Biological Potential and Mechanism of Prodigiosin from Serratia marcescens Subsp. lawsoniana in Human Choriocarcinoma and Prostate Cancer Cell Lines

Abstract

1. Introduction
2. Results
2.1. Identification of the Prodigiosin-Producing Strain HDZK-BYSB107
2.1.1. Morphological Characterization
2.1.2. Physiological and Biochemical Characteristics of Strain HDZK-BYSB107
2.1.3. Molecular Analysis
2.2. Isolation, Purification, and Identification of Prodigiosin from Strain HDZK-BYSB107
2.3. Physical and Chemical Properties of Prodigiosin from Strain HDZK-BYSB107
2.4. Activity Assays on the Prodigiosin from Strain HDZK-BYSB107
2.4.1. Antimicrobial Activity Assay
2.4.2. In Vitro Apoptotic Activity Assay
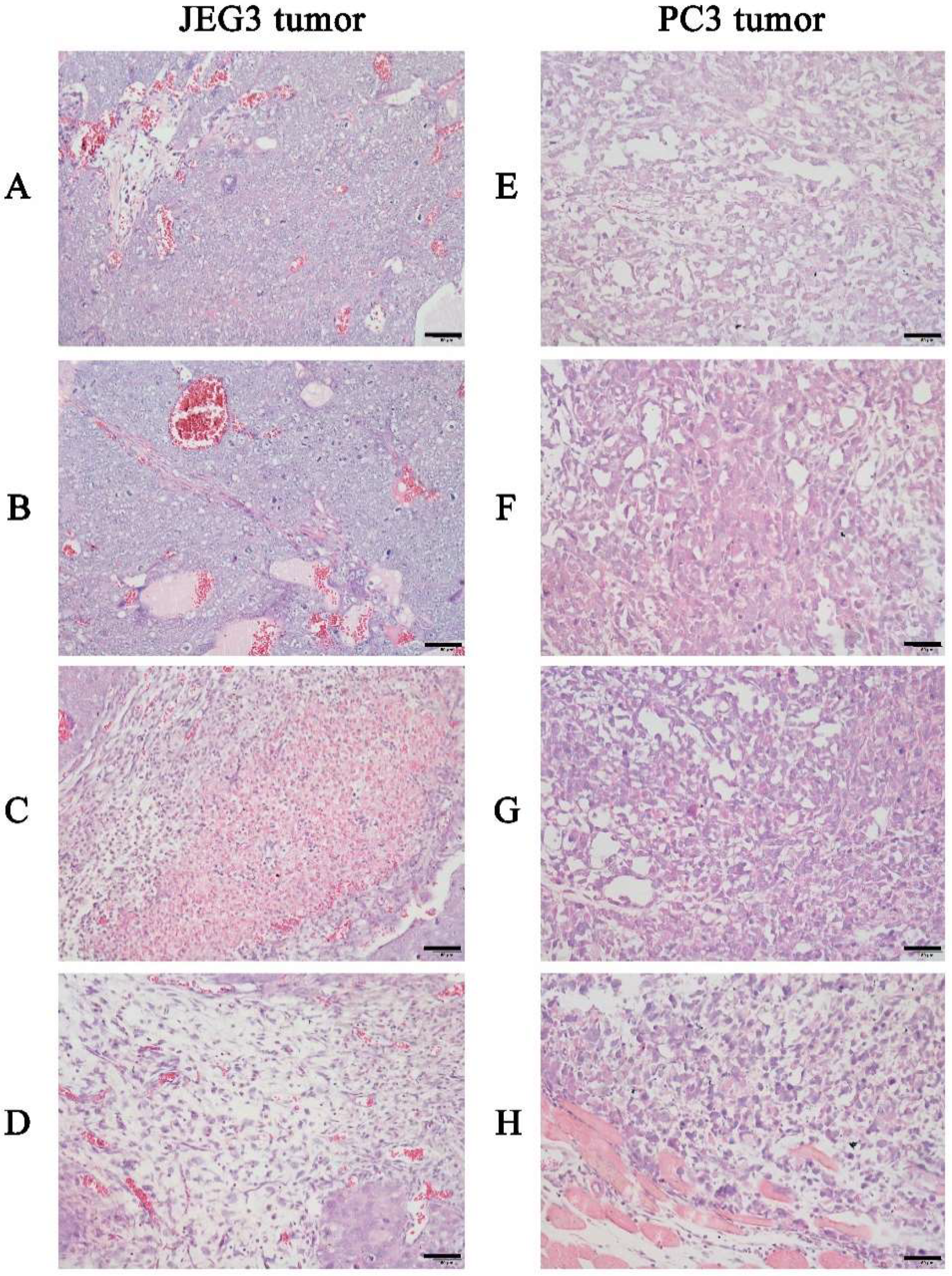
2.4.3. In Vivo Anticancer Activity Assay
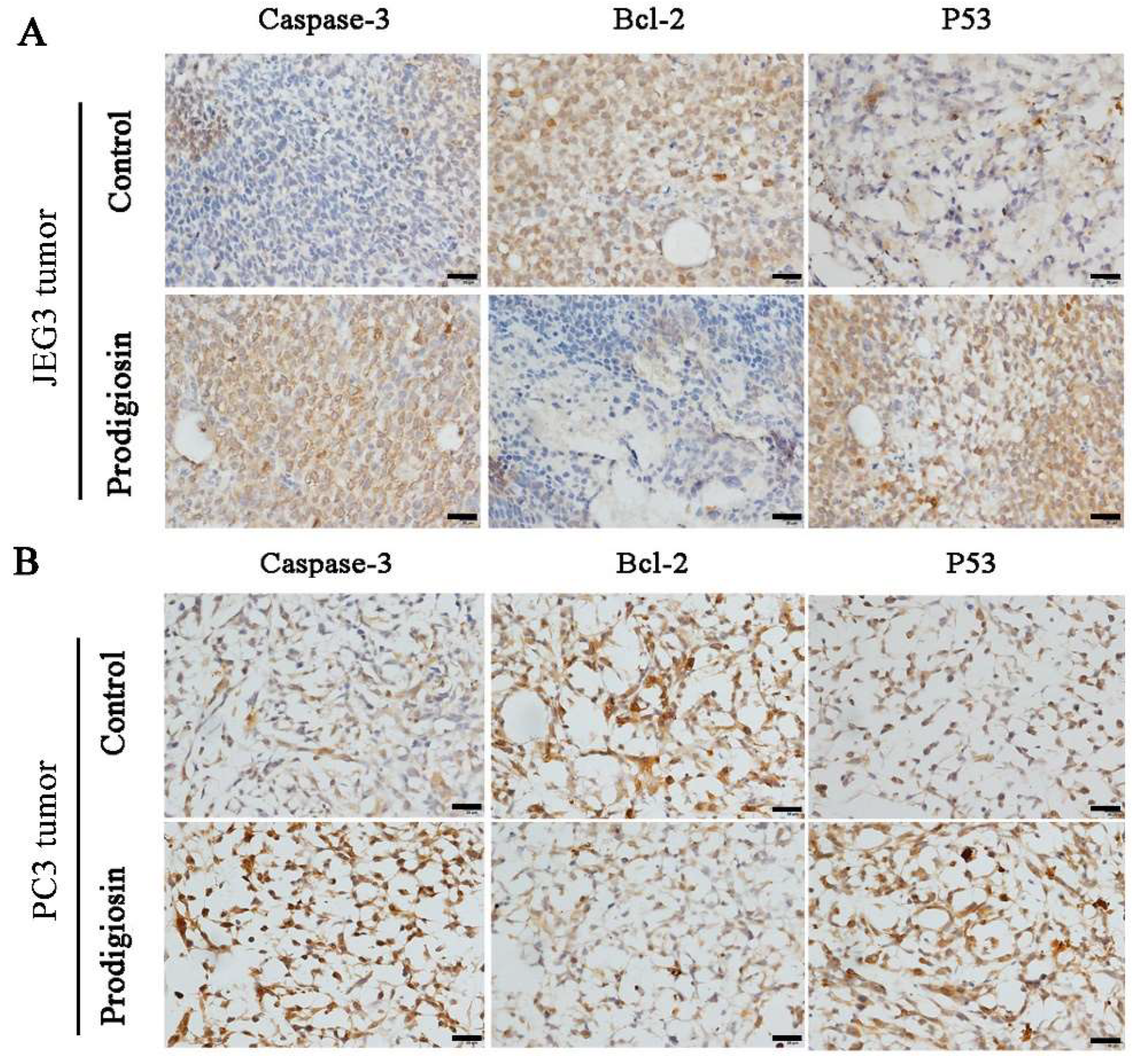
2.5. Anticancer Mechanism of the Prodigiosin from Strain HDZK-BYSB107
3. Discussion
4. Materials and Methods
4.1. Strains and Cells
4.2. Isolation and Culture of Strain HDZK-BYSB107
4.3. Identification of Strain HDZK-BYSB107
4.3.1. Morphological Examination of Strain HDZK-BYSB107
4.3.2. Physiological and Biochemical Characteristics of Strain HDZK-BYSB107
4.3.3. Molecular Analysis of Strain HDZK-BYSB107
4.4. Preparation and Isolation of Fermentation broth from Strain HDZK-BYSB107
4.5. Isolation, Purification, and Identification of Prodigiosin from Strain HDZK-BYSB107
4.6. Activity Assays on Prodigiosin from Strain HDZK-BYSB107
4.6.1. Antimicrobial Activity Assay
4.6.2. In Vitro Anticancer Activity Assay
4.6.3. In Vivo Anticancer Activity Assay
4.7. TUNEL Assay
4.8. Anticancer Mechanism of the Prodigiosin from Strain HDZK-BYSB107
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Orlikova, B.; Legrand, N.; Panning, J.; Dicato, M.; Diederich, M. Anti-inflammatory and anticancer drugs from nature. Cancer Res. Treat. 2014, 159, 123–143. [Google Scholar]
- Ratcliffe, N.A.; Mello, C.B.; Garcia, E.S.; Butt, T.M.; Azambuja, P. Insect natural products and processes: New treatments for human disease. Insect Biochem. Mol. Boil. 2011, 41, 747–769. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.K.; Khuda-Bukhsh, A.R. Molecular approaches towards development of purified natural products and their structurally known derivatives as efficient anti-cancer drugs: Current trends. Eur. J. Pharmacol. 2013, 714, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Cragg, G.M.; Grothaus, P.G.; Newman, D.J. Impact of natural products on developing new anti-cancer agents. Chem. Rev. 2009, 109, 3012–3043. [Google Scholar] [CrossRef] [PubMed]
- Han, S.B.; Kim, H.M.; Kim, Y.H.; Lee, C.W.; Jang, E.S.; Son, K.H.; Kim, S.U.; Kim, Y.K. T-cell specific immunosuppression by prodigiosin isolated from Serratia marcescens. Int. J. Immunopharmacol. 1998, 20, 1–13. [Google Scholar] [CrossRef]
- Zhang, M.M.; Qiao, Y.; Ang, E.L.; Zhao, H.M. Using natural products for drug discovery: The impact of the genomics era. Expert Opin. Drug Dis. 2017, 12, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Aydemir, G.; Kasiri, Y.; Birta, E.; Béke, G.; Garcia, A.L.; Bartók, E.M.; Rühl, R. Lycopene-derived bioactive retinoic acid receptors/retinoid-X receptors-activating metabolites may be relevant for lycopene’s anti-cancer potential. Mol. Nutr. Food Res. 2013, 57, 739–747. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.J.; Bao, J.L.; Wu, G.S.; Xu, W.S.; Huang, M.Q.; Chen, X.P.; Wang, Y.T. Quinones derived from plant secondary metabolites as anti-cancer agents. Anti-Cancer Agents Med. Chem. 2013, 13, 456–463. [Google Scholar]
- Rawat, D.S.; Singh, R. Plant derived secondary metabolites as anti-cancer agents. Anti-Cancer Agents Med. Chem. 2013, 13, 1551. [Google Scholar] [CrossRef]
- Samaan, N.; Zhong, Q.; Fernandez, J.; Chen, G.; Hussain, A.M.; Zheng, S.L.; Wang, G.; Chen, Q.H. Design, synthesis, and evaluation of novel heteroaromatic analogs of curcumin as anti-cancer agents. Eur. J. Med. Chem. 2014, 75, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Bernardes, N.; Chakrabarty, A.M.; Fialho, A.M. Engineering of bacterial strains and their products for cancer therapy. Appl. Microbiol. Biotechnol. 2013, 97, 5189–5199. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.G.; Kaufmann, S.H. Targeting bacteria to improve cancer therapy. Science 2010, 330, 766–767. [Google Scholar] [CrossRef] [PubMed]
- Mathuriya, A.S. Magnetotactic bacteria for cancer therapy. Biotechnol. Lett. 2015, 37, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.H.; Lin, S.C.; Yang, S.Y.; Pan, M.Y.; Lin, Y.W.; Hsu, C.Y.; Wei, Y.H.; Chang, J.S.; Chang, C.C. Prodigiosin-induced cytotoxicity involves RAD51 down-regulation through the JNK and p38 MAPK pathways in human breast carcinoma cell lines. Toxicol. Lett. 2012, 212, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, H.Y.; Shieh, J.J.; Chen, C.J.; Pan, M.Y.; Yang, S.Y.; Lin, S.C.; Chang, J.S.; Lee, A.Y.; Chang, C.C. Prodigiosin down-regulates SKP2 to induce p27(KIP1) stabilization and antiproliferation in human lung adenocarcinoma cells. Br. J. Pharmacol. 2012, 166, 2095–20108. [Google Scholar] [CrossRef] [PubMed]
- Elahian, F.; Moghimi, B.; Dinmohammadi, F.; Ghamghami, M.; Hamidi, M.; Mirzaei, S.A. The anticancer agent prodigiosin is not a multidrug resistance protein substrate. DNA Cell Biol. 2013, 32, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Suryawanshi, R.K.; Patil, C.D.; Borase, H.P.; Salunke, B.K.; Patil, S.V. Studies on production and biological potential of prodigiosin by Serratia marcescens. Appl. Biochem. Biotechnol. 2014, 173, 1209–1221. [Google Scholar] [CrossRef] [PubMed]
- Suryawanshi, R.K.; Patil, C.D.; Borase, H.P.; Narkhede, C.P.; Stevenson, A.; Hallsworth, J.E.; Patil, S.V. Towards an understanding of bacterial metabolites prodigiosin and violacein and their potential for use in commercial sunscreens. Int. J. Cosmet. Sci. 2015, 37, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Wang, Y.Y.; Qi, X.; Gu, Q.; Geng, M.; Li, J. Undecylprodigiosin induced apoptosis in P388 cancer cells is associated with its binding to ribosome. PLoS ONE 2013, 8, e65381. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, B.; Zhou, L.; Yu, S.; Su, Z.; Song, J.; Sun, Q.; Sha, O.; Wang, X.; Jiang, W.; et al. Prodigiosin inhibits Wnt/β-catenin signaling and exerts anticancer activity in breast cancer cells. Proc. Natl. Acad. Sci. USA 2016, 113, 13150–13155. [Google Scholar] [CrossRef] [PubMed]
- Hong, B.; Prabhu, V.V.; Zhang, S.; van den Heuvel, A.P.; Dicker, D.T.; Kopelovich, L.; El-Deiry, W.S. Prodigiosin rescues deficient p53 signaling and antitumor effects via upregulating p73 and disrupting its interaction with mutant p53. Cancer Res. 2014, 74, 1153–1165. [Google Scholar] [CrossRef] [PubMed]
- Chiu, W.J.; Lin, S.R.; Chen, Y.H.; Tsai, M.J.; Leong, M.K.; Weng, C.F. Prodigiosin-Emerged PI3K/Beclin-1-Independent Pathway Elicits Autophagic Cell Death in Doxorubicin-Sensitive and -Resistant Lung Cancer. J. Clin. Med. 2018, 7, 321. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, T.; Sakurai, F.; Hayakawa, Y. A prodigiosin from the roseophilin producer Streptomyces griseoviridis. J. Nat. Prod. 2008, 71, 1265–1267. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Liu, G.; Li, J.; Huang, H.; Zhang, X.; Zhang, H.; Ju, J. Cytotoxic and antibacterial angucycline- and prodigiosin-analogues from the deep-sea derived Streptomyces sp. SCSIO 11594. Mar. Drugs 2015, 13, 1304–1316. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Lee, J.S.; Park, Y.K.; Kim, J.F.; Jeong, H.; Oh, T.K.; Kim, B.S.; Lee, C.H. Biosynthesis of antibiotic prodiginines in the marine bacterium Hahella chejuensis KCTC 2396. J. Appl. Microbiol. 2007, 102, 937–944. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Ran, T.; Shen, X.; Xu, L.; Wang, W.; Xu, D. Expression, crystallization and preliminary crystallographic data analysis of PigF, an O-methyltransferase from the prodigiosin-synthetic pathway in Serratia. Acta Crystallogr. F 2012, 68, 898–901. [Google Scholar] [CrossRef] [PubMed]
- Salem, S.M.; Kancharla, P.; Florova, G.; Gupta, S.; Lu, W.; Reynolds, K.A. Elucidation of final steps of the marineosins biosynthetic pathway through identification and characterization of the corresponding gene cluster. J. Am. Chem. Soc. 2014, 136, 4565–4574. [Google Scholar] [CrossRef] [PubMed]
- Dalili, D.; Fouladdel, S.; Rastkari, N.; Samadi, N.; Ahmadkhaniha, R.; Ardavan, A.; Azizi, E. Prodigiosin, the red pigment of Serratia marcescens, shows cytotoxic effects and apoptosis induction in HT-29 and T47D cancer cell lines. Nat. Prod. Res. 2011, 26, 2078–2083. [Google Scholar] [PubMed]
- Ho, T.F.; Ma, C.J.; Lu, C.H.; Tsai, Y.T.; Wei, Y.H.; Chang, J.S.; Lai, J.K.; Cheuh, P.J.; Yeh, C.T.; Tang, P.C.; et al. Undecylprodigiosin selectively induces apoptosis in human breast carcinoma cells independent of p53. Toxicol. Appl. Pharmacol. 2007, 225, 318–328. [Google Scholar] [CrossRef] [PubMed]
- Sam, M.R.; Pourpak, R.S. Regulation of p53 and survivin by prodigiosin compound derived from Serratia marcescens contribute to caspase-3-dependent apoptosis in acute lymphoblastic leukemia cells. Hum. Exp. Toxicol. 2018, 37, 608–617. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S. Caspase function in programmed cell death. Cell Death Differ. 2007, 14, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sun, H.; Zhao, H.L.; Chen, X.X.; Li, J.J.; Li, B.Q. Early apoptosis of monocytes induced by Helicobacter pylori infection through multiple pathways. Dev. Comp. Immunol. 2017, 73, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.O.; Park, C.; Hwang, H.J.; Hong, S.H.; Kim, G.Y.; Cho, E.J.; Kim, W.J.; Choi, Y.H. Baicalein induces apoptosis via ROS-dependent activation of caspases in human bladder cancer 5637 cells. Int. J. Oncol. 2016, 49, 1009–1018. [Google Scholar] [CrossRef] [PubMed]
- Pluta, P.; Jeziorski, A.; Pluta, A.; Cebula-Obrzut, B.; Wierzbowska, A.; Piekarski, J.; Smolewski, P. Expression of IAP family proteins and its clinical importance in breast cancer patients. Neoplasma 2015, 62, 666–673. [Google Scholar] [CrossRef] [PubMed]
- Tamanini, E.; Buck, I.M.; Chessari, G.; Chiarparin, E.; Day, J.E.; Frederickson, M.; Griffiths-Jones, C.M.; Hearn, K.; Heightman, T.D.; Iqbal, A.; et al. Discovery of a potent nonpeptidomimetic, small-molecule antagonist of cellular inhibitor of apoptosis protein 1 (cIAP1) and X-linked inhibitor of apoptosis protein (XIAP). J. Med. Chem. 2017, 60, 4611–4625. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Yu, H.W.; Yan, C.; Zhao, W.Y.; Liu, Y.; Zhang, D.D.; Li, J.; Liu, N. X-linked inhibitor of apoptosis inhibits apoptosis and preserves the blood-brain barrier after experimental subarachnoid hemorrhage. Sci. Rep. 2017, 7, e44918. [Google Scholar] [CrossRef] [PubMed]
- Derakhshan, A.; Chen, Z.; Van Waes, C. Therapeutic small molecules target inhibitor of apoptosis proteins in cancers with deregulation of extrinsic and intrinsic cell death pathways. Clin. Cancer Res. 2017, 23, 1379–1387. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Kaufmann, T.; Biswas, S. Impact of inhibitor of apoptosis proteins on immune modulation and inflammation. Immunol. Cell Biol. 2017, 95, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Ferraris, D.V. Evolution of poly(ADP-ribose) polymerase-1 (PARP-1) inhibitors. From concept to clinic. J. Med. Chem. 2010, 53, 4561–4584. [Google Scholar] [CrossRef] [PubMed]
- He, J.X.; Yang, C.H.; Miao, Z.H. Poly(ADP-ribose) polymerase inhibitors as promising cancer therapeutics. Acta Pharmacol. Sin. 2010, 31, 1172–1180. [Google Scholar] [CrossRef] [PubMed]
- Vyas, S.; Chang, P. New PARP targets for cancer therapy. Nat. Rev. Cancer 2014, 14, 502–509. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.Y.; Shen, Y.C.; Lu, C.H.; Yang, S.Y.; Ho, T.F.; Pen, Y.T.; Chang, C.C. Prodigiosin activates endoplasmic reticulum stress cell death pathway in human breast carcinoma cell lines. Toxicol. Appl. Pharmacol. 2012, 265, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Hwang, W.J.; Kartal, S.N.; Yoshimura, T.; Imamura, Y. Synergistic effect of heartwood extractives and quaternary ammonium compounds on termite resistance of treated wood. Pest Manag. Sci. 2007, 63, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.H.; Chen, C.H.; Chien, S.C.; Lin, Y.L. Five new cadinane-type sesquiterpenes from the heartwood of Chamaecyparis obtusa var. formosana. J. Nat. Prod. 2002, 65, 25–28. [Google Scholar] [CrossRef] [PubMed]
- Song, M.J.; Bae, J.; Lee, D.S.; Kim, C.K.; Kim, J.S.; Kim, S.W.; Hong, S.I. Purification and characterization of prodigiosin produced by integrated bioreactor from Serratia sp. KH-95. J. Biosci. Bioeng. 2006, 101, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Hong, K.L.; Joung, H.Y. Statistical optimization of medium components for the production of prodigiosin by Hahella chejuensis KCTC 2396. J. Microbiol. Biotechnol. 2008, 18, 1903–1907. [Google Scholar] [PubMed]
- Lee, J.S.; Kim, Y.S.; Park, S.; Kim, J.; Kang, S.J.; Lee, M.H.; Ryu, S.; Choi, J.M.; Oh, T.K.; Yoon, J.H. Exceptional production of both prodigiosin and cycloprodigiosin as major metabolic constituents by a novel marine bacterium, Zooshikella rubidus S1-1. Appl. Environ. Microbiol. 2011, 77, 4967–4973. [Google Scholar] [CrossRef] [PubMed]
- Stankovic, N.; Senerovic, L.; Ilic-Tomic, T.; Vasiljevic, B.; Nikodinovic-Runic, J. Properties and applications of undecyl-prodigiosin and other bacterial prodigiosins. Appl. Microbiol. Biotechnol. 2014, 98, 3841–3858. [Google Scholar] [CrossRef] [PubMed]
- Francisco, R.; Perez-Tomas, R.; Gimenez-Bonafe, P.; Soto-Cerrato, V.; Gimenez-Xavier, P.; Ambrosio, S. Mechanisms of prodigiosin cytotoxicity in human neuroblastoma cell lines. Eur. J. Pharmacol. 2007, 572, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Pandey, R.; Chander, R.; Sainis, K.B. Prodigiosins as anti-cancer agents: Living upto their name. Curr. Pharm. Des. 2009, 15, 732–741. [Google Scholar] [CrossRef] [PubMed]
- Montaner, B.; Perez-Tomas, R. Prodigiosin induces caspase-9 and caspase-8 activation and cytochrome C release in Jurkat T cells. Ann. N. Y. Acad. Sci. 2002, 973, 246–249. [Google Scholar] [CrossRef] [PubMed]
- Perez-Tomas, R.; Montaner, B.; Llagostera, E.; Soto-Cerrato, V. The prodigiosins, proapoptotic drugs with anticancer properties. Biochem. Pharmacol. 2003, 66, 1447–1452. [Google Scholar] [CrossRef]
- Hu, Y.; Benedict, M.A.; Ding, L.; Núñez, G. Role of cytochrome c and dATP/ATP hydrolysis in Apaf-1-mediated caspase-9 acti-vation and apoptosis. EMBO J. 1999, 18, 3586–3595. [Google Scholar] [CrossRef] [PubMed]
- Thornberry, N.A.; Lazebnik, Y. Caspases: Enemies within. Science 1998, 281, 1312–1316. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.M.; Cory, S. Bcl-2-regulated apoptosis: Mechanism and therapeutic potential. Curr. Opin. Immunol. 2007, 19, 488–496. [Google Scholar] [CrossRef] [PubMed]
- Hyun, H.B.; Lee, W.S.; Go, S.I.; Nagappan, A.; Park, C.; Han, M.H.; Hong, S.H.; Kim, G.; Kim, G.Y.; Cheong, J.; et al. The flavonoid morin from moraceae induces apoptosis by modulation of Bcl-2 family members and Fas receptor in HCT 116 cells. Int. J. Oncol. 2015, 46, 2670–2678. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.C.; Wang, H.Z.; Zhang, B.X.; Chen, Y.M.; Zhang, Y.M.; Sun, X.; Xiao, G.; Nan, K.; Ren, H.; Qin, S. LCL161 increases paclitaxel-induced apoptosis by degrading cIAP1 and cIAP2 in NSCLC. J. Exp. Clin. Cancer Res. 2016, 35, e158. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.L.; Wang, M. Expression levels of survivin, Bcl-2, and KAI1 proteins in cervical cancer and their correlation with metastasis. Genet. Mol. Res. 2015, 14, 17059–17067. [Google Scholar] [CrossRef] [PubMed]
- Niemhom, N.; Chutrakul, C.; Suriyachadkun, C.; Thawai, C. Phytohabitans kaempferiae sp. nov., an endophytic actinomycete isolated from the leaf of Kaempferia larsenii. Int. J. Syst. Evol. Microboil. 2016, 66, 2917–2922. [Google Scholar] [CrossRef] [PubMed]
- Sacco, N.J.; Bonetto, M.C.; Corton, E. Isolation and characterization of a novel electrogenic bacterium, Dietzia sp. RNV-4. PLoS ONE 2017, 12, e0169955. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.; Liu, L.; Huang, Y.; Zhang, X.; Wang, Z. Production of Monascus pigments as extracellular crystals by cell suspension culture. Appl. Microbiol. Biotechnol. 2018, 102, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Shi, T.; Xu, M.; Brock, N.L.; Zhao, Y.L.; Wang, Y.; Deng, Z.; Pang, X.; Tao, M. Hybrubins: Bipyrrole Tetramic Acids Obtained by Crosstalk between a Truncated Undecylprodigiosin Pathway and Heterologous Tetramic Acid Biosynthetic Genes. Org. Lett. 2016, 18, 572–575. [Google Scholar] [CrossRef] [PubMed]
- Stankovic, N.; Radulovic, V.; Petkovic, M.; Vuckovic, I.; Jadranin, M.; Vasiljevic, B.; Nikodinovic-Runic, J. Streptomyces sp. JS520 produces exceptionally high quantities of undecylprodigiosin with antibacterial, antioxidative, and UV-protective properties. Appl. Microbiol. Biotechnol. 2012, 96, 1217–1231. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Yang, M.; Wang, M.; Liu, X.; Xu, H.; Xu, X.; Sun, G.; Sun, X. Total Saponins of Aralia Elata (Miq) Seem Alleviate Calcium Homeostasis Imbalance and Endoplasmic Reticulum Stress-Related Apoptosis Induced by Myocardial Ischemia/Reperfusion Injury. Cell. Physiol. Biochem. 2018, 50, 28–40. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Dosage of Prodigiosin (μg/kg) | Tumor Volume before Treatment (cm3) | Tumor Volume after 2 d Treatment (cm3) | Tumor Volume after 9 d Treatment (cm3) | Tumor Volume after 16 d Treatment (cm3) |
|---|---|---|---|---|---|
| JEG3 cells | 0 | 0.207 ± 0.031 | 0.225 ± 0.014 | 0.253 ± 0.017 | 0.281 ± 0.022 |
| PC3 cells | 0.212 ± 0.132 | 0.237 ± 0.168 | 0.261 ± 0.052 | 0.295 ± 0.085 | |
| JEG3 cells | 50 | 0.206 ± 0.142 | 0.225 ± 0.012 | 0.252 ± 0.005 | 0.279 ± 0.013 |
| PC3 cells | 0.212 ± 0.174 | 0.236 ± 0.053 | 0.259 ± 0.031 | 0.294 ± 0.047 | |
| JEG3 cells | 100 | 0.202 ± 0.016 a | 0.205 ± 0.043 a | 0.192 ± 0.192 a | 0.180 ± 0.142 a |
| PC3 cells | 0.216 ± 0.194 a | 0.208 ± 0.018 a | 0.195 ± 0.087 a | 0.182 ± 0.104 a | |
| JEG3 cells | 250 | 0.211 ± 0.067 a | 0.169 ± 0.069 b | 0.152 ± 0.172 b | 0.144 ± 0.067 b |
| PC3 cells | 0.209 ± 0.037 a | 0.163 ± 0.041 b | 0.155 ± 0.054 b | 0.149 ± 0.127 b |
| Groups | Dosage of Prodigiosin (μg/kg) | Tumor Weight (g) | Tumor Inhibition Rate (%) |
|---|---|---|---|
| JEG3 cells | 0 | 0.85 ± 0.13 | |
| PC3 cells | 0.87 ± 0.21 | ||
| JEG3 cells | 50 | 0.84 ± 0.34 | 1.17 ± 0.19 |
| PC3 cells | 0.86 ± 0.02 | 1.14 ± 0.25 | |
| JEG3 cells | 100 | 0.77 ± 0.01* | 9.41 ± 0.14* |
| PC3 cells | 0.81 ± 0.03* | 6.80 ± 0.32* | |
| JEG3 cells | 250 | 0.64 ± 0.05** | 24.71 ± 0.29** |
| PC3 cells | 0.69 ± 0.13** | 20.69 ± 0.17** |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, D.; Liu, J.; Wang, X.; Kong, D.; Du, W.; Li, H.; Hse, C.-Y.; Shupe, T.; Zhou, D.; Zhao, K. Biological Potential and Mechanism of Prodigiosin from Serratia marcescens Subsp. lawsoniana in Human Choriocarcinoma and Prostate Cancer Cell Lines. Int. J. Mol. Sci. 2018, 19, 3465. https://doi.org/10.3390/ijms19113465
Li D, Liu J, Wang X, Kong D, Du W, Li H, Hse C-Y, Shupe T, Zhou D, Zhao K. Biological Potential and Mechanism of Prodigiosin from Serratia marcescens Subsp. lawsoniana in Human Choriocarcinoma and Prostate Cancer Cell Lines. International Journal of Molecular Sciences. 2018; 19(11):3465. https://doi.org/10.3390/ijms19113465
Chicago/Turabian StyleLi, Dan, Jun Liu, Xin Wang, Di Kong, Wei Du, Hongbo Li, Chung-Yun Hse, Todd Shupe, Dongpo Zhou, and Kai Zhao. 2018. "Biological Potential and Mechanism of Prodigiosin from Serratia marcescens Subsp. lawsoniana in Human Choriocarcinoma and Prostate Cancer Cell Lines" International Journal of Molecular Sciences 19, no. 11: 3465. https://doi.org/10.3390/ijms19113465
APA StyleLi, D., Liu, J., Wang, X., Kong, D., Du, W., Li, H., Hse, C.-Y., Shupe, T., Zhou, D., & Zhao, K. (2018). Biological Potential and Mechanism of Prodigiosin from Serratia marcescens Subsp. lawsoniana in Human Choriocarcinoma and Prostate Cancer Cell Lines. International Journal of Molecular Sciences, 19(11), 3465. https://doi.org/10.3390/ijms19113465