The Interleukin-17 Family of Cytokines in Breast Cancer

 ,
, 
Abstract
1. Introduction
2. The IL-17/IL-17R Axis
2.1. The Interleukin-17 Family of Cytokine
2.2. The Interleukin-17 Receptor
2.3. Cellular Production of IL-17
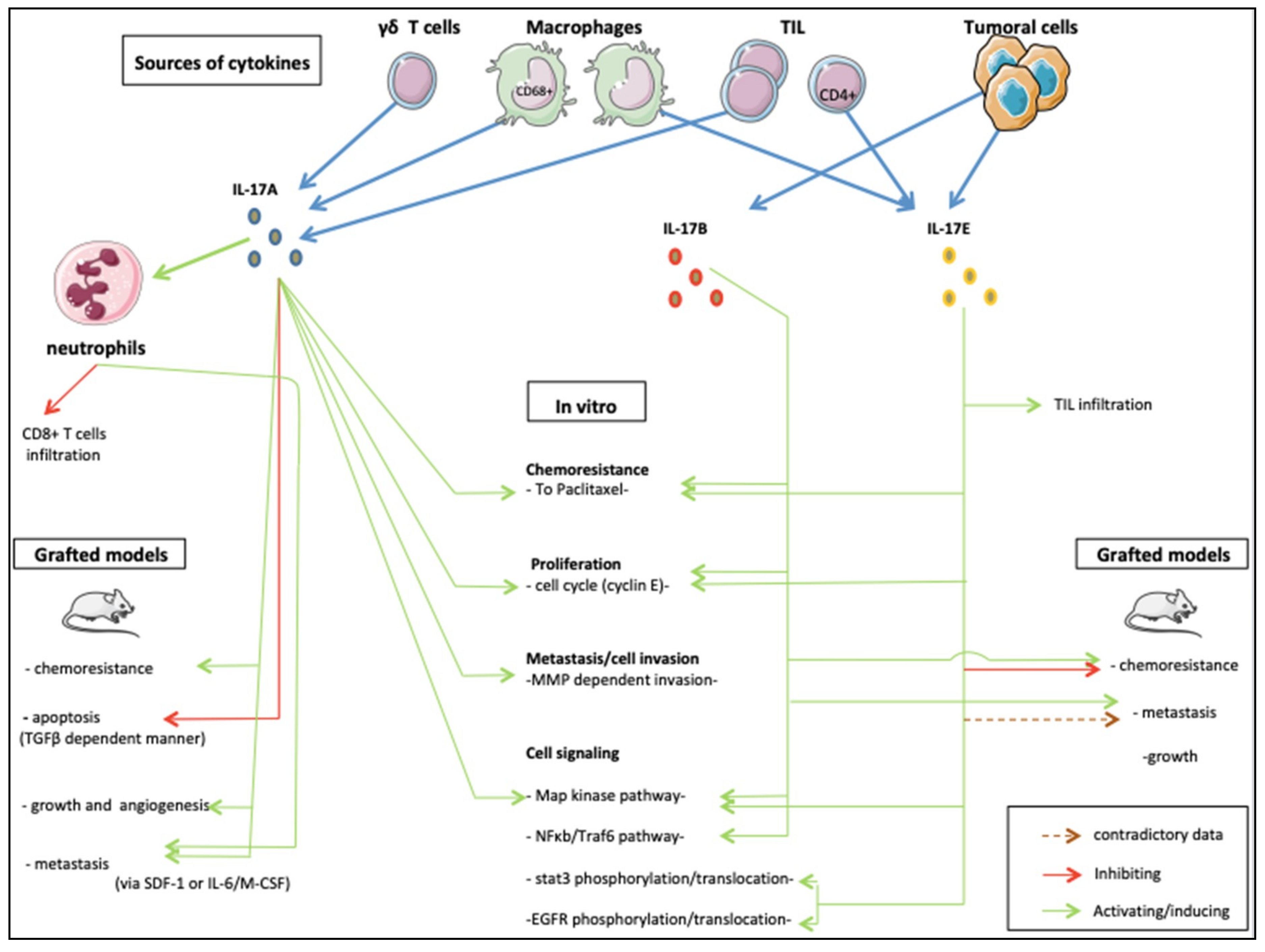
3. Roles of IL-17 in Breast Cancer Models
4. Discussion
5. Patents
Funding
Conflicts of Interest
Abbreviations
| BC | Breast cancer |
| CDK | Cyclin dependent kinase |
| mTOR | Mammalian target of rapamycin |
| IL | Interleukin |
| NSCLC | Nonsquamous cells lung cancer |
| IC-NST | Invasive carcinoma of non special type |
| ILC | Invasive lobular carcinoma |
| ER | Estrogen receptor |
| PR | Progesterone receptor |
| HER | Human epidermal growth factor receptor |
| TKI | Tyrosine kinase inhibitors |
| PARP | Poly adenosine diphosphate ribose polymerase |
| SEER | Surveillance epidemiology and end results |
| TNBC | Triple negative breast cancer |
| OS | Overall survival |
| CTLA8 | Cytotoxic T lymphocytes associated lymphocytes 8 |
| AUR | Adenylate uridylate rich |
| NF-kB | Nuclear factor kappa B |
| cDNA | Complentary desoxyribonucleic acid |
| TGF-β | Transforming growth factor β |
| IL-17R | Interleukin-17 receptor |
| SEFIR | Similar expression to fibroblast growth factor genes and IL-17R |
| TRAF | Tumor necrosing factor receptor associated factor |
| MAPK | Mitogen-activated protein kinase |
| ERK | Extracellular signal-regulated kinase |
| p38 | p38 kinase |
| JNK | c-Jun N-terminal kinase |
| IKKi | I kappa B kinase |
| TBK1 | TANK binding kinase 1 |
| mRNA | Messenger ribonucleic acid |
| HuR | Human antigen R |
| ASF/SF2 | Serine/arginine rich splicing factor 2 |
| TAK1 | Transforming growth factor β activated kinase 1 |
| SMURF2 | Smad ubiquitination regulatory factor |
| DAZAP2 | Deleted in azoospermia associated 2 |
| GM-CSF | Granulocyte-macrophage colony-stimulating factor |
| STAT | Signal transducer and activator of transcription |
| TYK | Tyrosine kinase |
| RORγ | RAR-related orphan receptor gamma |
| ITRC | Innate tissue resident cells |
| ILC3 | type 3 innate lymphoid cells |
| CCR | Chemokine receptor |
| SBR | Scarf Bloom and Richardson |
| BrdU | Bromodeoxyuridine |
| siRNA | Small interfering ribonucleic acid |
| c-RAF | RAF proto-oncogene serine/threonine-protein kinase |
| STAT | Signal transducer and activator of transcription |
| TPL2 | Tumor progression locus 2 |
| TIL | Tumor-infiltrating lymphocytes |
| MMP | Matrix metalloproteinase |
| SDF-1 | Stromal derived factor 1 |
| M-CSF | Macrophage colony-stimulating factor |
| SCID | Severe combined immunodeficiency |
| CM | Conditioned medium |
| MEC | Mammary epithelial cell |
| MDSC | Myeloid-derived suppressor cells |
| LMWCE | Low molecular weight cyclin E |
| MEKK | MAPK/extracellular signal-regulated kinase kinase |
| PrC | Prostate cancer |
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Dyba, T.; Randi, G.; Bettio, M.; Gavin, A.; Visser, O.; Bray, F. Cancer incidence and mortality patterns in europe: Estimates for 40 countries and 25 major cancers in 2018. Eur. J. Cancer 2018, 103, 356–387. [Google Scholar] [CrossRef]
- Senkus, E.; Kyriakides, S.; Ohno, S.; Penault-Llorca, F.; Poortmans, P.; Rutgers, E.; Zackrisson, S.; Cardoso, F.; Committee, E.G. Primary breast cancer: Esmo clinical practice guidelines for diagnosis, treatment and follow-up. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol./ESMO 2015, 26 (Suppl. 5), v8–v30. [Google Scholar] [CrossRef] [PubMed]
- Altundag, K. Have all triple-negative breast cancer patients worse breast cancer-specific survival? Breast 2017, 36, 102. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Spezia, M.; Huang, S.; Yuan, C.; Zeng, Z.; Zhang, L.; Ji, X.; Liu, W.; Huang, B.; Luo, W.; et al. Breast cancer development and progression: Risk factors, cancer stem cells, signaling pathways, genomics, and molecular pathogenesis. Genes Dis. 2018, 5, 77–106. [Google Scholar] [CrossRef]
- Lal, S.; McCart Reed, A.E.; de Luca, X.M.; Simpson, P.T. Molecular signatures in breast cancer. Methods 2017, 131, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Sorlie, T.; Perou, C.M.; Tibshirani, R.; Aas, T.; Geisler, S.; Johnsen, H.; Hastie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; et al. Gene expression patterns of breast carcinomas distinguish tumor subclasses with clinical implications. Proc. Natl. Acad. Sci. USA 2001, 98, 10869–10874. [Google Scholar] [CrossRef]
- Perou, C.M.; Sorlie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular portraits of human breast tumours. Nature 2000, 406, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Russnes, H.G.; Lingjaerde, O.C.; Borresen-Dale, A.L.; Caldas, C. Breast cancer molecular stratification: From intrinsic subtypes to integrative clusters. Am. J. Pathol. 2017, 187, 2152–2162. [Google Scholar] [CrossRef]
- Cardoso, F.; Senkus, E.; Costa, A.; Papadopoulos, E.; Aapro, M.; Andre, F.; Harbeck, N.; Aguilar Lopez, B.; Barrios, C.H.; Bergh, J.; et al. 4th eso-esmo international consensus guidelines for advanced breast cancer (abc 4)dagger. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol./ESMO 2018, 29, 1634–1657. [Google Scholar] [CrossRef] [PubMed]
- Ballinger, T.J.; Meier, J.B.; Jansen, V.M. Current landscape of targeted therapies for hormone-receptor positive, her2 negative metastatic breast cancer. Front. Oncol. 2018, 8, 308. [Google Scholar] [CrossRef]
- Westphal, T.; Gampenrieder, S.P.; Rinnerthaler, G.; Greil, R. Cure in metastatic breast cancer. Memo 2018, 11, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Eng, L.G.; Dawood, S.; Sopik, V.; Haaland, B.; Tan, P.S.; Bhoo-Pathy, N.; Warner, E.; Iqbal, J.; Narod, S.A.; Dent, R. Ten-year survival in women with primary stage iv breast cancer. Breast Cancer Res. Treat. 2016, 160, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Dirix, L.Y.; Takacs, I.; Jerusalem, G.; Nikolinakos, P.; Arkenau, H.T.; Forero-Torres, A.; Boccia, R.; Lippman, M.E.; Somer, R.; Smakal, M.; et al. Avelumab, an anti-pd-l1 antibody, in patients with locally advanced or metastatic breast cancer: A phase 1b javelin solid tumor study. Breast Cancer Res. Treat. 2018, 167, 671–686. [Google Scholar] [CrossRef] [PubMed]
- Wein, L.; Luen, S.J.; Savas, P.; Salgado, R.; Loi, S. Checkpoint blockade in the treatment of breast cancer: Current status and future directions. Br. J. Cancer 2018, 119, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Hui, L.; Chen, Y. Tumor microenvironment: Sanctuary of the devil. Cancer Lett. 2015, 368, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Genova, C.; Rossi, G.; Rijavec, E.; Biello, F.; Barletta, G.; Tagliamento, M.; Grossi, F. Releasing the brake: Safety profile of immune check-point inhibitors in non-small cell lung cancer. Expert Opin. Drug Saf. 2017, 16, 573–585. [Google Scholar] [CrossRef] [PubMed]
- Ascierto, P.A.; McArthur, G.A. Checkpoint inhibitors in melanoma and early phase development in solid tumors: What’s the future? J. Transl. Med. 2017, 15, 173. [Google Scholar] [CrossRef] [PubMed]
- Samadi, P.; Saki, S.; Dermani, F.K.; Pourjafar, M.; Saidijam, M. Emerging ways to treat breast cancer: Will promises be met? Cell. Oncol. (Dordr.) 2018, 41, 605–621. [Google Scholar] [CrossRef]
- Balkwill, F.; Mantovani, A. Inflammation and cancer: Back to virchow? Lancet 2001, 357, 539–545. [Google Scholar] [CrossRef]
- Diakos, C.I.; Charles, K.A.; McMillan, D.C.; Clarke, S.J. Cancer-related inflammation and treatment effectiveness. Lancet Oncol. 2014, 15, e493–e503. [Google Scholar] [CrossRef]
- DeNardo, D.G.; Coussens, L.M. Inflammation and breast cancer. Balancing immune response: Crosstalk between adaptive and innate immune cells during breast cancer progression. Breast Cancer Res. 2007, 9, 212. [Google Scholar] [CrossRef] [PubMed]
- Setrerrahmane, S.; Xu, H. Tumor-related interleukins: Old validated targets for new anti-cancer drug development. Mol. Cancer 2017, 16, 153. [Google Scholar] [CrossRef] [PubMed]
- Sloot, Y.J.E.; Smit, J.W.; Joosten, L.A.B.; Netea-Maier, R.T. Insights into the role of il-32 in cancer. Semin. Immunol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Holen, I.; Lefley, D.V.; Francis, S.E.; Rennicks, S.; Bradbury, S.; Coleman, R.E.; Ottewell, P. Il-1 drives breast cancer growth and bone metastasis in vivo. Oncotarget 2016, 7, 75571–75584. [Google Scholar] [CrossRef] [PubMed]
- Heo, T.H.; Wahler, J.; Suh, N. Potential therapeutic implications of il-6/il-6r/gp130-targeting agents in breast cancer. Oncotarget 2016, 7, 15460–15473. [Google Scholar] [CrossRef] [PubMed]
- Amatya, N.; Garg, A.V.; Gaffen, S.L. Il-17 signaling: The yin and the yang. Trends Immunol. 2017, 38, 310–322. [Google Scholar] [CrossRef]
- Song, Y.; Yang, J.M. Role of interleukin (il)-17 and t-helper (th)17 cells in cancer. Biochem. Biophys. Res. Commun. 2017, 493, 1–8. [Google Scholar] [CrossRef]
- Rouvier, E.; Luciani, M.F.; Mattei, M.G.; Denizot, F.; Golstein, P. Ctla-8, cloned from an activated t cell, bearing au-rich messenger rna instability sequences, and homologous to a herpesvirus saimiri gene. J. Immunol. 1993, 150, 5445–5456. [Google Scholar]
- Yao, Z.; Painter, S.L.; Fanslow, W.C.; Ulrich, D.; Macduff, B.M.; Spriggs, M.K.; Armitage, R.J. Human il-17: A novel cytokine derived from t cells. J. Immunol. 1995, 155, 5483–5486. [Google Scholar]
- Gaffen, S.L. Life before seventeen: Cloning of the il-17 receptor. J. Immunol. 2011, 187, 4389–4391. [Google Scholar] [CrossRef]
- Isailovic, N.; Daigo, K.; Mantovani, A.; Selmi, C. Interleukin-17 and innate immunity in infections and chronic inflammation. J. Autoimmun. 2015, 60, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Fabre, J.; Giustiniani, J.; Garbar, C.; Antonicelli, F.; Merrouche, Y.; Bensussan, A.; Bagot, M.; Al-Dacak, R. Targeting the tumor microenvironment: The protumor effects of il-17 related to cancer type. Int. J. Mol. Sci. 2016, 17, 1433. [Google Scholar] [CrossRef] [PubMed]
- Fabre, J.; Giustiniani, J.; Antonicelli, F.; Merrouche, Y.; Bagot, M.; Bensussan, A. A focus on il-17 targeting treatments in psoriasis. J. Dermatol. Res. 2016, 2, 1–5. [Google Scholar] [CrossRef]
- Giusti, D.; Le Jan, S.; Gatouillat, G.; Bernard, P.; Pham, B.N.; Antonicelli, F. Biomarkers related to bullous pemphigoid activity and outcome. Exp. Dermatol. 2017, 26, 1240–1247. [Google Scholar] [CrossRef]
- Kim, G.; Khanal, P.; Lim, S.C.; Yun, H.J.; Ahn, S.G.; Ki, S.H.; Choi, H.S. Interleukin-17 induces ap-1 activity and cellular transformation via upregulation of tumor progression locus 2 activity. Carcinogenesis 2013, 34, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Le Jan, S.; Plee, J.; Vallerand, D.; Dupont, A.; Delanez, E.; Durlach, A.; Jackson, P.L.; Edwin Blalock, J.; Bernard, P.; Antonicelli, F. Innate immune cell-produced il-17 sustains inflammation in bullous pemphigoid. J. Investig. Dermatol. 2014, 134, 2908–2917. [Google Scholar] [CrossRef] [PubMed]
- Plee, J.; Le Jan, S.; Giustiniani, J.; Barbe, C.; Joly, P.; Bedane, C.; Vabres, P.; Truchetet, F.; Aubin, F.; Antonicelli, F.; et al. Integrating longitudinal serum il-17 and il-23 follow-up, along with autoantibodies variation, contributes to predict bullous pemphigoid outcome. Sci. Rep. 2015, 5, 18001. [Google Scholar] [CrossRef]
- Fan, Z.; Yang, J.; Yang, C.; Guo, X. Il-17: A promising therapeutic target for atherosclerosis. Int. J. Cardiol. 2016, 202, 930–931. [Google Scholar] [CrossRef]
- Fossiez, F.; Djossou, O.; Chomarat, P.; Flores-Romo, L.; Ait-Yahia, S.; Maat, C.; Pin, J.J.; Garrone, P.; Garcia, E.; Saeland, S.; et al. T cell interleukin-17 induces stromal cells to produce proinflammatory and hematopoietic cytokines. J. Exp. Med. 1996, 183, 2593–2603. [Google Scholar] [CrossRef]
- Hymowitz, S.G.; Filvaroff, E.H.; Yin, J.P.; Lee, J.; Cai, L.; Risser, P.; Maruoka, M.; Mao, W.; Foster, J.; Kelley, R.F.; et al. Il-17s adopt a cystine knot fold: Structure and activity of a novel cytokine, il-17f, and implications for receptor binding. EMBO J. 2001, 20, 5332–5341. [Google Scholar] [CrossRef]
- McDonald, N.Q.; Hendrickson, W.A. A structural superfamily of growth factors containing a cystine knot motif. Cell 1993, 73, 421–424. [Google Scholar] [CrossRef]
- Moseley, T.A.; Haudenschild, D.R.; Rose, L.; Reddi, A.H. Interleukin-17 family and il-17 receptors. Cytokine Growth Factor Rev. 2003, 14, 155–174. [Google Scholar] [CrossRef]
- Chang, S.H.; Dong, C. A novel heterodimeric cytokine consisting of il-17 and il-17f regulates inflammatory responses. Cell Res. 2007, 17, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.F.; Guo, Y.; Quazi, A.; Luxenberg, D.P.; Bennett, F.; Ross, J.F.; Qiu, Y.; Whitters, M.J.; Tomkinson, K.N.; Dunussi-Joannopoulos, K.; et al. Identification of an interleukin 17f/17a heterodimer in activated human cd4+ t cells. J. Biol. Chem. 2007, 282, 13447–13455. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.D.; Zhang, H.; He, M.X. Comparative and evolutionary analysis of the interleukin 17 gene family in invertebrates. PLoS ONE 2015, 10, e0132802. [Google Scholar] [CrossRef] [PubMed]
- Kolls, J.K.; Linden, A. Interleukin-17 family members and inflammation. Immunity 2004, 21, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Iwakura, Y.; Ishigame, H.; Saijo, S.; Nakae, S. Functional specialization of interleukin-17 family members. Immunity 2011, 34, 149–162. [Google Scholar] [CrossRef]
- Bie, Q.; Jin, C.; Zhang, B.; Dong, H. Il-17b: A new area of study in the il-17 family. Mol. Immunol. 2017, 90, 50–56. [Google Scholar] [CrossRef]
- Zhang, B.; Liu, C.; Qian, W.; Han, Y.; Li, X.; Deng, J. Structure of the unique sefir domain from human interleukin 17 receptor a reveals a composite ligand-binding site containing a conserved alpha-helix for act1 binding and il-17 signaling. Acta Crystallogr. D Biol. Crystallogr. 2014, 70, 1476–1483. [Google Scholar] [CrossRef]
- Patel, D.D.; Lee, D.M.; Kolbinger, F.; Antoni, C. Effect of il-17a blockade with secukinumab in autoimmune diseases. Ann. Rheum. Dis. 2013, 72 (Suppl. 2) (Suppl. 2), ii116–ii123. [Google Scholar] [CrossRef]
- Wright, J.F.; Bennett, F.; Li, B.; Brooks, J.; Luxenberg, D.P.; Whitters, M.J.; Tomkinson, K.N.; Fitz, L.J.; Wolfman, N.M.; Collins, M.; et al. The human il-17f/il-17a heterodimeric cytokine signals through the il-17ra/il-17rc receptor complex. J. Immunol. 2008, 181, 2799–2805. [Google Scholar] [CrossRef] [PubMed]
- Zrioual, S.; Toh, M.L.; Tournadre, A.; Zhou, Y.; Cazalis, M.A.; Pachot, A.; Miossec, V.; Miossec, P. Il-17ra and il-17rc receptors are essential for il-17a-induced elr+ cxc chemokine expression in synoviocytes and are overexpressed in rheumatoid blood. J. Immunol. 2008, 180, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Kuestner, R.E.; Taft, D.W.; Haran, A.; Brandt, C.S.; Brender, T.; Lum, K.; Harder, B.; Okada, S.; Ostrander, C.D.; Kreindler, J.L.; et al. Identification of the il-17 receptor related molecule il-17rc as the receptor for il-17f. J. Immunol. 2007, 179, 5462–5473. [Google Scholar] [CrossRef] [PubMed]
- Monin, L.; Gaffen, S.L. Interleukin 17 family cytokines: Signaling mechanisms, biological activities, and therapeutic implications. Cold Spring Harb. Perspect. Biol. 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T.R.; Cherwinski, H.; Bond, M.W.; Giedlin, M.A.; Coffman, R.L. Two types of murine helper t cell clone. I. Definition according to profiles of lymphokine activities and secreted proteins. J. Immunol. 1986, 136, 2348–2357. [Google Scholar]
- Aarvak, T.; Chabaud, M.; Miossec, P.; Natvig, J.B. Il-17 is produced by some proinflammatory th1/th0 cells but not by th2 cells. J. Immunol. 1999, 162, 1246–1251. [Google Scholar]
- Steinman, L. A brief history of t(h)17, the first major revision in the t(h)1/t(h)2 hypothesis of t cell-mediated tissue damage. Nat. Med. 2007, 13, 139–145. [Google Scholar] [CrossRef]
- Park, H.; Li, Z.; Yang, X.O.; Chang, S.H.; Nurieva, R.; Wang, Y.H.; Wang, Y.; Hood, L.; Zhu, Z.; Tian, Q.; et al. A distinct lineage of cd4 t cells regulates tissue inflammation by producing interleukin 17. Nat. Immunol. 2005, 6, 1133–1141. [Google Scholar] [CrossRef]
- Korn, T.; Oukka, M.; Kuchroo, V.; Bettelli, E. Th17 cells: Effector t cells with inflammatory properties. Semin. Immunol. 2007, 19, 362–371. [Google Scholar] [CrossRef]
- Liang, S.C.; Tan, X.Y.; Luxenberg, D.P.; Karim, R.; Dunussi-Joannopoulos, K.; Collins, M.; Fouser, L.A. Interleukin (il)-22 and il-17 are coexpressed by th17 cells and cooperatively enhance expression of antimicrobial peptides. J. Exp. Med. 2006, 203, 2271–2279. [Google Scholar] [CrossRef]
- Nurieva, R.; Yang, X.O.; Martinez, G.; Zhang, Y.; Panopoulos, A.D.; Ma, L.; Schluns, K.; Tian, Q.; Watowich, S.S.; Jetten, A.M.; et al. Essential autocrine regulation by il-21 in the generation of inflammatory t cells. Nature 2007, 448, 480–483. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.A. The bench-to-bedside story of il-17 and the therapeutic efficacy of its targeting in spondyloarthritis. Curr. Rheumatol. Rep. 2016, 18, 33. [Google Scholar] [CrossRef] [PubMed]
- Veldhoen, M.; Hocking, R.J.; Atkins, C.J.; Locksley, R.M.; Stockinger, B. Tgfbeta in the context of an inflammatory cytokine milieu supports de novo differentiation of il-17-producing t cells. Immunity 2006, 24, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Bystrom, J.; Taher, T.E.; Muhyaddin, M.S.; Clanchy, F.I.; Mangat, P.; Jawad, A.S.; Williams, R.O.; Mageed, R.A. Harnessing the therapeutic potential of th17 cells. Med. Inflamm. 2015, 2015, 205156. [Google Scholar] [CrossRef] [PubMed]
- Ciofani, M.; Madar, A.; Galan, C.; Sellars, M.; Mace, K.; Pauli, F.; Agarwal, A.; Huang, W.; Parkhurst, C.N.; Muratet, M.; et al. A validated regulatory network for th17 cell specification. Cell 2012, 151, 289–303. [Google Scholar] [CrossRef] [PubMed]
- Huber, M.; Heink, S.; Pagenstecher, A.; Reinhard, K.; Ritter, J.; Visekruna, A.; Guralnik, A.; Bollig, N.; Jeltsch, K.; Heinemann, C.; et al. Il-17a secretion by cd8+ t cells supports th17-mediated autoimmune encephalomyelitis. J. Clin. Investig. 2013, 123, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Kronenberg, M. Toward an understanding of nkt cell biology: Progress and paradoxes. Annu. Rev. Immunol. 2005, 23, 877–900. [Google Scholar] [CrossRef] [PubMed]
- Marks, B.R.; Nowyhed, H.N.; Choi, J.Y.; Poholek, A.C.; Odegard, J.M.; Flavell, R.A.; Craft, J. Thymic self-reactivity selects natural interleukin 17-producing t cells that can regulate peripheral inflammation. Nat. Immunol. 2009, 10, 1125–1132. [Google Scholar] [CrossRef]
- Cua, D.J.; Sherlock, J.; Chen, Y.; Murphy, C.A.; Joyce, B.; Seymour, B.; Lucian, L.; To, W.; Kwan, S.; Churakova, T.; et al. Interleukin-23 rather than interleukin-12 is the critical cytokine for autoimmune inflammation of the brain. Nature 2003, 421, 744–748. [Google Scholar] [CrossRef]
- Noordenbos, T.; Blijdorp, I.; Chen, S.; Stap, J.; Mul, E.; Canete, J.D.; Lubberts, E.; Yeremenko, N.; Baeten, D. Human mast cells capture, store, and release bioactive, exogenous il-17a. J. Leukoc. Biol. 2016, 100, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.M.; Rubin, C.J.; Khandpur, R.; Wang, J.Y.; Riblett, M.; Yalavarthi, S.; Villanueva, E.C.; Shah, P.; Kaplan, M.J.; Bruce, A.T. Mast cells and neutrophils release il-17 through extracellular trap formation in psoriasis. J. Immunol. 2011, 187, 490–500. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Karin, M. The il-23 to il-17 cascade inflammation-related cancers. Clin. Exp. Rheumatol. 2015, 33, S87–S90. [Google Scholar] [PubMed]
- Morita, H.; Arae, K.; Unno, H.; Toyama, S.; Motomura, K.; Matsuda, A.; Suto, H.; Okumura, K.; Sudo, K.; Takahashi, T.; et al. Il-25 and il-33 contribute to development of eosinophilic airway inflammation in epicutaneously antigen-sensitized mice. PLoS ONE 2015, 10, e0134226. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.H.; Angkasekwinai, P.; Lu, N.; Voo, K.S.; Arima, K.; Hanabuchi, S.; Hippe, A.; Corrigan, C.J.; Dong, C.; Homey, B.; et al. Il-25 augments type 2 immune responses by enhancing the expansion and functions of tslp-dc-activated th2 memory cells. J. Exp. Med. 2007, 204, 1837–1847. [Google Scholar] [CrossRef] [PubMed]
- Manni, M.L.; Robinson, K.M.; Alcorn, J.F. A tale of two cytokines: Il-17 and il-22 in asthma and infection. Expert Rev. Respir. Med. 2014, 8, 25–42. [Google Scholar] [CrossRef] [PubMed]
- Corrigan, C.J.; Wang, W.; Meng, Q.; Fang, C.; Eid, G.; Caballero, M.R.; Lv, Z.; An, Y.; Wang, Y.H.; Liu, Y.J.; et al. Allergen-induced expression of il-25 and il-25 receptor in atopic asthmatic airways and late-phase cutaneous responses. J. Allergy Clin. Immunol. 2011, 128, 116–124. [Google Scholar] [CrossRef]
- Kleinschek, M.A.; Owyang, A.M.; Joyce-Shaikh, B.; Langrish, C.L.; Chen, Y.; Gorman, D.M.; Blumenschein, W.M.; McClanahan, T.; Brombacher, F.; Hurst, S.D.; et al. Il-25 regulates th17 function in autoimmune inflammation. J. Exp. Med. 2007, 204, 161–170. [Google Scholar] [CrossRef]
- Ramirez-Carrozzi, V.; Sambandam, A.; Luis, E.; Lin, Z.; Jeet, S.; Lesch, J.; Hackney, J.; Kim, J.; Zhou, M.; Lai, J.; et al. Il-17c regulates the innate immune function of epithelial cells in an autocrine manner. Nat. Immunol. 2011, 12, 1159–1166. [Google Scholar] [CrossRef]
- Hwang, S.Y.; Kim, H.Y. Expression of il-17 homologs and their receptors in the synovial cells of rheumatoid arthritis patients. Mol. Cells 2005, 19, 180–184. [Google Scholar]
- Li, H.; Chen, J.; Huang, A.; Stinson, J.; Heldens, S.; Foster, J.; Dowd, P.; Gurney, A.L.; Wood, W.I. Cloning and characterization of il-17b and il-17c, two new members of the il-17 cytokine family. Proc. Natl. Acad. Sci. USA 2000, 97, 773–778. [Google Scholar] [CrossRef]
- Starnes, T.; Broxmeyer, H.E.; Robertson, M.J.; Hromas, R. Cutting edge: Il-17d, a novel member of the il-17 family, stimulates cytokine production and inhibits hemopoiesis. J. Immunol. 2002, 169, 642–646. [Google Scholar] [CrossRef] [PubMed]
- Salvator Fabre, J.A. The multifaceted immune system. MOJ Immunol. 2016, 3. [Google Scholar] [CrossRef]
- Bonnet, M.C.; Bagot, M.; Bensussan, A. Monoclonal antibodies targeting il-17a or its receptor in psoriasis: A new therapeutic approach? Med. Sci. (Paris) 2012, 28, 1035–1037. [Google Scholar] [CrossRef] [PubMed]
- Van Uden, D.J.; van Laarhoven, H.W.; Westenberg, A.H.; de Wilt, J.H.; Blanken-Peeters, C.F. Inflammatory breast cancer: An overview. Crit. Rev. Oncol./Hematol. 2015, 93, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Mulcahy, L.A.; Mohammed, R.A.; Lee, A.H.; Franks, H.A.; Kilpatrick, L.; Yilmazer, A.; Paish, E.C.; Ellis, I.O.; Patel, P.M.; et al. Il-17 expression by breast-cancer-associated macrophages: Il-17 promotes invasiveness of breast cancer cell lines. Breast Cancer Res. 2008, 10, R95. [Google Scholar] [CrossRef]
- Cochaud, S.; Giustiniani, J.; Thomas, C.; Laprevotte, E.; Garbar, C.; Savoye, A.M.; Cure, H.; Mascaux, C.; Alberici, G.; Bonnefoy, N.; et al. Il-17a is produced by breast cancer tils and promotes chemoresistance and proliferation through erk1/2. Sci. Rep. 2013, 3, 3456. [Google Scholar] [CrossRef] [PubMed]
- Du, J.W.; Xu, K.Y.; Fang, L.Y.; Qi, X.L. Interleukin-17, produced by lymphocytes, promotes tumor growth and angiogenesis in a mouse model of breast cancer. Mol. Med. Rep. 2012, 6, 1099–1102. [Google Scholar] [CrossRef] [PubMed]
- Eiro, N.; Fernandez-Garcia, B.; Gonzalez, L.O.; Vizoso, F.J. Cytokines related to mmp-11 expression by inflammatory cells and breast cancer metastasis. Oncoimmunology 2013, 2, e24010. [Google Scholar] [CrossRef]
- Merrouche, Y.; Fabre, J.; Cure, H.; Garbar, C.; Fuselier, C.; Bastid, J.; Antonicelli, F.; Al-Daccak, R.; Bensussan, A.; Giustiniani, J. Il-17e synergizes with egf and confers in vitro resistance to egfr-targeted therapies in tnbc cells. Oncotarget 2016, 7, 53350–53361. [Google Scholar] [CrossRef]
- Nam, J.S.; Terabe, M.; Kang, M.J.; Chae, H.; Voong, N.; Yang, Y.A.; Laurence, A.; Michalowska, A.; Mamura, M.; Lonning, S.; et al. Transforming growth factor beta subverts the immune system into directly promoting tumor growth through interleukin-17. Cancer Res. 2008, 68, 3915–3923. [Google Scholar] [CrossRef]
- Roy, L.D.; Ghosh, S.; Pathangey, L.B.; Tinder, T.L.; Gruber, H.E.; Mukherjee, P. Collagen induced arthritis increases secondary metastasis in mmtv-pyv mt mouse model of mammary cancer. BMC Cancer 2011, 11, 365. [Google Scholar] [CrossRef] [PubMed]
- Roy, L.D.; Sahraei, M.; Schettini, J.L.; Gruber, H.E.; Besmer, D.M.; Mukherjee, P. Systemic neutralization of il-17a significantly reduces breast cancer associated metastasis in arthritic mice by reducing cxcl12/sdf-1 expression in the metastatic niches. BMC Cancer 2014, 14, 225. [Google Scholar] [CrossRef] [PubMed]
- Mombelli, S.; Cochaud, S.; Merrouche, Y.; Garbar, C.; Antonicelli, F.; Laprevotte, E.; Alberici, G.; Bonnefoy, N.; Eliaou, J.F.; Bastid, J.; et al. Il-17a and its homologs il-25/il-17e recruit the c-raf/s6 kinase pathway and the generation of pro-oncogenic lmw-e in breast cancer cells. Sci. Rep. 2015, 5, 11874. [Google Scholar] [CrossRef] [PubMed]
- Laprevotte, E.; Cochaud, S.; du Manoir, S.; Lapierre, M.; Dejou, C.; Philippe, M.; Giustiniani, J.; Frewer, K.A.; Sanders, A.J.; Jiang, W.G.; et al. The il-17b-il-17 receptor b pathway promotes resistance to paclitaxel in breast tumors through activation of the erk1/2 pathway. Oncotarget 2017, 8, 113360–113372. [Google Scholar] [CrossRef] [PubMed]
- Keyomarsi, K.; Tucker, S.L.; Buchholz, T.A.; Callister, M.; Ding, Y.; Hortobagyi, G.N.; Bedrosian, I.; Knickerbocker, C.; Toyofuku, W.; Lowe, M.; et al. Cyclin e and survival in patients with breast cancer. The N. Engl. J. Med. 2002, 347, 1566–1575. [Google Scholar] [CrossRef] [PubMed]
- Bonner, J.A.; Maihle, N.J.; Folven, B.R.; Christianson, T.J.; Spain, K. The interaction of epidermal growth factor and radiation in human head and neck squamous cell carcinoma cell lines with vastly different radiosensitivities. Int. J. Radiat. Oncol. Biol. Phys. 1994, 29, 243–247. [Google Scholar] [CrossRef]
- Huang, C.K.; Yang, C.Y.; Jeng, Y.M.; Chen, C.L.; Wu, H.H.; Chang, Y.C.; Ma, C.; Kuo, W.H.; Chang, K.J.; Shew, J.Y.; et al. Autocrine/paracrine mechanism of interleukin-17b receptor promotes breast tumorigenesis through nf-kappab-mediated antiapoptotic pathway. Oncogene 2014, 33, 2968–2977. [Google Scholar] [CrossRef]
- Coffelt, S.B.; Kersten, K.; Doornebal, C.W.; Weiden, J.; Vrijland, K.; Hau, C.S.; Verstegen, N.J.; Ciampricotti, M.; Hawinkels, L.J.; Jonkers, J.; et al. Il-17-producing gammadelta t cells and neutrophils conspire to promote breast cancer metastasis. Nature 2015, 522, 345–348. [Google Scholar] [CrossRef]
- Jiang, Z.; Chen, J.; Du, X.; Cheng, H.; Wang, X.; Dong, C. Il-25 blockade inhibits metastasis in breast cancer. Protein Cell 2017, 8, 191–201. [Google Scholar] [CrossRef]
- Eiro, N.; Gonzalez, L.; Gonzalez, L.O.; Fernandez-Garcia, B.; Lamelas, M.L.; Marin, L.; Gonzalez-Reyes, S.; del Casar, J.M.; Vizoso, F.J. Relationship between the inflammatory molecular profile of breast carcinomas and distant metastasis development. PLoS ONE 2012, 7, e49047. [Google Scholar] [CrossRef]
- Benatar, T.; Cao, M.Y.; Lee, Y.; Lightfoot, J.; Feng, N.; Gu, X.; Lee, V.; Jin, H.; Wang, M.; Wright, J.A.; et al. Il-17e, a proinflammatory cytokine, has antitumor efficacy against several tumor types in vivo. Cancer Immunol. Immunother. 2010, 59, 805–817. [Google Scholar] [CrossRef] [PubMed]
- Furuta, S.; Jeng, Y.M.; Zhou, L.; Huang, L.; Kuhn, I.; Bissell, M.J.; Lee, W.H. Il-25 causes apoptosis of il-25r-expressing breast cancer cells without toxicity to nonmalignant cells. Sci. Transl. Med. 2011, 3, 78ra31. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Huang, W.; Kong, D. Il-17 inhibits the accumulation of myeloid-derived suppressor cells in breast cancer via activating stat3. Int. Immunopharmacol. 2018, 59, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Nam, J.S.; Terabe, M.; Mamura, M.; Kang, M.J.; Chae, H.; Stuelten, C.; Kohn, E.; Tang, B.; Sabzevari, H.; Anver, M.R.; et al. An anti-transforming growth factor beta antibody suppresses metastasis via cooperative effects on multiple cell compartments. Cancer Res. 2008, 68, 3835–3843. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Kang, H.; Fung, A.; Zhao, H.; Wang, T.; Ma, D. The role of interleukin 17 in tumour proliferation, angiogenesis, and metastasis. Mediat. Inflamm. 2014, 2014, 623759. [Google Scholar] [CrossRef] [PubMed]
- Qian, X.; Chen, H.; Wu, X.; Hu, L.; Huang, Q.; Jin, Y. Interleukin-17 acts as double-edged sword in anti-tumor immunity and tumorigenesis. Cytokine 2015, 89, 34–44. [Google Scholar] [CrossRef]
- Honorati, M.C.; Neri, S.; Cattini, L.; Facchini, A. Il-17 enhances the susceptibility of u-2 os osteosarcoma cells to nk cell lysis. Clin. Exp. Immunol. 2003, 133, 344–349. [Google Scholar] [CrossRef]
- O’Sullivan, T.; Saddawi-Konefka, R.; Gross, E.; Tran, M.; Mayfield, S.P.; Ikeda, H.; Bui, J.D. Interleukin-17d mediates tumor rejection through recruitment of natural killer cells. Cell Rep. 2014, 7, 989–998. [Google Scholar] [CrossRef]
- Parajuli, P.; Mittal, S. Role of il-17 in glioma progression. J. Spine Neurosurg. 2013, (Suppl. 1), S1–S4. [Google Scholar] [CrossRef]
- Wu, F.; Xu, J.; Huang, Q.; Han, J.; Duan, L.; Fan, J.; Lv, Z.; Guo, M.; Hu, G.; Chen, L.; et al. The role of interleukin-17 in lung cancer. Mediat. Inflamm. 2016, 2016, 8494079. [Google Scholar] [CrossRef]
- Gu, K.; Li, M.M.; Shen, J.; Liu, F.; Cao, J.Y.; Jin, S.; Yu, Y. Interleukin-17-induced emt promotes lung cancer cell migration and invasion via nf-kappab/zeb1 signal pathway. Am. J. Cancer Res. 2015, 5, 1169–1179. [Google Scholar] [PubMed]
- Feng, M.; Wang, Y.; Chen, K.; Bian, Z.; Jinfang, W.; Gao, Q. Il-17a promotes the migration and invasiveness of cervical cancer cells by coordinately activating mmps expression via the p38/nf-kappab signal pathway. PLoS ONE 2014, 9, e108502. [Google Scholar] [CrossRef] [PubMed]
- Tartour, E.; Fossiez, F.; Joyeux, I.; Galinha, A.; Gey, A.; Claret, E.; Sastre-Garau, X.; Couturier, J.; Mosseri, V.; Vives, V.; et al. Interleukin 17, a t-cell-derived cytokine, promotes tumorigenicity of human cervical tumors in nude mice. Cancer Res. 1999, 59, 3698–3704. [Google Scholar] [PubMed]
- Pan, B.; Shen, J.; Cao, J.; Zhou, Y.; Shang, L.; Jin, S.; Cao, S.; Che, D.; Liu, F.; Yu, Y. Interleukin-17 promotes angiogenesis by stimulating vegf production of cancer cells via the stat3/giv signaling pathway in non-small-cell lung cancer. Sci. Rep. 2015, 5, 16053. [Google Scholar] [CrossRef]
- Proverbs-Singh, T.; Feldman, J.L.; Morris, M.J.; Autio, K.A.; Traina, T.A. Targeting the androgen receptor in prostate and breast cancer: Several new agents in development. Endocr. Relat. Cancer 2015, 22, R87–R106. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Liu, S.; Ge, D.; Zhang, Q.; Xue, Y.; Xiong, Z.; Abdel-Mageed, A.B.; Myers, L.; Hill, S.M.; Rowan, B.G.; et al. Interleukin-17 promotes formation and growth of prostate adenocarcinoma in mouse models. Cancer Res. 2012, 72, 2589–2599. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Liu, S.; Parajuli, K.R.; Zhang, W.; Zhang, K.; Mo, Z.; Liu, J.; Chen, Z.; Yang, S.; Wang, A.R.; et al. Interleukin-17 promotes prostate cancer via mmp7-induced epithelial-to-mesenchymal transition. Oncogene 2016, 36, 687–699. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zhang, Q.; Liu, S.; Parajuli, K.R.; Qu, Y.; Mei, J.; Chen, Z.; Zhang, H.; Khismatullin, D.B.; You, Z. Il-17 and insulin/igf1 enhance adhesion of prostate cancer cells to vascular endothelial cells through cd44-vcam-1 interaction. Prostate 2015, 75, 883–895. [Google Scholar] [CrossRef] [PubMed]
- Welte, T.; Zhang, X.H. Interleukin-17 could promote breast cancer progression at several stages of the disease. Mediat. Inflamm. 2015, 2015, 804347. [Google Scholar] [CrossRef]
- Loget, J.; Plee, J.; Antonicelli, F.; Bernard, P. A successful treatment with ustekinumab in a case of relapsing bullous pemphigoid associated with psoriasis. J. Eur. Acad. Dermatol. Venereol. 2017, 31, e228–e230. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Type of IL-17 | Type of Study | Breast Cancer Cell Lines | Effect of IL-17 Exposure | Murine Model | Mechanism | Reference |
|---|---|---|---|---|---|---|
| IL-17A | Clinical and preclinical | No | Anti | BALB/c Nude | Induction of differentiation and apoptosis, inhibition of proliferation of MDSC via STAT3 | [103] |
| IL-17A | Preclinical | No | Pro | KEP, Tcrδ−/− | Production of IL-17A by γδ T cells induces neutrophils to suppress CD8+ T Cells and promotes distant metastases. | [98] |
| IL-17A | Clinical and preclinical | MCF7, T47D, BT20, MDA-MB468, MD-MB157, MDA-MB231 | Pro | No | Activation or ERK1/2 pathway induces proliferation, migration, invasion and chemoresistance | [86] |
| IL-17A | Clinical | No | Pro | No | IL-17A associated to MMP-11+ mononuclear infiltrating cells which are correlated to metastasis | [88,100] |
| IL-17A | Preclinical | MCF7 | Pro | No | Activation of MAPK: MEKK, ERK, JNK, cJun, STAT3.Cell proliferation. | [35] |
| IL-17A | Preclinical | MA782, 4T1 | Pro | BALB/c | Increase in tumor volume and microvascular density | [87] |
| IL-17A | Preclinical | 4T1, PyV MT cell line | Pro | PyV MT, arthritic PyV MT | Upregulation of SDF1, IL-6, G-CSF. Promotion of bone and lung metastases | [91,92] |
| IL-17A | Clinical and preclinical | MCF-7, T47D, MDA-MB435, MDA-MB231 | Pro | No | Recruitment of macrophages, activation of MMP | [85] |
| IL-17A | Preclinical | 4T1, MDA-MB231, EM6, MDA-MB435, Hs578t | Pro | BALB/c | TGF- β dependent tumor growth, inhibition of apoptosis | [104] |
| IL-17E | Clinical and preclinical | No | Pro | MMTV-PyMT | Production of IL-17E by tumor- infiltrating macrophages | [99] |
| IL-17E | Preclinical | MCF7, MDA-MB468, MDA-MB 435-S, MDA-MB231,SKBR3, T47D, ZR75, Hs578t, HCC1937, MDA-MB175-7 | Anti | Nude | Induction of apoptosis, decrease in colony formation and tumor growth | [102] |
| IL-17E | Preclinical | MDA-MB-435 | Anti | CD1-nude | Decrease in tumor volume. B cells mandatory | [101] |
| IL-17A and Il-17E | Preclinical | MDA-MB468, BT20, IJG-1731 | Pro | No | Activation of STAT3, PYK-2, Src and HER-1. Nuclear translocation of pSTAT3 and pHER-1. Resistance to TKI. | [89] |
| IL-17A and IL-17E | Clinical and preclinical | T47D, MCF7, BT20, IJG-1731 | Pro | No | Activation of cRAF and S6 kinases. Chemoresistance and generation of LMWCE | [93] |
| IL-17B | Clinical and preclinical | BT20, MDA-MB-468, MCF7 | Pro | Nude | Resistance to paclitaxel in cell lines and xenografts via ERK pathway. Upregulation of BCL2. | [94] |
| IL-17B | Clinical and preclinical | MCF7, MDA-MB-157, MDA-MB-231, MDA-MB-361, MDA-MB-468, SKBR3, SKBR3-hr | Pro | NOD/SCID/γnull | Promotion of proliferation and tumor growth through IL-17RB via NF-kB and TRAF6 | [97] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fabre, J.A.S.; Giustiniani, J.; Garbar, C.; Merrouche, Y.; Antonicelli, F.; Bensussan, A. The Interleukin-17 Family of Cytokines in Breast Cancer. Int. J. Mol. Sci. 2018, 19, 3880. https://doi.org/10.3390/ijms19123880
Fabre JAS, Giustiniani J, Garbar C, Merrouche Y, Antonicelli F, Bensussan A. The Interleukin-17 Family of Cytokines in Breast Cancer. International Journal of Molecular Sciences. 2018; 19(12):3880. https://doi.org/10.3390/ijms19123880
Chicago/Turabian StyleFabre, Joseph Antoine Salvator, Jérôme Giustiniani, Christian Garbar, Yacine Merrouche, Frank Antonicelli, and Armand Bensussan. 2018. "The Interleukin-17 Family of Cytokines in Breast Cancer" International Journal of Molecular Sciences 19, no. 12: 3880. https://doi.org/10.3390/ijms19123880
APA StyleFabre, J. A. S., Giustiniani, J., Garbar, C., Merrouche, Y., Antonicelli, F., & Bensussan, A. (2018). The Interleukin-17 Family of Cytokines in Breast Cancer. International Journal of Molecular Sciences, 19(12), 3880. https://doi.org/10.3390/ijms19123880