Expression and Regulation of Thymic Stromal Lymphopoietin and Thymic Stromal Lymphopoietin Receptor Heterocomplex in the Innate–Adaptive Immunity of Pediatric Asthma


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Pediatric Asthma
3. Thymic Stromal Lymphopoietin
4. TSLPR Heterocomplex
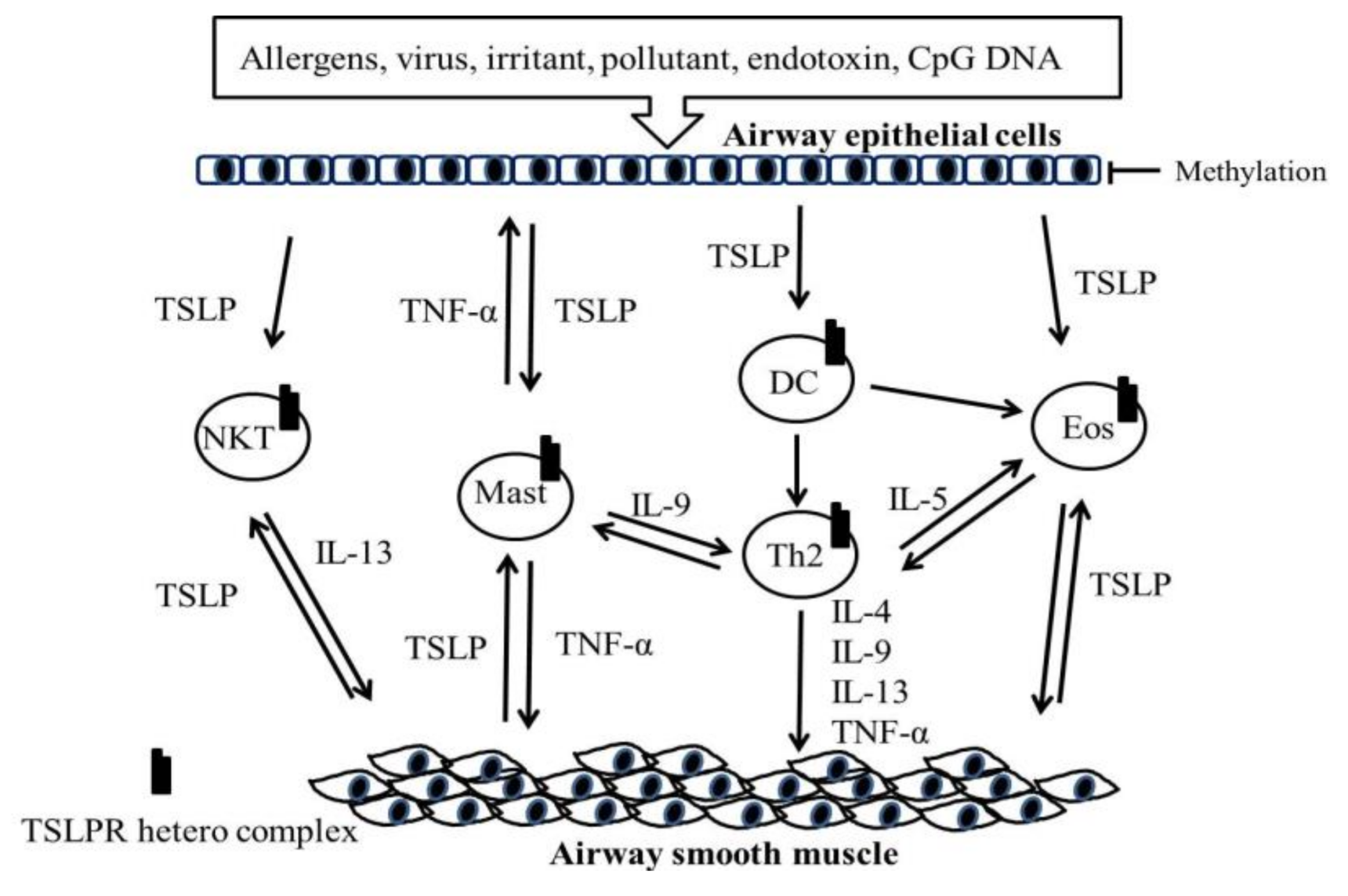
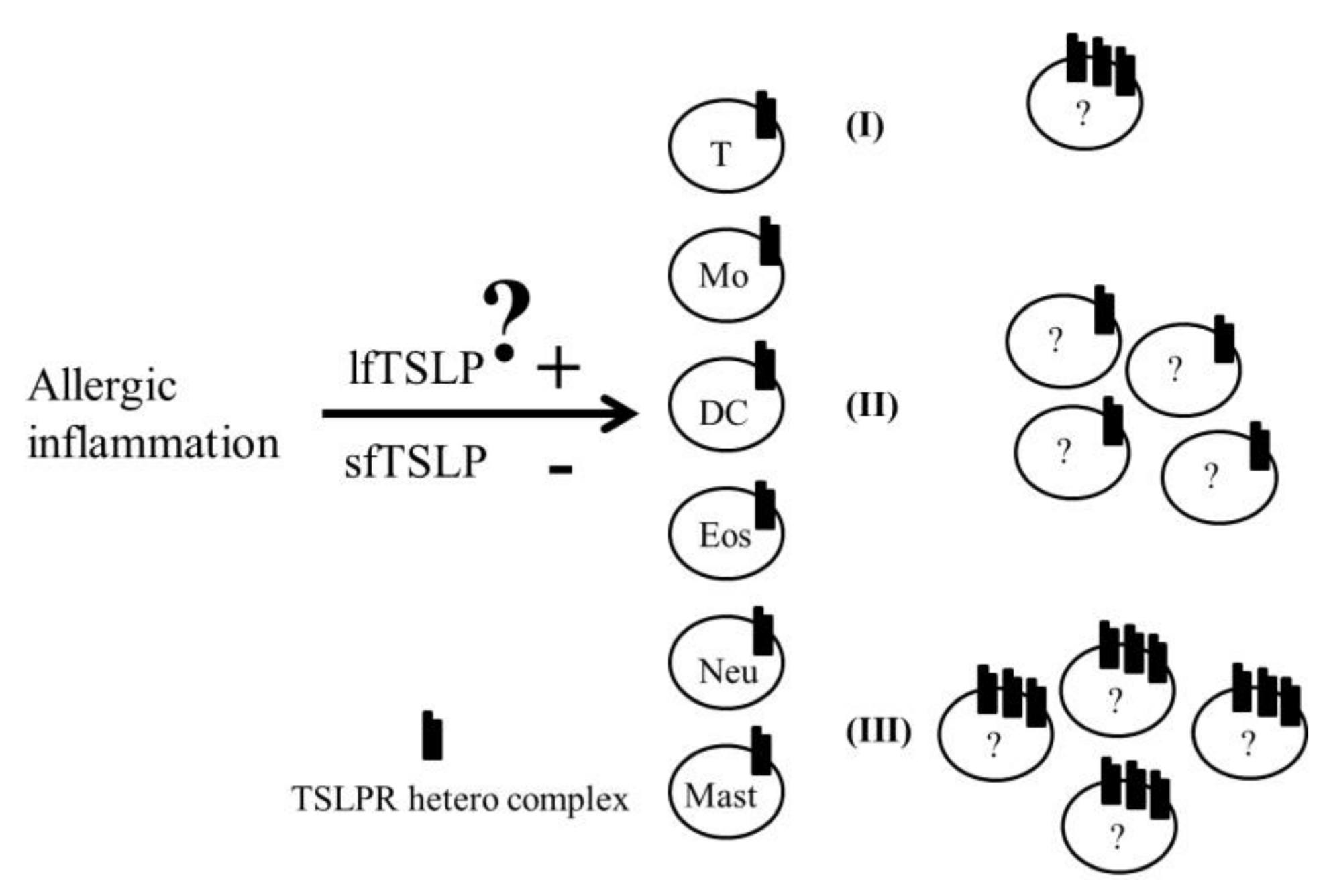
5. Role of the TSLP and TSLPR Heterocomplex in Asthma
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Koczulla, A.R.; Vogelmeier, C.F.; Garn, H.; Renz, H. New concepts in asthma: Clinical phenotypes and pathophysiological mechanisms. Drug Discov. Today 2017, 22, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Mauad, T.; Bel, E.H.; Sterk, P.J. Asthma therapy and airway remodeling. J. Allergy Clin. Immunol. 2007, 120, 997–1009. [Google Scholar] [CrossRef] [PubMed]
- Boulet, L.; Sterk, P. Airway remodelling: The future. Eur. Respir. Soc. 2007, 30, 831–834. [Google Scholar] [CrossRef] [PubMed]
- Royce, S.; Lim, C.; Patel, K.; Wang, B.; Samuel, C.; Tang, M. Intranasally administered serelaxin abrogates airway remodelling and attenuates airway hyperresponsiveness in allergic airways disease. Clin. Exp. Allergy 2014, 44, 1399–1408. [Google Scholar] [CrossRef] [PubMed]
- Bergeron, C.; Al-Ramli, W.; Hamid, Q. Remodeling in asthma. Proc. Am. Thorac. Soc. 2009, 6, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Holgate, S.T. Innate and adaptive immune responses in asthma. Nat. Med. 2012, 18, 673–683. [Google Scholar] [CrossRef] [PubMed]
- McWilliam, A.S.; Nelson, D.; Thomas, J.A.; Holt, P.G. Rapid dendritic cell recruitment is a hallmark of the acute inflammatory response at mucosal surfaces. J. Exp. Med. 1994, 179, 1331–1336. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Liu, Y.-J.; Arima, K. Cellular and molecular mechanisms of TSLP function in human allergic disorders-TSLP programs the “Th2 code” in dendritic cells. Allergol. Int. 2012, 61, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Comeau, M.; Ziegler, S. The influence of TSLP on the allergic response. Mucosal Immunol. 2010, 3, 138. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.K.; Hu, S.; Cheung, P.F.; Lam, C.W. Thymic stromal lymphopoietin induces chemotactic and prosurvival effects in eosinophils: Implications in allergic inflammation. Am. J. Respir. Cell Mol. Biol. 2010, 43, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.H.; Liu, Y.J. Thymic stromal lymphopoietin, OX40-ligand, and interleukin-25 in allergic responses. Clin. Exp. Allergy 2009, 39, 798–806. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, C.M.; Saglani, S. T cells in asthma: Influences of genetics, environment, and T-cell plasticity. J. Allergy Clin. Immunol. 2013, 131, 1267–1274. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.-K.; Kim, T.S.; Hufford, M.M.; Braciale, T.J. Viral infection of the lung: Host response and sequelae. J. Allergy Clin. Immunol. 2013, 132, 1263–1276. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Geha, R.S. Thymic stromal lymphopoietin. Ann. N. Y. Acad. Sci. 2010, 1183, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.Q.; Liao, W. Screening and functional pathway analysis of genes associated with pediatric allergic asthma using a DNA microarray. Mol. Med. Rep. 2015, 11, 4197–4203. [Google Scholar] [CrossRef] [PubMed]
- Szefler, S.J. Advances in pediatric asthma in 2013: Coordinating asthma care. J. Allergy Clin. Immunol. 2014, 133, 654–661. [Google Scholar] [CrossRef] [PubMed]
- Harpsøe, M.C.; Basit, S.; Bager, P.; Wohlfahrt, J.; Benn, C.S.; Nøhr, E.A.; Linneberg, A.; Jess, T. Maternal obesity, gestational weight gain, and risk of asthma and atopic disease in offspring: A study within the Danish National Birth Cohort. J. Allergy Clin. Immunol. 2013, 131, 1033–1040. [Google Scholar] [CrossRef] [PubMed]
- Rzehak, P.; Wijga, A.H.; Keil, T.; Eller, E.; Bindslev-Jensen, C.; Smit, H.A.; Weyler, J.; Dom, S.; Sunyer, J.; Mendez, M. Body mass index trajectory classes and incident asthma in childhood: Results from 8 European Birth Cohorts—A Global Allergy and Asthma European Network initiative. J. Allergy Clin. Immunol. 2013, 131, 1528–1536.e13. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.-W.; Lin, S.-C. Environmental factors association between asthma and acute bronchiolitis in young children—A perspective cohort study. Eur. J. Pediatr. 2012, 171, 1645–1650. [Google Scholar] [CrossRef] [PubMed]
- Dalt, L.D.; Callegaro, S.; Carraro, S.; Andreola, B.; Corradi, M.; Baraldi, E. Nasal lavage leukotrienes in infants with RSV bronchiolitis. Pediatr. Allergy Immunol. 2007, 18, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.-K.; Choi, J.; Kim, H.B.; Callaway, Z.; Shin, B.M.; Kim, J.-T.; Fujisawa, T.; Koh, Y.Y. A randomized intervention of montelukast for post-bronchiolitis: Effect on eosinophil degranulation. J. Pediatr. 2010, 156, 749–754. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-C.; Headley, M.B.; Loo, Y.-M.; Berlin, A.; Gale, M.; Debley, J.S.; Lukacs, N.W.; Ziegler, S.F. Thymic stromal lymphopoietin is induced by respiratory syncytial virus–infected airway epithelial cells and promotes a type 2 response to infection. J. Allergy Clin. Immunol. 2012, 130, 1187–1196.e5. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.-C.; Lin, H.-W.; Chiang, B.-L. Association of croup with asthma in children: A cohort study. Medicine 2017, 96, e7667. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.-J.; Söderhäll, C.; Bustamante, M.; Baïz, N.; Gruzieva, O.; Gehring, U.; Mason, D.; Chatzi, L.; Basterrechea, M.; Llop, S. DNA methylation in childhood asthma: An epigenome-wide meta-analysis. Lancet Respir. Med. 2018. [Google Scholar] [CrossRef]
- Tse, S.M.; Gold, D.R.; Sordillo, J.E.; Hoffman, E.B.; Gillman, M.W.; Rifas-Shiman, S.L.; Fuhlbrigge, A.L.; Tantisira, K.G.; Weiss, S.T.; Litonjua, A.A. Diagnostic accuracy of the bronchodilator response in children. J. Allergy Clin. Immunol. 2013, 132, 554.e5–559.e5. [Google Scholar] [CrossRef] [PubMed]
- National, A.E.; Prevention, P. Expert Panel Report 3 (EPR-3): Guidelines for the Diagnosis and Management of Asthma-Summary Report 2007. J. Allergy Clin. Immunol. 2007, 120 (Suppl. S5), S94–S138. [Google Scholar]
- Malinovschi, A.; Fonseca, J.A.; Jacinto, T.; Alving, K.; Janson, C. Exhaled nitric oxide levels and blood eosinophil counts independently associate with wheeze and asthma events in National Health and Nutrition Examination Survey subjects. J. Allergy Clin. Immunol. 2013, 132, 821.e5–827.e5. [Google Scholar] [CrossRef] [PubMed]
- Oh, M.; Shim, J.Y.; Jung, Y.H.; Seo, J.H.; Young Kim, H.; Kwon, J.W.; Kim, B.J.; Kim, H.B.; Kim, W.K.; Lee, S.Y. Fraction of exhaled nitric oxide and wheezing phenotypes in preschool children. Pediatr. Pulmonol. 2013, 48, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Hoch, H.E.; Szefler, S.J. Intermittent steroid inhalation for the treatment of childhood asthma. Expert Rev. Clin. Immunol. 2016, 12, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Wark, P.A.; McDonald, V.M.; Gibson, P.G. Adjusting prednisone using blood eosinophils reduces exacerbations and improves asthma control in difficult patients with asthma. Respirology 2015, 20, 1282–1284. [Google Scholar] [CrossRef] [PubMed]
- Yoneda, T.; Katayama, N.; Kimura, H.; Fujimura, M. A case of Churg-Strauss syndrome with methylprednisolone sodium succinate allergy. Nihon Kokyuki Gakkai Zasshi 2009, 47, 1147–1150. [Google Scholar] [PubMed]
- Eid, N.S.; O’Hagan, A.; Bickel, S.; Morton, R.; Jacobson, S.; Myers, J.A. Anti-inflammatory dosing of theophylline in the treatment of status asthmaticus in children. J. Asthma Allergy 2016, 9, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Harada, M.; Hirota, T.; Jodo, A.I.; Hitomi, Y.; Sakashita, M.; Tsunoda, T.; Miyagawa, T.; Doi, S.; Kameda, M.; Fujita, K. Thymic stromal lymphopoietin gene promoter polymorphisms are associated with susceptibility to bronchial asthma. Am. J. Respir. Cell Mol. Biol. 2011, 44, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Muchao, F.P.; Torres, H.C.C.; De Lalibera, I.B.; de Souza, A.V.; Rodrigues, J.C.; Schvartsman, C.; da Silva Filho, L.V.R. Albuterol via metered-dose inhaler in children: Lower doses are effective, and higher doses are safe. Pediatr. Pulmonol. 2016, 51, 1122–1130. [Google Scholar] [CrossRef] [PubMed]
- Chou, Y.-L.; Wu, C.-C.; Wang, H.-W. Effects of bambuterol and terbutaline on isolated rat’s tracheal smooth muscle. Eur. Arch. Otorhinolaryngol. 2010, 267, 1305–1311. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Lu, J.Y.-L.; Wu, X.; Summer, S.; Whoriskey, J.; Saris, C.; Reagan, J.D. G-protein-coupled receptor 35 is a target of the asthma drugs cromolyn disodium and nedocromil sodium. Pharmacology 2010, 86, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Horvath, G.; Wanner, A. Inhaled corticosteroids: Effects on the airway vasculature in bronchial asthma. Eur. Respir. J. 2006, 27, 172–187. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.J. Inhaled corticosteroids. Pharmaceuticals 2010, 3, 514–540. [Google Scholar] [CrossRef] [PubMed]
- Gotshall, R.W. Exercise-induced bronchoconstriction. Drugs 2002, 62, 1725–1739. [Google Scholar] [CrossRef] [PubMed]
- Kew, K.M.; Quinn, M.; Quon, B.S.; Ducharme, F.M. Increased versus stable doses of inhaled corticosteroids for exacerbations of chronic asthma in adults and children. Cochrane Libr. 2016. [Google Scholar] [CrossRef] [PubMed]
- Raissy, H.H.; Blake, K. Does use of inhaled corticosteroid for management of asthma in children make them shorter adults? Pediatr. Allergy Immunol. Pulmonol. 2013, 26, 99–101. [Google Scholar] [CrossRef] [PubMed]
- Chee, C.; Sellahewa, L.; Pappachan, J.M. Inhaled corticosteroids and bone health. Open Respir. Med. J. 2014, 8, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Sannarangappa, V.; Jalleh, R. Inhaled corticosteroids and secondary adrenal insufficiency. Open Respir. Med. J. 2014, 8, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Egbuonu, F.; Antonio, F.A.; Edavalath, M. Effect of inhaled corticosteroids on glycemic status. Open Respir. Med. J. 2014, 8, 101–105. [Google Scholar] [PubMed]
- Stanbrook, M.B. Adding salmeterol to fluticasone does not increase serious asthma events or reduce exacerbations in children. Ann. Intern. Med. 2016, 165, Jc58. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.C.; Chuang, Y.H.; Yang, Y.H.; Chiang, B.L. Decrease in interleukin-21 in children suffering with severe atopic dermatitis. Pediatr. Allergy Immunol. 2011, 22, 869–875. [Google Scholar] [CrossRef] [PubMed]
- Vicente, C.T.; Revez, J.A.; Ferreira, M.A. Lessons from ten years of genome-wide association studies of asthma. Clin. Transl. Immunol. 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Reche, P.A.; Soumelis, V.; Gorman, D.M.; Clifford, T.; Liu, M.-R.; Travis, M.; Zurawski, S.M.; Johnston, J.; Liu, Y.-J.; Spits, H. Human thymic stromal lymphopoietin preferentially stimulates myeloid cells. J. Immunol. 2001, 167, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Quentmeier, H.; Drexler, H.; Fleckenstein, D.; Zaborski, M.; Armstrong, A.; Sims, J.; Lyman, S. Cloning of human thymic stromal lymphopoietin (TSLP) and signaling mechanisms leading to proliferation. Leukemia 2001, 15, 1286–1292. [Google Scholar] [CrossRef] [PubMed]
- Sims, J.E.; Williams, D.E.; Morrissey, P.J.; Garka, K.; Foxworthe, D.; Price, V.; Friend, S.L.; Farr, A.; Bedell, M.A.; Jenkins, N.A. Molecular cloning and biological characterization of a novel murine lymphoid growth factor. J. Exp. Med. 2000, 192, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Ray, R.J.; Furlonger, C.; Williams, D.E.; Paige, C.J. Characterization of thymic stromal-derived lymphopoietin (TSLP) in murine B cell development in vitro. Eur. J. Immunol. 1996, 26, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-J. TSLP in epithelial cell and dendritic cell cross talk. Adv. Immunol. 2009, 101, 1–25. [Google Scholar] [PubMed]
- Liu, Y.-J.; Soumelis, V.; Watanabe, N.; Ito, T.; Wang, Y.-H.; de Waal Malefyt, R.; Omori, M.; Zhou, B.; Ziegler, S.F. TSLP: An epithelial cell cytokine that regulates T cell differentiation by conditioning dendritic cell maturation. Annu. Rev. Immunol. 2007, 25, 193–219. [Google Scholar] [CrossRef] [PubMed]
- Moon, P.-D.; Choi, I.-H.; Kim, H.-M. Berberine inhibits the production of thymic stromal lymphopoietin by the blockade of caspase-1/NF-κB pathway in mast cells. Int. Immunopharmacol. 2011, 11, 1954–1959. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, M.; Rochman, Y.; Spolski, R.; Samsel, L.; Leonard, W.J. Thymic stromal lymphopoietin is produced by dendritic cells. J. Immunol. 2011, 187, 1207–1211. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-J. Thymic stromal lymphopoietin: Master switch for allergic inflammation. J. Exp. Med. 2006, 203, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Messaddeq, N.; Teletin, M.; Pasquali, J.-L.; Metzger, D.; Chambon, P. Retinoid X receptor ablation in adult mouse keratinocytes generates an atopic dermatitis triggered by thymic stromal lymphopoietin. Proc. Natl. Acad. Sci. USA 2005, 102, 14795–14800. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, A.; Ebihara, N.; Yokoi, N.; Kawasaki, S.; Tanioka, H.; Inatomi, T.; de Waal Malefyt, R.; Hamuro, J.; Kinoshita, S.; Murakami, A. Functional role of thymic stromal lymphopoietin in chronic allergic keratoconjunctivitis. Investig. Ophthalmol. Vis. Sci. 2010, 51, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Mou, Z.; Xia, J.; Tan, Y.; Wang, X.; Zhang, Y.; Zhou, B.; Li, H.; Han, D. Overexpression of thymic stromal lymphopoietin in allergic rhinitis. Acta Otolaryngol. 2009, 129, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Hartgring, S.; Willis, C.; Dean, C.; Broere, F.; van Eden, W.; Bijlsma, J.; Lafeber, F.; van Roon, J. Critical proinflammatory role of thymic stromal lymphopoietin and its receptor in experimental autoimmune arthritis. Arthritis Rheumatol. 2011, 63, 1878–1887. [Google Scholar] [CrossRef] [PubMed]
- Kuan, E.L.; Ziegler, S.F. Thymic stromal lymphopoietin and cancer. J. Immunol. 2014, 193, 4283–4288. [Google Scholar] [CrossRef] [PubMed]
- Yue, W.; Lin, Y.; Yang, X.; Li, B.; Liu, J.; He, R. Thymic stromal lymphopoietin (TSLP) inhibits human colon tumor growth by promoting apoptosis of tumor cells. Oncotarget 2016, 7, 16840–16854. [Google Scholar] [CrossRef] [PubMed]
- Harada, M.; Hirota, T.; Jodo, A.I.; Doi, S.; Kameda, M.; Fujita, K.; Miyatake, A.; Enomoto, T.; Noguchi, E.; Yoshihara, S. Functional analysis of the thymic stromal lymphopoietin variants in human bronchial epithelial cells. Am. J. Respir. Cell Mol. Biol. 2009, 40, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Bjerkan, L.; Schreurs, O.; Engen, S.; Jahnsen, F.; Baekkevold, E.; Blix, I.J.; Schenck, K. The short form of TSLP is constitutively translated in human keratinocytes and has characteristics of an antimicrobial peptide. Mucosal Immunol. 2015, 8, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Takai, T.; Chen, X.; Okumura, K.; Ogawa, H. Long TSLP transcript expression and release of TSLP induced by TLR ligands and cytokines in human keratinocytes. J. Dermatol. Sci. 2012, 66, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Fornasa, G.; Tsilingiri, K.; Caprioli, F.; Botti, F.; Mapelli, M.; Meller, S.; Kislat, A.; Homey, B.; Di Sabatino, A.; Sonzogni, A. Dichotomy of short and long thymic stromal lymphopoietin isoforms in inflammatory disorders of the bowel and skin. J. Allergy Clin. Immunol. 2015, 136, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Hu, Y.; Liu, L.; Zou, M.; Huang, C.; Luo, L.; Yu, C.; Wan, X.; Zhao, H.; Chen, J. Distinct roles of short and long thymic stromal lymphopoietin isoforms in house dust mite-induced asthmatic airway epithelial barrier disruption. Sci. Rep. 2016, 6, 39559. [Google Scholar] [CrossRef] [PubMed]
- Hunninghake, G.M.; Lasky-Su, J.; Soto-Quirós, M.E.; Avila, L.; Liang, C.; Lake, S.L.; Hudson, T.J.; Spesny, M.; Fournier, E.; Sylvia, J.S. Sex-stratified linkage analysis identifies a female-specific locus for IgE to cockroach in Costa Ricans. Am. J. Respir. Crit. Care Med. 2008, 177, 830–836. [Google Scholar] [CrossRef] [PubMed]
- Vercelli, D. Discovering susceptibility genes for asthma and allergy. Nat. Rev. Immunol. 2008, 8, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Adapa, D.; Sai, Y.; Anand, S.; Mehaboobi, S.; Aramalla, E. A Brief Review on Immune Mediated Diseases. J. Clin. Cell. Immunol. 2011, 11, 2. [Google Scholar]
- Borriello, F.; Iannone, R.; Di Somma, S.; Vastolo, V.; Petrosino, G.; Visconte, F.; Raia, M.; Scalia, G.; Loffredo, S.; Varricchi, G. Lipopolysaccharide-elicited TSLPR expression enriches a functionally discrete subset of human CD14+ CD1c+ monocytes. J. Immunol. 2017, 198, 3426–3435. [Google Scholar] [CrossRef] [PubMed]
- Kummola, L.; Ortutay, Z.; Chen, X.; Caucheteux, S.; Hämäläinen, S.; Aittomäki, S.; Yagi, R.; Zhu, J.; Pesu, M.; Paul, W.E. IL-7Rα Expression Regulates Murine Dendritic Cell Sensitivity to Thymic Stromal Lymphopoietin. J. Immunol. 2017, 198, 3909–3918. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, S.F.; Liu, Y.-J. Thymic stromal lymphopoietin in normal and pathogenic T cell development and function. Nat. Immunol. 2006, 7, 709–714. [Google Scholar] [CrossRef] [PubMed]
- Morshed, M.; Yousefi, S.; Stöckle, C.; Simon, H.U.; Simon, D. Thymic stromal lymphopoietin stimulates the formation of eosinophil extracellular traps. Allergy 2012, 67, 1127–1137. [Google Scholar] [CrossRef] [PubMed]
- Geering, B.; Stoeckle, C.; Conus, S.; Simon, H.-U. Living and dying for inflammation: Neutrophils, eosinophils, basophils. Trends Immunol. 2013, 34, 398–409. [Google Scholar] [CrossRef] [PubMed]
- Han, N.-R.; Oh, H.-A.; Nam, S.-Y.; Moon, P.-D.; Kim, D.-W.; Kim, H.-M.; Jeong, H.-J. TSLP induces mast cell development and aggravates allergic reactions through the activation of MDM2 and STAT6. J. Investig. Dermatol. 2014, 134, 2521–2530. [Google Scholar] [CrossRef] [PubMed]
- Jia, M.; Wang, Z.-J.; Zhao, H.-Z.; Shen, H.-P.; Cheng, Y.-P.; Luo, Z.-B.; Tang, Y.-M. Prognostic significance of cytokine receptor-like factor 2 alterations in acute lymphoblastic leukemia: A meta-analysis. World J. Pediatr. 2015, 11, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Pandey, A. Site-directed mutagenesis reveals a unique requirement for tyrosine residues in IL-7Rα and TSLPR cytoplasmic domains in TSLP-dependent cell proliferation. BMC Immunol. 2010, 11, 5. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-Y.; Lin, C.-C.; Lin, C.G.-J.; Hsiao, Y.-H.; Wu, L.S.-H. Polymorphisms of interleukin 7 receptor are associated with mite-sensitive allergic asthma in children in Taiwan. Tzu Chi Med. J. 2010, 22, 18–23. [Google Scholar] [CrossRef]
- Shamim, Z.; Müller, K.; Svejgaard, A.; Poulsen, L.K.; Bodtger, U.; Ryder, L. Association between genetic polymorphisms in the human interleukin-7 receptor α-chain and inhalation allergy. Int. J. Immunogenet. 2007, 34, 149–151. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Wang, L.; Wu, X. A potential link between TSLP/TSLPR/STAT5 and TLR2/MyD88/NFκB-p65 in human corneal epithelial cells for Aspergillus fumigatus tolerance. Mol. Immunol. 2016, 71, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Redhu, N.; Gounni, A. Function and mechanisms of TSLP/TSLPR complex in asthma and COPD. Clin. Exp. Allergy 2012, 42, 994–1005. [Google Scholar] [CrossRef] [PubMed]
- Durum, S.K. IL-7 and TSLP receptors: Twisted sisters. Blood 2014, 124, 4–5. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Ma, P.; de Paiva, C.S.; Cunningham, M.A.; Hwang, C.S.; Pflugfelder, S.C.; Li, D.-Q. TSLP and downstream molecules in experimental mouse allergic conjunctivitis. Investig. Ophthalmol. Vis. Sci. 2010, 51, 3076–3082. [Google Scholar] [CrossRef] [PubMed]
- Bjerkan, L.; Sonesson, A.; Schenck, K. Multiple functions of the new cytokine-based antimicrobial peptide thymic stromal lymphopoietin (TSLP). Pharmaceuticals 2016, 9, 41. [Google Scholar] [CrossRef] [PubMed]
- Verstraete, K.; Peelman, F.; Braun, H.; Lopez, J.; Van Rompaey, D.; Dansercoer, A.; Vandenberghe, I.; Pauwels, K.; Tavernier, J.; Lambrecht, B.N. Structure and antagonism of the receptor complex mediated by human TSLP in allergy and asthma. Nat. Commun. 2017, 8, 14937. [Google Scholar] [CrossRef] [PubMed]
- Sinha, S.; Singh, J.; Jindal, S.K. Association of interleukin 7 receptor (rs1494555 and rs6897932) gene polymorphisms with asthma in a north Indian population. Allergy Rhinol. 2015, 6, e168. [Google Scholar] [CrossRef] [PubMed]
- Sherrill, J.D.; Gao, P.-S.; Stucke, E.M.; Blanchard, C.; Collins, M.H.; Putnam, P.E.; Franciosi, J.P.; Kushner, J.P.; Abonia, J.P.; Assa’ad, A.H. Variants of thymic stromal lymphopoietin and its receptor associate with eosinophilic esophagitis. J. Allergy Clin. Immunol. 2010, 126, 160–165.e3. [Google Scholar] [CrossRef] [PubMed]
- Gao, P.-S.; Rafaels, N.M.; Mu, D.; Hand, T.; Murray, T.; Boguniewicz, M.; Hata, T.; Schneider, L.; Hanifin, J.M.; Gallo, R.L. Genetic variants in thymic stromal lymphopoietin are associated with atopic dermatitis and eczema herpeticum. J. Allergy Clin. Immunol. 2010, 125, 1403–1407. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Huang, G.; Hu, B.; Song, Y.; Shi, Y. A soluble thymic stromal lymphopoietin (TSLP) antagonist, TSLPR-immunoglobulin, reduces the severity of allergic disease by regulating pulmonary dendritic cells. Clin. Exp. Immunol. 2011, 164, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Alves, N.L.; van Leeuwen, E.M.; Derks, I.A.; van Lier, R.A. Differential regulation of human IL-7 receptor α expression by IL-7 and TCR signaling. J. Immunol. 2008, 180, 5201–5210. [Google Scholar] [CrossRef] [PubMed]
- Ashbaugh, J.J.; Brambilla, R.; Karmally, S.A.; Cabello, C.; Malek, T.R.; Bethea, J.R. IL7Rα contributes to experimental autoimmune encephalomyelitis through altered T cell responses and nonhematopoietic cell lineages. J. Immunol. 2013, 190, 4525–4534. [Google Scholar] [CrossRef] [PubMed]
- Ying, S.; O’Connor, B.; Ratoff, J.; Meng, Q.; Mallett, K.; Cousins, D.; Robinson, D.; Zhang, G.; Zhao, J.; Lee, T.H. Thymic stromal lymphopoietin expression is increased in asthmatic airways and correlates with expression of Th2-attracting chemokines and disease severity. J. Immunol. 2005, 174, 8183–8190. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, K.D.; Vanichsarn, C.; Nadeau, K.C. TSLP directly impairs pulmonary Treg function: Association with aberrant tolerogenic immunity in asthmatic airway. Allergy Asthma Clin. Immunol. 2010, 6, 4. [Google Scholar] [CrossRef] [PubMed]
- Semlali, A.; Jacques, E.; Koussih, L.; Gounni, A.S.; Chakir, J. Thymic stromal lymphopoietin–induced human asthmatic airway epithelial cell proliferation through an IL-13–dependent pathway. J. Allergy Clin. Immunol. 2010, 125, 844–850. [Google Scholar] [CrossRef] [PubMed]
- Uller, L.; Leino, M.; Bedke, N.; Sammut, D.; Green, B.; Lau, L.; Howarth, P.H.; Holgate, S.T.; Davies, D.E. Double-stranded RNA induces disproportionate expression of thymic stromal lymphopoietin versus interferon-β in bronchial epithelial cells from donors with asthma. Thorax 2010, 65, 626–632. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.-I.; Kang, I.-H.; Chun, S.-W.; Yun, K.-J.; Moon, H.-B.; Chae, S.-C. Identifying the polymorphisms in the thymic stromal lymphopoietin receptor (TSLPR) and their association with asthma. BMB Rep. 2010, 43, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Myers, J.M.B.; Martin, L.J.; Kovacic, M.B.; Mersha, T.B.; He, H.; Pilipenko, V.; Lindsey, M.A.; Ericksen, M.B.; Bernstein, D.I.; LeMasters, G.K. Epistasis between serine protease inhibitor Kazal-type 5 (SPINK5) and thymic stromal lymphopoietin (TSLP) genes contributes to childhood asthma. J. Allergy Clin. Immunol. 2014, 134, 891.e3–899.e3. [Google Scholar] [CrossRef] [PubMed]
- Kabata, H.; Moro, K.; Fukunaga, K.; Suzuki, Y.; Miyata, J.; Masaki, K.; Betsuyaku, T.; Koyasu, S.; Asano, K. Thymic stromal lymphopoietin induces corticosteroid resistance in natural helper cells during airway inflammation. Nat. Commun. 2013, 4, 2675. [Google Scholar] [CrossRef] [PubMed]
- Gauvreau, G.M.; O’byrne, P.M.; Boulet, L.-P.; Wang, Y.; Cockcroft, D.; Bigler, J.; FitzGerald, J.M.; Boedigheimer, M.; Davis, B.E.; Dias, C. Effects of an anti-TSLP antibody on allergen-induced asthmatic responses. N. Engl. J. Med. 2014, 370, 2102–2110. [Google Scholar] [CrossRef] [PubMed]
- Corren, J.; Parnes, J.R.; Wang, L.; Mo, M.; Roseti, S.L.; Griffiths, J.M.; van der Merwe, R. Tezepelumab in Adults with Uncontrolled Asthma. N. Engl. J. Med. 2017, 377, 936–946. [Google Scholar] [CrossRef] [PubMed]
- Oksel, C.; Custovic, A. Development of allergic sensitization and its relevance to paediatric asthma. Curr. Opin. Allergy Clin. Immunol. 2018, 18, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.-C.; Huang, J.-J.; Wang, J.-Y.; Chuang, H.-C.; Chiang, B.-L.; Ye, Y.-L. Upregulated thymic stromal lymphopoietin receptor expression in children with asthma. Eur. J. Clin. Investig. 2016, 46, 511–519. [Google Scholar] [CrossRef] [PubMed]
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, S.-C.; Cheng, F.-Y.; Liu, J.-J.; Ye, Y.-L. Expression and Regulation of Thymic Stromal Lymphopoietin and Thymic Stromal Lymphopoietin Receptor Heterocomplex in the Innate–Adaptive Immunity of Pediatric Asthma. Int. J. Mol. Sci. 2018, 19, 1231. https://doi.org/10.3390/ijms19041231
Lin S-C, Cheng F-Y, Liu J-J, Ye Y-L. Expression and Regulation of Thymic Stromal Lymphopoietin and Thymic Stromal Lymphopoietin Receptor Heterocomplex in the Innate–Adaptive Immunity of Pediatric Asthma. International Journal of Molecular Sciences. 2018; 19(4):1231. https://doi.org/10.3390/ijms19041231
Chicago/Turabian StyleLin, Sheng-Chieh, Fang-Yi Cheng, Jun-Jen Liu, and Yi-Ling Ye. 2018. "Expression and Regulation of Thymic Stromal Lymphopoietin and Thymic Stromal Lymphopoietin Receptor Heterocomplex in the Innate–Adaptive Immunity of Pediatric Asthma" International Journal of Molecular Sciences 19, no. 4: 1231. https://doi.org/10.3390/ijms19041231
APA StyleLin, S.-C., Cheng, F.-Y., Liu, J.-J., & Ye, Y.-L. (2018). Expression and Regulation of Thymic Stromal Lymphopoietin and Thymic Stromal Lymphopoietin Receptor Heterocomplex in the Innate–Adaptive Immunity of Pediatric Asthma. International Journal of Molecular Sciences, 19(4), 1231. https://doi.org/10.3390/ijms19041231