miR-211 Plays a Critical Role in Cnidium officinale Makino Extract-Induced, ROS/ER Stress-Mediated Apoptosis in U937 and U266 Cells



 ,
, 
Abstract
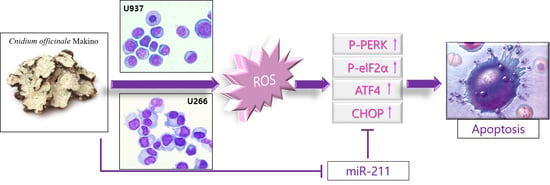
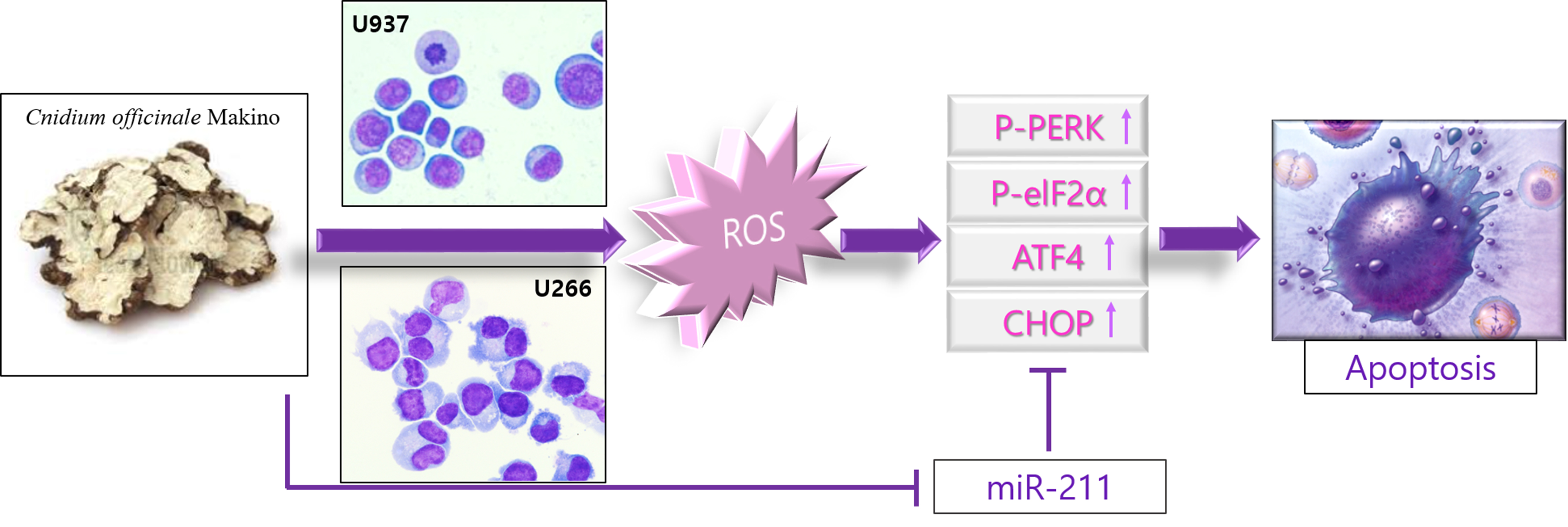{kind=link}
{kind=link}
{kind=link}
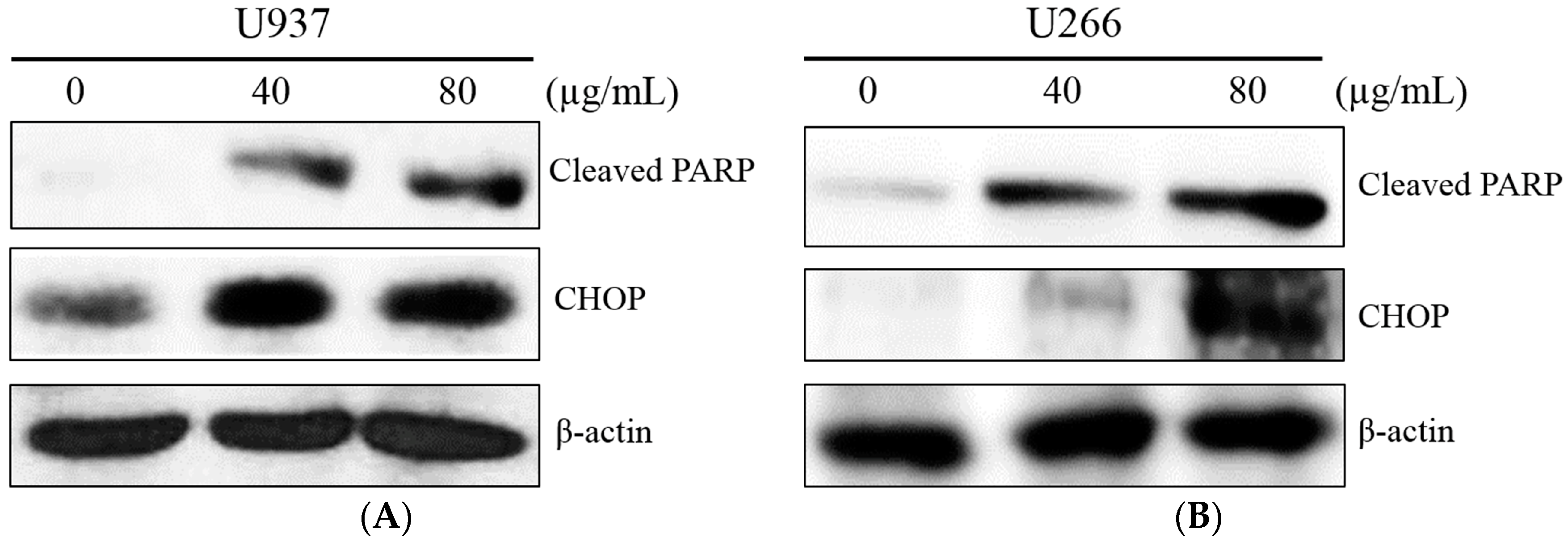{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
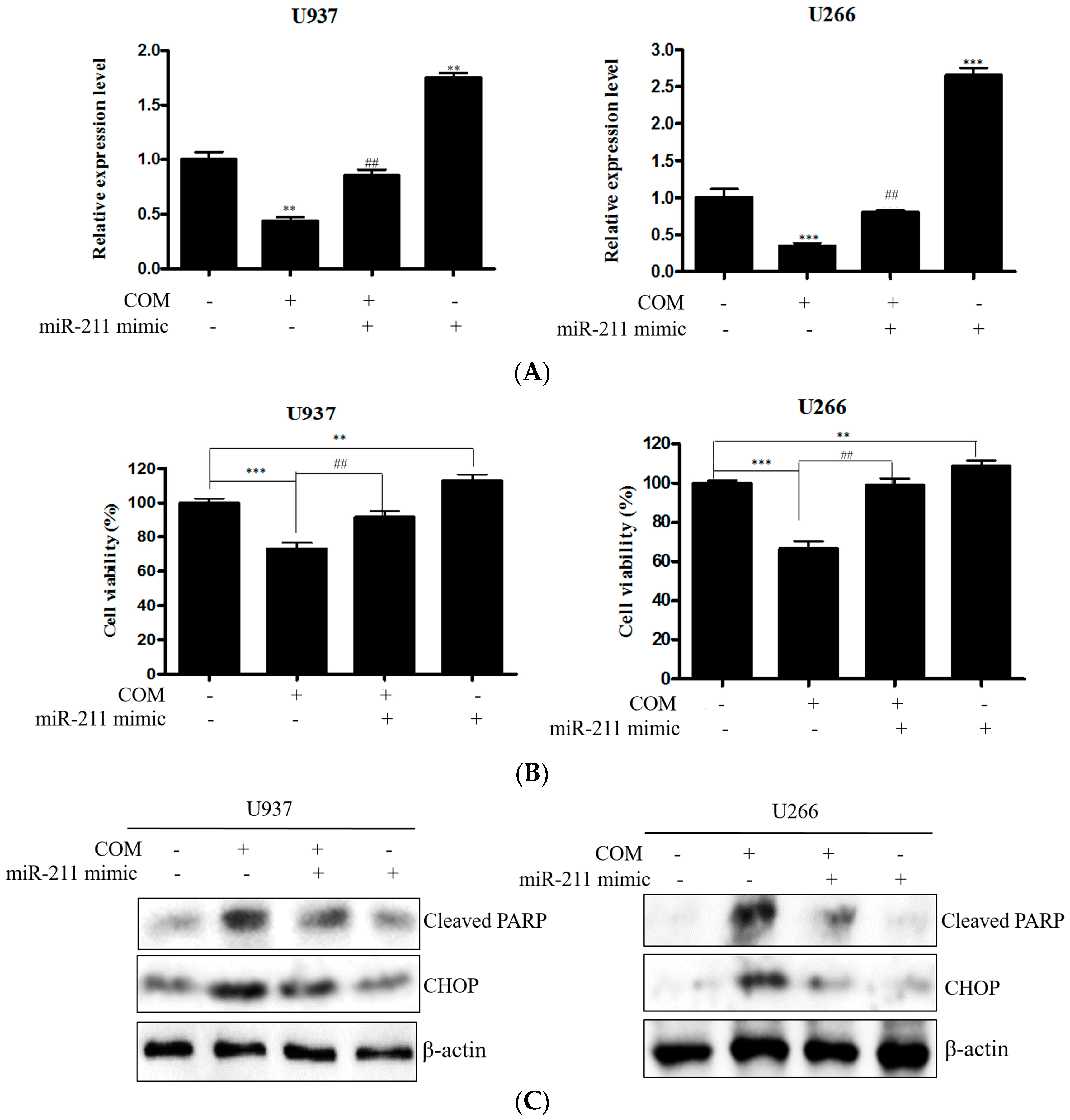{kind=link}
1. Introduction
2. Results
2.1. COM Exerted Cytotoxicity in U937 and U266 Cells
2.2. COM Treatment Increased ER Stress-Related Proteins in U937 and U266 Cells
2.3. COM Treatment Induced Apoptosis in U937 and U266 Cells
2.4. COM Treatment Induced Apoptosis via ROS Generation in U937 and U266 Cells
2.5. COM Downregulated the Expression of miR-211 in U937 and U266 Cells
2.6. miR-211 Plays a Critical Role in COM-Induced Apoptosis in U937 and U266 Cells
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Culture
4.3. Cytotoxicity Assay
4.4. Western Blot Analysis
4.5. Measurement of ROS Generation
4.6. qRT-PCR
4.7. miRNA Mimic Transfection
4.8. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Agarwal, A.; Ghobrial, I.M. Monoclonal gammopathy of undetermined significance and smoldering multiple myeloma: A review of the current understanding of epidemiology, biology, risk stratification, and management of myeloma precursor disease. Clin. Cancer Res. 2012, 19, 985–994. [Google Scholar] [CrossRef] [PubMed]
- Barlogie, B.; Shaughnessy, J.; Tricot, G.; Jacobson, J.; Zangari, M.; Anaissie, E.; Walker, R.; Crowley, J. Treatment of multiple myeloma. Blood 2004, 103, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.; Xu, W.; Reed, J.C. Cell death and endoplasmic reticulum stress: Disease relevance and therapeutic opportunities. Nat. Rev. Drug Discov. 2008, 7, 1013–1030. [Google Scholar] [CrossRef] [PubMed]
- Hetz, C. The unfolded protein response: Controlling cell fate decisions under ER stress and beyond. Nat. Rev. Mol. Cell Biol. 2012, 13, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Schröder, M.; Kaufman, R.J. ER stress and the unfolded protein response. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2005, 569, 29–63. [Google Scholar] [CrossRef] [PubMed]
- Ron, D.; Walter, P. Signal integration in the endoplasmic reticulum unfolded protein response. Nat. Rev. Mol. Cell Biol. 2007, 8, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Sano, R.; Reed, J.C. ER stress-induced cell death mechanisms. BBA Mol. Cell Res. 2013, 1833, 3460–3470. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Bailly-Maitre, B.; Reed, J.C. Endoplasmic reticulum stress: Cell life and death decisions. J. Clin. Investig. 2005, 115, 2656–2664. [Google Scholar] [CrossRef] [PubMed]
- Ri, M.; Tashiro, E.; Oikawa, D.; Shinjo, S.; Tokuda, M.; Yokouchi, Y.; Narita, T.; Masaki, A.; Ito, A.; Ding, J.; et al. Identification of Toyocamycin, an agent cytotoxic for multiple myeloma cells, as a potent inhibitor of ER stress-induced XBP1 mRNA splicing. Blood Cancer J. 2012, 2, e79. [Google Scholar] [CrossRef] [PubMed]
- Obeng, E.A.; Carlson, L.M.; Gutman, D.M.; Harrington, W.J.; Lee, K.P.; Boise, L.H. Proteasome inhibitors induce a terminal unfolded protein response in multiple myeloma cells. Blood 2006, 107, 4907–4916. [Google Scholar] [CrossRef] [PubMed]
- Verfaillie, T.; Rubio, N.; Garg, A.D.; Bultynck, G.; Rizzuto, R.; Decuypere, J.P.; Piette, J.; Linehan, C.; Gupta, S.; Samali, A.; et al. PERK is required at the ER-mitochondrial contact sites to convey apoptosis after ROS-based ER stress. Cell Death Differ. 2012, 19, 1880–1891. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Srivastava, S.K.; Kim, S.H. Caspase-9 as a therapeutic target for treating cancer. Expert Opin. Ther. Targets 2015, 19, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Chitnis, N.S.; Pytel, D.; Bobrovnikova-Marjon, E.; Pant, D.; Zheng, H.; Maas, N.L.; Frederick, B.; Kushner, J.A.; Chodosh, L.A.; Koumenis, C.; et al. miR-211 is a prosurvival microRNA that regulates chop expression in a PERK-dependent manner. Mol. Cell 2012, 48, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Song, H.-S.; Park, H.; Kim, B. Activation of ER stress dependent-miR-216b has critical role in Salvia miltiorrhiza Ethanol Extract Induced Apoptosis in U266 and U937 cells. Int. J. Mol. Sci. 2018. under revision. [Google Scholar]
- Hong, H.; An, J.C.; de la Cruz, J.F.; Hwang, S.G. Cnidium officinale Makino extract induces apoptosis through activation of caspase-3 and p53 in human liver cancer HepG2 cells. Exp. Ther. Med. 2017, 14, 3191–3197. [Google Scholar] [CrossRef] [PubMed][Green Version]
- De la Cruz, J.; Kim, D.H.; Hwang, S.G. Anti cancer effects of Cnidium officinale Makino extract mediated through apoptosis and cell cycle arrest in the HT-29 human colorectal cancer cell line. Asian Pac. J. Cancer Prev. APJCP 2014, 15, 5117–5121. [Google Scholar] [PubMed]
- Lee, K.E.; Shin, J.A.; Hong, I.S.; Cho, N.P.; Cho, S.D. Effect of methanol extracts of Cnidium officinale Makino and Capsella bursa-pastoris on the apoptosis of HSC-2 human oral cancer cells. Exp. Ther. Med. 2013, 5, 789–792. [Google Scholar] [CrossRef] [PubMed]
- Joo, H.; Lee, H.J.; Shin, E.A.; Kim, H.; Seo, K.H.; Baek, N.I.; Kim, B.; Kim, S.H. c-Jun N-terminal Kinase-Dependent Endoplasmic Reticulum Stress Pathway is Critically Involved in Arjunic Acid Induced Apoptosis in Non-Small Cell Lung Cancer Cells. Phytother. Res. PTR 2016, 30, 596–603. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Shin, E.A.; Kim, C.G.; Lee, D.Y.; Kim, B.; Baek, N.I.; Kim, S.H. Obovatol Induces Apoptosis in Non-small Cell Lung Cancer Cells via C/EBP Homologous Protein Activation. Phytother. Res. PTR 2016, 30, 1841–1847. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Yun, M.; Kim, E.O.; Jung, D.B.; Won, G.; Kim, B.; Jung, J.H.; Kim, S.H. Decursin enhances TRAIL-induced apoptosis through oxidative stress mediated- endoplasmic reticulum stress signalling in non-small cell lung cancers. Br. J. Pharmacol. 2016, 173, 1033–1044. [Google Scholar] [CrossRef] [PubMed]
- Kyle, R.A.; Jacobus, S.; Friedenberg, W.R.; Slabber, C.F.; Rajkumar, S.V.; Greipp, P.R. The treatment of multiple myeloma using vincristine, carmustine, melphalan, cyclophosphamide, and prednisone (VBMCP) alternating with high-dose cyclophosphamide and α2β interferon versus VBMCP: Results of a phase III Eastern Cooperative Oncology Group Study E5A93. Cancer 2009, 115, 2155–2164. [Google Scholar] [PubMed]
- Facon, T.; Mary, J.Y.; Hulin, C.; Benboubker, L.; Attal, M.; Pegourie, B.; Renaud, M.; Harousseau, J.L.; Guillerm, G.; Chaleteix, C.; et al. Melphalan and prednisone plus thalidomide versus melphalan and prednisone alone or reduced-intensity autologous stem cell transplantation in elderly patients with multiple myeloma (IFM 99-06): A randomised trial. Lancet 2007, 370, 1209–1218. [Google Scholar] [CrossRef]
- Aghajanian, C.; Soignet, S.; Dizon, D.S.; Pien, C.S.; Adams, J.; Elliott, P.J.; Sabbatini, P.; Miller, V.; Hensley, M.L.; Pezzulli, S.; et al. A phase I trial of the novel proteasome inhibitor PS341 in advanced solid tumor malignancies. Clin. Cancer Res. 2002, 8, 2505–2511. [Google Scholar] [PubMed]
- Ailawadhi, S.; Frank, R.D.; Sharma, M.; Menghani, R.; Temkit, M.; Paulus, S.; Khera, N.; Hashmi, S.; Advani, P.; Swaika, A.; et al. Trends in multiple myeloma presentation, management, cost of care, and outcomes in the Medicare population: A comprehensive look at racial disparities. Cancer 2018. [Google Scholar] [CrossRef] [PubMed]
- Nomura, S.; Ito, T.; Yoshimura, H.; Hotta, M.; Nakanishi, T.; Fujita, S.; Nakaya, A.; Satake, A.; Ishii, K. Evaluation of thrombosis-related biomarkers before and after therapy in patients with multiple myeloma. J. Blood Med. 2018, 9, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.M.; Lee, Y.R.; Kim, C.S.; Jo, K.; Sohn, E.; Kim, J.S.; Kim, J. Cnidium officinale extract and butylidenephthalide inhibits retinal neovascularization in vitro and in vivo. BMC Complement. Altern. Med. 2016, 16, 231. [Google Scholar] [CrossRef] [PubMed]
- Bae, K.E.; Choi, Y.W.; Kim, S.T.; Kim, Y.K. Components of rhizome extract of Cnidium officinale Makino and their in vitro biological effects. Molecules 2011, 16, 8833–8847. [Google Scholar] [CrossRef] [PubMed]
- Kwak, D.H.; Kim, J.K.; Kim, J.Y.; Jeong, H.Y.; Keum, K.S.; Han, S.H.; Rho, Y.I.; Woo, W.H.; Jung, K.Y.; Choi, B.K.; et al. Anti-angiogenic activities of Cnidium officinale Makino and Tabanus bovinus. J. Ethnopharmacol. 2002, 81, 373–379. [Google Scholar] [CrossRef]
- Cao, S.S.; Kaufman, R.J. Endoplasmic Reticulum Stress and Oxidative Stress in Cell Fate Decision and Human Disease. Antioxid. Redox Signal. 2014, 21, 396–413. [Google Scholar] [CrossRef] [PubMed]
- Belmont, P.J.; Chen, W.J.; Thuerauf, D.J.; Glembotski, C.C. Regulation of microRNA expression in the heart by the ATF6 branch of the ER stress response. J. Mol. Cell. Cardiol. 2012, 52, 1176–1182. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Zhang, L.; Wang, F.; Wang, Y.; Huo, X.-S.; Yin, Y.-X.; Wang, Y.-Q.; Zhang, L.; Sun, S.-H. Modulation of the unfolded protein response is the core of microRNA-122-involved sensitivity to chemotherapy in hepatocellular carcinoma. Neoplasia 2011, 13. [Google Scholar] [CrossRef]
- Dai, R.; Li, J.; Liu, Y.; Yan, D.; Chen, S.; Duan, C.; Liu, X.; He, T.; Li, H. miR-221/222 suppression protects against endoplasmic reticulum stress-induced apoptosis via p27Kip1-and MEK/ERK-mediated cell cycle regulation. Biol. Chem. 2010, 391, 791–801. [Google Scholar] [CrossRef] [PubMed]








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cha, J.A.; Song, H.-S.; Kang, B.; Park, M.N.; Park, K.S.; Kim, S.-H.; Shim, B.-S.; Kim, B. miR-211 Plays a Critical Role in Cnidium officinale Makino Extract-Induced, ROS/ER Stress-Mediated Apoptosis in U937 and U266 Cells. Int. J. Mol. Sci. 2018, 19, 865. https://doi.org/10.3390/ijms19030865
Cha JA, Song H-S, Kang B, Park MN, Park KS, Kim S-H, Shim B-S, Kim B. miR-211 Plays a Critical Role in Cnidium officinale Makino Extract-Induced, ROS/ER Stress-Mediated Apoptosis in U937 and U266 Cells. International Journal of Molecular Sciences. 2018; 19(3):865. https://doi.org/10.3390/ijms19030865
Chicago/Turabian StyleCha, Jin Ah, Hyo-Sook Song, Beomku Kang, Moon Nyeo Park, Kyoung Sun Park, Sung-Hoon Kim, Bum-Sang Shim, and Bonglee Kim. 2018. "miR-211 Plays a Critical Role in Cnidium officinale Makino Extract-Induced, ROS/ER Stress-Mediated Apoptosis in U937 and U266 Cells" International Journal of Molecular Sciences 19, no. 3: 865. https://doi.org/10.3390/ijms19030865
APA StyleCha, J. A., Song, H.-S., Kang, B., Park, M. N., Park, K. S., Kim, S.-H., Shim, B.-S., & Kim, B. (2018). miR-211 Plays a Critical Role in Cnidium officinale Makino Extract-Induced, ROS/ER Stress-Mediated Apoptosis in U937 and U266 Cells. International Journal of Molecular Sciences, 19(3), 865. https://doi.org/10.3390/ijms19030865