The Xanthine Oxidase Inhibitor Febuxostat Suppresses the Progression of IgA Nephropathy, Possibly via Its Anti-Inflammatory and Anti-Fibrotic Effects in the gddY Mouse Model


Abstract
1. Introduction
2. Results
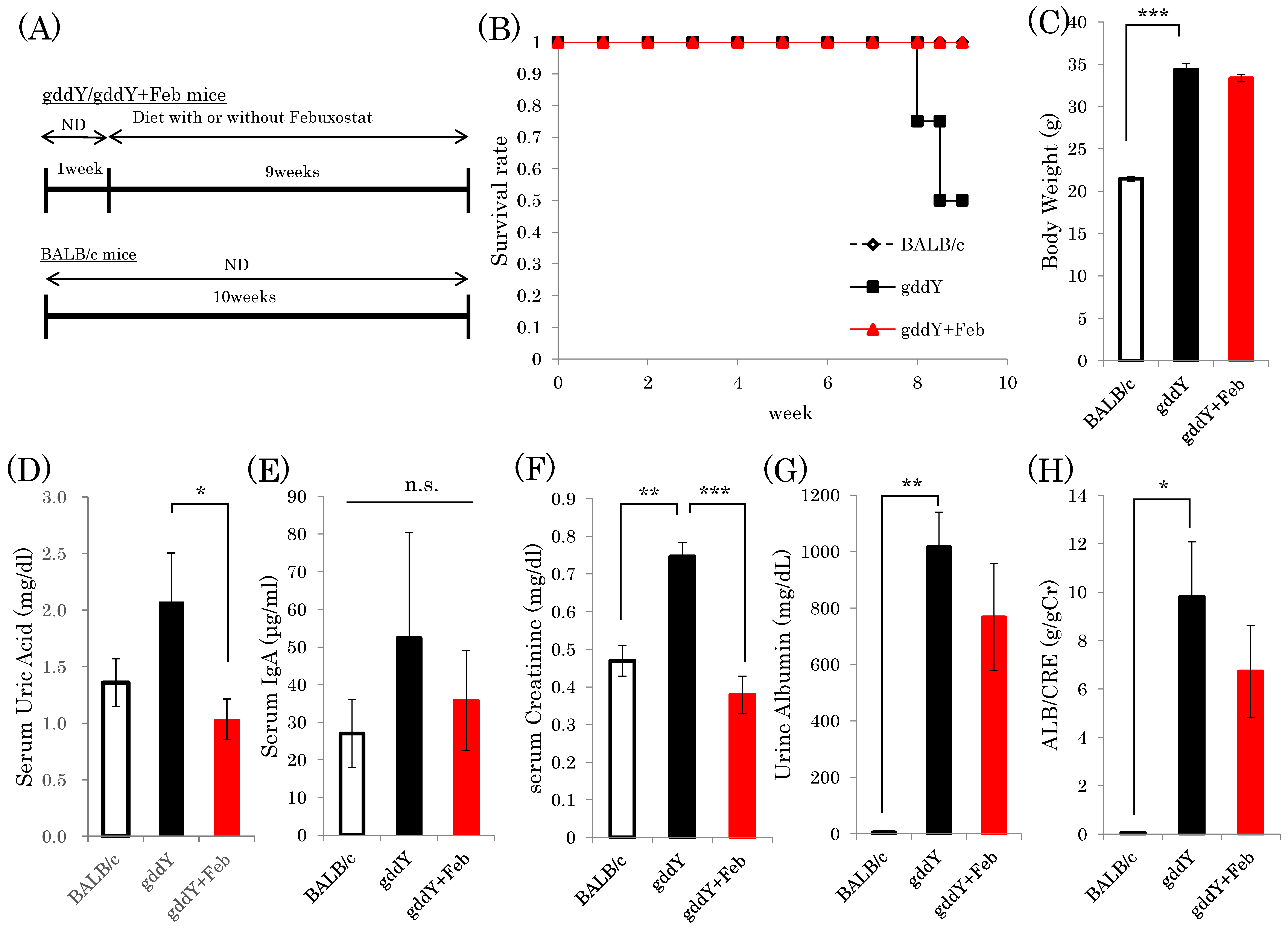
2.1. Xanthine Oxidase Inhibitor Febuxostat Prevents Progression of IgA Nephropathy
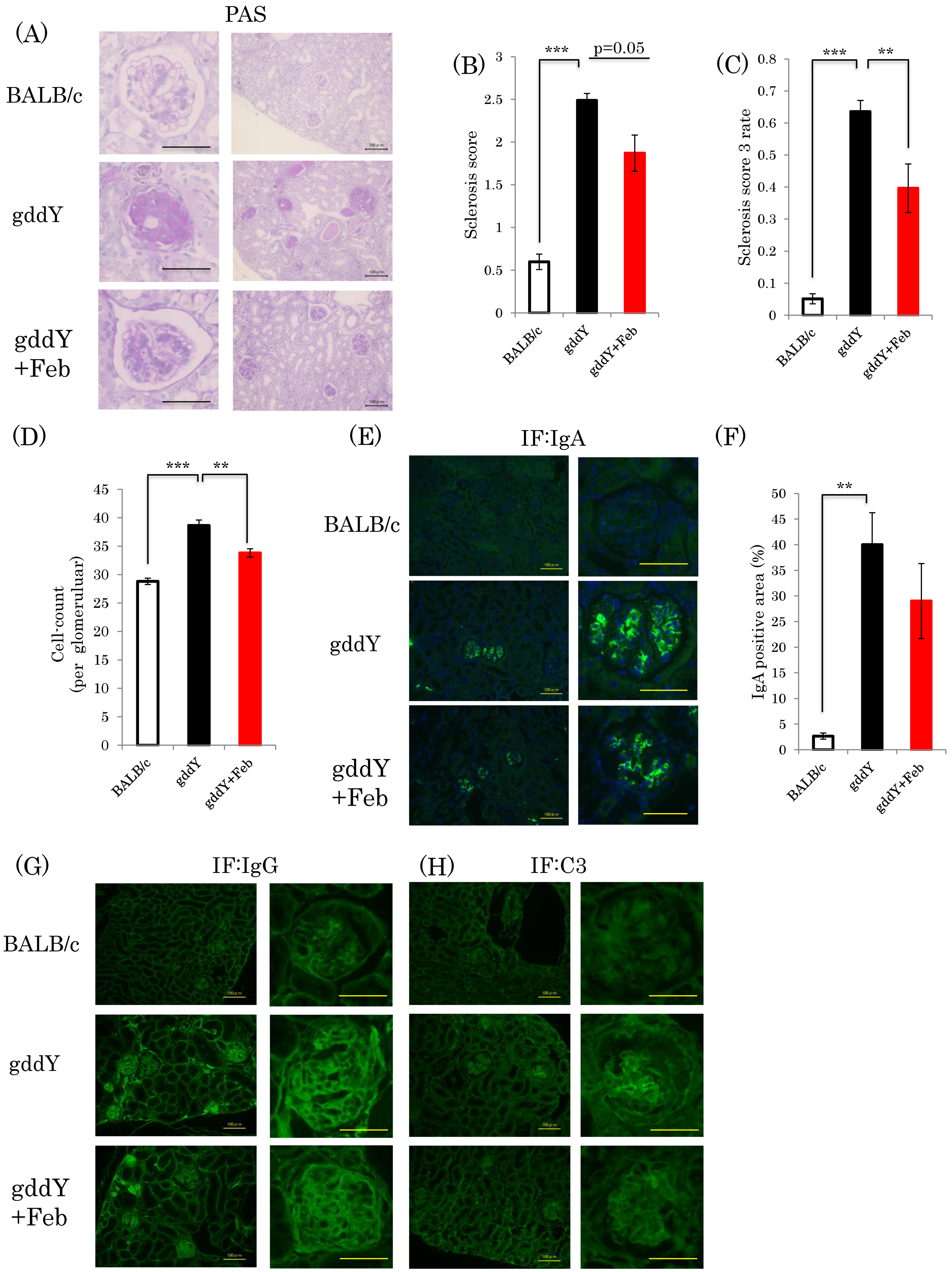
2.2. Febuxostat Suppresses Nephropathy Development without Significantly Affecting Glomerular IgA Deposition
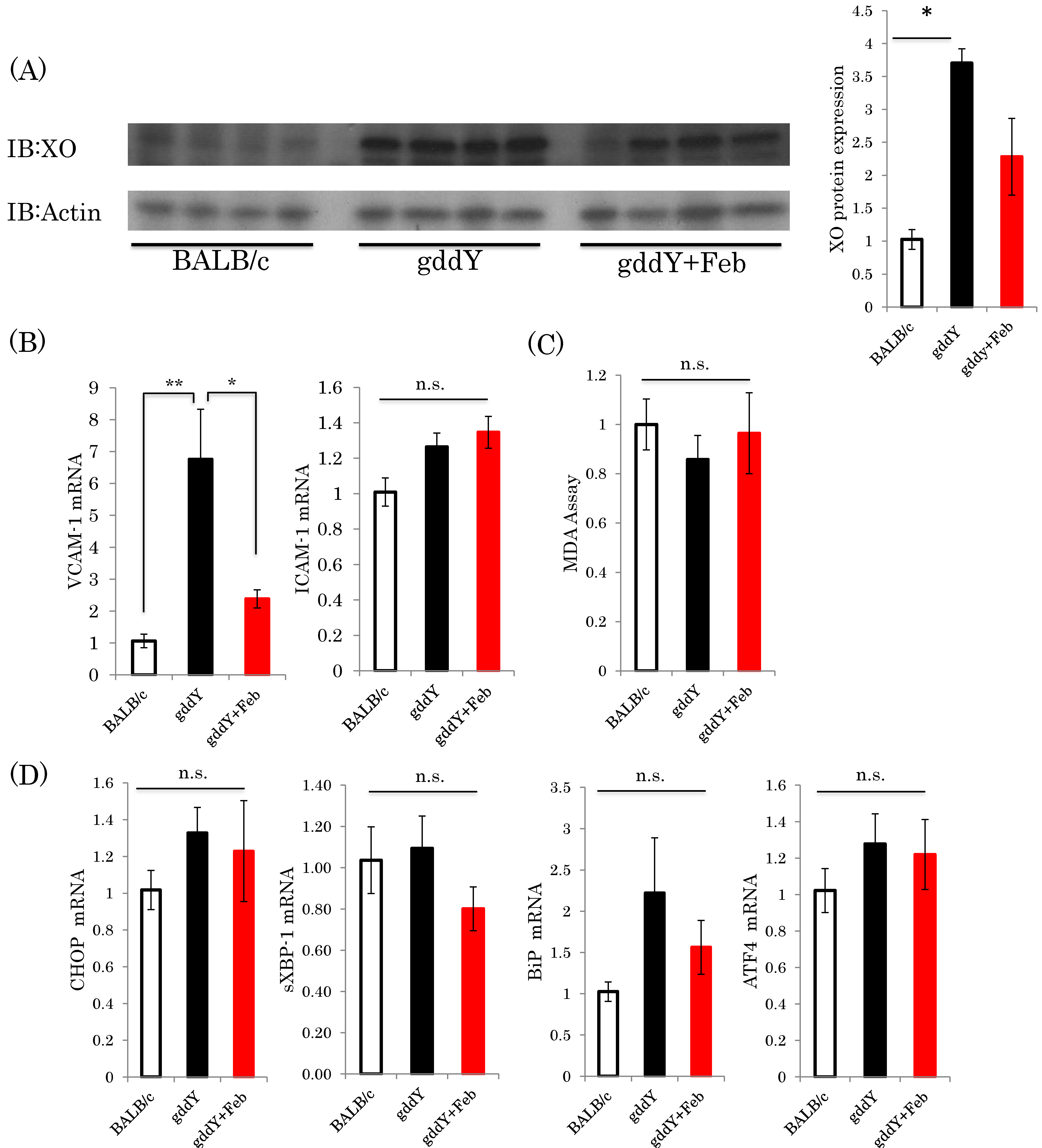
2.3. Febuxostat Suppresses Inflammation—Related Gene Expressions in the Kidneys of gddY Mice
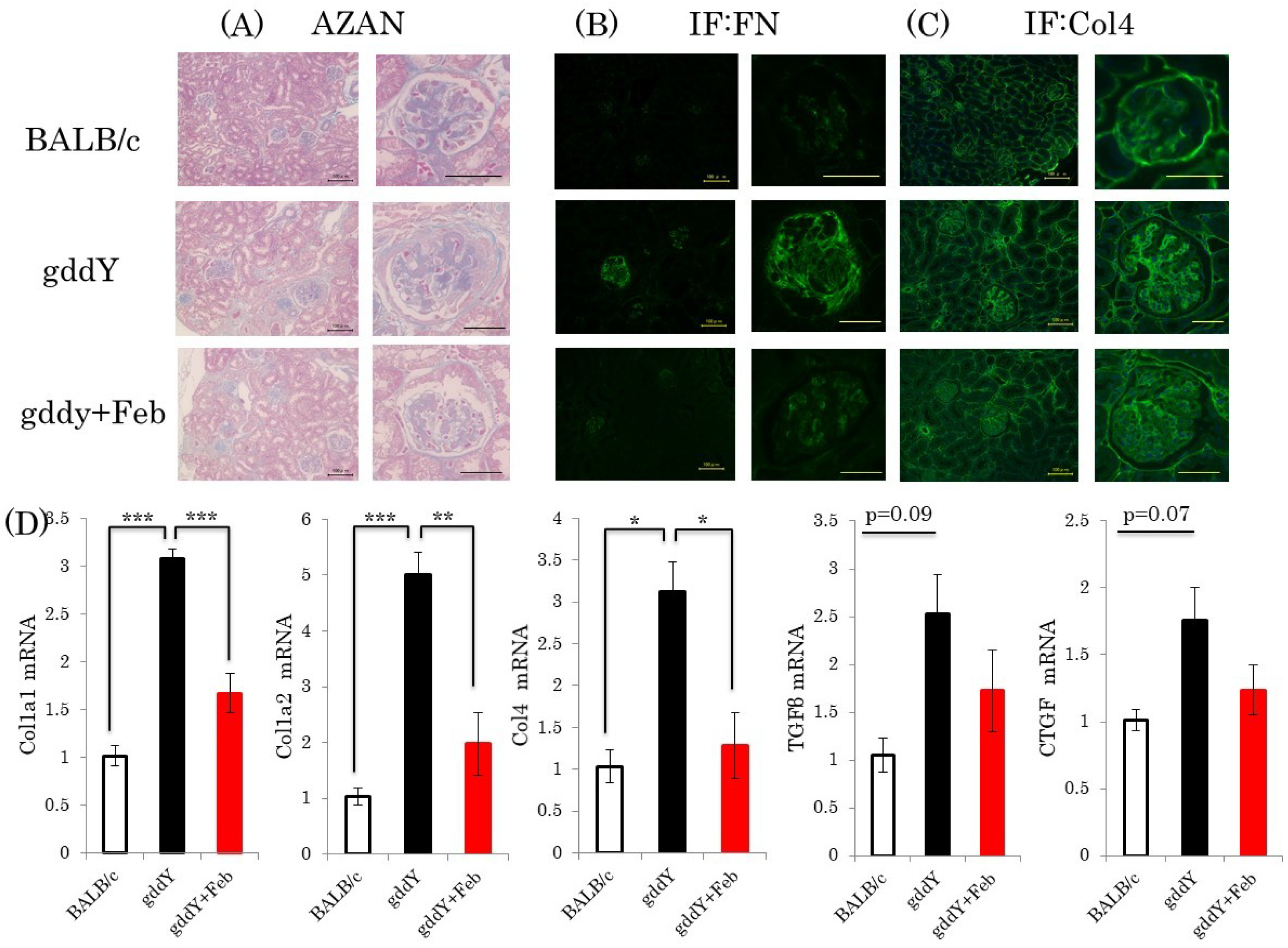
2.4. Febuxostat Ameliorates Kidney Fibrosis
2.5. Febuxostat Prevents Progression of IgA Nephropathy by Inhibiting Xanthine Oxidase
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Reagents
4.3. Metabolic Analysis
4.4. Histological Study
4.5. Measurement of Glomerular IgA Positive Area
4.6. MDA Assay
4.7. Western Blotting
4.8. Real-Time PCR
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- DuBose, T.D., Jr. American Society of Nephrology Presidential Address 2006: Chronic kidney disease as a public health threat—New strategy for a growing problem. J. Am. Soc. Nephrol. 2007, 8, 1038–1045. [Google Scholar] [CrossRef] [PubMed]
- Vejakama, P.; Atiporn, I.; McKay, G.J.; Maxwell, A.P.; McEvoy, M.; Attia, J.; Thakkinstian, A. Treatment effects of renin-angiotensin aldosterone system blockade on kidney failure and mortality in chronic kidney disease patients. BMC Nephrol. 2017, 18, 342. [Google Scholar] [CrossRef] [PubMed]
- Riccio, E.; Di Nuzzi, A.; Pisani, A. Nutritional treatment in chronic kidney disease: The concept of nephroprotection. Clin. Exp. Nephrol. 2015, 19, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Ritz, E.; Schwenger, V. Lifestyle modification and progressive renal failure. Nephrology 2005, 10, 387–392. [Google Scholar] [CrossRef]
- Tanaka, K.; Nakayama, M.; Kanno, M.; Kimura, H.; Watanabe, K.; Tani, Y.; Hayashi, Y.; Asahi, K.; Terawaki, H.; Watanabe, T. Renoprotective effects of febuxostat in hyperuricemic patients with chronic kidney disease: A parallel-group, randomized, controlled trial. Clin. Exp. Nephrol. 2015, 19, 1044–1053. [Google Scholar] [CrossRef]
- Sanchez-Lozada, L.G.; Tapia, E.; Soto, V.; Avila-Casado, C.; Franco, M.; Zhao, L.; Johnson, R.J. Treatment with the xanthine oxidase inhibitor febuxostat lowers uric acid and alleviates systemic and glomerular hypertension in experimental hyperuricaemia. Nephrol. Dial. Transpl. 2008, 23, 1179–1185. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Jeong, K.H.; Kim, Y.G.; Moon, J.Y.; Lee, S.H.; Ihm, C.G.; Sung, J.Y.; Lee, T.W. Febuxostat ameliorates diabetic renal injury in a streptozotocin-induced diabetic rat model. Am. J. Nephrol. 2014, 40, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Akimoto, T.; Morishita, Y.; Ito, T.; Iimura, O.; Tsunematsu, S.; Watanabe, Y.; Kusano, Y.; Nagata, D. Febuxostat for hyperuricemia in patients with advanced chronic kidney disease. Drug Target Insights 2014, 8, 39–43. [Google Scholar] [CrossRef]
- Becker, M.A.; Schumacher, H.R.; Espinoza, L.R.; Wells, A.F.; MacDonald, P.; Lloyd, E.; Lademacher, C. The urate-lowering efficacy and safety of febuxostat in the treatment of the hyperuricemia of gout: The CONFIRMS trial. Arthritis Res. Ther. 2010, 12, R63. [Google Scholar] [CrossRef]
- Wyatt, R.J.; Julian, B.A. The pathophysiology of IgA nephropathy. J. Am. Soc. Nephrol. 2013, 368, 2402–2414. [Google Scholar]
- Schena, F.P. A retrospective analysis of the natural history of primary IgA nephropathy worldwide. Am. J. Med. 1990, 89, 209–215. [Google Scholar] [CrossRef]
- Vogt, L.; Waanders, F.; Boomsma, F.; de Zeeuw, D.; Navis, G. Effects of dietary sodium and hydrochlorothiazide on the antiproteinuric efficacy of losartan. J. Am. Soc. Nephrol. 2008, 19, 999–1007. [Google Scholar] [CrossRef] [PubMed]
- Slagman, M.C.; Waanders, F.; Hemmelder, M.H.; Woittiez, A.J.; Janssen, W.M.; Lambers Heerspink, H.J.; Navis, G.; Laverman, G.D. Moderate dietary sodium restriction added to angiotensin converting enzyme inhibition compared with dual blockade in lowering proteinuria and blood pressure: Randomised controlled trial. BMJ 2011, 343, d4366. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Zhang, X.; Tian, J.; Li, Q.; Chen, J. Combination therapy an ACE inhibitor and an angiotensin receptor blocker for IgA nephropathy: A meta-analysis. Int. J. Clin. Pract. 2012, 66, 917–923. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Xu, D.; Perkovic, V.; Ma, X.; Johnson, D.W.; Woodward, M.; Levin, A.; Zhang, H.; Wang, H. Corticosteroid therapy in IgA nephropathy. J. Am. Soc. Nephrol. 2012, 23, 1108–1116. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Suzuki, Y.; Novak, J.; Tomino, Y. Development of animal models of human IgA nephropathy. Drug Discov. Today Dis. Models 2014, 11, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, K.; Suzuki, Y.; Otsuji, M.; Suzuki, H.; Kihara, M.; Kajiyama, T.; Hashimoto, A.; Nishimura, H.; Brown, R.; Hall, S. Development of a model of early-onset IgA nephropathy. J. Am. Soc. Nephrol. 2012, 23, 1364–1374. [Google Scholar] [CrossRef]
- El Karoui, K.; Hill, G.S.; Karras, A.; Moulonguet, L.; Caudwell, V.; Loupy, A.; Bruneval, P.; Jacquot, C.; Nochy, D. Focal segmental glomerulosclerosis plays a major role in the progression of IgA nephropathy. II. Light microscopic and clinical studies. Kidney Int. 2011, 79, 643–654. [Google Scholar] [CrossRef]
- Lim, C.S.; Yoon, H.J.; Kim, Y.S.; Ahn, C.; Han, J.S.; Kim, S.; Lee, J.S.; Lee, H.S.; Chae, D.W. Clinicopathological correlation of intrarenal cytokines and chemokines in IgA nephropathy. Nephrology 2003, 8, 21–27. [Google Scholar] [CrossRef]
- Henricus, A.; Mutsaers, M.; Stribos Elisabeth, G.D.; Glorieux, G.; Vanholder, R.; Olinga, P. Chronic kidney disease and fibrosis: The role of uremic retention solutes. Front. Med. 2015, 2, 60. [Google Scholar]
- Raij, L.; Azar, S.; Keane, W. Mesangial immune injury, hypertension, and progressive glomerular damage in Dahl rats. Kidney Int. 1984, 26, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, J.; Namba, T.; Takabatake, Y.; Kimura, T.; Takahashi, A.; Yamamoto, T.; Minami, S.; Sakai, S.; Fujimura, R.; Kaimori, J.Y. Antioxidant role of autophagy in maintaining the integrity of glomerular capillaries. Autophagy 2018, 14, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Kiryluk, K.; Novak, J.; Moldoveanu, Z.; Herr, A.B.; Renfrow, M.B.; Wyatt, R.J.; Scolari, F.; Mestecky, J.; Gharavi, A.G. The pathophysiology of IgA nephropathy. J. Am. Soc. Nephrol. 2011, 22, 1795–1803. [Google Scholar] [CrossRef] [PubMed]
- Lai, K.N.; Leung, J.C.; Chan, L.Y.; Saleem, M.A.; Mathieson, P.W.; Tam, K.Y.; Xiao, J.; Lai, F.M.; Tang, S.C. Podocyte injury induced by mesangial-derived cytokines in IgA nephropathy. Nephrol. Dial. Transp. 2009, 24, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Lai, K.N.; Leung, J.C.; Chan, L.Y.; Saleem, M.A.; Mathieson, P.W.; Lai, F.M.; Tang, S.C. Activation of podocytes by mesangial-derived TNF-alpha: Glomerulo-podocytic communication in IgA nephropathy. Am. J. Physiol. Ren. Physiol. 2008, 294, F945–F955. [Google Scholar] [CrossRef] [PubMed]
- Menon, M.C.; Chuang, P.Y.; He, J.C. Role of podocyte injury in IgA nephropathy. Contrib. Nephrol. 2013, 181, 41–51. [Google Scholar] [PubMed]
- Kushiyama, A.; Okubo, H.; Sakoda, H.; Kikuchi, T.; Fujishiro, M.; Sato, H.; Kushiyama, S.; Iwashita, M.; Nishimura, F.; Fukushima, T. Xanthine oxidoreductase is involved in macrophage foam cell formation and atherosclerosis development. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 291–298. [Google Scholar] [CrossRef]
- Nakatsu, Y.; Seno, Y.; Kushiyama, A.; Sakoda, H.; Fujishiro, M.; Katasako, A.; Mori, K.; Matsunaga, Y.; Fukushima, T.; Kanaoka, R. The xanthine oxidase inhibitor febuxostat suppresses development of nonalcoholic steatohepatitis in a rodent model. Am. J. Physiol. Gastroint. Liver Physiol. 2015, 309, G42–G51. [Google Scholar] [CrossRef]
- Nomura, J.; Busso, N.; Ives, A.; Matsui, C.; Tsujimoto, S.; Shirakura, T.; Tamura, M.; Kobayashi, T.; So, A.; Yamanaka, Y. Xanthine oxidase inhibition by febuxostat attenuates experimental atherosclerosis in mice. Sci. Rep. 2014, 4, 4554. [Google Scholar] [CrossRef]
- Yang, C.C.; Ma, M.C.; Chien, C.T.; Wu, M.S.; Sun, W.K.; Chen, C.F. Hypoxic preconditioning attenuates lipopolysaccharide-induced oxidative stress in rat kidneys. J. Physiol. 2007, 582, 407–419. [Google Scholar] [CrossRef]
- Landmesser, U.; Spiekermann, S.; Preuss, C.; Sorrentino, S.; Fischer, D.; Manes, C.; Mueller, M.; Drexler, H. Angiotensin II induces endothelial xanthine oxidase activation: Role for endothelial dysfunction in patients with coronary disease. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 943–948. [Google Scholar] [CrossRef] [PubMed]
- Ohtsubo, T.; Matsumura, K.; Sakagami, K.; Fujii, K.; Tsuruya, K.; Noguchi, H.; Rovira, II; Finkel, T.; Iida, M. Xanthine oxidoreductase depletion induces renal interstitial fibrosis through aberrant lipid and purine accumulation in renal tubules. Hypertension 2009, 5, 868–876. [Google Scholar] [CrossRef] [PubMed]
- Shibagaki, Y.; Ohno, I.; Hosoya, T.; Kimura, K. Safety, efficacy and renal effect of febuxostat in patients with moderate-to-severe kidney dysfunction. Hypertens. Res. 2014. 37, 919. [CrossRef]
- Nakamura, T.; Murase, T.; Nampei, M.; Morimoto, N.; Ashizawa, N.; Iwanaga, T.; Sakamoto, R. Effects of topiroxostat and febuxostat on urinary albumin excretion and plasma xanthine oxidoreductase activity in db/db mice. Eur. J. Pharmacol. 2016, 780, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Kosugi, T.; Nakayama, T.; Heinig, M.; Zhang, L.; Yuzawa, Y.; Sanchez-Lozada, L.G.; Roncal, C.; Johnson, R.J.; Nakagawa, T. Effect of lowering uric acid on renal disease in the type 2 diabetic db/db mice. Am. J. Physiol. Ren. Physiol. 2009, 297, F481–F488. [Google Scholar] [CrossRef] [PubMed]
- Hosoya, T.; Ohno, I.; Nomura, S.; Hisatome, I.; Uchida, S.; Fujimori, S.; Yamamoto, T.; Hara, S. Effects of topiroxostat on the serum urate levels and urinary albumin excretion in hyperuricemic stage 3 chronic kidney disease patients with or without gout. Clin. Exp. Nephrol. 2014, 18, 876–884. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, A.; Harada, M.; Yamada, Y.; Hashimoto, K.; Kamijo, Y. Identification of chronic kidney disease patient characteristics influencing the renoprotective effects of febuxostat therapy: A retrospective follow-up study. BMC Nephrol. 2017, 18, 162. [Google Scholar] [CrossRef] [PubMed]
- Szalay, C.I.; Erdélyi, K.; Kökény, G.; Lajtár, E.; Godó, M.; Révész, C.; Kaucsár, T.; Kiss, N.; Sárközy, M.; Csont, T. Oxidative/nitrative stress and inflammation drive progression of doxorubicin-induced renal fibrosis in rats as revealed by comparing a normal and a fibrosis-resistant rat strain. PLoS ONE 2015, 10, e0127090. [Google Scholar] [CrossRef]
- Lu, Y.; Wen, J.; Chen, D.P.; Wu, L.L.; Li, Q.G.; Xie, Y.; Wu, D.; Liu, X.; Chen, X.M. Modulation of cyclins and p53 in mesangial cell proliferation and apoptosis during Habu nephritis. Clin. Exp. Nephrol. 2016, 20, 178–186. [Google Scholar] [CrossRef]
- Satake, K.; Shimizu, Y.; Sasaki, Y.; Yanagawa, H.; Suzuki, H.; Suzuki, Y.; Horikoshi, S.; Honda, S.; Shibuya, K.; Shibuya, A. Serum under-O-glycosylated IgA1 level is not correlated with glomerular IgA deposition based upon heterogeneity in the composition of immune complexes in IgA nephropathy. BMC Nephrol. 2014, 15, 89. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| mCHOP | CCACCACACCTGAAGCAGAA | AGGTGAAAGGCAGGGACTCA |
| mCTGF | CAAAGCAGCTGCAAATACCA | GGCCAAATGTGTCTTCCAGT |
| mCol1a1 | GAGCGGAGAGTACTGGATCG | GCTTCTTTTCCTTGGGGTTC |
| mCol1a2 | CCGTGCTTCTCAGAACATCA | GAGCAGCCATCGACTAGGAC |
| mCol4a1 | GGCTCTGGCTGTGGAAAA | CCAGGTTCTCCAGCATCACC |
| mIL-1 | AGCTGCCACAGCTTCTCCA | TTGACGGACCCCAAAAGATG |
| mIL-6 | CCATCCAGTTGCCTTCTTGG | TGCAAGTGCATCATCGTTGT |
| mMCP1 | AGGTCCCTGTCATGCTTCTG | TCTGGACCCATTCCTTCTTG |
| mTNFa | GAACTGGCAGAAGAGGCACT | AGGGTCTGGGCCATAGAACT |
| mTGFb1 | TTGCTTCAGCTCCACAGAGA | TGGTTGTAGAGGGCAAGGAC |
| mLCN2 | CCAGTTCGCCATGGTATTTT | TCCTTCAGTTCAGGGGACAG |
| mCXCL1 | CACCCAAACCGAAGTCATAG | AAGCCAGCGTTCACCAGA |
| mCXCL2 | TCCAGAGCTTGAGTGTGACGC | TGGATGATTTTCTGAACCAGGG |
| mCXCL5 | GGTCCACAGTGCCCTACG | GCGAGTGCATTCCGCTTA |
| mF4/80 | TCTGGGGAGCTTACGATGGA | TAGGAATCCCGCAATGATGG |
| mVCAM | TCTTGGGAGCCTCAACGGTA | TGACAGGCTCCATGGTCAGA |
| msXBP1 | CTGAGTCCGAATCAGGTCCAG | GTCCATGGGAAGATGTTCTGG |
| mBiP | TTCAGCCAATTATCAGCAAACTC | TTTTCTGATGTATCCTCTTCACC |
| mATF4 | GGGTTCTGTCTTCCACTCCA | AAGCAGCAGAGTCAGGCTTCC |
| mGAPDH | TGATGGGTGTGAACCACG | GGGCCATCCACAGTCTTCTG |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Inoue, M.-K.; Yamamotoya, T.; Nakatsu, Y.; Ueda, K.; Inoue, Y.; Matsunaga, Y.; Sakoda, H.; Fujishiro, M.; Ono, H.; Morii, K.; et al. The Xanthine Oxidase Inhibitor Febuxostat Suppresses the Progression of IgA Nephropathy, Possibly via Its Anti-Inflammatory and Anti-Fibrotic Effects in the gddY Mouse Model. Int. J. Mol. Sci. 2018, 19, 3967. https://doi.org/10.3390/ijms19123967
Inoue M-K, Yamamotoya T, Nakatsu Y, Ueda K, Inoue Y, Matsunaga Y, Sakoda H, Fujishiro M, Ono H, Morii K, et al. The Xanthine Oxidase Inhibitor Febuxostat Suppresses the Progression of IgA Nephropathy, Possibly via Its Anti-Inflammatory and Anti-Fibrotic Effects in the gddY Mouse Model. International Journal of Molecular Sciences. 2018; 19(12):3967. https://doi.org/10.3390/ijms19123967
Chicago/Turabian StyleInoue, Masa-Ki, Takeshi Yamamotoya, Yusuke Nakatsu, Koji Ueda, Yuki Inoue, Yasuka Matsunaga, Hideyuki Sakoda, Midori Fujishiro, Hiraku Ono, Kenichi Morii, and et al. 2018. "The Xanthine Oxidase Inhibitor Febuxostat Suppresses the Progression of IgA Nephropathy, Possibly via Its Anti-Inflammatory and Anti-Fibrotic Effects in the gddY Mouse Model" International Journal of Molecular Sciences 19, no. 12: 3967. https://doi.org/10.3390/ijms19123967
APA StyleInoue, M.-K., Yamamotoya, T., Nakatsu, Y., Ueda, K., Inoue, Y., Matsunaga, Y., Sakoda, H., Fujishiro, M., Ono, H., Morii, K., Sasaki, K., Masaki, T., Suzuki, Y., Asano, T., & Kushiyama, A. (2018). The Xanthine Oxidase Inhibitor Febuxostat Suppresses the Progression of IgA Nephropathy, Possibly via Its Anti-Inflammatory and Anti-Fibrotic Effects in the gddY Mouse Model. International Journal of Molecular Sciences, 19(12), 3967. https://doi.org/10.3390/ijms19123967