, A.; Barrier, B.; Nagel, S.C. A Multi-Omics Informatics Approach for Identifying Molecular Mechanisms and Biomarkers in Clinical Patients with Endometriosis. In Proceedings of the IEEE International Conference on Bioinformatics and Biomedicine (BIBM), Kansas City, MO, USA, 13–16 November 2017; pp. 2221–2223. [Google Scholar]
- Daraï, E.; Ploteau, S.; Ballester, M.; Bendifallah, S. Endométriose: Physiopathologie, facteurs génétiques etdiagnostic clinique. La Presse Médicale 2017, 46, 1156–1165. [Google Scholar] [CrossRef] [PubMed]
- Aghajanova, L.; Burney, R.O.; Tran, N.D.; Giudice, L.C. mRNA and miRNA Biomarkers for Endometriosis In Biomarkers for Endometriosis; Springer International Publishing AG: Berlin, Germany, 2017; pp. 165–183. ISBN 978-3-319-59856-7. [Google Scholar]
- Saare, M.; Rekker, K.; Laisk-podar, T.; Rahmioglu, N.; Salumets, A.; Martin, G.; Peters, M. Challenges in Endometriosis MiRNA Studies—From Tissue Heterogeneity to Disease Specific MiRNAs. Biochim. Biophys. Acta 2017, 1863, 2282–2292. [Google Scholar] [CrossRef] [PubMed]
- Krishnamoorthy, K.; Decherney, A.H. Genetics of Endometriosis. Clin. Obstet. Gynecol. 2017, 60, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Sapkota, Y.; Steinthorsdottir, V.; Morris, A.P.; Fassbender, A.; Rahmioglu, N.; De Vivo, I.; Buring, J.E.; Zhang, F.; Edwards, T.L.; Jones, S.; et al. Meta-analysis identifies five novel loci associated with endometriosis highlighting key genes involved in hormone metabolism. Nat. Commun. 2017, 8, 15539. [Google Scholar] [CrossRef] [PubMed]
- Stockard, C.R. Developmental rate and structural expression: An experimental study of twins ‘double monsters’ and single deformities, and the interaction among embryonic organs during their origin and development. Dev. Dyn. 1921, 28, 115–277. [Google Scholar] [CrossRef]
- Saxen, L.; Rapila, J. Sensitive periods in development. In Congenital Defects, 1st ed.; Ebert, J.D., Ed.; Holt, Rinehart, Winston: New York, NY, USA, 1969; pp. 112–139. [Google Scholar]
- Baranov, V.S.; Ivaschenko, T.E.; Liehr, T.; Yarmolinskaya, M.I. Systems genetics view of endometriosis: A common complex disorder. Eur. J. Obstet. Gynecol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Gargett, C.E.; Gurung, S. Endometrial Mesenchymal Stem/Stromal Cells, Their Fibroblast Progeny in Endometriosis, and More. Biol. Reprod. 2016, 94, 1291–1295. [Google Scholar] [CrossRef] [PubMed]
- Santamaria, X.; Massasa, E.E.; Taylor, H.S. Migration of cells from experimental endometriosis to the uterine endometrium. Endocrinology 2012, 153, 5566–5574. [Google Scholar] [CrossRef] [PubMed]
- Proestling, K.; Birner, P.; Balendran, S.; Nirtl, N.; Marton, E.; Yerlikaya, G.; Kuessel, L.; Reischer, T.; Wenzl, R.; Streubel, B.; et al. Enhanced expression of the stemness-related factors OCT4, OX15 and TWIST1 in ectopic endometrium of endometriosis patients. Reprod. Biol. Endocrinol. 2016, 14, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lagana, A.S.; Salmeri, F.M.; Vitale, S.G.; Triolo, O.; Gotte, M. Stem Cell Trafficking During Endometriosis: May Epigenetics Play a Pivotal Role? Reprod. Sci. 2017. [Google Scholar] [CrossRef] [PubMed]
- Valentijn, A.J.; Palial, K.; Al-Lamee, H.; Tempest, N.; Drury, J.; Von Zglinicki, T.; Saretzki, G.; Murray, P.; Gargett, C.E.; Hapangama, D.K. SSEA-1 isolates human endometrial basal glandular epithelial cells: Phenotypic and functional characterization and implications in the pathogenesis of endometriosis. Hum. Reprod. 2013, 28, 2695–2708. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Ichim, T.E.; Zhong, J.; Rogers, A.; Yin, Z.; Jackson, J.; Wang, H.; Ge, W.; Bogin, V.; Chan, K.W.; et al. Endometrial regenerative cells: A novel stem cell population. J. Transl. Med. 2007, 5, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Baranov, V.S.; Ivaschenko, T.E.; Yarmolinskaya, M.I. Comparative systems genetics view of endometriosis and uterine leiomyoma: Two sides of the same coin? Syst. Biol. Reprod. Med. 2016, 62, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Mamillapalli, R.; Mutlu, L.; Du, H.; Taylor, H.S. Chemoattraction of bone marrow-derived stem cells towards human endometrial stromal cells is mediated by estradiol regulated CXCL12 and CXCR4 expression. Stem Cell Res. 2015, 15, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Laganà, A.S.; Vitale, S.G.; Salmeri, F.M.; Triolo, O.; Ban Frangež, H.; Vrtačnik-Bokal, E.; Stojanovska, L.; Apostolopoulos, V.; Granese, R.; Sofo, V.; et al. Unus pro omnibus, omnes pro uno: A novel, evidence-based, unifying theory for the pathogenesis of endometriosis. Med. Hypotheses 2017, 103, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Sampson, J.A. Peritoneal endometriosis due to menstrual dissemination of endometrial tissue into peritoneal cavity. Am. J. Obstet. Gynaecol. 1927, 14, 422–469. [Google Scholar] [CrossRef]
- Yang, Y.M.; Yang, W.X. Epithelial-to-mesenchymal transition in the development of endometriosis. Oncotarget 2017, 8, 41679–41689. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef] [PubMed]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef] [PubMed]
- Young, V.J.; Brown, J.K.; Saunders, P.T.; Duncan, W.C.; Horne, A.W. The peritoneum is both a source and target of TGF-β in women with endometriosis. PLoS ONE 2014, 9, e106773. [Google Scholar] [CrossRef] [PubMed]
- Tseng, J.C.; Chen, H.F.; Wu, K.J. A twist tale of cancer metastasis and tumor angiogenesis. Histol. Histopathol. 2015, 30, 1283–1294. [Google Scholar] [PubMed]
- Brunet, T.; Bouclet, A.; Ahmadi, P.; Mitrossilis, D.; Driquez, B.; Brunet, A.C.; Henry, L.; Serman, F.; Béalle, G.; Ménager, C.; et al. Evolutionary conservation of early mesoderm specification by mechanotransduction in Bilateria. Nat. Commun. 2013, 4, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Ito, F.; Yamada, Y.; Shigemitsu, A.; Akinishi, M.; Kaniwa, H.; Miyake, R.; Yamanaka, S.; Kobayashi, H. Role of oxidative stress and epigenetic modification in endometriosis. Reprod. Sci. 2017. [Google Scholar] [CrossRef] [PubMed]
- Capobianco, A.; Rovere-Querini, P. Endometriosis, a disease of the macrophage. Front. Immunol. 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, H.; Iwai, K.; Niiro, E.; Morioka, S.; Yamada, Y. Fetal programming theory: Implication for the understanding of endometriosis. Hum. Immunol. 2014, 75, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Koippallil Gopalakrishnan Nair, A.R.; Pandit, H.; Warty, N.; Madan, T. Endometriotic mesenchymal stem cells exhibit a distinct immune phenotype. Int. Immunol. 2015, 27, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Young, V.J.; Ahmad, S.F.; Brown, J.K.; Duncan, W.C.; Horne, A.W. Peritoneal VEGF-A expression is regulated by TGF-β1 through an ID1 pathway in women withendometriosis. Sci. Rep. 2015, 5, 16859. [Google Scholar] [CrossRef] [PubMed]
- Barragan, F.; Irwin, J.C.; Balayan, S.; Erikson, D.W.; Chen, J.C.; Houshdaran, S.; Piltonen, T.T.; Spitzer, T.L.; George, A.; Rabban, J.T.; et al. Human Endometrial Fibroblasts Derived from Mesenchymal Progenitors Inherit Progesterone Resistance and Acquire an Inflammatory Phenotype in the Endometrial Niche in Endometriosis. Biol. Reprod. 2016, 94, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Pluchino, N.; Taylor, H.S. Endometriosis and Stem Cell Trafficking. Reprod. Sci. 2016, 23, 1616–1619. [Google Scholar] [CrossRef] [PubMed]
- Koukoura, O.; Sifakis, S.; Spandidos, D.A. DNA methylation in endometriosis. Mol. Med. Rep. 2016, 13, 2939–2948. [Google Scholar] [CrossRef] [PubMed]
- Bulun, S.E.; Monsivais, D.; Kakinuma, T.; Furukava, Y.; Barnardi, L.; Pavone, M.E.; Dyson, M. Molecular biology of endometriosis^ from aromatase to genomic abnormalities. Semin. Reprod. Med. 2015, 33, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Waddington, C.H. Tendency towards regularity of development and their genetical control. In International Workshop Teratology; WHO: Copenhagen, Dennmark, 1968; pp. 66–75. [Google Scholar]
- Carlson, B.M. Human Embryology and Developmental Biology, 4th ed.; MOSBY: Maryland Heights, MO, USA, 2009; p. 533. ISBN 9780323082792. [Google Scholar]
- Gaetje, R.; Holtrich, U.; Engels, K.; Kissler, S.; Rody, A.; Karn, T.; Kaufmann, M. Endometriosis may be generated by mimicking the ontogenetic development of the female genital tract. Fertil. Steril. 2007, 87, 651–665. [Google Scholar] [CrossRef] [PubMed]
- Signorile, P.G.; Baldi, F.; Bussani, R.; Viceconte, R.; Bulzomi, P.; D’Armiento, M.; D’Avino, A.; Baldi, A. Embryonic origin of endometriosis: Analysis of 101 human fetuses. J. Cell. Physiol. 2012, 227, 1653–1656. [Google Scholar] [CrossRef] [PubMed]
- Matalliotakis, M.; Zervou, M.I.; Matalliotaki, C.; Rahmioglu, N.; Koumantakis, G.; Kalogiannidis, I.; Prapas, I.; Zondervan, K.; Spandidos, D.A.; Matalliotakis, I.; et al. The role of gene polymorphisms in endometriosis. Mol. Med. Rep. 2017, 16, 5881–5886. [Google Scholar] [CrossRef] [PubMed]
- McKinnon, B.D.; Bertschi, D.; Wanner, J.; Bersinger, N.A.; Mueller, M.D. Hormonal Contraceptive Use and the Prevalence of Endometriotic Lesions at Different Regions within the Peritoneal Cavity. Biomed. Res. Int. 2014, 2014, 590950. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Gu, C.; Ye, M.; Zhang, Z.; Han, W.; Fan, W.; Meng, Y. Identification of global transcriptome abnormalities and potential biomarkers in eutopic endometria of women with endometriosis: A preliminary study. Biomed. Rep. 2017, 6, 654–662. [Google Scholar] [CrossRef] [PubMed]
- Fassbender, A.; Rahimoglu, N.; Vitonis, A.F.; Viganò, P.; Giudice, L.C.; D’Hooghe, T.M.; Hummelshoj, L.; Adamson, G.D.; Becker, C.M.; Missmer, S.A.; et al. World Endometriosis Research Foundation Endometriosis Phenome and Biobanking Harmonisation Project: IV. Tissue collection, processing, and storage in endometriosis research. Fertil. Steril. 2014, 102, 1244–1253. [Google Scholar] [CrossRef] [PubMed]

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baranov, V.; Malysheva, O.; Yarmolinskaya, M. Pathogenomics of Endometriosis Development. Int. J. Mol. Sci. 2018, 19, 1852. https://doi.org/10.3390/ijms19071852
Baranov V, Malysheva O, Yarmolinskaya M. Pathogenomics of Endometriosis Development. International Journal of Molecular Sciences. 2018; 19(7):1852. https://doi.org/10.3390/ijms19071852
Chicago/Turabian StyleBaranov, Vladislav, Olga Malysheva, and Maria Yarmolinskaya. 2018. "Pathogenomics of Endometriosis Development" International Journal of Molecular Sciences 19, no. 7: 1852. https://doi.org/10.3390/ijms19071852
APA StyleBaranov, V., Malysheva, O., & Yarmolinskaya, M. (2018). Pathogenomics of Endometriosis Development. International Journal of Molecular Sciences, 19(7), 1852. https://doi.org/10.3390/ijms19071852