BDNF, Brain, and Regeneration: Insights from Zebrafish

 ,
,  ,
, {kind=link}
{kind=link}
Abstract
:1. The Brain of the Zebrafish Is a Model for Neuroscientific Studies
2. The Evolution of the Brain Derived Neurotrophic Factor (BDNF) Gene and Protein
3. Different Receptors Mediate the Multiple Roles of BDNF in the Brain
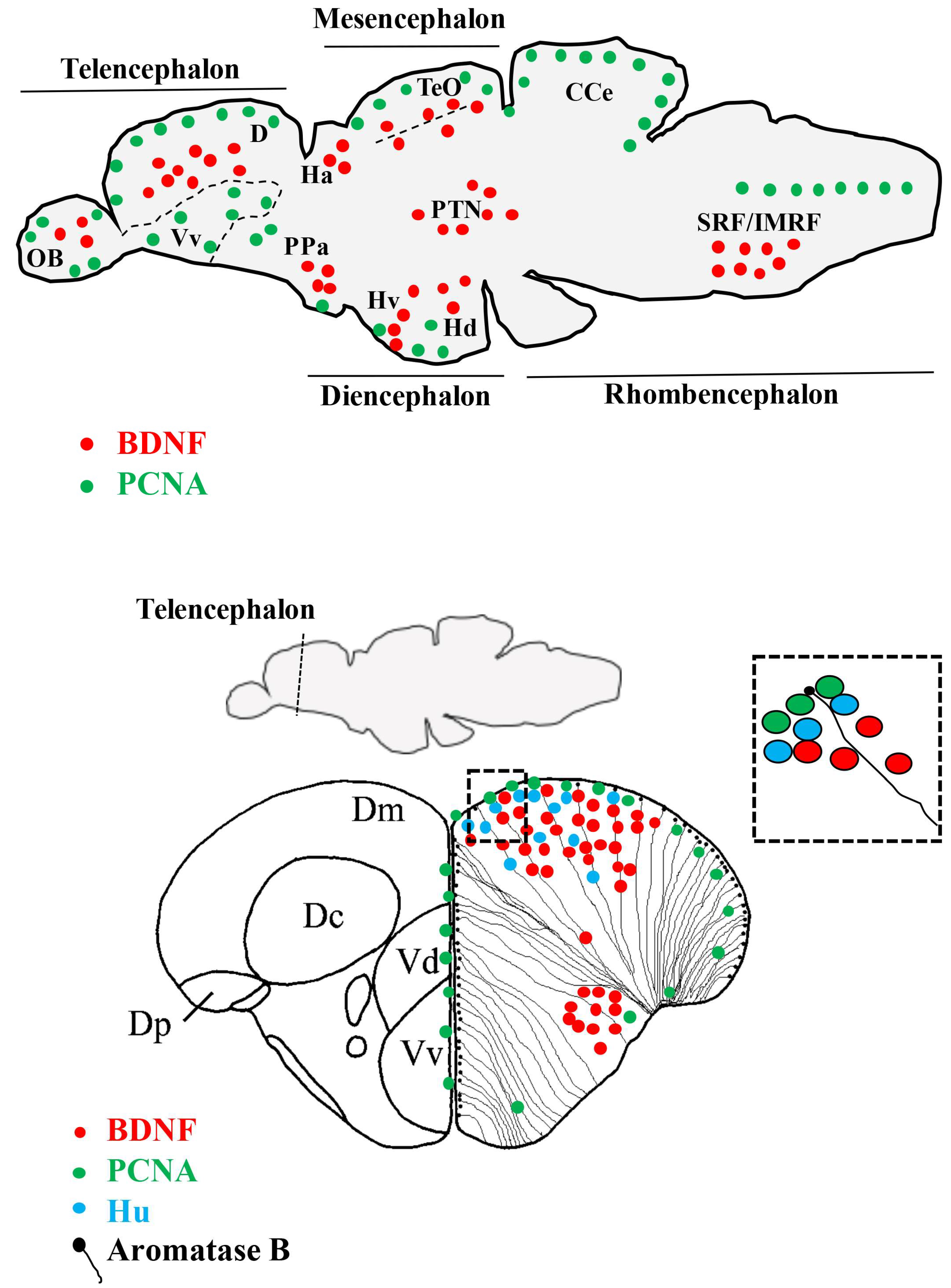
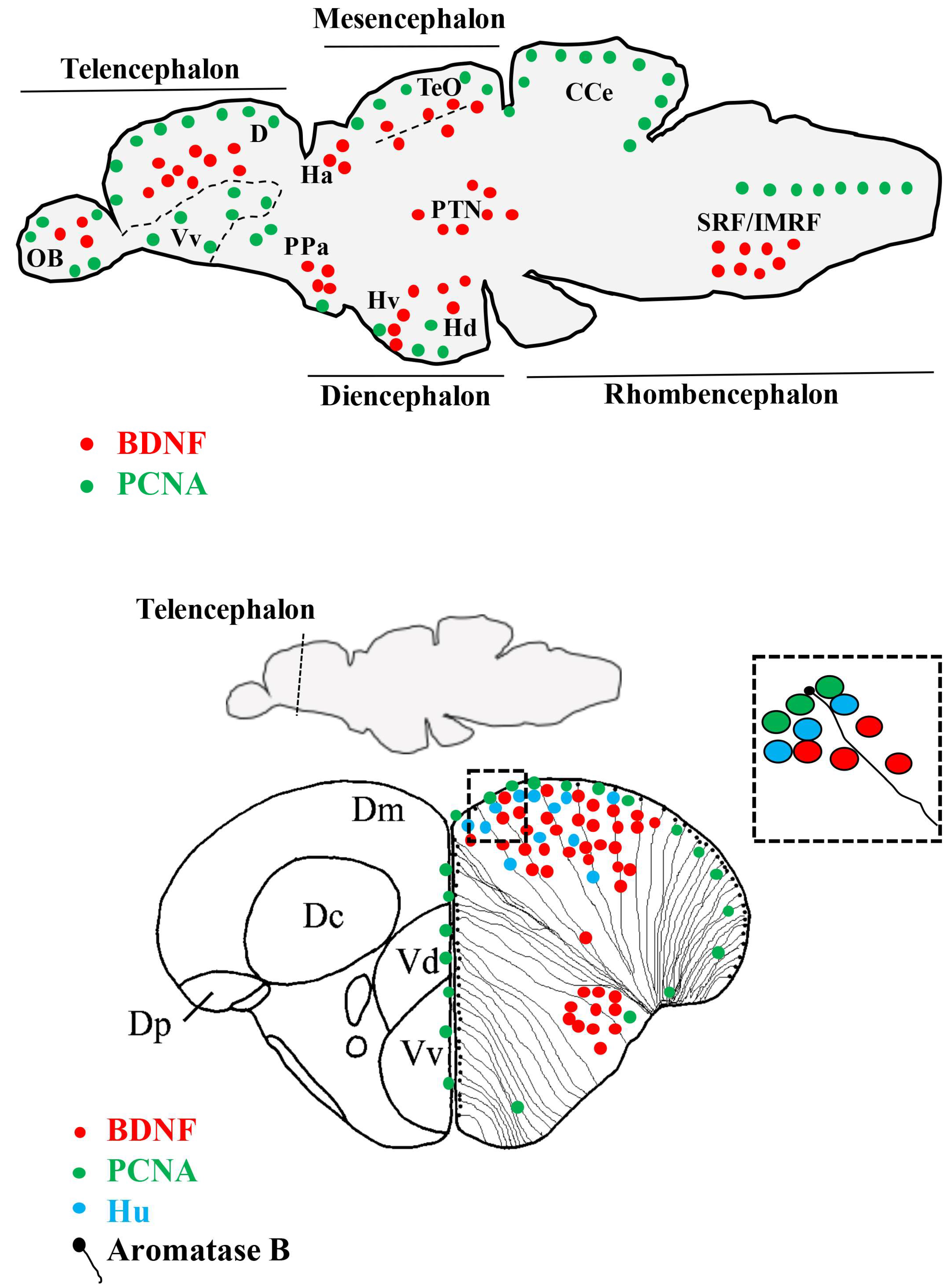
4. The Distribution of BDNF in the Brain of Vertebrates
5. The Distribution of BDNF in the Brain of Zebrafish
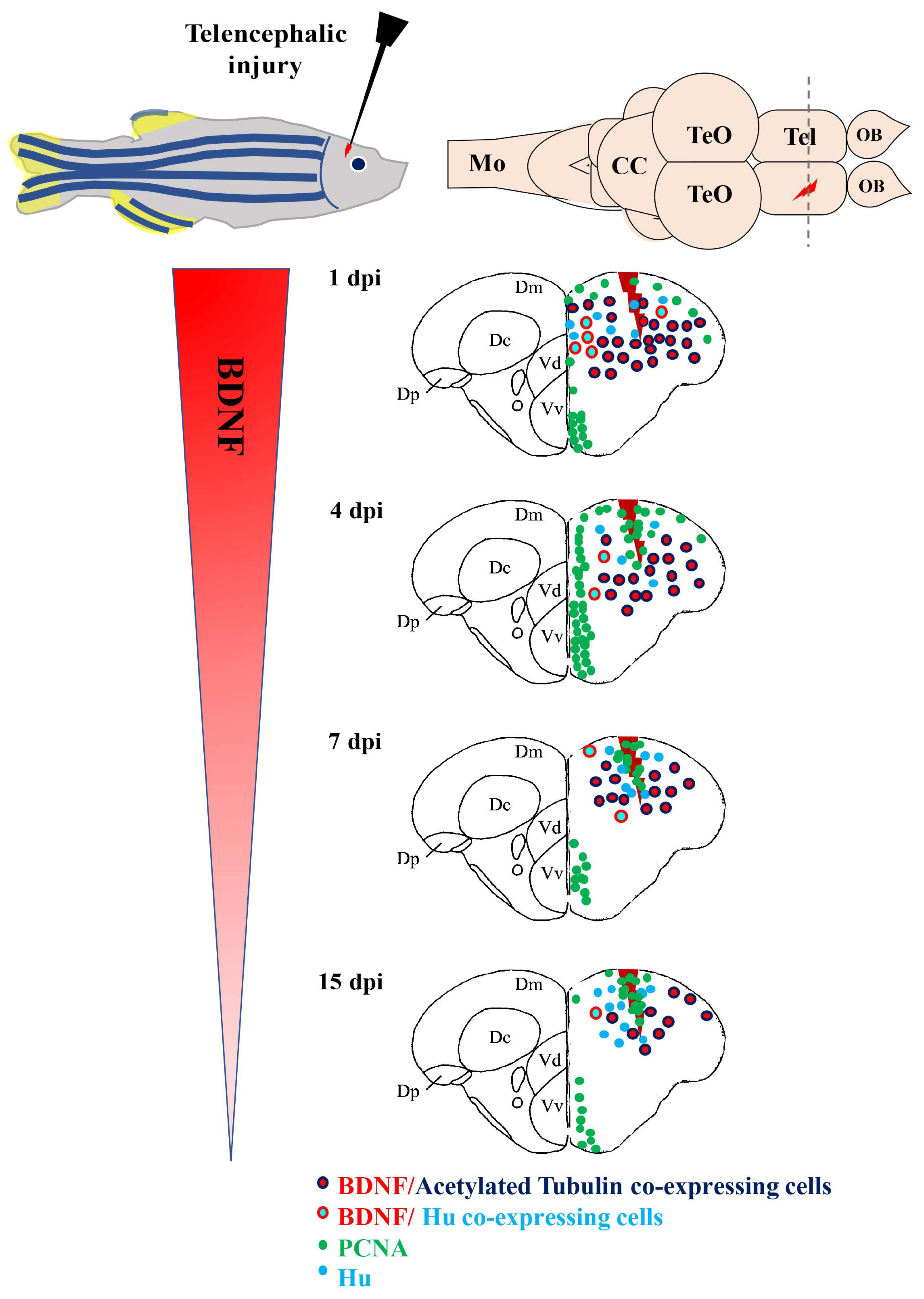
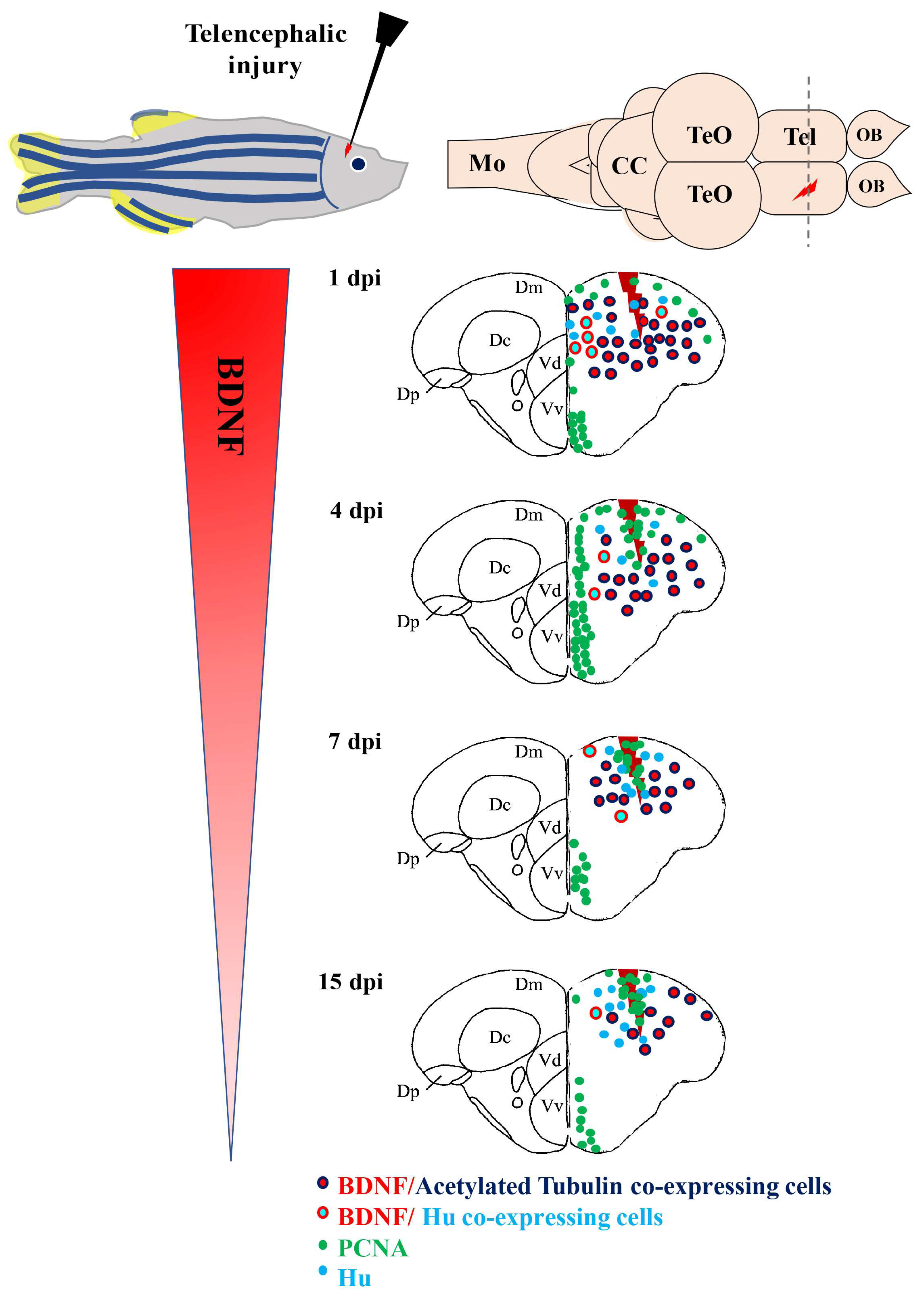
6. BDNF and Adult Neurogenesis, Neuroregeneration in Vertebrate Brain
7. Concluding Remarks
Acknowledgments
Conflicts of Interest
References
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiso, N.; Moro, E.; Argenton, F. Zebrafish pancreas development. Mol. Cell. Endocrinol. 2009, 312, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Jagannathan-Bogdan, M.; Zon, L.I. Hematopoiesis. Development 2013, 140, 2463–2467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vornanen, M.; Hassinen, M. Zebrafish heart as a model for human cardiac electrophysiology. Channels 2016, 10, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Wullimann, M.F.; Rupp, B.; Reichert, H. Neuroanatomy of the Zebrafish Brain: A Topological Atlas; Birkhauser: Berlin, Germany, 1996; ISBN 978-3-0348-8979-7. [Google Scholar]
- Mueller, T.; Wullimann, M.F. Atlas of Early Zebrafish Brain Development; Elsevier: Amsterdam, The Netherlands, 2016; ISBN 978-0-12-418669-9. [Google Scholar]
- Butler, A.B.; Hodos, W. Comparative Vertebrate Neuroanatomy. Evolution and Adaptation, 2nd ed.; John Wiley: Hoboken, NJ, USA, 2015; ISBN1 9780471210054. ISBN2 9780471733843. [Google Scholar] [CrossRef]
- Panula, P.; Chen, Y.C.; Priyadarshini, M.; Kudo, H.; Semenova, S.; Sundvik, M.; Sallinen, V. The comparative neuroanatomy and neurochemistry of zebrafish CNS systems of relevance to human neuropsychiatric diseases. Neurobiol. Dis. 2010, 40, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Randlett, O.; Wee, C.L.; Naumann, E.A.; Nnaemeka, O.; Schoppik, D.; Fitzgerald, J.E.; Portugues, R.; Lacoste, A.M.B.; Riegler, F.; Engert, F.; et al. Whole-brain activity mapping onto a zebrafish brain atlas. Nat. Methods 2015, 12, 1039–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perathoner, S.; Cordero-Maldonado, M.L.; Crawford, A.D. Potential of zebrafish as a model for exploring the role of the amygdala in emotional memory and motivational behavior. J. Neurosci. Res. 2016, 94, 445–462. [Google Scholar] [CrossRef] [PubMed]
- Stewart, A.M.; Braubach, O.; Spitsbergen, J.; Gerlai, R.; Kalueffl, A.V. Zebrafish models for translational neuroscience research: From tank to bedside. Trends Neurosci. 2014, 37, 264–278. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, L.; Lossi, L.; Merighi, A.; de Girolamo, P. Anatomical features for the adequate choice of experimental animal models in biomedicine: I. Fishes. Ann. Anat. Anat. Anz. 2016, 205, 75–84. [Google Scholar] [CrossRef]
- Fontana, B.D.; Mezzomo, N.J.; Kalueff, A.V.; Rosemberg, D.B. The developing utility of zebrafish models of neurological and neuropsychiatric disorders: A critical review. Exp. Neurol. 2018, 299, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Sakai, C.; Ijaz, S.; Hoffman, E.J. Zebrafish Models of Neurodevelopmental Disorders: Past, Present, and Future. Front. Mol. Neurosci. 2018, 11, 294. [Google Scholar] [CrossRef] [PubMed]
- Griffin, A.; Krasniak, C.; Baraban, S.C. Advancing epilepsy treatment through personalized genetic zebrafish models. In Neurobiology of Epilepsy: From Genes to Networks; Rossignol, E., Carmant, L., Lacaille, J.C., Eds.; Progress in Brain Research: Amsterdam, The Netherlands, 2016; Volume 226, pp. 195–207. [Google Scholar]
- Cho, S.J.; Byun, D.; Nam, T.S.; Choi, S.Y.; Lee, B.G.; Kim, M.K.; Kim, S. Zebrafish as an animal model in epilepsy studies with multichannel EEG recordings (vol 7, 3099, 2017). Sci. Rep. 2017, 7, 3099. [Google Scholar] [CrossRef] [PubMed]
- Newman, M.; Ebrahimie, E.; Lardelli, M. Using the zebrafish model for Alzheimer’s disease research. Front. Genet. 2014, 5, 189. [Google Scholar] [CrossRef] [PubMed]
- Babu, N.S.; Murthy, C.L.N.; Kakara, S.; Sharma, R.; Swamy, C.V.B.; Idris, M.M. 1-Methyl-4-phenyl-1,2,3,6-tetrahydropyridine induced Parkinson’s disease in zebrafish. Proteomics 2016, 16, 1407–1420. [Google Scholar] [CrossRef] [PubMed]
- Fonseka, T.M.; Wen, X.Y.; Foster, J.A.; Kennedy, S.H. Zebrafish Models of Major Depressive Disorders. J. Neurosci. Res. 2016, 94, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Giacomotto, J.; Carroll, A.P.; Rinkwitz, S.; Mowry, B.; Cairns, M.J.; Becker, T.S. Developmental suppression of schizophrenia-associated miR-137 alters sensorimotor function in zebrafish. Transl. Psychiatry 2016, 6, e818. [Google Scholar] [CrossRef] [PubMed]
- Meshalkina, D.; Kizlyk, M.N.; Kysil, E.V.; Collier, A.D.; Echevarria, D.J.; Abreu, M.S.; Barcellos, L.J.G.; Song, C.; Warnick, J.E.; Kyzar, E.J.; et al. Zebrafish models of autism spectrum disorder. Exp. Neurol. 2018, 299, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Dorsemans, A.-C.; Soule, S.; Weger, M.; Bourdon, E.; d’Hellencourt, C.L.; Meilhac, O.; Diotel, N. Impaired Constitutive and Regenerative Neurogenesis in Adult Hyperglycemic Zebrafish. J. Comp. Neurol. 2017, 525, 442–458. [Google Scholar] [CrossRef] [PubMed]
- Van Kesteren, R.E.; Fainzilber, M.; Hauser, G.; van Minnen, J.; Vreugdenhil, E.; Smit, A.B.; Ibañez, C.F.; Geraerts, W.P.; Bulloch, A.G. Early evolutionary origin of the neurotrophin receptor family. EMBO J. 1998, 17, 2534–2542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck, G.; Munno, D.W.; Levy, Z.; Dissel, H.M.; Van-Minnen, J.; Syed, N.I.; Fainzilber, M. Neurotrophic activities of trk receptors conserved over 600 million years of evolution. J. Neurobiol. 2004, 60, 12–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bothwell, M. Evolution of the neurotrophin signaling system in invertebrates. Brain Behav. Evol. 2006, 68, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Lapraz, F.; Rottinger, E.; Duboc, V.; Range, R.; Duloquin, L.; Walton, K.; Wu, K.; Bradham, C.; Loza, M.A.; Wilson, K. RTK and TGF-beta signaling pathways genes in the sea urchin genome. Dev. Boil. 2006, 300, 132–152. [Google Scholar] [CrossRef] [PubMed]
- Sossin, W.S. Tracing the evolution and function of the Trk superfamily of receptor tyrosine kinases. Brain Behav. Evol. 2006, 68, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.F.; Pennack, J.A.; McQuilton, P.; Forero, M.G.; Mizuguchi, K.; Sutcliffe, B.; Gu, C.-J.; Fenton, J.C.; Hidalgo, A. Drosophila Neurotrophins Reveal a Common Mechanism for Nervous System Formation. PLoS Boil. 2008, 6, 2476–2495. [Google Scholar] [CrossRef] [PubMed]
- Wilson, K.H.S. The genome sequence of the protostome Daphnia pulex encodes respective orthologues of a neurotrophin, a Trk and a p75NTR: Evolution of neurotrophin signaling components and related proteins in the bilateria. BMC Evol. Biol. 2009, 9. [Google Scholar] [CrossRef] [PubMed]
- Kassabov, S.R.; Choi, Y.B.; Karl, K.A.; Vishwasrao, H.D.; Bailey, C.H.; Kandel, E.R. A Single Aplysia Neurotrophin Mediates Synaptic Facilitation via Differentially Processed Isoforms. Cell Rep. 2013, 3, 1213–1227. [Google Scholar] [CrossRef] [PubMed]
- Lauri, A.; Bertucci, P.; Arendt, D. Neurotrophin, p75, and Trk Signaling Module in the Developing Nervous System of the Marine Annelid Platynereis dumerilii. BioMed Res. Int. 2016, 2016, 2456062. [Google Scholar] [CrossRef]
- Benito-Gutierrez, E.; Garcia-Fernandez, J.; Comella, J.X. Origin and evolution of the Trk family of neurotrophic receptors. Mol. Cell. Neurosci. 2006, 31, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Hallbook, F. Evolution of the vertebrate neurotrophin and Trk receptor gene families. Curr. Opin. Neurobiol. 1999, 9, 616–621. [Google Scholar] [CrossRef]
- Lanave, C.; Colangelo, A.M.; Saccone, C.; Alberghina, L. Molecular evolution of the neurotrophin family members and their Trk receptors. Gene 2007, 394, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Gotz, R.; Schartl, M. The conservation of neurotrophic factors during vertebrate evolution. Comp. Biochem. Physiol. C Pharmacol. Toxicol. Endocrinol. 1994, 108, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, A.S.; Fainzilber, M.; Falck, P.; Ibanez, C.F. Neurotrophin-7: A novel member of the neurotrophin family from the zebrafish. FEBS Lett. 1998, 424, 285–290. [Google Scholar] [CrossRef]
- Lai, K.O.; Fu, W.Y.; Ip, F.C.F.; Ip, N.Y. Cloning and expression of a novel neurotrophin, NT-7, from carp. Mol. Cell. Neurosci. 1998, 11, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, G.; Pagtakhan, C.J. Both 5’ and 3’ flanks regulate Zebrafish brain-derived neurotrophic factor gene expression. BMC Neurosci. 2004, 5, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hempstead, B.L. Brain-Derived Neurotrophic Factor: Three ligands, many actions. Trans. Am. Clin. Climatol. Assoc. 2015, 126, 9–19. [Google Scholar] [PubMed]
- Anastasia, A.; Deinhardt, K.; Chao, M.V.; Will, N.E.; Irmady, K.; Lee, F.S.; Hempstead, B.L.; Bracken, C. Val66Met polymorphism of BDNF alters prodomain structure to induce neuronal growth cone retraction. Nat. Commun. 2013, 4, 2490. [Google Scholar] [CrossRef] [PubMed]
- Tettamanti, G.; Cattaneo, A.G.; Gornati, R.; de Eguileor, M.; Bernardini, G.; Binelli, G. Phylogenesis of brain-derived neurotrophic factor (BDNF) in vertebrates. Gene 2010, 450, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Gotz, R.; Raulf, F.; Schartl, M. Brain-derived neurotrophic factor is more highly conserved in structure and function than nerve growth-factor during vertebrate evolution. J. Neurochem. 1992, 59, 432–442. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Heinrich, G. Brain-derived neurotrophic factor gene expression in the developing zebrafish. Int. J. Dev. Neurosci. 1997, 15, 983–997. [Google Scholar] [CrossRef]
- Heinrich, G.; Lum, T. Fish neurotrophins and Trk receptors. Int. J. Dev. Neurosci. 2000, 18, 1–27. [Google Scholar] [CrossRef]
- Fenner, B.M. Truncated TrkB: Beyond a dominant negative receptor. Cytokine Growth Factor Rev. 2012, 23, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Longo, F.M.; Massa, S.M. Small-molecule modulation of neurotrophin receptors: A strategy for the treatment of neurological disease. Nat. Rev. Drug Discov. 2013, 12, 507–525. [Google Scholar] [CrossRef] [PubMed]
- Harrington, A.W.; Li, Q.M.; Tep, C.; Park, J.B.; He, Z.G.; Yoon, S.O. The Role of Kalirin9 in p75/Nogo Receptor-mediated RhoA Activation in Cerebellar Granule Neurons. J. Boil. Chem. 2008, 283, 24690–24697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunet, F.G.; Roest Crollius, H.; Paris, M.; Aury, J.M.; Gibert, P.; Jaillon, O. Gene loss and evolutionary rates following whole-genome duplication in teleost fishes. Mol. Boil. Evol. 2006, 23, 1808–1816. [Google Scholar] [CrossRef] [PubMed]
- Han, H.W.; Chou, C.M.; Chu, C.Y.; Cheng, C.H.; Yang, C.H.; Hung, C.C.; Hwang, P.P.; Lee, S.J.; Liao, Y.F.; Huang, C.J. The Nogo-C2/Nogo Receptor Complex Regulates the Morphogenesis of Zebrafish Lateral Line Primordium through Modulating the Expression of dkk1b, a Wnt Signal Inhibitor. PLoS ONE 2014, 9, e86345. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.C.; Marazzi, G.; Sandell, J.H.; Heinrich, G. 5 Trk receptors in the zebrafish. Dev. Boil. 1995, 169, 745–758. [Google Scholar] [CrossRef] [PubMed]
- Ohta, K.; Suzuki, S.; Warita, K.; Kaji, T.; Kusaka, T.; Miki, T. Prolonged maternal separation attenuates BDNF-ERK signaling correlated with spine formation in the hippocampus during early brain development. J. Neurochem. 2017, 141, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Kojima, M.; Mizui, T. BDNF Propeptide: A Novel Modulator of Synaptic Plasticity. In Vitamins and Hormones; Litwack, G., Ed.; Academic Press: Amsterdam, The Netherlands, 2017; Volume 104, pp. 19–28. [Google Scholar]
- Leal, G.; Bramham, C.R.; Duarte, C.B. BDNF and Hippocampal Synaptic Plasticity. In Vitamins and Hormones; Litwack, G., Ed.; Academic Press: Amsterdam, The Netherlands, 2017; Volume 104, pp. 153–195. [Google Scholar]
- Skibinska, M.; Groszewska, A.; Kapelski, P.; Rajewska-Rager, A.; Pawlak, J.; Dmitrzak-Weglarz, M.; Szczepankiewicz, A.; Twarowska-Hauser, J. Val66Met functional polymorphism and serum protein level of brain-derived neurotrophic factor (BDNF) in acute episode of schizophrenia and depression. Pharmacol. Rep. 2018, 70, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Youssef, M.M.; Underwood, M.D.; Huang, Y.Y.; Hsiung, S.C.; Liu, Y.; Simpson, N.R.; Akalian, M.J.; Rosoklija, G.B.; Dwork, A.J.; Arango, V.; et al. Association of BDNF Va166Met Polymorphism and Brain BDNF Levels with Major Depression and Suicide. International. J. Neuropsychopharmacol. 2018, 21, 528–538. [Google Scholar] [CrossRef] [PubMed]
- Wysokinski, A. Serum levels of brain-derived neurotrophic factor (BDNF) and neurotrophin-3 (NT-3) in depressed patients with schizophrenia. Nord. J. Psychiatry 2016, 70, 267–271. [Google Scholar] [CrossRef] [PubMed]
- Gulyaeva, N.V. Interplay between brain BDNF and glutamatergic systems: A brief state of the evidence and association with the pathogenesis of depression. Biochem. Mosc. 2017, 82, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Molendijk, M.L.; Spinhoven, P.; Polak, M.; Bus, B.A.A.; Penninx, B.; Elzinga, B.M. Serum BDNF concentrations as peripheral manifestations of depression: Evidence from a systematic review and meta-analyses on 179 associations (N = 9484). Mol. Psychiatry 2014, 19, 791–800. [Google Scholar] [CrossRef] [PubMed]
- Francis, K.; Dougali, A.; Sideri, K.; Kroupis, C.; Vasdekis, V.; Dima, K.; Douzenis, A. Brain-derived neurotrophic factor (BDNF) in children with ASD and their parents: A 3-year follow-up. Acta Psychiatr. Scand. 2018, 137, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Tapia-Arancibia, L.; Aliaga, E.; Silhol, M.; Arancibia, S. New insights into brain BDNF function in normal aging and Alzheimer disease. Brain Res. Rev. 2008, 59, 201–220. [Google Scholar] [CrossRef] [PubMed]
- Khorshid Ahmad, T.; Acosta, C.; Cortes, C.; Lakowski, T.M.; Gangadaran, S.; Namaka, M. Transcriptional Regulation of Brain-Derived Neurotrophic Factor (BDNF) by Methyl CpG Binding Protein 2 (MeCP2): A Novel Mechanism for Re-Myelination and/or Myelin Repair Involved in the Treatment of Multiple Sclerosis (MS). Mol. Neurobiol. 2016, 53, 1092–1107. [Google Scholar] [CrossRef] [PubMed]
- Maisonpierre, P.C.; Belluscio, L.; Friedman, B.; Alderson, R.F.; Wiegand, S.J.; Furth, M.E.; Lindsay, R.M.; Yancopopulos, G.D. NT-3, BDNF, and NGF in the developing rat nervous-system—Parallel as well as reciprocal patterns of expression. Neuron 1990, 5, 501–509. [Google Scholar] [CrossRef]
- Hofer, M.; Pagliusi, S.R.; Hohn, A.; Leibrock, J.; Barde, Y.A. Regional distribution of Brain-Derived Neurotrophic Factor messenger-RNA in the adult-mouse brain. EMBO J. 1990, 9, 2459–2464. [Google Scholar] [CrossRef] [PubMed]
- Castren, E.; Thoenen, H.; Lindholm, D. Brain-Derived Neurotrophic Factor messenger-RNA is expressed in the septum, hypothalamus and in adrenergic brain-stem nuclei of adult-rat brain and is increased by osmotic stimulation in the paraventricular nucleus. Neuroscience 1995, 64, 71–80. [Google Scholar] [CrossRef]
- Yan, Q.; Rosenfeld, R.D.; Matheson, C.R.; Hawkins, N.; Lopez, O.T.; Bennett, L.; Welcher, A.A. Expression of brain-derived neurotrophic factor protein in the adult rat central nervous system. Neuroscience 1997, 78, 431–448. [Google Scholar] [CrossRef]
- Conner, J.M.; Lauterborn, J.C.; Yan, Q.; Gall, C.M.; Varon, S. Distribution of brain-derived neurotrophic factor (BDNF) protein and mRNA in the normal adult rat CNS: Evidence for anterograde axonal transport. J. Neurosci. 1997, 17, 2295–2313. [Google Scholar] [CrossRef] [PubMed]
- Murer, M.G.; Raisman-Vozari, R.; Yan, Q.; Ruberg, M.; Agid, Y.; Michel, P.P. Survival factors promote BDNF protein expression in mesencephalic dopaminergic neurons. Neuroreport 1999, 10, 801–805. [Google Scholar] [CrossRef] [PubMed]
- Quartu, M.; Serra, M.P.; Boi, M.; Melis, T.; Ambu, R.; Del Fiacco, M. Brain-derived neurotrophic factor (BDNF) and polysialylated-neural cell adhesion molecule (PSA-NCAM): Codistribution in the human brainstem precerebellar nuclei from prenatal to adult age. Brain Res. 2010, 1363, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.P.; Wade, J. 17 beta-Estradiol Regulates the Sexually Dimorphic Expression of BDNF and TrkB Proteins in the Song System of Juvenile Zebra Finches. PLoS ONE 2012, 7, e43687. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.P.; Wade, J. Sex and age differences in brain-derived neurotrophic factor and vimentin in the zebra finch song system: Relationships to newly generated cells. J. Comp. Neurol. 2016, 524, 1081–1096. [Google Scholar] [CrossRef] [PubMed]
- Brenowitz, E.A. Testosterone and Brain-Derived Neurotrophic Factor interactions in the avian song control system. Neuroscience 2013, 239, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Theiss, C.; Gunturkun, O. Distribution of BDNF, NT-3, trkB and trkC in the developing retino-tectal system of the pigeon (Columba livia). Anat. Embryol. 2001, 204, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Duprey-Diaz, M.V.; Soto, I.; Blagburn, J.M.; Blanco, R.E. Changes in brain-derived neurotrophic factor and trkB receptor in the adult Rana pipiens retina and optic tectum after optic nerve injury. J. Comp. Neurol. 2002, 454, 456–469. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Humbel, B.M.; Roubos, E.W. High-pressure freezing followed by cryosubstitution as a tool for preserving high-quality ultrastructure and immunoreactivity in the Xenopus laevis pituitary gland. Brain Res. Brain Res. Protoc. 2005, 15, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Vissio, P.G.; Canepa, M.M.; Maggese, M.C. Brain-derived neurotrophic factor (BDNF)-like immunoreactivity localization in the retina and brain of Cichlasoma dimerus (Teleostei, Perciformes). Tissue Cell 2008, 40, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Dalton, V.S.; Roberts, B.L.; Borich, S.M. Brain derived neurotrophic factor and trk B mRNA expression in the brain of a brain stem-spinal cord regenerating model, the European eel, after spinal cord injury. Neurosci. Lett. 2009, 461, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Dalton, V.S.; Borich, S.M.; Murphy, P.; Roberts, B.L. Brain-Derived Neurotrophic Factor mRNA Expression in the Brain of the Teleost Fish, Anguilla anguilla, the European Eel. Brain Behav. Evol. 2009, 73, 43–58. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, L.; De Girolamo, P.; Lucini, C.; Terzibasi, E.T.; Baumgart, M.; Castaldo, L.; Cellerino, A. Brain-Derived Neurotrophic Factor: mRNA Expression and Protein Distribution in the Brain of the Teleost Nothobranchius furzeri. J. Comp. Neurol. 2014, 522, 1004–1030. [Google Scholar] [CrossRef] [PubMed]
- Catania, S.; Germana, A.; Cabo, R.; Ochoa-Erena, F.J.; Guerrera, M.C.; Hannestad, J.; Represa, J.; Vega, J.A. Neurotrophin and Trk neurotrophin receptors in the inner ear of Salmo salar and Salmo trutta. J. Anat. 2007, 210, 78–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Germana, A.; Catania, S.; Cavallaro, M.; Gonzalez-Martinez, T.; Ciriaco, E.; Hannestad, J.; Vega, J.A. Immunohistochemical localization of BDNF-, TrkB- and TrkA-like proteins in the teleost lateral line system. J. Anat. 2002, 200, 477–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cellerino, A.; Kohler, K. Brain-derived neurotrophic factor neurotrophin-4 receptor TrkB is localized on ganglion cells and dopaminergic amacrine cells in the vertebrate retina. J. Comp. Neurol. 1997, 386, 149–160. [Google Scholar] [CrossRef]
- Caminos, E.; Becker, E.; Martin-Zanca, D.; Vecino, E. Neurotrophins and their receptors in the tench retina during optic nerve regeneration. J. Comp. Neurol. 1999, 404, 321–331. [Google Scholar] [CrossRef]
- Gatta, C.; Castaldo, L.; Cellerino, A.; de Girolamo, P.; Lucini, C.; D’Angelo, L. Brain derived neurotrophic factor in the retina of the teleost, N. furzeri. Ann. Anat. Anat. Anz. 2014, 196, 192–196. [Google Scholar] [CrossRef] [PubMed]
- Cacialli, P.; D’Angelo, L.; de Girolamo, P.; Avallone, L.; Lucini, C.; Pellegrini, E.; Castaldo, L. Morpho-Functional Features of the Gonads of Danio rerio: The Role of Brain-Derived Neurotrophic Factor. Anat. Rec. 2018, 301, 140–147. [Google Scholar] [CrossRef] [PubMed]
- De Felice, E.; Porreca, I.; Alleva, E.; Girolamo, P.; Ambrosino, C.; Ciriaco, E.; Germanà, A.; Sordino, P. Localization of BDNF expression in the developing brain of zebrafish. J. Anat. 2014, 224, 564–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nittoli, V.; Sepe, R.M.; Coppola, U.; D’Agostino, Y.; De Felice, E.; Palladino, A.; Vassalli, Q.A.; Locascio, A.; Ristoratore, F.; Spagnuolo, A.; et al. A comprehensive analysis of neurotrophins and neurotrophin tyrosine kinase receptors expression during development of zebrafish. J. Comp. Neurol. 2018, 526, 1057–1072. [Google Scholar] [CrossRef] [PubMed]
- Cacialli, P.; Gueguen, M.M.; Coumailleau, P.; D’Angelo, L.; Kah, O.; Lucini, C.; Pellegrini, E. BDNF Expression in Larval and Adult Zebrafish Brain: Distribution and Cell Identification. PLoS ONE 2016, 11, e0158057. [Google Scholar] [CrossRef] [PubMed]
- Hill, R.A. Interaction of Sex Steroid Hormones and Brain-Derived Neurotrophic Factor-Tyrosine Kinase B Signalling: Relevance to Schizophrenia and Depression. J. Neuroendocr. 2012, 24, 1553–1561. [Google Scholar] [CrossRef] [PubMed]
- Hill, R.A.; Wu, Y.W.C.; Kwek, P.; van den Buuse, M. Modulatory Effects of Sex Steroid Hormones on Brain-Derived Neurotrophic Factor-Tyrosine Kinase B Expression during Adolescent Development in C57Bl/6 Mice. J. Neuroendocr. 2012, 24, 774–788. [Google Scholar] [CrossRef] [PubMed]
- Carbone, D.L.; Handa, R.J. Sex and stress hormone influences on the expression and activity of Brain-Derived Neurotrophic Factor. Neuroscience 2013, 239, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.H.; Ueshima, E.; Muraoka, O.; Tanaka, H.; Yeo, S.Y.; Huh, T.L.; Miki, N. Zebrafish elav/HuC homologue as a very early neuronal marker. Neurosci. Lett. 1996, 216, 109–112. [Google Scholar] [CrossRef]
- Ilieva, M.; Della Vedova, P.; Hansen, O.; Dufva, M. Tracking neuronal marker expression inside living differentiating cells using molecular beacons. Front. Cell. Neurosci. 2013, 7, 266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janke, C.; Kneussel, M. Tubulin post-translational modifications: Encoding functions on the neuronal microtubule cytoskeleton. Trends Neurosci. 2010, 33, 362–372. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.; Strahle, U.; Scholpp, S. Neurogenesis in zebrafish—From embryo to adult. Neural Dev. 2013, 8, 3. [Google Scholar] [CrossRef] [PubMed]
- Tokumine, J.; Kakinohana, O.; Cizkova, D.; Smith, D.W.; Marsala, M. Changes in spinal GDNF, BDNF, and NT-3 expression after transient ischemia in the rat. J. Neurosci. Res. 2003, 74, 552–561. [Google Scholar] [CrossRef] [PubMed]
- Burbach, G.J.; Hellweg, R.; Haas, C.A.; Del Turco, D.; Deicke, U.; Abramowski, D.; Jucker, M.; Staufenbiel, M.; Deller, T. Induction of brain-derived neurotrophic factor in plaque-associated glial cells of aged APP23 transgenic mice. J. Neurosci. 2004, 24, 2421–2430. [Google Scholar] [CrossRef] [PubMed]
- Gatta, C.; Altamura, G.; Avallone, L.; Castaldo, L.; Corteggio, A.; D’Angelo, L.; de Girolamo, P.; Lucini, C. Neurotrophins and Their Trk-Receptors in the Cerebellum of Zebrafish. J. Morphol. 2016, 277, 725–736. [Google Scholar] [CrossRef] [PubMed]
- Northcutt, R.G. Do Teleost Fishes Possess a Homolog of Mammalian Isocortex? Brain Behav. Evol. 2011, 78, 136–138. [Google Scholar] [CrossRef] [PubMed]
- Mueller, T.; Dong, Z.Q.; Berberoglu, M.A.; Guo, S. The dorsal pallium in zebrafish, Danio rerio (Cyprinidae, Teleostei). Brain Res. 2011, 1381, 95–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barker, J.M.; Boonstra, R.; Wojtowicz, J.M. From pattern to purpose: How comparative studies contribute to understanding the function of adult neurogenesis. Eur. J. Neurosci. 2011, 34, 963–977. [Google Scholar] [CrossRef] [PubMed]
- Sorrells, S.F.; Paredes, M.F.; Ebrian-Silla, A.C.; Sandoval, K.; Qi, D.S.; Kelley, K.W.; James, D.; Mayer, S.; Chanh, J.; Auguste, K.I.; et al. Human hippocampal neurogenesis drops sharply in children to undetectable levels in adults. Nature 2018, 555, 377. [Google Scholar] [CrossRef] [PubMed]
- Boldrini, M.; Fulmore, C.A.; Tartt, A.N.; Simeon, L.R.; Pavlova, I.; Poposka, V.; Rosoklija, G.B.; Stankov, A.; Arango, V.; Dwork, A.l.; et al. Human Hippocampal Neurogenesis Persists throughout Aging. Cell Stem Cell 2018, 22, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Kempermann, G.; Gage, F.H.; Aigner, L.; Song, H.; Curtis, M.A.; Thuret, S.; Kuhn, H.G.; Jessberger, S.; Frankland, P.W.; Cameron, H.A.; et al. Human Adult Neurogenesis: Evidence and Remaining Questions. Cell Stem Cell 2018, 23, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Grandel, H.; Kaslin, J.; Ganz, J.; Wenzel, I.; Brand, M. Neural stem cells and neurogenesis in the adult zebrafish brain: Origin, proliferation dynamics, migration and cell fate. Dev. Boil. 2006, 295, 263–277. [Google Scholar] [CrossRef] [PubMed]
- Zupanc, G.K.H.; Hinsch, K.; Gage, F.H. Proliferation, migration, neuronal differentiation, and long-term survival of new cells in the adult zebrafish brain. J. Comp. Neurol. 2005, 488, 290–319. [Google Scholar] [CrossRef] [PubMed]
- Adolf, B.; Chapouton, P.; Lam, C.S.; Topp, S.; Tannhauser, B.; Strahle, U.; Gotz, M.; Bally-Cuif, L. Conserved and acquired features of adult neurogenesis in the zebrafish telencephalon. Dev. Boil. 2006, 295, 278–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapouton, P.; Jagasia, R.; Bally-Cuif, L. Adult neurogenesis in non-mammalian vertebrates. Bioessays 2007, 29, 745–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellegrini, E.; Mouriec, K.; Anglade, I.; Menuet, A.; Le Page, Y.; Gueguen, M.M.; Marmignon, M.H.; Brion, F.; Pakdel, F.; Kah, O. Identification of aromatase-positive radial glial cells as progenitor cells in the ventricular layer of the forebrain in zebrafish. J. Comp. Neurol. 2007, 501, 150–167. [Google Scholar] [CrossRef] [PubMed]
- Kizil, C.; Kaslin, J.; Kroehne, V.; Brand, M. Adult neurogenesis and brain regeneration in zebrafish. Dev. Neurobiol. 2012, 72, 429–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diotel, N.; Vaillant, C.; Gabbero, C.; Mironov, S.; Fostier, A.; Gueguen, M.M.; Anglade, I.; Kah, O.; Pellegrini, E. Effects of estradiol in adult neurogenesis and brain repair in zebrafish. Horm. Behav. 2013, 63, 193–207. [Google Scholar] [CrossRef] [PubMed]
- Zigova, T.; Pencea, V.; Wiegand, S.J.; Luskin, M.B. Intraventricular administration of BDNF increases the number of newly generated neurons in the adult olfactory bulb. Mol. Cell. Neurosci. 1998, 11, 234–245. [Google Scholar] [CrossRef] [PubMed]
- Pencea, V.; Bingaman, K.D.; Wiegand, S.J.; Luskin, M.B. Infusion of brain-derived neurotrophic factor into the lateral ventricle of the adult rat leads to new neurons in the parenchyma of the striatum, septum, thalamus, and hypothalamus. J. Neurosci. 2001, 21, 6706–6717. [Google Scholar] [CrossRef] [PubMed]
- Lindholm, D.; Carroll, P.; Tzimagiorgis, G.; Thoenen, H. Autocrine-paracrine regulation of hippocampal neuron survival by IGF-1 and the neurotrophins BDNF, NT-3 and NT-4. Eur. J. Neurosci. 1996, 8, 1452–1460. [Google Scholar] [CrossRef] [PubMed]
- Galvao, R.P.; Garcia-Verdugo, J.M.; Alvarez-Buylla, A. Brain-Derived Neurotrophic Factor Signaling Does Not Stimulate Subventricular Zone Neurogenesis in Adult Mice and Rats. J. Neurosci. 2008, 28, 13368–13383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bath, K.G.; Akins, M.R.; Lee, F.S. BDNF control of adult SVZ neurogenesis. Dev. Psychobiol. 2012, 54, 578–589. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Duan, W.; Mattson, M.P. Evidence that brain-derived neurotrophic factor is required for basal neurogenesis and mediates, in part, the enhancement of neurogenesis by dietary restriction in the hippocampus of adult mice. J. Neurochem. 2002, 82, 1367–1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, J.P.; Cordeira, J.; Calderon, G.A.; Iyer, L.K.; Rios, M. Depletion of central BDNF in mice impedes terminal differentiation of new granule neurons in the adult hippocampus. Mol. Cell. Neurosci. 2008, 39, 372–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Cao, M.; Liu, A.Q.; Di, W.Y.; Zhao, F.; Tian, Y.J.; Jia, J. Changes of Inflammatory Cytokines and Neurotrophins Emphasized Their Roles in Hypoxic-Ischemic Brain Damage. Int. J. Neurosci. 2013, 123, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, G.B.; Vasterling, J.J.; Vedak, P.C. Brain-derived neurotrophic factor in traumatic brain injury, post-traumatic stress disorder, and their comorbid conditions: Role in pathogenesis and treatment. Behav. Pharmacol. 2010, 21, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Rostami, E.; Krueger, F.; Plantman, S.; Davidsson, J.; Agoston, D.; Grafman, J.; Risling, M. Alteration in BDNF and its receptors, full-length and truncated TrkB and p75(NTR) following penetrating traumatic brain injury. Brain Res. 2014, 1542, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Schober, M.E.; Block, B.; Requena, D.F.; Hale, M.A.; Lane, R.H. Developmental traumatic brain injury decreased brain derived neurotrophic factor expression late after injury. Metab. Brain Dis. 2012, 27, 167–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skoglosa, Y.; Lewen, A.; Takei, N.; Hillered, L.; Lindholm, D. Regulation of pituitary adenylate cyclase activating polypeptide and its receptor type 1 after traumatic brain injury: Comparison with brain-derived neurotrophic factor and the induction of neuronal cell death. Neuroscience 1999, 90, 235–247. [Google Scholar] [CrossRef]
- Griesbach, G.S.; Hovda, D.A.; Molteni, R.; Gomez-Pinilla, F. Alterations in BDNF and synapsin I within the occipital cortex and hippocampus after mild traumatic brain injury in the developing rat: Reflections of injury-induced neuroplasticity. J. Neurotrauma 2002, 19, 803–814. [Google Scholar] [CrossRef] [PubMed]
- Hicks, R.R.; Li, C.Y.; Zhang, L.X.; Dhillon, H.S.; Prasad, M.R.; Seroogy, K.B. Alterations in BDNF and trkB mRNA levels in the cerebral cortex following experimental brain trauma in rats. J. Neurotrauma 1999, 16, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Felderhoff-Mueser, U.; Sifringer, M.; Pesditschek, S.; Kuckuck, H.; Moysich, A.; Bittigau, P.; Ikonomidou, C. Pathways leading to apoptotic neurodegeneration following trauma to the developing rat brain. Neurobiol. Dis. 2002, 11, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Hicks, R.R.; Numan, S.; Dhillon, H.S.; Prasad, M.R.; Seroogy, K.B. Alterations in BDNF and NT-3 mRNAs in rat hippocampus after experimental brain trauma. Mol. Brain Res. 1997, 48, 401–406. [Google Scholar] [CrossRef]
- Alder, J.; Fujioka, W.; Giarratana, A.; Wissocki, J.; Thakkar, K.; Vuong, P.; Patel, B.; Chakraborty, T.; Elsabeh, R.; Parikh, A.; et al. Genetic and pharmacological intervention of the p75NTR pathway alters morphological and behavioural recovery following traumatic brain injury in mice. Brain Inj. 2016, 30, 48–65. [Google Scholar] [CrossRef] [PubMed]
- Delbary-Gossart, S.; Lee, S.; Baroni, M.; Lamarche, I.; Arnone, M.; Canolle, B.; Lin, A.; Sacramento, J.; Salegio, E.A.; Castel, M.N.; et al. A novel inhibitor of p75-neurotrophin receptor improves functional outcomes in two models of traumatic brain injury. Brain 2016, 139, 1762–1782. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, I.; Ballabriga, J.; Marti, E.; Pozas, E.; Planas, A.M.; Blasi, J. BDNF and TrkB co-localize in CA1 neurons resistant to transient forebrain ischemia in the adult gerbil. J. Neuropathol. Exp. Neurol. 1997, 56, 790–797. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.L.; Hu, Y.; Zhang, P.; Zhang, Z.; Li, L.H.; Gao, G.D.; Zhou, X.F.; Wang, T.H. Neural Stem Cell Transplantation Promotes Functional Recovery from Traumatic Brain Injury via Brain Derived Neurotrophic Factor-Mediated Neuroplasticity. Mol. Neurobiol. 2018, 55, 2696–2711. [Google Scholar] [CrossRef] [PubMed]
- Xuan, W.J.; Agrawal, T.; Huang, L.Y.; Gupta, G.K.; Hamblin, M.R. Low-level laser therapy for traumatic brain injury in mice increases brain derived neurotrophic factor (BDNF) and synaptogenesis. J. Biophotonics 2015, 8, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Su, W.S.; Wu, C.H.; Chen, S.F.; Yang, F.Y. Transcranial ultrasound stimulation promotes brain-derived neurotrophic factor and reduces apoptosis in a mouse model of traumatic brain injury. Brain Stimul. 2017, 10, 1032–1041. [Google Scholar] [CrossRef] [PubMed]
- Griesbach, G.S.; Hovda, D.A.; Gomez-Pinilla, F. Exercise-induced improvement in cognitive performance after traumatic brain injury in rats is dependent on BDNF activation. Brain Res. 2009, 1288, 105–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wurzelmann, M.; Romeika, J.; Sun, D. Therapeutic potential of brain-derived neurotrophic factor (BDNF) and a small molecular mimics of BDNF for traumatic brain injury. Neural Regen. Res. 2017, 12, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Kishimoto, N.; Shimizu, K.; Sawamoto, K. Neuronal regeneration in a zebrafish model of adult brain injury. Dis. Model. Mech. 2012, 5, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Cacialli, P.; D’Angelo, L.; Kah, O.; Coumailleau, P.; Gueguen, M.M.; Pellegrini, E.; Lucini, C. Neuronal expression of brain derived neurotrophic factor in the injured telencephalon of adult zebrafish. J. Comp. Neurol. 2018, 526, 569–582. [Google Scholar] [CrossRef] [PubMed]
- Cacialli, P.; Palladino, A.; Lucini, C. Role of brain-derived neurotrophic factor during the regenerative response after traumatic brain injury in adult zebrafish. Neural Regen. Res. 2018, 13, 941–944. [Google Scholar] [CrossRef] [PubMed]
- Salas, C.; Broglio, C.; Rodriguez, F. Evolution of forebrain and spatial cognition in vertebrates: Conservation across diversity. Brain Behav. Evol. 2003, 62, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Lum, T.; Huynh, G.; Heinrich, G. Brain-derived neurotrophic factor and TrkB tyrosine kinase receptor gene expression in zebrafish embryo and larva. Int. J. Dev. Neurosci. 2001, 19, 569–587. [Google Scholar] [CrossRef]
- Abbate, F.; Guerrera, M.C.; Montalbano, G.; Levanti, M.B.; Germanà, G.P.; Navarra, M.; Laurà, R.; Vega, J.A.; Ciriaco, E.; Germanà, A. Expression and anatomical distribution of trkB in the encephalon of the adult zebrafish. Neurosci. Lett. 2014, 20, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Anand, S.K.; Mondel, A.C. TrkB receptor antagonism inhibits stab injury induced proliferative response in adult zebrafish (Danio rerio) brain. Neurosci. Lett. 2018, 672, 28–33. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, L.; de Girolamo, P.; Cellerino, A.; Tozzini, E.T.; Castaldo, L.; Lucini, C. Neurotrophin Trk receptors in the brain of a teleost fish, Nothobranchius furzeri. Microsc. Res. Tech. 2012, 75, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Kyritsis, N.; Kizil, C.; Zocher, S.; Kroehne, V.; Kaslin, J.; Freudenreich, D.; Iltzsche, A.; Brand, M. Acute Inflammation Initiates the Regenerative Response in the Adult Zebrafish Brain. Science 2012, 338, 1353–1356. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Sheng, W.; Han, L.; He, Q.; Ji, X.; Liu, K. Activation of BDNF-TrkB signaling pathway-regulated brain inflammation in pentylenetetrazole-induced seizures in zebrafish. Fish Shellfish Immunol. 2018, 83, 26–36. [Google Scholar] [CrossRef] [PubMed]
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lucini, C.; D’Angelo, L.; Cacialli, P.; Palladino, A.; De Girolamo, P. BDNF, Brain, and Regeneration: Insights from Zebrafish. Int. J. Mol. Sci. 2018, 19, 3155. https://doi.org/10.3390/ijms19103155
Lucini C, D’Angelo L, Cacialli P, Palladino A, De Girolamo P. BDNF, Brain, and Regeneration: Insights from Zebrafish. International Journal of Molecular Sciences. 2018; 19(10):3155. https://doi.org/10.3390/ijms19103155
Chicago/Turabian StyleLucini, Carla, Livia D’Angelo, Pietro Cacialli, Antonio Palladino, and Paolo De Girolamo. 2018. "BDNF, Brain, and Regeneration: Insights from Zebrafish" International Journal of Molecular Sciences 19, no. 10: 3155. https://doi.org/10.3390/ijms19103155
APA StyleLucini, C., D’Angelo, L., Cacialli, P., Palladino, A., & De Girolamo, P. (2018). BDNF, Brain, and Regeneration: Insights from Zebrafish. International Journal of Molecular Sciences, 19(10), 3155. https://doi.org/10.3390/ijms19103155