Genetic Variants in STAT3 Promoter Regions and Their Application in Molecular Breeding for Body Size Traits in Qinchuan Cattle

 , ,
, ,
Abstract
:
1. Introduction
2. Results
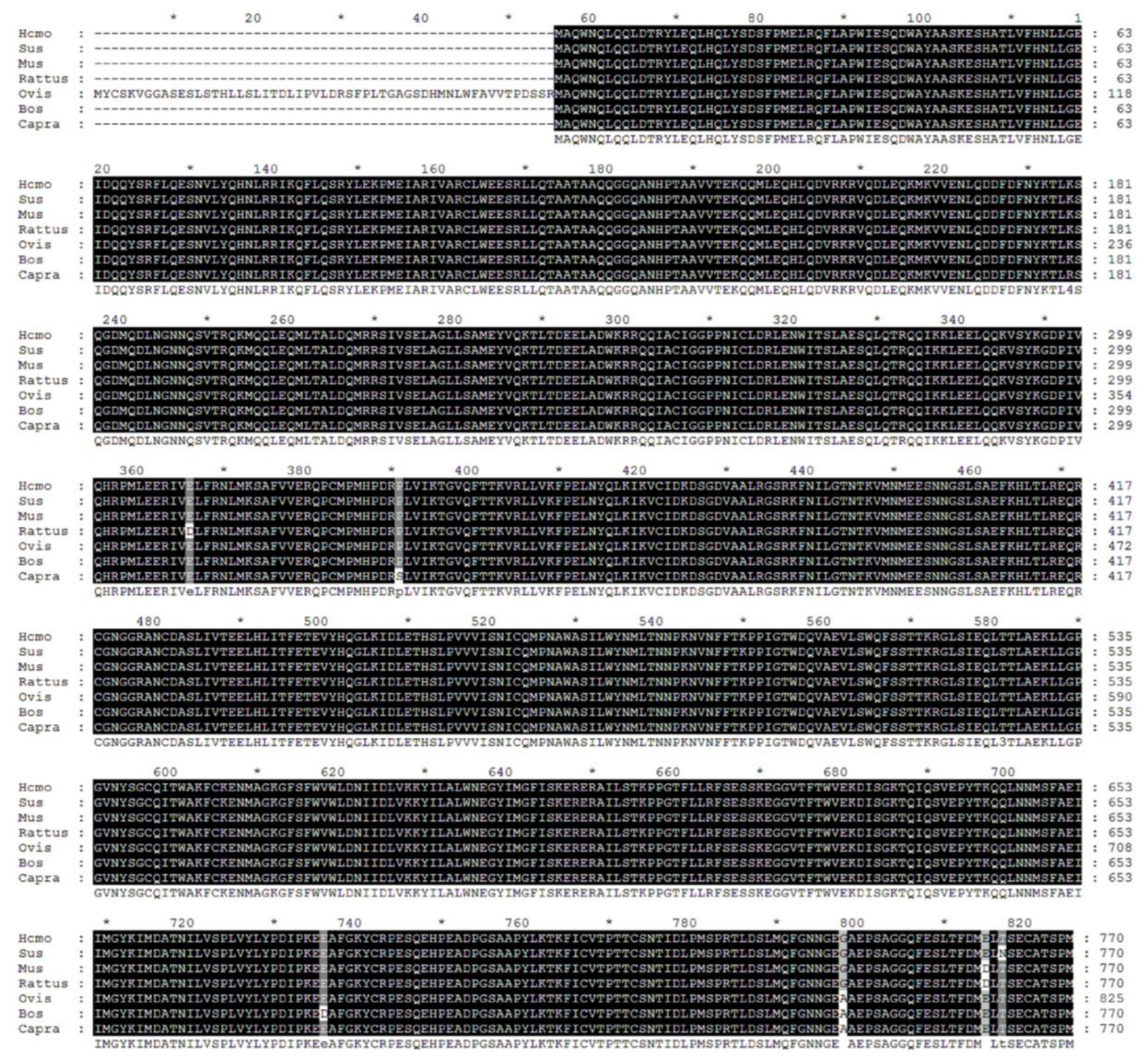
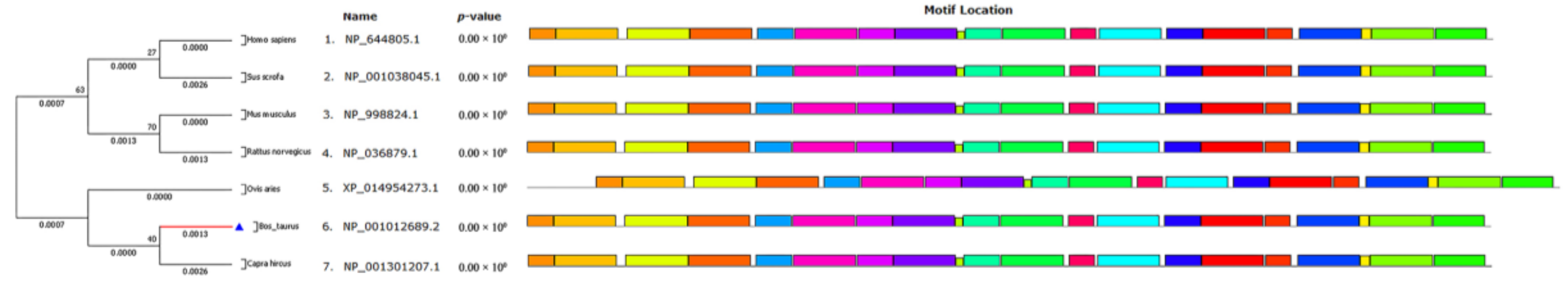
2.1. Biological Evolution and Estimates of Conservation
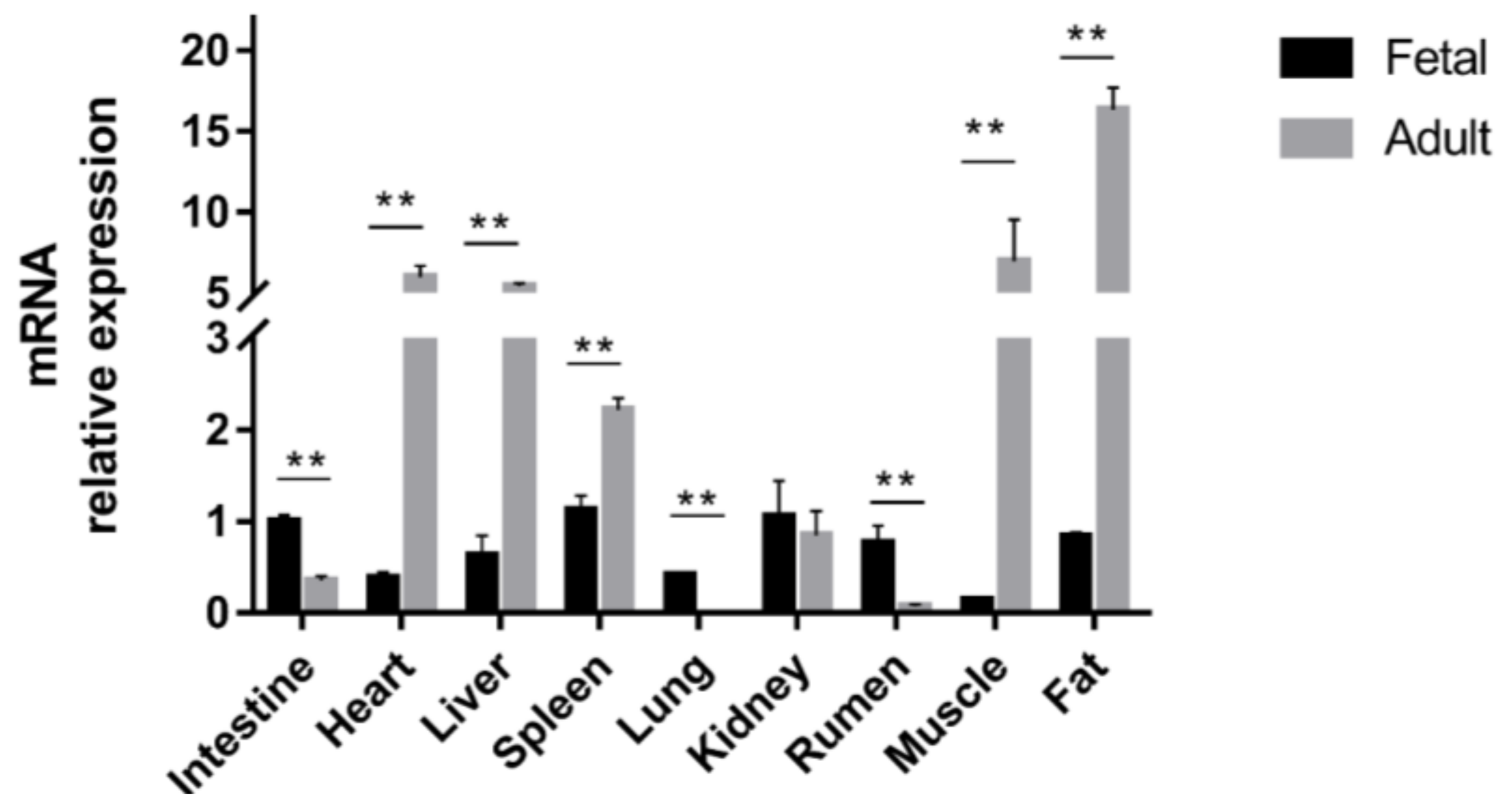
2.2. Differential Expression of STAT3 across Development
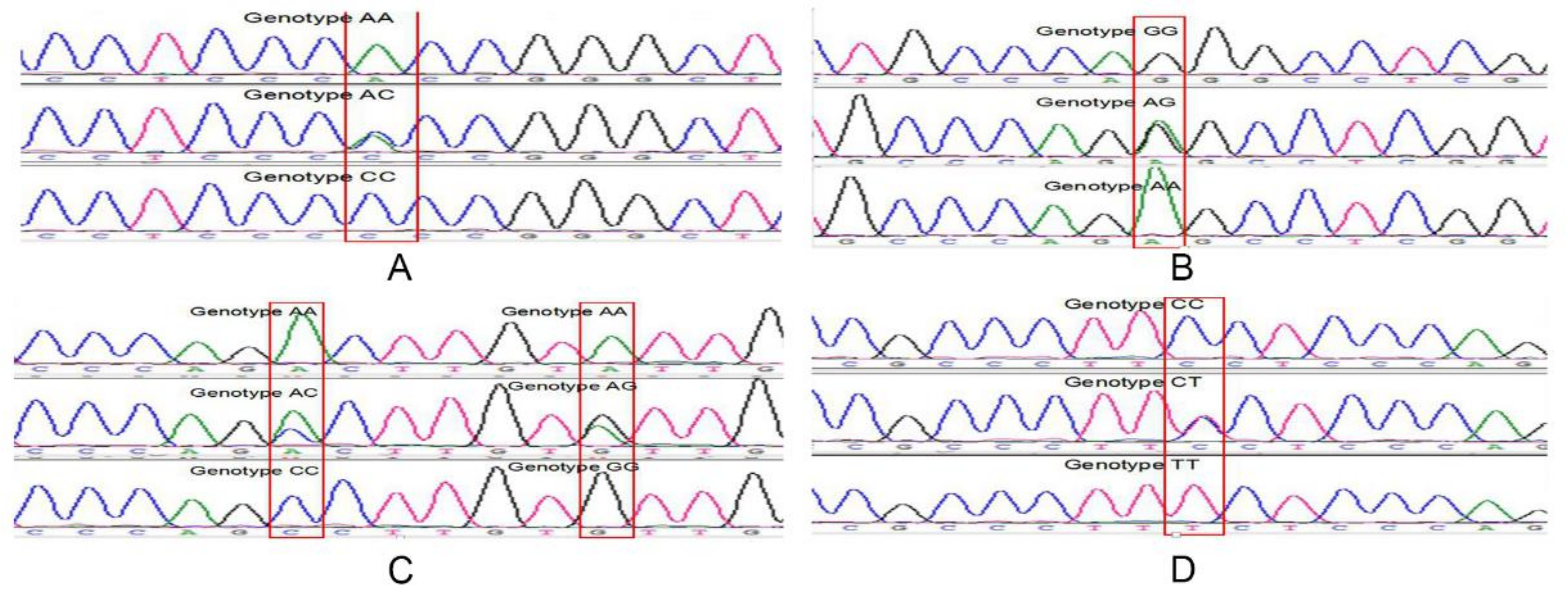
2.3. Identification of Sequence Variants and Prediction of Transcription Factors in Qinchuan Cattle
2.4. Linkage Disequilibrum (LD) and Haplotype Analysis
2.5. Effects of SNPs and Haplotype Combinations on Body Size Traits
3. Discussion
4. Materials and Methods
4.1. Bioinformatics Analyses
4.2. Subject Animals
4.3. RNA Preparation and Real-Time PCR
4.4. DNA Isolation, and Phenotypic Data
4.5. PCR Amplification and Genotyping
4.6. Data Analyses
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CDD | Conserved Domains Database |
| FQC | fetal Qinchuan cattle |
| AQC | adult Qinchuan cattle |
| LD | linkage disequilibrium |
| MAS | marker-assisted selection |
| QTL | quantitative trait locus |
| HWE | Hardy-Weinberg equilibrium |
References
- Zuo, B.; Yang, H.; Lei, M.; Li, F.; Deng, C.; Jiang, S.; Xiong, Y. Association of the polymorphism in GYS1 and ACOX1 genes with meat quality traits in pigs. Animal 2007. [Google Scholar] [CrossRef] [PubMed]
- Brito, L.F.; Clarke, S.M.; McEwan, J.C.; Miller, S.P.; Pickering, N.K.; Bain, W.E.; Dodds, K.G.; Sargolzaei, M.; Schenkel, F.S. Prediction of genomic breeding values for growth, carcass and meat quality traits in a multi-breed sheep population using a HD SNP chip. BMC Genet. 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Gui, L.S.; Hao, R.J.; Zhang, Y.R.; Zhao, X.L.; Zan, L.S. Haplotype distribution in the class I sirtuin genes and their associations with ultrasound carcass traits in Qinchuan cattle (Bos taurus). Mol. Cell. Probes 2015, 29, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.C.; Wang, Y.N.; Fu, C.Z.; Zan, L.S. Association study and expression analysis of MTNR1A as a candidate gene for body measurement and meat quality traits in Qinchuan cattle. Gene 2015, 570, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhang, S.; Wei, S.; Zhang, Y.; Li, Y.; Fu, C.; Zhao, C.; Zan, L. Novel polymorphisms of SIX4 gene and their association with body measurement traits in Qinchuan cattle. Gene 2014, 539, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Fontanesi, L.; Scotti, E.; Buttazzoni, L.; Dall’Olio, S.; Bagnato, A.; Fiego, D.P.L.; Davoli, R.; Russo, V. Confirmed association between a single nucleotide polymorphism in the FTO gene and obesity-related traits in heavy pigs. Mol. Biol. Rep. 2010, 37, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Alvarado, J.J.; Betts, L.; Moroco, J.A.; Smithgall, T.E.; Yeh, J.I. Crystal Structure of the Src Family Kinase Hck SH3-SH2 Linker Regulatory Region Supports an SH3-dominant Activation Mechanism. J. Biol. Chem. 2010, 285, 35455–35461. [Google Scholar] [CrossRef] [PubMed]
- Kamran, M.Z.; Patil, P.; Gude, R.P. Role of STAT3 in Cancer Metastasis and Translational Advances. BioMed Res. Int. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Cui, G.; Sun, L.; Wang, S.J.; Li, Y.L.; Meng, Y.G.; Guan, Z.; Fan, W.S.; Li, L.A.; Yang, Y.Z.; et al. STAT3 acetylation-induced promoter methylation is associated with downregulation of the ARHI tumor-suppressor gene in ovarian cancer. Oncol. Rep. 2013, 30, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, S.R.; Fan, M.Y.; Du, Z.Y.; Yang, C.H.; Pfeffer, L.M. Unphosphorylated STAT3 regulates the antiproliferative, antiviral, and gene-inducing actions of type I interferons. Biochem. Biophys. Res. Commun. 2017, 490, 739–745. [Google Scholar] [CrossRef] [PubMed]
- Limagne, E.; Thibaudin, M.; Euvrard, R.; Berger, H.; Chalons, P.; Vegan, F.; Humblin, E.; Boidot, R.; Rebe, C.; Derangere, V.; et al. Sirtuin-1 Activation Controls Tumor Growth by Impeding Th17 Differentiation via STAT3 Deacetylation. Cell Rep. 2017, 19, 746–759. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.; Xie, Y.X.; Gou, Q.H.; Guo, X.Q.; Yao, Q.; Gou, X.J. JAK/STAT3 and Smad3 activities are required for the wound healing properties of Periplaneta americana extracts. Int. J. Mol. Med. 2017, 40, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Vaisse, C.; Halaas, J.L.; Horvath, C.M.; Darnell, J.E.; Stoffel, M.; Friedman, J.M. Leptin activation of Stat3 in the hypothalamus of wild–type and ob/ob mice but not db/db mice. Nat. Genet. 1996, 14, 95–97. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.X.; Huang, L.; Elefteriou, F.; Yang, G.Q.; Shelton, J.M.; Giles, J.E.; Oz, O.K.; Pourbahrami, T.; Lu, C.Y.H.; Richardson, J.A.; et al. Essential role of STAT3 in body weight and glucose homeostasis. Mol. Cell. Biol. 2004, 24, 258–269. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Wolfgang, M.J.; Neschen, S.; Morino, K.; Horvath, T.L.; Shulman, G.I.; Fu, X.Y. Disruption of neural signal transducer and activator of transcription 3 causes obesity, diabetes, infertility, and thermal dysregulation. Proc. Natl. Acad. Sci. USA 2004, 101, 4661–4666. [Google Scholar] [CrossRef] [PubMed]
- Gorogawa, S.; Fujitani, Y.; Kaneto, H.; Hazama, Y.; Watada, H.; Miyamoto, Y.; Takeda, K.; Akira, S.; Magnuson, M.A.; Yamasaki, Y.; et al. Insulin secretory defects and impaired islet architecture in pancreatic beta-cell-specific STAT3 knockout mice. Biochem. Biophys. Res. Commun. 2004, 319, 1159–1170. [Google Scholar] [CrossRef] [PubMed]
- Derecka, M.; Gornicka, A.; Koralov, S.B.; Szczepanek, K.; Morgan, M.; Raje, V.; Sisler, J.; Zhang, Q.F.; Otero, D.; Cichy, J.; et al. Tyk2 and Stat3 Regulate Brown Adipose Tissue Differentiation and Obesity. Cell Metab. 2012, 16, 814–824. [Google Scholar] [CrossRef] [PubMed]
- Song, N.; Gui, L.S.; Xu, H.C.; Wu, S.; Zan, L.S. Identification of single nucleotide polymorphisms of the signal transducer and activator of transcription 3 gene (STAT3) associated with body measurement and carcass quality traits in beef cattle. Genet. Mol. Res. GMR 2015, 14, 11242–11249. [Google Scholar] [CrossRef] [PubMed]
- Pang, Y.H.; Wang, J.Q.; Zhang, C.L.; Lei, C.Z.; Lan, X.Y.; Yue, W.P.; Gu, C.W.; Chen, D.X.; Chen, H. The polymorphisms of bovine VEGF gene and their associations with growth traits in Chinese cattle. Mol. Biol. Rep. 2011, 38, 755–759. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zan, L.; Zhao, S.; Xin, Y.; Li, L.; Cui, W.; Tang, Z.; Li, K. Molecular characterization, polymorphism of bovine ZBTB38 gene and association with body measurement traits in native Chinese cattle breeds. Mol. Biol. Rep. 2010, 37, 4041–4049. [Google Scholar] [CrossRef] [PubMed]
- Dekkers, J.C. Commercial application of marker-and gene-assisted selection in livestock: Strategies and lessons. J. Anim. Sci. 2004, 82 (Suppl. 13), E313–E328. [Google Scholar] [PubMed]
- de Camargo, G.M.F.; Cardoso, D.F.; Gil, F.M.M.; Fonseca, P.D.D.; Zetouni, L.; Braz, C.U.; de Freitas, A.C.; de Souza, F.R.P.; Aspilcueta-Borquis, R.R.; Baldi, F.; et al. First polymorphisms in JY-1 gene in cattle (Bos taurus indicus) and their association with sexual precocity and growth traits. Mol. Biol. Rep. 2012, 39, 10105–10109. [Google Scholar] [CrossRef] [PubMed]
- Alim, M.A.; Fan, Y.P.; Wu, X.P.; Xie, Y.; Zhang, Y.; Zhang, S.L.; Sun, D.X.; Zhang, Y.; Zhang, Q.; Liu, L.; et al. Genetic effects of stearoyl-coenzyme A desaturase (SCD) polymorphism on milk production traits in the Chinese dairy population. Mol. Biol. Rep. 2012, 39, 8733–8740. [Google Scholar] [CrossRef] [PubMed]
- Jamshidi, Y.; Kyriakou, T.; Gooljar, S.B.; Collins, L.J.; Lane, C.A.; Snieder, H.; Wang, X.L.; Spector, T.D.; O’Dell, S.D. Common STAT3 variants are not associated with obesity or insulin resistance in female twins. Obesity 2007, 15, 1634–1639. [Google Scholar] [CrossRef] [PubMed]
- Gianotti, T.F.; Sookoian, S.; Gemma, C.; Burgueno, A.L.; Gonzalez, C.D.; Pirola, C.J. Study of genetic variation in the STAT3 on obesity and insulin resistance in male adults. Obesity 2008, 16, 1702–1707. [Google Scholar] [CrossRef] [PubMed]
- Phillips, C.M.; Goumidi, L.; Bertrais, S.; Field, M.R.; Peloso, G.M.; Shen, J.; McManus, R.; Hercberg, S.; Lairon, D.; Planells, R.; et al. Dietary Saturated Fat Modulates the Association between STAT3 Polymorphisms and Abdominal Obesity in Adults. J. Nutr. 2009, 139, 2011–2017. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Tiwari, H.K.; Lin, W.Y.; Allison, D.B.; Chung, W.K.; Leibel, R.L.; Yi, N.; Liu, N. Genetic association analysis of 30 genes related to obesity in a European American population. Int. J. Obes. 2014, 38, 724–729. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liu, X.; Wang, D.; Ning, C.; Wang, H.F.; Zhang, Q.; Jiang, L. Functional validation of GPIHBP1 and identification of a functional mutation in GPIHBP1 for milk fat traits in dairy cattle. Sci. Rep. 2017, 7, 10. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.L.; Yin, H.W.; Li, C.; Qin, C.H.; Cai, W.T.; Cao, M.Y.; Zhang, S.L. Genetic effects of PDGFRB and MARCH1 identified in GWAS revealing strong associations with semen production traits in Chinese Holstein bulls. BMC Genet. 2017, 18, 10. [Google Scholar] [CrossRef] [PubMed]
- Gui, L.S.; Hong, J.Y.; Raza, S.H.A.; Zan, L.S. Genetic variants in SIRT3 transcriptional regulatory region affect promoter activity and fat deposition in three cattle breeds. Mol. Cell. Probes 2017, 32, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.J.; Liu, Y.F.; Xu, Q.; Liu, C.K.; Wang, J.; Ding, C.; Fang, M.Y. A post-GWAS confirming GPAT3 gene associated with pig growth and a significant SNP influencing its promoter activity. Anim. Genet. 2017, 48, 478–482. [Google Scholar] [CrossRef] [PubMed]
- Marty, A.; Amigues, Y.; Servin, B.; Renand, G.; Leveziel, H.; Rocha, D. Genetic Variability and Linkage Disequilibrium Patterns in the Bovine DNAJA1 Gene. Mol. Biotechnol. 2010, 44, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Nettleton, D.; Dekkers, J.C.M. Evaluation of linkage disequilibrium measures between multi-allelic markers as predictors of linkage disequilibrium between single nucleotide polymorphisms. Genet. Res. 2007, 89, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Zhang, R.J.; Jiang, Y.S.; Wang, X.; Li, J.; Lv, H.C.; Tang, G.P.; Guo, X.D.; Meng, X.W.; Zhang, H.K.; et al. Assessing the patterns of linkage disequilibrium in genic regions of the human genome. FEBS J. 2011, 278, 3748–3755. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.C.; Cha, M.H.; Kim, D.M.; Lee, H.; Moon, J.S.; Choi, S.M.; Kim, K.S.; Yoon, Y. A functional promoter polymorphism−607 G > C of WNT10B is associated with abdominal fat in Korean female subjects. J. Nutr. Biochem. 2011, 22, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.Z.; He, H.; Sun, J.J.; Wang, J.; Li, Z.J.; Lan, X.Y.; Lei, C.Z.; Zhang, C.L.; Zhang, E.P.; Wang, J.Q.; et al. Haplotype combination of SREBP-1c gene sequence variants is associated with growth traits in cattle. Genome Natl. Res. Counc. Can. 2011, 54, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Posada, D.; Crandall, K.A. Intraspecific gene genealogies: Trees grafting into networks. Trends Ecol. Evol. 2001, 16, 37–45. [Google Scholar] [CrossRef]
- Bates, S.H.; Stearns, W.H.; Dundon, T.A.; Schubert, M.; Tso, A.W.; Wang, Y.; Banks, A.S.; Lavery, H.J.; Haq, A.K.; Maratos-Flier, E. STAT3 signalling is required for leptin regulation of energy balance but not reproduction. Nature 2003, 421, 856–859. [Google Scholar] [CrossRef] [PubMed]
- Cernkovich, E.R.; Deng, J.; Bond, M.C.; Combs, T.P.; Harp, J.B. Adipose-specific disruption of signal transducer and activator of transcription 3 increases body weight and adiposity. Endocrinology 2008, 149, 1581–1590. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.C.; Wu, X.F.; Li, X.C.; Xia, T.; Lei, C.Z.; Chen, H.; Pan, C.Y.; Lan, X.Y. Novel genetic variants associated with mRNA expression of signal transducer and activator of transcription 3(STAT3) gene significantly affected goat growth traits. Small Rumin. Res. 2015, 129, 25–36. [Google Scholar] [CrossRef]
- Pan, C.; Wu, C.; Jia, W.; Xu, Y.; Lei, C.; Hu, S.; Lan, X.; Chen, H. A critical functional missense mutation (H173R) in the bovine PROP1 gene significantly affects growth traits in cattle. Gene 2013, 531, 398–402. [Google Scholar] [CrossRef] [PubMed]
- Garieri, M.; Delaneau, O.; Santoni, F.; Fish, R.J.; Mull, D.; Carninci, P.; Dermitzakis, E.T.; Antonarakis, S.E.; Fort, A. The effect of genetic variation on promoter usage and enhancer activity. Nat. Commun. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Dall’Olio, S.; Wang, Y.; Sartori, C.; Fontanesi, L.; Mantovani, R. Association of myostatin (MSTN) gene polymorphisms with morphological traits in the Italian Heavy Draft Horse breed. Livest. Sci. 2014, 160, 29–36. [Google Scholar] [CrossRef]
- Chou, J.L.; Wei, K.L.; Shie, C.C.; Wu, C.S.; Chan, M.W.Y. Infection of H. pylori is associated with promoter methylation of a putative STAT3-targeted microRNA, miR-193a, in gastric cancer. J. Gastroenterol. Hepatol. 2015, 30, 86. [Google Scholar]
- Ahluwalia, A.; Busse, B.A.; Thiruvengadam, S.S.; Tarnawski, A.S. Importins Are Critical for Colorectal Cancer (CRC) Growth and Are Novel Biomarkers of CRC. Underlying Mechanisms Include: Increased Nuclear Transport of P-CREB and P-STAT3, VEGF Gene Promoter Activation and Aberrant VEGF Expression. Gastroenterology 2011, 140, S184. [Google Scholar]
- Lindemann, C.; Hackmann, O.; Delic, S.; Schmidt, N.; Reifenberger, G.; Riemenschneider, M.J. SOCS3 promoter methylation is mutually exclusive to EGFR amplification in gliomas and promotes glioma cell invasion through STAT3 and FAK activation. Acta Neuropathol. 2011, 122, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Farren, M.R.; Spangler, H.; Carlson, L.M.; Lee, K.P. STAT3 Activity Negatively Regulates PKC beta Promoter Activity, Impairing Dendritic Cell Differentiation. Blood 2011, 118, 89. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Marchlerbauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2017, 45, D200. [Google Scholar] [CrossRef] [PubMed]
- Marchlerbauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I. CDD: NCBI’s conserved domain database. Nucleic Acids Res. 2015, 43, D222. [Google Scholar] [CrossRef] [PubMed]
- Richardson, C.; Jones, P.C.; Barnard, V.; Hebert, C.N.; Terlecki, S.; Wijeratne, W.V. Estimation of the developmental age of the bovine fetus and newborn calf. Vet. Rec. 1990, 126, 279–284. [Google Scholar] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Denman, A. Molecular cloning: A laboratory manual. Immunology 1983, 49, 411. [Google Scholar]
- Gilbert, R.; Bailey, D.; Shannon, N. Linear body measurements of cattle before and after 20 years of selection for postweaning gain when fed two different diets. J. Anim. Sci. 1993, 71, 1712–1720. [Google Scholar] [CrossRef] [PubMed]
- Yeh, F.; Yang, R.-C.; Boyle, T. POPGENE Version1. 32, Microsoft Window-Base Software for Population Genetic Analysis: A Quick User’s Guide; University of Alberta, Center for International Forestry Research: Edmonton, AB, Canada, 1999. [Google Scholar]
- Nei, M.; Roychoudhury, A. Sampling variances of heterozygosity and genetic distance. Genetics 1974, 76, 379–390. [Google Scholar] [PubMed]
- Gui, L.S.; Zhang, Y.R.; Liu, G.Y.; Zan, L.S. Expression of the SIRT2 Gene and Its Relationship with Body Size Traits in Qinchuan Cattle (Bos taurus). Int. J. Mol. Sci. 2015, 16, 2458–2471. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.R.; Gui, L.S.; Li, Y.K.; Jiang, B.J.; Wang, H.C.; Zhang, Y.Y.; Zan, L.S. Molecular Characterization of Bovine SMO Gene and Effects of Its Genetic Variations on Body Size Traits in Qinchuan Cattle (Bos taurus). Int. J. Mol. Sci. 2015, 16, 16966–16980. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; He, L. SHEsis, a powerful software platform for analyses of linkage disequilibrium, haplotype construction, and genetic association at polymorphism loci. Am. J. Med. Genet. B 2005, 138b, 143. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Loci | Genotype Frequency | Allele Frequency | HWE | PIC | He | Ne | |||
|---|---|---|---|---|---|---|---|---|---|
| SNP1 | AA | AC | CC | A | C | ||||
| 0.2405 | 0.1833 | 0.5762 | 0.3321 | 0.6679 | 144.6004 | 0.3452 | 0.4436 | 1.7974 | |
| SNP2 | AA | AG | GG | A | G | ||||
| 0.2857 | 0.1429 | 0.5714 | 0.3571 | 0.6429 | 199.3185 | 0.3538 | 0.4592 | 1.8491 | |
| SNP3/4 | AA/GG | AC/AG | CC/AA | A/G | C/A | ||||
| 0.2143 | 0.5952 | 0.1905 | 0.5119 | 0.4881 | 15.3463 | 0.3749 | 0.4997 | 1.9989 | |
| SNP5 | CC | CT | TT | C | T | ||||
| 0.5238 | 0.3095 | 0.1667 | 0.6786 | 0.3214 | 35.4313 | 0.3411 | 0.4362 | 1.7738 | |
| SNPs | Variation | Predicted Transcription Factor Binding Site | |||
|---|---|---|---|---|---|
| Gain a | Loss b | Transcription Factor Binding Sequence c | Score | ||
| SNP1 | −304A>C | - | Sp1 | CCCCC | 100 |
| SNP2 | −285G>A | AP-2 | - | CCCAGGG | 100 |
| SNP3 | −209A>C | - | MyoD | CAGCC | 100 |
| SNP4 | −203A>G | CP1 | - | ATTGC | 100 |
| SNP5 | −188T>C | - | SRF | TTCCTC | 100 |
| SNP | QC | |
|---|---|---|
| r2 | D′ | |
| SNP1-SNP2 | 0.211 | 0.486 |
| SNP1-SNP3 | 0.072 | 0.391 |
| SNP1-SNP4 | 0.060 | 0.340 |
| SNP1-SNP5 | 0.052 | 0.235 |
| SNP2-SNP3 | 0.149 | 0.530 |
| SNP2-SNP4 | 0.091 | 0.369 |
| SNP2-SNP4 | 0.276 | 0.569 |
| SNP3-SNP4 | 0.452 | 1.000 |
| SNP4-SNP5 | 0.332 | 0.817 |
| Haplotypes | SNP1 | SNP2 | SNP3 | SNP4 | SNP5 | Frequencies |
|---|---|---|---|---|---|---|
| Hap1 | A | A | A | A | T | 0.151 |
| Hap2 | C | G | A | A | C | 0.112 |
| Hap3 | C | G | C | G | C | 0.332 |
| Loci | Genotypes | Body Size Traits (Mean ± SD) | |||||
|---|---|---|---|---|---|---|---|
| Body Length (cm) | Wither Height (cm) | Hip Height (cm) | Hip Width (cm) | Chest Depth (cm) | Chest Circumference (cm) | ||
| SNP1 | AA | 127.683 ± 1.051 | 117.990 ± 0.623 | 119.119 ± 0.434 | 34.297 ± 0.617 | 59.446 ± 0.729 a | 155.574 ± 1.775 |
| AC | 127.753 ± 1.203 | 117.325 ± 0.714 | 119.636 ± 0.497 | 34.260 ± 0.707 | 57.961 ± 0.835 | 152.740 ± 2.033 | |
| CC | 127.839 ± .679 | 116.855 ± 0.403 | 119.326 ± 0.280 | 34.083 ± 0.399 | 57.430 ± 0.471 b | 154.694 ± 1.147 | |
| SNP2 | AA | 127.875 ± 0.969 | 118.567 ± 0.564 A | 119.667 ± 0.419 | 34.833 ± 0.566 | 58.167 ± 0.672 | 155.750 ± 1.641 |
| AG | 128.667 ± 1.370 | 118.683 ± 0.798 A | 119.583 ± 0.593 | 33.333 ± 0.801 | 59.333 ± 0.951 | 154.833 ± 2.321 | |
| GG | 127.146 ± 0.685 | 115.833 ± 0.399 B | 118.500 ± 0.297 | 33.917 ± 0.400 | 57.479 ± 0.475 | 153.583 ± 1.161 | |
| SNP3/4 | AA/AA | 129.056 ± 1.100 | 119.278 ± 0.656 A,a | 120.000 ± 0.482 | 34.778 ± 0.645 | 60.222 ± 0.765 A | 157.556 ± 1.876 a |
| AC/AG | 125.984 ± 0.661 | 116.279 ± 0.394 B | 118.681 ± 0.290 | 33.209 ± 0.388 B | 56.787 ± 0.460 B | 152.185 ± 1.128 b | |
| CC/GG | 130.938 ± 1.167 | 116.938 ± 0.696 b | 118.938 ± 0.511 | 36.125 ± 0.684 A | 59.000 ± 0.811 | 157.875 ± 1.990 a | |
| SNP5 | TT | 128.143 ± 1.269 | 118.071 ± 0.748 a | 119.962 ± 0.400 A | 35.273 ± 0.411 A | 59.714 ± 0.873 a | 156.000 ± 1.575 |
| TC | 127.885 ± 0.931 | 118.038 ± 0.549 | 119.500 ± 0.545 | 33.429 ± 0.728 | 58.692 ± 0.641 | 155.143 ± 2.147 | |
| CC | 127.205 ± 0.716 | 116.114 ± 0.422 b | 118.250 ± 0.307 B | 32.462 ± 0.534 B | 56.932 ± 0.493 b | 153.182 ± 1.211 | |
| Haplotype Combination | Body Size Traits (Mean ± SD) | ||||||
|---|---|---|---|---|---|---|---|
| Frequency | Body Length | Wither Height | Hip Height (cm) | Hip Width (cm) | Chest Circumference (cm) | Chest Depth (cm) | |
| H1H1 (AA-AA-AA-AA-TT) | 12% (50) | 125.600 ± 1.128 B | 116.700 ± 0.711 A | 118.500 ± 0.596 a | 32.600 ± 0.778 B | 59.200 ± 0.998 A,b | 152.600 ± 2.243 B |
| H2H3 (CC-GG-AC-GA-CC) | 19% (80) | 124.125 ± 0.892 B | 113.750 ± 0.562 B | 116.563 ± 0.471 B,b | 33.625 ± 0.615 B | 55.688 ± 0.789 B | 146.750 ± 1.773 B |
| H3H3 (CC-GG-CC-GG-CC) | 9% (40) | 137.625 ± 1.262 A | 118.625 ± 0.795 A | 120.125 ± 0.667 A | 38.250 ± 0.870 A | 63.250 ± 1.115 A,a | 167.500 ± 2.507 A |
| Item | Function | Primer Sequences | Tm (◦C) | Production Size |
|---|---|---|---|---|
| STAT3 | RT-PCR | F: 5-CACTTCTGCCAAGGGAGACT-3 | 59.5 | 261 bp |
| R: 5-CGCGTATGCCCAATCTTGAC-3 | ||||
| β-actin | Internal control | F: 5-CACCAACTGGGACGACAT-3 | 61 | 202 bp |
| R: 5-ATACAGGGACAGCACAGC-3 | ||||
| GAPDH | Internal control | F: 5-CCAACGTGTCTGTTGTGGAT-3 | 61 | 80 bp |
| R: 5-CTGCTTCACCACCTTCTTGA-3 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, S.; Wang, Y.; Ning, Y.; Guo, H.; Wang, X.; Zhang, L.; Khan, R.; Cheng, G.; Wang, H.; Zan, L. Genetic Variants in STAT3 Promoter Regions and Their Application in Molecular Breeding for Body Size Traits in Qinchuan Cattle. Int. J. Mol. Sci. 2018, 19, 1035. https://doi.org/10.3390/ijms19041035
Wu S, Wang Y, Ning Y, Guo H, Wang X, Zhang L, Khan R, Cheng G, Wang H, Zan L. Genetic Variants in STAT3 Promoter Regions and Their Application in Molecular Breeding for Body Size Traits in Qinchuan Cattle. International Journal of Molecular Sciences. 2018; 19(4):1035. https://doi.org/10.3390/ijms19041035
Chicago/Turabian StyleWu, Sen, Yaning Wang, Yue Ning, Hongfang Guo, Xiaoyu Wang, Le Zhang, Rajwali Khan, Gong Cheng, Hongbao Wang, and Linsen Zan. 2018. "Genetic Variants in STAT3 Promoter Regions and Their Application in Molecular Breeding for Body Size Traits in Qinchuan Cattle" International Journal of Molecular Sciences 19, no. 4: 1035. https://doi.org/10.3390/ijms19041035
APA StyleWu, S., Wang, Y., Ning, Y., Guo, H., Wang, X., Zhang, L., Khan, R., Cheng, G., Wang, H., & Zan, L. (2018). Genetic Variants in STAT3 Promoter Regions and Their Application in Molecular Breeding for Body Size Traits in Qinchuan Cattle. International Journal of Molecular Sciences, 19(4), 1035. https://doi.org/10.3390/ijms19041035