Proteoglycans Are Attractive Biomarkers and Therapeutic Targets in Hepatocellular Carcinoma


Abstract
1. Introduction
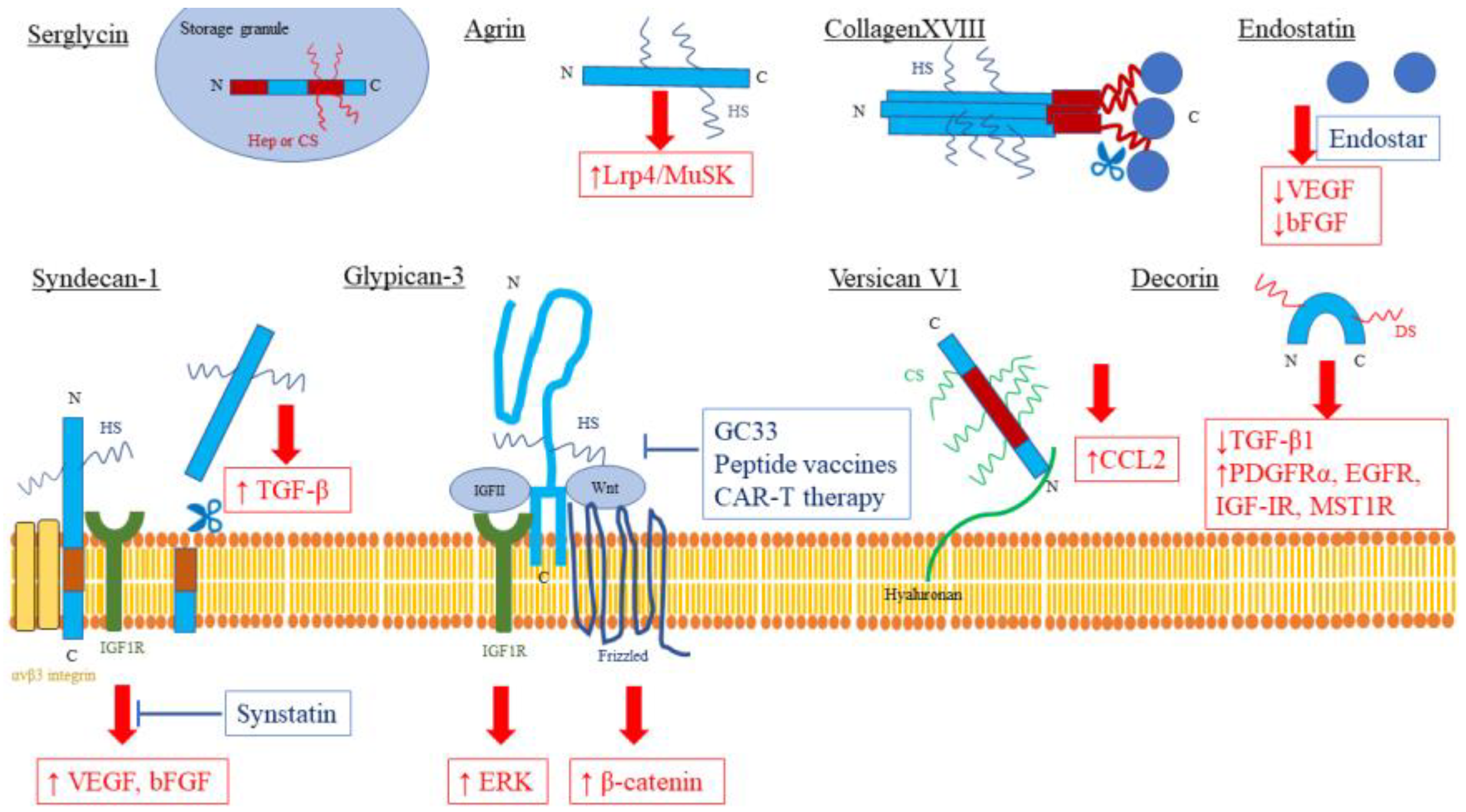
2. Intracellular Proteoglycan: Serglycin
3. Cell Surface Proteoglycan (Transmembrane): Syndecan-1/CD138
4. Cell Surface Proteoglycan (Glycosylphosphatidylinositol-Anchored): Glypican-3
5. Pericellular Proteoglycan: Agrin
6. Pericellular Proteoglycan: Collagen XVIII/Endostatin
7. Extracellular Proteoglycan (Hyalectan): Versican
8. Extracellular Proteoglycan (Small Leucine-Rich Proteoglycans): Decorin
9. Future Perspectives
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Shiina, S.; Tateishi, R.; Arano, T.; Uchino, K.; Enooku, K.; Nakagawa, H.; Asaoka, Y.; Sato, T.; Masuzaki, R.; Kondo, Y.; et al. Radiofrequency ablation for hepatocellular carcinoma: 10-year outcome and prognostic factors. Am. J. Gastroenterol. 2012, 107, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Gea, V.; Toffanin, S.; Friedman, S.L.; Llovet, J.M. Role of the microenvironment in the pathogenesis and treatment of hepatocellular carcinoma. Gastroenterology 2013, 144, 512–527. [Google Scholar] [CrossRef] [PubMed]
- Iozzo, R.V.; Schaefer, L. Proteoglycan form and function: A comprehensive nomenclature of proteoglycans. Matrix Biol. 2015, 42, 11–55. [Google Scholar] [CrossRef] [PubMed]
- Theocharis, A.D.; Skandalis, S.S.; Tzanakakis, G.N.; Karamanos, N.K. Proteoglycans in health and disease: Novel roles for proteoglycans in malignancy and their pharmacological targeting. FEBS J. 2010, 277, 3904–3923. [Google Scholar] [CrossRef] [PubMed]
- Pejler, G.; Abrink, M.; Wernersson, S. Serglycin proteoglycan: Regulating the storage and activities of hematopoietic proteases. Biofactors 2009, 35, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Korpetinou, A.; Papachristou, D.J.; Lampropoulou, A.; Bouris, P.; Labropoulou, V.T.; Noulas, A.; Karamanos, N.K.; Theocharis, A.D. Increased Expression of Serglycin in Specific Carcinomas and Aggressive Cancer Cell Lines. BioMed Res. Int. 2015, 2015, 690721. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.Y.; Hsu, H.S.; Tyan, S.W.; Li, F.Y.; Shew, J.Y.; Lee, W.H.; Chen, J.Y. Serglycin in tumor microenvironment promotes non-small cell lung cancer aggressiveness in a CD44-dependent manner. Oncogene 2017, 36, 2457–2471. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Zhou, X.; Qu, C.; Tang, Y.; Zhang, Q.; Hong, J. Serglycin (SRGN) overexpression predicts poor prognosis in hepatocellular carcinoma patients. Med. Oncol. 2013, 30, 707. [Google Scholar] [CrossRef] [PubMed]
- Theocharis, A.D.; Seidel, C.; Borset, M.; Dobra, K.; Baykov, V.; Labropoulou, V.; Kanakis, I.; Dalas, E.; Karamanos, N.K.; Sundan, A.; et al. Serglycin constitutively secreted by myeloma plasma cells is a potent inhibitor of bone mineralization in vitro. J. Biol. Chem. 2006, 281, 35116–35128. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Zeng, Z.C.; Xiang, Z.L.; Yang, P. Mass spectrometry-based serum peptide profiling in hepatocellular carcinoma with bone metastasis. World J. Gastroenterol. 2014, 20, 3025–3032. [Google Scholar] [CrossRef] [PubMed]
- Chu, Q.; Huang, H.; Huang, T.; Cao, L.; Peng, L.; Shi, S.; Zheng, L.; Xu, L.; Zhang, S.; Huang, J.; et al. Extracellular serglycin upregulates the CD44 receptor in an autocrine manner to maintain self-renewal in nasopharyngeal carcinoma cells by reciprocally activating the MAPK/β-catenin axis. Cell Death Dis. 2016, 7, e2456. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Deng, Y.; Zheng, G.; Jia, X.; Xiong, Y.; Luo, K.; Qiu, Q.; Qiu, N.; Yin, J.; Lu, M.; et al. SRGN-TGFβ2 regulatory loop confers invasion and metastasis in triple-negative breast cancer. Oncogenesis 2017, 6, e360. [Google Scholar] [CrossRef] [PubMed]
- Saunders, S.; Jalkanen, M.; O’Farrell, S.; Bernfield, M. Molecular cloning of syndecan, an integral membrane proteoglycan. J. Cell. Biol. 1989, 108, 1547–1556. [Google Scholar] [CrossRef] [PubMed]
- Tatrai, P.; Egedi, K.; Somoracz, A.; van Kuppevelt, T.H.; Ten Dam, G.; Lyon, M.; Deakin, J.A.; Kiss, A.; Schaff, Z.; Kovalszky, I. Quantitative and qualitative alterations of heparan sulfate in fibrogenic liver diseases and hepatocellular cancer. J. Histochem. Cytochem. 2010, 58, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, A.; Ono, M.; Fujimoto, Y.; Gallo, R.L.; Bernfield, M.; Kohgo, Y. Reduced expression of syndecan-1 in human hepatocellular carcinoma with high metastatic potential. Int. J. Cancer 1997, 74, 482–491. [Google Scholar] [CrossRef]
- Li, H.G.; Xie, D.R.; Shen, X.M.; Li, H.H.; Zeng, H.; Zeng, Y.J. Clinicopathological significance of expression of paxillin, syndecan-1 and EMMPRIN in hepatocellular carcinoma. World J. Gastroenterol. 2005, 11, 1445–1451. [Google Scholar] [CrossRef] [PubMed]
- Ding, K.; Lopez-Burks, M.; Sanchez-Duran, J.A.; Korc, M.; Lander, A.D. Growth factor-induced shedding of syndecan-1 confers glypican-1 dependence on mitogenic responses of cancer cells. J. Cell. Biol. 2005, 171, 729–738. [Google Scholar] [CrossRef] [PubMed]
- Zvibel, I.; Halfon, P.; Fishman, S.; Penaranda, G.; Leshno, M.; Or, A.B.; Halpern, Z.; Oren, R. Syndecan 1 (CD138) serum levels: A novel biomarker in predicting liver fibrosis stage in patients with hepatitis C. Liver Int. 2009, 29, 208–212. [Google Scholar] [CrossRef] [PubMed]
- Metwaly, H.A.; Al-Gayyar, M.M.; Eletreby, S.; Ebrahim, M.A.; El-Shishtawy, M.M. Relevance of serum levels of interleukin-6 and syndecan-1 in patients with hepatocellular carcinoma. Sci. Pharm. 2012, 80, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Nault, J.C.; Guyot, E.; Laguillier, C.; Chevret, S.; Ganne-Carrie, N.; N’Kontchou, G.; Beaugrand, M.; Seror, O.; Trinchet, J.C.; Coelho, J.; et al. Serum proteoglycans as prognostic biomarkers of hepatocellular carcinoma in patients with alcoholic cirrhosis. Cancer Epidemiol. Biomark. Prev. 2013, 22, 1343–1352. [Google Scholar] [CrossRef] [PubMed]
- Bass, M.D.; Morgan, M.R.; Humphries, M.J. Syndecans shed their reputation as inert molecules. Sci. Signal. 2009, 64, pe18. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Yao, X.; Chen, L.; Yan, Z.; Liu, J.; Zhang, Y.; Feng, T.; Wu, J.; Liu, X. Sphingosine-1-phosphate induced epithelial-mesenchymal transition of hepatocellular carcinoma via an MMP-7/syndecan-1/TGF-beta autocrine loop. Oncotarget 2016, 7, 63324–63337. [Google Scholar] [CrossRef] [PubMed]
- Bao, M.; Chen, Z.; Xu, Y.; Zhao, Y.; Zha, R.; Huang, S.; Liu, L.; Chen, T.; Li, J.; Tu, H.; et al. Sphingosine kinase 1 promotes tumour cell migration and invasion via the S1P/EDG1 axis in hepatocellular carcinoma. Liver Int. 2012, 32, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Wu, X.Z. Heparanase and hepatocellular carcinoma: Promoter or inhibitor? World J. Gastroenterol. 2010, 16, 306–311. [Google Scholar] [CrossRef] [PubMed]
- Ramani, V.C.; Purushothaman, A.; Stewart, M.D.; Thompson, C.A.; Vlodavsky, I.; Au, J.L.; Sanderson, R.D. The heparanase/syndecan-1 axis in cancer: Mechanisms and therapies. FEBS J. 2013, 280, 2294–2306. [Google Scholar] [CrossRef] [PubMed]
- Rapraeger, A.C. Synstatin: A selective inhibitor of the syndecan-1-coupled IGF1R-αvβ3 integrin complex in tumorigenesis and angiogenesis. FEBS J. 2013, 280, 2207–2215. [Google Scholar] [CrossRef] [PubMed]
- Beauvais, D.M.; Ell, B.J.; McWhorter, A.R.; Rapraeger, A.C. Syndecan-1 regulates αvβ3 and αvβ5 integrin activation during angiogenesis and is blocked by synstatin, a novel peptide inhibitor. J. Exp. Med. 2009, 206, 691–705. [Google Scholar] [CrossRef] [PubMed]
- Metwaly, H.A.; El-Gayar, A.M.; El-Shishtawy, M.M. Inhibition of the signaling pathway of syndecan-1 by synstatin: A promising anti-integrin inhibitor of angiogenesis and proliferation in HCC in rats. Arch. Biochem. Biophys. 2018, 652, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Okabe, H.; Satoh, S.; Kato, T.; Kitahara, O.; Yanagawa, R.; Yamaoka, Y.; Tsunoda, T.; Furukawa, Y.; Nakamura, Y. Genome-wide analysis of gene expression in human hepatocellular carcinomas using cDNA microarray: Identification of genes involved in viral carcinogenesis and tumor progression. Cancer Res. 2001, 61, 2129–2137. [Google Scholar] [PubMed]
- Midorikawa, Y.; Ishikawa, S.; Iwanari, H.; Imamura, T.; Sakamoto, H.; Miyazono, K.; Kodama, T.; Makuuchi, M.; Aburatani, H. Glypican-3, overexpressed in hepatocellular carcinoma, modulates FGF2 and BMP-7 signaling. Int. J. Cancer 2003, 103, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Sung, Y.K.; Hwang, S.Y.; Park, M.K.; Farooq, M.; Han, I.S.; Bae, H.I.; Kim, J.C.; Kim, M. Glypican-3 is overexpressed in human hepatocellular carcinoma. Cancer Sci. 2003, 94, 259–262. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.W.; Friess, H.; Wang, L.; Abou-Shady, M.; Zimmermann, A.; Lander, A.D.; Korc, M.; Kleeff, J.; Buchler, M.W. Enhanced glypican-3 expression differentiates the majority of hepatocellular carcinomas from benign hepatic disorders. Gut 2001, 48, 558–564. [Google Scholar] [CrossRef] [PubMed]
- Capurro, M.; Wanless, I.R.; Sherman, M.; Deboer, G.; Shi, W.; Miyoshi, E.; Filmus, J. Glypican-3: A novel serum and histochemical marker for hepatocellular carcinoma. Gastroenterology 2003, 125, 89–97. [Google Scholar] [CrossRef]
- Libbrecht, L.; Severi, T.; Cassiman, D.; Vander Borght, S.; Pirenne, J.; Nevens, F.; Verslype, C.; van Pelt, J.; Roskams, T. Glypican-3 expression distinguishes small hepatocellular carcinomas from cirrhosis, dysplastic nodules, and focal nodular hyperplasia-like nodules. Am. J. Surg. Pathol. 2006, 30, 1405–1411. [Google Scholar] [CrossRef] [PubMed]
- Di Tommaso, L.; Franchi, G.; Park, Y.N.; Fiamengo, B.; Destro, A.; Morenghi, E.; Montorsi, M.; Torzilli, G.; Tommasini, M.; Terracciano, L.; et al. Diagnostic value of HSP70, glypican 3, and glutamine synthetase in hepatocellular nodules in cirrhosis. Hepatology 2007, 45, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Di Tommaso, L.; Destro, A.; Seok, J.Y.; Balladore, E.; Terracciano, L.; Sangiovanni, A.; Iavarone, M.; Colombo, M.; Jang, J.J.; Yu, E.; et al. The application of markers (HSP70 GPC3 and GS) in liver biopsies is useful for detection of hepatocellular carcinoma. J. Hepatol. 2009, 50, 746–754. [Google Scholar] [CrossRef] [PubMed]
- Shirakawa, H.; Suzuki, H.; Shimomura, M.; Kojima, M.; Gotohda, N.; Takahashi, S.; Nakagohri, T.; Konishi, M.; Kobayashi, N.; Kinoshita, T.; et al. Glypican-3 expression is correlated with poor prognosis in hepatocellular carcinoma. Cancer Sci. 2009, 100, 1403–1407. [Google Scholar] [CrossRef] [PubMed]
- Kaseb, A.O.; Hassan, M.; Lacin, S.; Abdel-Wahab, R.; Amin, H.M.; Shalaby, A.; Wolff, R.A.; Yao, J.; Rashid, A.; Vennapusa, B.; et al. Evaluating clinical and prognostic implications of Glypican-3 in hepatocellular carcinoma. Oncotarget 2016, 7, 69916–69926. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.J.; Qi, C.Y.; Xiao, W.K.; Li, S.Q.; Peng, B.G.; Liang, L.J. Glypican-3 is a potential prognostic biomarker for hepatocellular carcinoma after curative resection. Surgery 2013, 154, 536–544. [Google Scholar] [CrossRef] [PubMed]
- Ning, S.; Bin, C.; Na, H.; Peng, S.; Yi, D.; Xiang-hua, Y.; Fang-yin, Z.; Da-yong, Z.; Rong-cheng, L. Glypican-3, a novel prognostic marker of hepatocellular cancer, is related with postoperative metastasis and recurrence in hepatocellular cancer patients. Mol. Biol. Rep. 2012, 39, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.P.; Ariizumi, S.; Nakano, M.; Yamamoto, M. Positive glypican-3 expression in early hepatocellular carcinoma predicts recurrence after hepatectomy. J. Gastroenterol. 2014, 49, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.L.; Zhu, Z.J.; Teng, D.H.; Yao, Z.; Gao, W.; Shen, Z.Y. Glypican-3 expression and its relationship with recurrence of HCC after liver transplantation. World J. Gastroenterol. 2012, 18, 2408–2414. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, M.; Ma, H.; Song, X.; He, L.; Ye, X.; Li, X. Overexpression of glypican-3 is a predictor of poor prognosis in hepatocellular carcinoma: An updated meta-analysis. Medicine 2018, 97, e11130. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Yang, C.; Lu, W.; Zeng, Y. Prognostic significance of glypican-3 expression in hepatocellular carcinoma: A meta-analysis. Medicine 2018, 97, e9702. [Google Scholar] [CrossRef] [PubMed]
- Hippo, Y.; Watanabe, K.; Watanabe, A.; Midorikawa, Y.; Yamamoto, S.; Ihara, S.; Tokita, S.; Iwanari, H.; Ito, Y.; Nakano, K.; et al. Identification of soluble NH2-terminal fragment of glypican-3 as a serological marker for early-stage hepatocellular carcinoma. Cancer Res. 2004, 64, 2418–2423. [Google Scholar] [CrossRef] [PubMed]
- Traister, A.; Shi, W.; Filmus, J. Mammalian Notum induces the release of glypicans and other GPI-anchored proteins from the cell surface. Biochem. J. 2008, 410, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Nakatsura, T.; Yoshitake, Y.; Senju, S.; Monji, M.; Komori, H.; Motomura, Y.; Hosaka, S.; Beppu, T.; Ishiko, T.; Kamohara, H.; et al. Glypican-3, overexpressed specifically in human hepatocellular carcinoma, is a novel tumor marker. Biochem. Biophys. Res. Commun. 2003, 306, 16–25. [Google Scholar] [CrossRef]
- Jia, X.; Liu, J.; Gao, Y.; Huang, Y.; Du, Z. Diagnosis accuracy of serum glypican-3 in patients with hepatocellular carcinoma: A systematic review with meta-analysis. Arch. Med. Res. 2014, 45, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Capurro, M.I.; Xu, P.; Shi, W.; Li, F.; Jia, A.; Filmus, J. Glypican-3 inhibits Hedgehog signaling during development by competing with patched for Hedgehog binding. Dev. Cell. 2008, 14, 700–711. [Google Scholar] [CrossRef] [PubMed]
- Cano-Gauci, D.F.; Song, H.H.; Yang, H.; McKerlie, C.; Choo, B.; Shi, W.; Pullano, R.; Piscione, T.D.; Grisaru, S.; Soon, S.; et al. Glypican-3-deficient mice exhibit developmental overgrowth and some of the abnormalities typical of Simpson-Golabi-Behmel syndrome. J. Cell. Biol. 1999, 146, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Chen, N.; Chen, Y.; Sun, L.; Li, L.; Liu, H. Elevated GPC3 level promotes cell proliferation in liver cancer. Oncol. Lett. 2018, 16, 970–976. [Google Scholar] [CrossRef] [PubMed]
- Capurro, M.I.; Xiang, Y.Y.; Lobe, C.; Filmus, J. Glypican-3 promotes the growth of hepatocellular carcinoma by stimulating canonical Wnt signaling. Cancer Res. 2005, 65, 6245–6254. [Google Scholar] [CrossRef] [PubMed]
- Capurro, M.; Martin, T.; Shi, W.; Filmus, J. Glypican-3 binds to Frizzled and plays a direct role in the stimulation of canonical Wnt signaling. J. Cell Sci. 2014, 127, 1565–1575. [Google Scholar] [CrossRef] [PubMed]
- Pez, F.; Lopez, A.; Kim, M.; Wands, J.R.; Caron de Fromentel, C.; Merle, P. Wnt signaling and hepatocarcinogenesis: Molecular targets for the development of innovative anticancer drugs. J. Hepatol. 2013, 59, 1107–1117. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Tseng, C.J.; Lin, T.T.; Cheng, I.; Pan, H.W.; Hsu, H.C.; Lee, Y.M. Glypican-3-mediated oncogenesis involves the Insulin-like growth factor-signaling pathway. Carcinogenesis 2008, 29, 1319–1326. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Liu, H.; Weng, H.; Zhang, X.; Li, P.; Fan, C.L.; Li, B.; Dong, P.L.; Li, L.; Dooley, S.; et al. Glypican-3 promotes epithelial-mesenchymal transition of hepatocellular carcinoma cells through ERK signaling pathway. Int. J. Oncol. 2015, 46, 1275–1285. [Google Scholar] [CrossRef] [PubMed]
- Ishiguro, T.; Sugimoto, M.; Kinoshita, Y.; Miyazaki, Y.; Nakano, K.; Tsunoda, H.; Sugo, I.; Ohizumi, I.; Aburatani, H.; Hamakubo, T.; et al. Anti-glypican 3 antibody as a potential antitumor agent for human liver cancer. Cancer Res. 2008, 68, 9832–9838. [Google Scholar] [CrossRef] [PubMed]
- Nakano, K.; Orita, T.; Nezu, J.; Yoshino, T.; Ohizumi, I.; Sugimoto, M.; Furugaki, K.; Kinoshita, Y.; Ishiguro, T.; Hamakubo, T.; et al. Anti-glypican 3 antibodies cause ADCC against human hepatocellular carcinoma cells. Biochem. Biophys. Res. Commun. 2009, 378, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Zhu, A.X.; Gold, P.J.; El-Khoueiry, A.B.; Abrams, T.A.; Morikawa, H.; Ohishi, N.; Ohtomo, T.; Philip, P.A. First-in-man phase I study of GC33, a novel recombinant humanized antibody against glypican-3, in patients with advanced hepatocellular carcinoma. Clin. Cancer Res. 2013, 19, 920–928. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, M.; Ohkawa, S.; Okusaka, T.; Mitsunaga, S.; Kobayashi, S.; Morizane, C.; Suzuki, I.; Yamamoto, S.; Furuse, J. Japanese phase I study of GC33, a humanized antibody against glypican-3 for advanced hepatocellular carcinoma. Cancer Sci. 2014, 105, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Nakatsura, T.; Komori, H.; Kubo, T.; Yoshitake, Y.; Senju, S.; Katagiri, T.; Furukawa, Y.; Ogawa, M.; Nakamura, Y.; Nishimura, Y. Mouse homologue of a novel human oncofetal antigen, glypican-3, evokes T-cell-mediated tumor rejection without autoimmune reactions in mice. Clin. Cancer Res. 2004, 10, 8630–8640. [Google Scholar] [CrossRef] [PubMed]
- Komori, H.; Nakatsura, T.; Senju, S.; Yoshitake, Y.; Motomura, Y.; Ikuta, Y.; Fukuma, D.; Yokomine, K.; Harao, M.; Beppu, T.; et al. Identification of HLA-A2- or HLA-A24-restricted CTL epitopes possibly useful for glypican-3-specific immunotherapy of hepatocellular carcinoma. Clin. Cancer Res. 2006, 12, 2689–2697. [Google Scholar] [CrossRef] [PubMed]
- Motomura, Y.; Ikuta, Y.; Kuronuma, T.; Komori, H.; Ito, M.; Tsuchihara, M.; Tsunoda, Y.; Shirakawa, H.; Baba, H.; Nishimura, Y.; et al. HLA-A2 and -A24-restricted glypican-3-derived peptide vaccine induces specific CTLs: Preclinical study using mice. Int. J. Oncol. 2008, 32, 985–990. [Google Scholar] [CrossRef] [PubMed]
- Sawada, Y.; Yoshikawa, T.; Nobuoka, D.; Shirakawa, H.; Kuronuma, T.; Motomura, Y.; Mizuno, S.; Ishii, H.; Nakachi, K.; Konishi, M.; et al. Phase I trial of a glypican-3-derived peptide vaccine for advanced hepatocellular carcinoma: Immunologic evidence and potential for improving overall survival. Clin. Cancer Res. 2012, 18, 3686–3696. [Google Scholar] [CrossRef] [PubMed]
- Sawada, Y.; Yoshikawa, T.; Ofuji, K.; Yoshimura, M.; Tsuchiya, N.; Takahashi, M.; Nobuoka, D.; Gotohda, N.; Takahashi, S.; Kato, Y.; et al. Phase II study of the GPC3-derived peptide vaccine as an adjuvant therapy for hepatocellular carcinoma patients. Oncoimmunology 2016, 5, e1129483. [Google Scholar] [CrossRef] [PubMed]
- Jackson, H.J.; Rafiq, S.; Brentjens, R.J. Driving CAR T-cells forward. Nat Rev Clin Oncol 2016, 13, 370–383. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Li, K.; Tu, H.; Pan, X.; Jiang, H.; Shi, B.; Kong, J.; Wang, H.; Yang, S.; Gu, J.; et al. Development of T cells redirected to glypican-3 for the treatment of hepatocellular carcinoma. Clin. Cancer Res. 2014, 20, 6418–6428. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Li, K.; Jiang, H.; Song, F.; Gao, H.; Pan, X.; Shi, B.; Bi, Y.; Wang, H.; Wang, H.; et al. Development of T cells carrying two complementary chimeric antigen receptors against glypican-3 and asialoglycoprotein receptor 1 for the treatment of hepatocellular carcinoma. Cancer Immunol. Immunother. 2017, 66, 475–489. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Di, S.; Shi, B.; Jiang, H.; Shi, Z.; Liu, Y.; Wang, Y.; Luo, H.; Yu, M.; Wu, X.; et al. Increased antitumor activities of glypican-3-specific chimeric antigen receptor-modified T cells by coexpression of a soluble PD1-CH3 fusion protein. Cancer Immunol. Immunother. 2018, 67, 1621–1634. [Google Scholar] [CrossRef] [PubMed]
- Bezakova, G.; Ruegg, M.A. New insights into the roles of agrin. Nat. Rev. Mol. Cell Biol. 2003, 4, 295–308. [Google Scholar] [CrossRef] [PubMed]
- Gautam, M.; Noakes, P.G.; Moscoso, L.; Rupp, F.; Scheller, R.H.; Merlie, J.P.; Sanes, J.R. Defective neuromuscular synaptogenesis in agrin-deficient mutant mice. Cell 1996, 85, 525–535. [Google Scholar] [CrossRef]
- Tatrai, P.; Dudas, J.; Batmunkh, E.; Mathe, M.; Zalatnai, A.; Schaff, Z.; Ramadori, G.; Kovalszky, I. Agrin, a novel basement membrane component in human and rat liver, accumulates in cirrhosis and hepatocellular carcinoma. Lab. Investig. 2006, 86, 1149–1160. [Google Scholar] [CrossRef] [PubMed]
- Batmunkh, E.; Tatrai, P.; Szabo, E.; Lodi, C.; Holczbauer, A.; Paska, C.; Kupcsulik, P.; Kiss, A.; Schaff, Z.; Kovalszky, I. Comparison of the expression of agrin, a basement membrane heparan sulfate proteoglycan, in cholangiocarcinoma and hepatocellular carcinoma. Hum. Pathol. 2007, 38, 1508–1515. [Google Scholar] [CrossRef] [PubMed]
- Tatrai, P.; Somoracz, A.; Batmunkh, E.; Schirmacher, P.; Kiss, A.; Schaff, Z.; Nagy, P.; Kovalszky, I. Agrin and CD34 immunohistochemistry for the discrimination of benign versus malignant hepatocellular lesions. Am. J. Surg. Pathol. 2009, 33, 874–885. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Lakshmanan, M.; Swa, H.L.; Chen, J.; Zhang, X.; Ong, Y.S.; Loo, L.S.; Akincilar, S.C.; Gunaratne, J.; Tergaonkar, V.; et al. An oncogenic role of Agrin in regulating focal adhesion integrity in hepatocellular carcinoma. Nat. Commun. 2015, 6, 6184. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.; Fang, C.; Yin, R.; Qiao, B.; Shang, R.; Wang, J.; Song, W.; He, Y.; Chen, Y. Agrin para-secreted by PDGF-activated human hepatic stellate cells promotes hepatocarcinogenesis in vitro and in vivo. Oncotarget 2017, 8, 105340–105355. [Google Scholar] [CrossRef] [PubMed]
- Seppinen, L.; Pihlajaniemi, T. The multiple functions of collagen XVIII in development and disease. Matrix Biol. 2011, 30, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Muragaki, Y.; Timmons, S.; Griffith, C.M.; Oh, S.P.; Fadel, B.; Quertermous, T.; Olsen, B.R. Mouse Col18a1 is expressed in a tissue-specific manner as three alternative variants and is localized in basement membrane zones. Proc. Natl. Acad. Sci. USA 1995, 92, 8763–8767. [Google Scholar] [CrossRef] [PubMed]
- Lietard, J.; Theret, N.; Rehn, M.; Musso, O.; Dargere, D.; Pihlajaniemi, T.; Clement, B. The promoter of the long variant of collagen XVIII, the precursor of endostatin, contains liver-specific regulatory elements. Hepatology 2000, 32, 1377–1385. [Google Scholar] [CrossRef] [PubMed]
- Musso, O.; Rehn, M.; Saarela, J.; Theret, N.; Lietard, J.; Hintikka, E.; Lotrian, D.; Campion, J.P.; Pihlajaniemi, T.; Clement, B. Collagen XVIII is localized in sinusoids and basement membrane zones and expressed by hepatocytes and activated stellate cells in fibrotic human liver. Hepatology 1998, 28, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Musso, O.; Theret, N.; Heljasvaara, R.; Rehn, M.; Turlin, B.; Campion, J.P.; Pihlajaniemi, T.; Clement, B. Tumor hepatocytes and basement membrane-Producing cells specifically express two different forms of the endostatin precursor, collagen XVIII, in human liver cancers. Hepatology 2001, 33, 868–876. [Google Scholar] [CrossRef] [PubMed]
- Musso, O.; Rehn, M.; Theret, N.; Turlin, B.; Bioulac-Sage, P.; Lotrian, D.; Campion, J.P.; Pihlajaniemi, T.; Clement, B. Tumor progression is associated with a significant decrease in the expression of the endostatin precursor collagen XVIII in human hepatocellular carcinomas. Cancer Res. 2001, 61, 45–49. [Google Scholar] [PubMed]
- Ferreras, M.; Felbor, U.; Lenhard, T.; Olsen, B.R.; Delaisse, J. Generation and degradation of human endostatin proteins by various proteinases. FEBS Lett. 2000, 486, 247–251. [Google Scholar] [CrossRef]
- O’Reilly, M.S.; Boehm, T.; Shing, Y.; Fukai, N.; Vasios, G.; Lane, W.S.; Flynn, E.; Birkhead, J.R.; Olsen, B.R.; Folkman, J. Endostatin: An endogenous inhibitor of angiogenesis and tumor growth. Cell 1997, 88, 277–285. [Google Scholar] [CrossRef]
- Kim, Y.M.; Hwang, S.; Kim, Y.M.; Pyun, B.J.; Kim, T.Y.; Lee, S.T.; Gho, Y.S.; Kwon, Y.G. Endostatin blocks vascular endothelial growth factor-mediated signaling via direct interaction with KDR/Flk-1. J. Biol. Chem. 2002, 277, 27872–27879. [Google Scholar] [CrossRef] [PubMed]
- Reis, R.C.; Schuppan, D.; Barreto, A.C.; Bauer, M.; Bork, J.P.; Hassler, G.; Coelho-Sampaio, T. Endostatin competes with bFGF for binding to heparin-like glycosaminoglycans. Biochem. Biophys. Res. Commun. 2005, 333, 976–983. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.H.; Huang, C.C.; Wu, C.L.; Lin, P.R.; Liu, S.Y.; Lin, J.W.; Chuang, J.H.; Tai, M.H. Increased endostatin/collagen XVIII expression correlates with elevated VEGF level and poor prognosis in hepatocellular carcinoma. Mod. Pathol. 2005, 18, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Dhar, D.K.; Ono, T.; Yamanoi, A.; Soda, Y.; Yamaguchi, E.; Rahman, M.A.; Kohno, H.; Nagasue, N. Serum endostatin predicts tumor vascularity in hepatocellular carcinoma. Cancer 2002, 95, 2188–2195. [Google Scholar] [CrossRef] [PubMed]
- Poon, R.T.; Ho, J.W.; Tong, C.S.; Lau, C.; Ng, I.O.; Fan, S.T. Prognostic significance of serum vascular endothelial growth factor and endostatin in patients with hepatocellular carcinoma. Br. J. Surg. 2004, 91, 1354–1360. [Google Scholar] [CrossRef] [PubMed]
- Feldman, A.L.; Alexander, H.R., Jr.; Bartlett, D.L.; Kranda, K.C.; Miller, M.S.; Costouros, N.G.; Choyke, P.L.; Libutti, S.K. A prospective analysis of plasma endostatin levels in colorectal cancer patients with liver metastases. Ann. Surg. Oncol. 2001, 8, 741–745. [Google Scholar] [CrossRef] [PubMed]
- Graepler, F.; Verbeek, B.; Graeter, T.; Smirnow, I.; Kong, H.L.; Schuppan, D.; Bauer, M.; Vonthein, R.; Gregor, M.; Lauer, U.M. Combined endostatin/sFlt-1 antiangiogenic gene therapy is highly effective in a rat model of HCC. Hepatology 2005, 41, 879–886. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Sham, J.; Yang, J.; Su, C.; Xue, H.; Chua, D.; Sun, L.; Zhang, Q.; Cui, Z.; Wu, M.; et al. Potent antitumor efficacy of an E1B 55kDa-deficient adenovirus carrying murine endostatin in hepatocellular carcinoma. Int. J. Cancer 2005, 113, 640–648. [Google Scholar] [CrossRef] [PubMed]
- Fu, G.F.; Li, X.; Hou, Y.Y.; Fan, Y.R.; Liu, W.H.; Xu, G.X. Bifidobacterium longum as an oral delivery system of endostatin for gene therapy on solid liver cancer. Cancer Gene. Ther. 2005, 12, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Tan, G.; Li, J.; Dong, X.; Krissansen, G.W.; Sun, X. Gene transfer of endostatin enhances the efficacy of doxorubicin to suppress human hepatocellular carcinomas in mice. Cancer Sci. 2007, 98, 1381–1387. [Google Scholar] [CrossRef] [PubMed]
- Ling, Y.; Yang, Y.; Lu, N.; You, Q.D.; Wang, S.; Gao, Y.; Chen, Y.; Guo, Q.L. Endostar, a novel recombinant human endostatin, exerts antiangiogenic effect via blocking VEGF-induced tyrosine phosphorylation of KDR/Flk-1 of endothelial cells. Biochem. Biophys. Res. Commun. 2007, 361, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wang, J.W.; Liu, Y.Y.; Yu, Q.T.; Zhang, Y.P.; Li, K.; Xu, L.Y.; Luo, S.X.; Qin, F.Z.; Chen, Z.T.; et al. Long-term results of a randomized, double-blind, and placebo-controlled phase III trial: Endostar (rh-endostatin) versus placebo in combination with vinorelbine and cisplatin in advanced non-small cell lung cancer. Thorac. Cancer 2013, 4, 440–448. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Qin, S.; Liu, Y.; Feng, J.; Wu, Q.; Qu, W.; Yin, X. Inhibitory effect of endostar on specific angiogenesis induced by human hepatocellular carcinoma. Gastroenterol. Res. Pract. 2015, 2015, 957574. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Gu, J.; Lv, Y.; Yuan, J.; Yang, N.; Chen, J.; Wang, C.; Hou, X.; Jia, X.; Feng, L.; et al. Angiogenesis for tumor vascular normalization of Endostar on hepatoma 22 tumor-bearing mice is involved in the immune response. Oncol. Lett. 2018, 15, 3437–3446. [Google Scholar] [CrossRef] [PubMed]
- Du, W.W.; Yang, W.; Yee, A.J. Roles of versican in cancer biology—Tumorigenesis, progression and metastasis. Histol. Histopathol. 2013, 28, 701–713. [Google Scholar] [PubMed]
- Schmalfeldt, M.; Dours-Zimmermann, M.T.; Winterhalter, K.H.; Zimmermann, D.R. Versican V2 is a major extracellular matrix component of the mature bovine brain. J. Biol. Chem. 1998, 273, 15758–15764. [Google Scholar] [CrossRef] [PubMed]
- Bukong, T.N.; Maurice, S.B.; Chahal, B.; Schaeffer, D.F.; Winwood, P.J. Versican: A novel modulator of hepatic fibrosis. Lab. Investig. 2016, 96, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Ramnath, D.; Irvine, K.M.; Lukowski, S.W.; Horsfall, L.U.; Loh, Z.; Clouston, A.D.; Patel, P.J.; Fagan, K.J.; Iyer, A.; Lampe, G.; et al. Hepatic expression profiling identifies steatosis-independent and steatosis-driven advanced fibrosis genes. JCI Insight 2018, 3, e120274. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Huang, W.; Tian, D.; Zhang, L.; Qi, X.; Chen, Z.; Shang, X.; Nie, Y.; Wu, K. Forkhead box Q1 promotes hepatocellular carcinoma metastasis by transactivating ZEB2 and VersicanV1 expression. Hepatology 2014, 59, 958–973. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Tateishi, K.; Nakatsuka, T.; Kudo, Y.; Takahashi, R.; Miyabayashi, K.; Yamamoto, K.; Asaoka, Y.; Ijichi, H.; Tateishi, R.; et al. Sharpin promotes hepatocellular carcinoma progression via transactivation of Versican expression. Oncogenesis 2016, 5, e277. [Google Scholar] [CrossRef] [PubMed]
- Naboulsi, W.; Megger, D.A.; Bracht, T.; Kohl, M.; Turewicz, M.; Eisenacher, M.; Voss, D.M.; Schlaak, J.F.; Hoffmann, A.C.; Weber, F.; et al. Quantitative Tissue Proteomics Analysis Reveals Versican as Potential Biomarker for Early-Stage Hepatocellular Carcinoma. J. Proteome Res. 2016, 15, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Ricciardelli, C.; Sakko, A.J.; Ween, M.P.; Russell, D.L.; Horsfall, D.J. The biological role and regulation of versican levels in cancer. Cancer Metastasis Rev. 2009, 28, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cao, L.; Yang, B.L.; Yang, B.B. The G3 domain of versican enhances cell proliferation via epidermial growth factor-like motifs. J. Biol. Chem. 1998, 273, 21342–21351. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.L.; Zhang, Y.; Cao, L.; Yang, B.B. Cell adhesion and proliferation mediated through the G1 domain of versican. J. Cell. Biochem. 1999, 72, 210–220. [Google Scholar] [CrossRef]
- Cattaruzza, S.; Schiappacassi, M.; Kimata, K.; Colombatti, A.; Perris, R. The globular domains of PG-M/versican modulate the proliferation-apoptosis equilibrium and invasive capabilities of tumor cells. FASEB J. 2004, 18, 779–781. [Google Scholar] [CrossRef] [PubMed]
- Sheng, W.; Wang, G.; Wang, Y.; Liang, J.; Wen, J.; Zheng, P.S.; Wu, Y.; Lee, V.; Slingerland, J.; Dumont, D.; et al. The roles of versican V1 and V2 isoforms in cell proliferation and apoptosis. Mol. Biol. Cell 2005, 16, 1330–1340. [Google Scholar] [CrossRef] [PubMed]
- Sakko, A.J.; Ricciardelli, C.; Mayne, K.; Suwiwat, S.; LeBaron, R.G.; Marshall, V.R.; Tilley, W.D.; Horsfall, D.J. Modulation of prostate cancer cell attachment to matrix by versican. Cancer Res. 2003, 63, 4786–4791. [Google Scholar] [PubMed]
- Ricciardelli, C.; Russell, D.L.; Ween, M.P.; Mayne, K.; Suwiwat, S.; Byers, S.; Marshall, V.R.; Tilley, W.D.; Horsfall, D.J. Formation of hyaluronan- and versican-rich pericellular matrix by prostate cancer cells promotes cell motility. J. Biol. Chem. 2007, 282, 10814–10825. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Takahashi, H.; Lin, W.W.; Descargues, P.; Grivennikov, S.; Kim, Y.; Luo, J.L.; Karin, M. Carcinoma-produced factors activate myeloid cells through TLR2 to stimulate metastasis. Nature 2009, 457, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.; Liyanarachchi, S.; Wright, F.A.; Davuluri, R.; Lockman, J.C.; de la Chapelle, A.; Pellegata, N.S. Gene expression profiling of isogenic cells with different TP53 gene dosage reveals numerous genes that are affected by TP53 dosage and identifies CSPG2 as a direct target of p53. Proc. Natl. Acad. Sci. USA 2002, 99, 15632–15637. [Google Scholar] [CrossRef] [PubMed]
- Domenzain-Reyna, C.; Hernandez, D.; Miquel-Serra, L.; Docampo, M.J.; Badenas, C.; Fabra, A.; Bassols, A. Structure and regulation of the versican promoter: The versican promoter is regulated by AP-1 and TCF transcription factors in invasive human melanoma cells. J. Biol. Chem. 2009, 284, 12306–12317. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Kim, J.M.; Park, B.; Cheon, Y.; Lee, B.; Choo, S.H.; Koh, S.S.; Lee, S. Newly identified tumor-associated role of human Sharpin. Mol. Cell. Biochem. 2010, 340, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Tateishi, R.; Koike, K. Emerging role of SHARPIN in hepatocellular carcinoma progression. Cancer Cell Microenviron. 2017, 4, e1540. [Google Scholar]
- Morton, S.U.; Scherz, P.J.; Cordes, K.R.; Ivey, K.N.; Stainier, D.Y.; Srivastava, D. microRNA-138 modulates cardiac patterning during embryonic development. Proc. Natl. Acad. Sci. USA 2008, 105, 17830–17835. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Hu, G.; Zhou, J. Repression of versican expression by microRNA-143. J. Biol. Chem. 2010, 285, 23241–23250. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Jeyapalan, Z.; Fang, L.; Yang, J.; Zhang, Y.; Yee, A.Y.; Li, M.; Du, W.W.; Shatseva, T.; Yang, B.B. Expression of versican 3′-untranslated region modulates endogenous microRNA functions. PLoS ONE 2010, 5, e13599. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Du, W.W.; Yang, X.; Chen, K.; Ghanekar, A.; Levy, G.; Yang, W.; Yee, A.J.; Lu, W.Y.; Xuan, J.W.; et al. Versican 3′-untranslated region (3′-UTR) functions as a ceRNA in inducing the development of hepatocellular carcinoma by regulating miRNA activity. FASEB J. 2013, 27, 907–919. [Google Scholar] [CrossRef] [PubMed]
- Meyer, D.H.; Krull, N.; Dreher, K.L.; Gressner, A.M. Biglycan and decorin gene expression in normal and fibrotic rat liver: Cellular localization and regulatory factors. Hepatology 1992, 16, 204–216. [Google Scholar] [CrossRef] [PubMed]
- Dudas, J.; Kovalszky, I.; Gallai, M.; Nagy, J.O.; Schaff, Z.; Knittel, T.; Mehde, M.; Neubauer, K.; Szalay, F.; Ramadori, G. Expression of decorin, transforming growth factor-β1, tissue inhibitor metalloproteinase 1 and 2, and type IV collagenases in chronic hepatitis. Am. J. Clin. Pathol. 2001, 115, 725–735. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Mann, D.M.; Ruoslahti, E. Negative regulation of transforming growth factor-beta by the proteoglycan decorin. Nature 1990, 346, 281–284. [Google Scholar] [CrossRef] [PubMed]
- Danielson, K.G.; Baribault, H.; Holmes, D.F.; Graham, H.; Kadler, K.E.; Iozzo, R.V. Targeted disruption of decorin leads to abnormal collagen fibril morphology and skin fragility. J. Cell. Biol. 1997, 136, 729–743. [Google Scholar] [CrossRef] [PubMed]
- Baghy, K.; Dezso, K.; Laszlo, V.; Fullar, A.; Peterfia, B.; Paku, S.; Nagy, P.; Schaff, Z.; Iozzo, R.V.; Kovalszky, I. Ablation of the decorin gene enhances experimental hepatic fibrosis and impairs hepatic healing in mice. Lab. Investig. 2011, 91, 439–451. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.F.; Zhang, Q.; Cheung, P.Y.; Shi, L.; Fong, C.C.; Zhang, Y.; Tzang, C.H.; Chan, B.P.; Fong, W.F.; Chun, J.; et al. Effects of rhDecorin on TGF-beta1 induced human hepatic stellate cells LX-2 activation. Biochim. Biophys. Acta 2006, 1760, 1587–1595. [Google Scholar] [CrossRef] [PubMed]
- Horvath, Z.; Kovalszky, I.; Fullar, A.; Kiss, K.; Schaff, Z.; Iozzo, R.V.; Baghy, K. Decorin deficiency promotes hepatic carcinogenesis. Matrix Biol. 2014, 35, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Terry, S.F. Obama’s Precision Medicine Initiative. Genet. Test. Mol. Biomark. 2015, 19, 113–114. [Google Scholar] [CrossRef] [PubMed]
- Pellegatta, S.; Savoldo, B.; Di Ianni, N.; Corbetta, C.; Chen, Y.; Patane, M.; Sun, C.; Pollo, B.; Ferrone, S.; DiMeco, F.; et al. Constitutive and TNFα-inducible expression of chondroitin sulfate proteoglycan 4 in glioblastoma and neurospheres: Implications for CAR-T cell therapy. Sci. Transl. Med. 2018, 10, eaao2731. [Google Scholar] [CrossRef] [PubMed]
- Neelapu, S.S.; Tummala, S.; Kebriaei, P.; Wierda, W.; Locke, F.L.; Lin, Y.; Jain, N.; Daver, N.; Gulbis, A.M.; Adkins, S.; et al. Toxicity management after chimeric antigen receptor T cell therapy: One size does not fit ‘ALL’. Nat. Rev. Clin. Oncol. 2018, 15, 218. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Location | Classification | Eponym | Predominant GAG |
|---|---|---|---|
| Intracellular | Secretory granules | Serglycin | Hep |
| Cell surface | Transmembrane | Syndecan, 1–4 | HS |
| NG2 | CS | ||
| Betaglycan | CS/HS | ||
| Phosphacan | CS | ||
| GPI-anchored | Glypican, 1–6 | HS | |
| Pericellular | Basement membrane zone | Perlecan | HS |
| Agrin | HS | ||
| Collagen XVIII | HS | ||
| Collagen XV | CS/HS | ||
| Extracellular | Hyalectan Lectican | Aggrecan | CS/KS |
| Versican | CS | ||
| Neurocan | CS | ||
| Brevican | CS | ||
| SLRPs:canonical class I | Biglycan | CS | |
| Decorin | DS | ||
| Asporin | |||
| ECM2 | |||
| ECMX | |||
| SLRPs:canonical class II | Fibromodulin | KS | |
| Lumican | KS | ||
| PRELP | |||
| Keratocan | KS | ||
| Osteoadherin | KS | ||
| SLRPs:canonical class III | Epiphycan | DS/CS | |
| Optican | |||
| Osteoglycin | |||
| SLRPs:non-canonical class VI | Chondroadherin | ||
| Nyctalopin | |||
| Tsukushi | |||
| SLRPs:non-canonical class V | Podocan | ||
| Podocan-Like 1 | |||
| SPOCK | Testican, 1–3 | HS |
| Proteoglycan | Sample | Biomarker |
|---|---|---|
| Serglycin | Tissue | ↑ HCC with vascular invasion, advanced BCLC staging, and unfavorable prognosis [9] |
| Serum | ↑ HCC with bone metastasis [11] | |
| Syndecan-1 | Tissue | ↓ HCC with poor differentiation and high metastatic potential [16,17] |
| Serum | ↑ CLD with fibrosis [19] ↑ HCC with advanced BCLC staging [20] ↑ HCC recurrence [21] | |
| Glypican-3 * | Tissue | ↑ HCC with poor prognosis [38,39] ↑ HCC recurrence after operation [40,41,42] ↑ HCC recurrence after liver transplantation [43] |
| Serum | ↑ HCC (Independent to AFP) [34,48] | |
| Agrin | Tissue | ↑ CLD and HCC [73] |
| Serum | N/A | |
| CollagenXVIII/Endostatin | Tissue | N/A |
| Serum | N/A | |
| Versican | Tissue | ↑ CLD with fibrosis [102,103,105] ↑ HCC with poor prognosis [104,105] ↑ early-stage HCC [106] |
| Serum | ↑ CLD with fibrosis [103] | |
| Decorin | Tissue | ↑ CLD with fibrosis [123] |
| Serum | N/A |
| Proteoglycan | Therapeutic Target and Agent | |
|---|---|---|
| Serglycin | Therapeutic target | N/A |
| Agent | N/A | |
| Syndecan-1 * | Therapeutic target | ↑ MMP-7/syndecan-1/TGF-β [23] ↑ Syndecan-1-IGF1R-αvβ3 integrin complex / VEGF, bFGF [27,28,29] |
| Agent | Synstatin (Syndecan-1-IGF1R-αvβ3 integrin complex inhibitor) (Preclinical) [27,28,29] | |
| Glypican-3 ** | Therapeutic target | ↑ Wnt/Frizzled/β-catenin [53,54] ↑ IGF-II/IGF-1R/ERK [56,57] ↓ SHH/Patched-1 [52] |
| Agent | GC33(recombinant humanized monoclonal antibody) (Phase II) [60,61] Peptide vaccines (Phase II) [65,66] CAR-T therapy (preclinical) [68,69,70] | |
| Agrin | Therapeutic target | ↑ Lrp4/MuSK [76] |
| Agent | N/A | |
| CollagenXVIII/Endostatin * | Therapeutic target | ↓ VEGF [86] ↓ bFGF/FGF-2 [87] |
| Agent | Gene therapy (preclinical) [92,93,94,95] Endostar (Recombinant human endostatin) (preclinical) [98,99] | |
| Versican | Therapeutic target | ↑ Versican/CCL2 [104] |
| Agent | N/A | |
| Decorin | Therapeutic target | ↓ TGF-β1 [125,126] ↑ PDGFRα, EGFR, IGF-IR, MST1R [129] |
| Agent | N/A | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanaka, Y.; Tateishi, R.; Koike, K. Proteoglycans Are Attractive Biomarkers and Therapeutic Targets in Hepatocellular Carcinoma. Int. J. Mol. Sci. 2018, 19, 3070. https://doi.org/10.3390/ijms19103070
Tanaka Y, Tateishi R, Koike K. Proteoglycans Are Attractive Biomarkers and Therapeutic Targets in Hepatocellular Carcinoma. International Journal of Molecular Sciences. 2018; 19(10):3070. https://doi.org/10.3390/ijms19103070
Chicago/Turabian StyleTanaka, Yasuo, Ryosuke Tateishi, and Kazuhiko Koike. 2018. "Proteoglycans Are Attractive Biomarkers and Therapeutic Targets in Hepatocellular Carcinoma" International Journal of Molecular Sciences 19, no. 10: 3070. https://doi.org/10.3390/ijms19103070
APA StyleTanaka, Y., Tateishi, R., & Koike, K. (2018). Proteoglycans Are Attractive Biomarkers and Therapeutic Targets in Hepatocellular Carcinoma. International Journal of Molecular Sciences, 19(10), 3070. https://doi.org/10.3390/ijms19103070