B Type and Complex A/B Type Epicatechin Trimers Isolated from Litchi pericarp Aqueous Extract Show High Antioxidant and Anticancer Activity


Abstract
1. Introduction
2. Results and Discussion
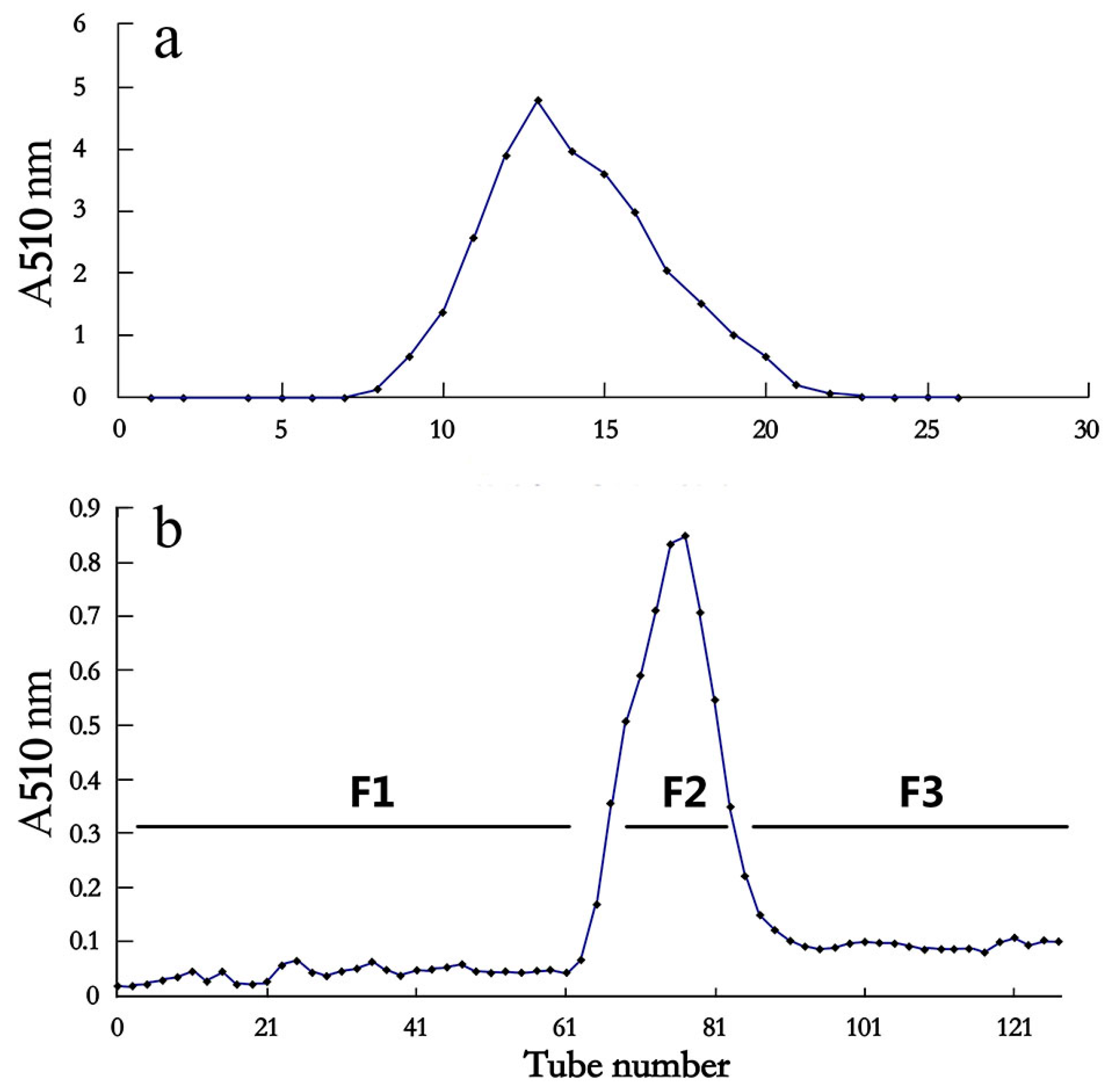
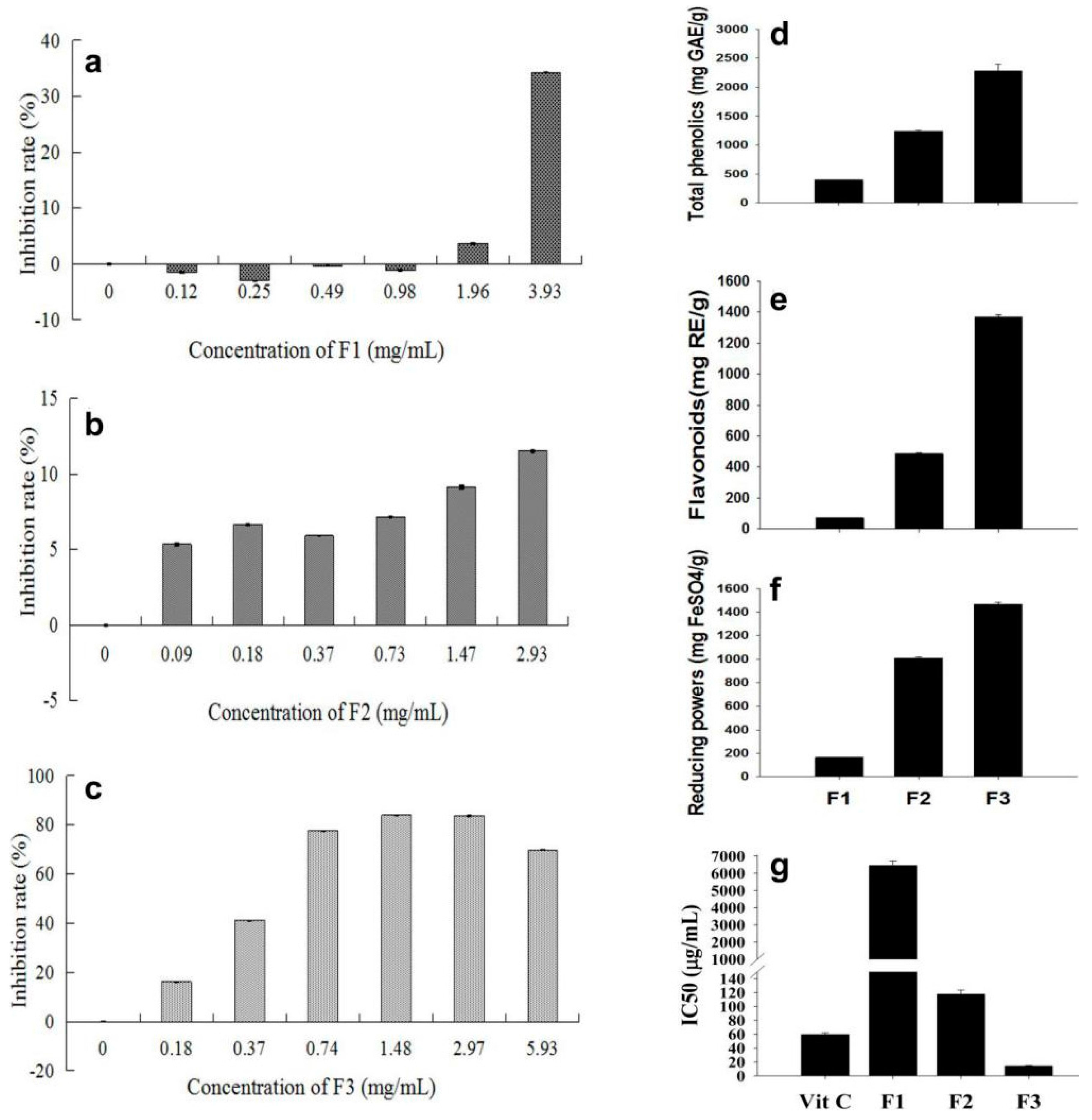
2.1. A Fraction from Litchi phenolic Extract Displayed Strong Inhibition on Cancer Cell Proliferation and Antioxidant Activity
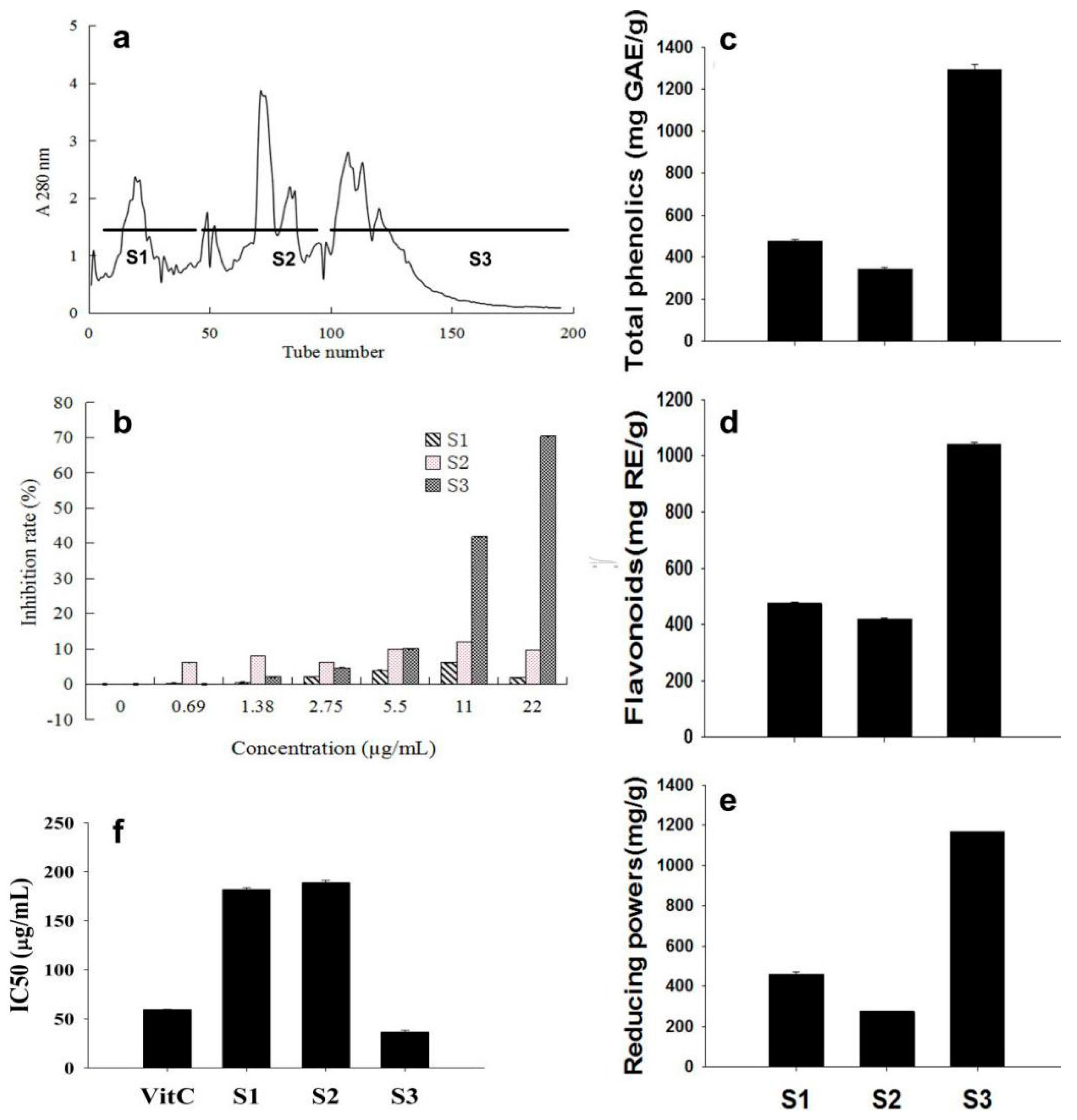
2.2. The Fraction with High Anti-Cancer Activity was Further Separated, and a Sub-Fraction S3 Was Isolated with High Activity
2.3. The Sub-Fraction with High Anti-Cancer Activity Was Further Separated and a Fraction Contained a B-Type Trimer Proanthocyanidin Showed High Antioxidant and Anticancer Activities
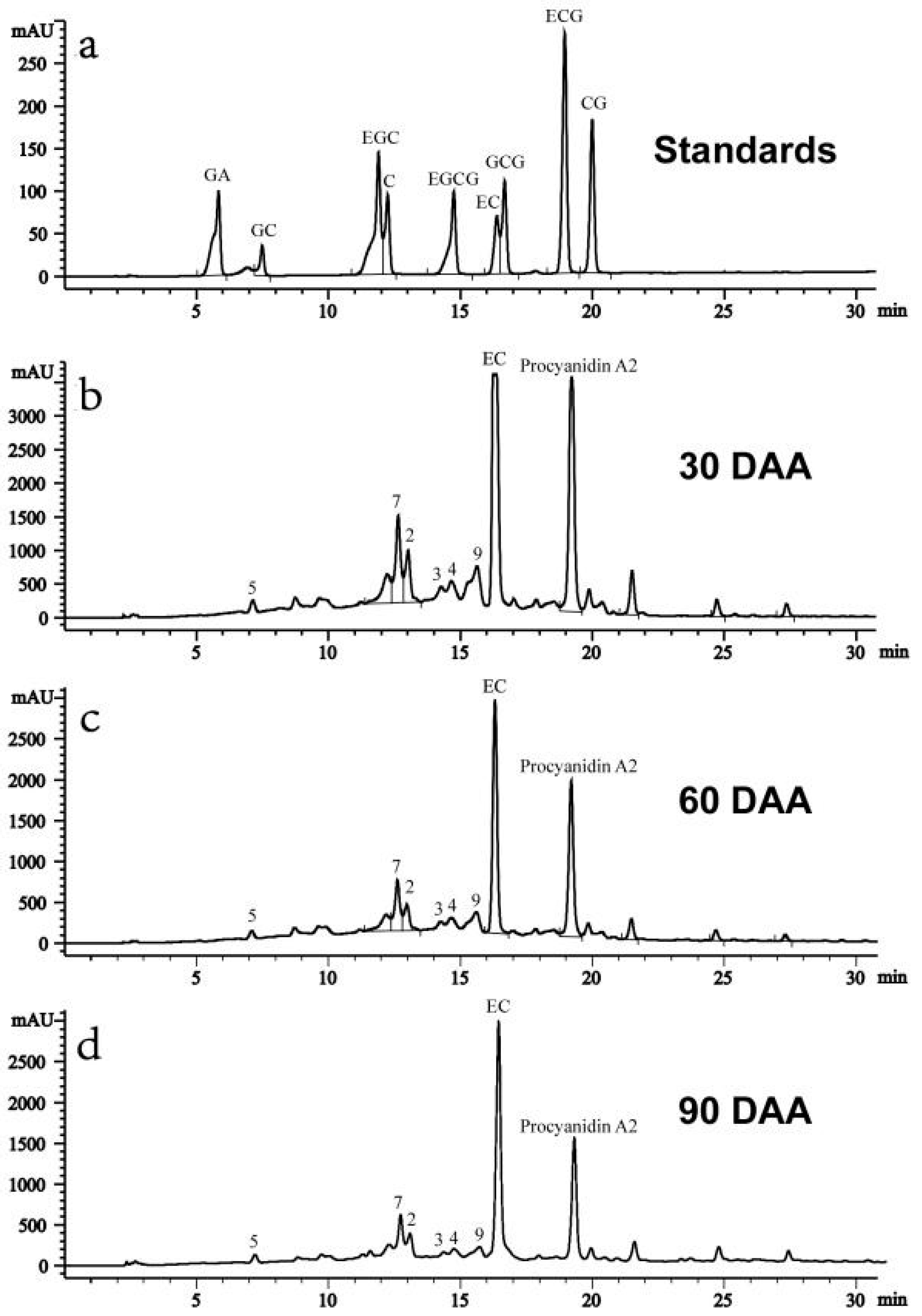
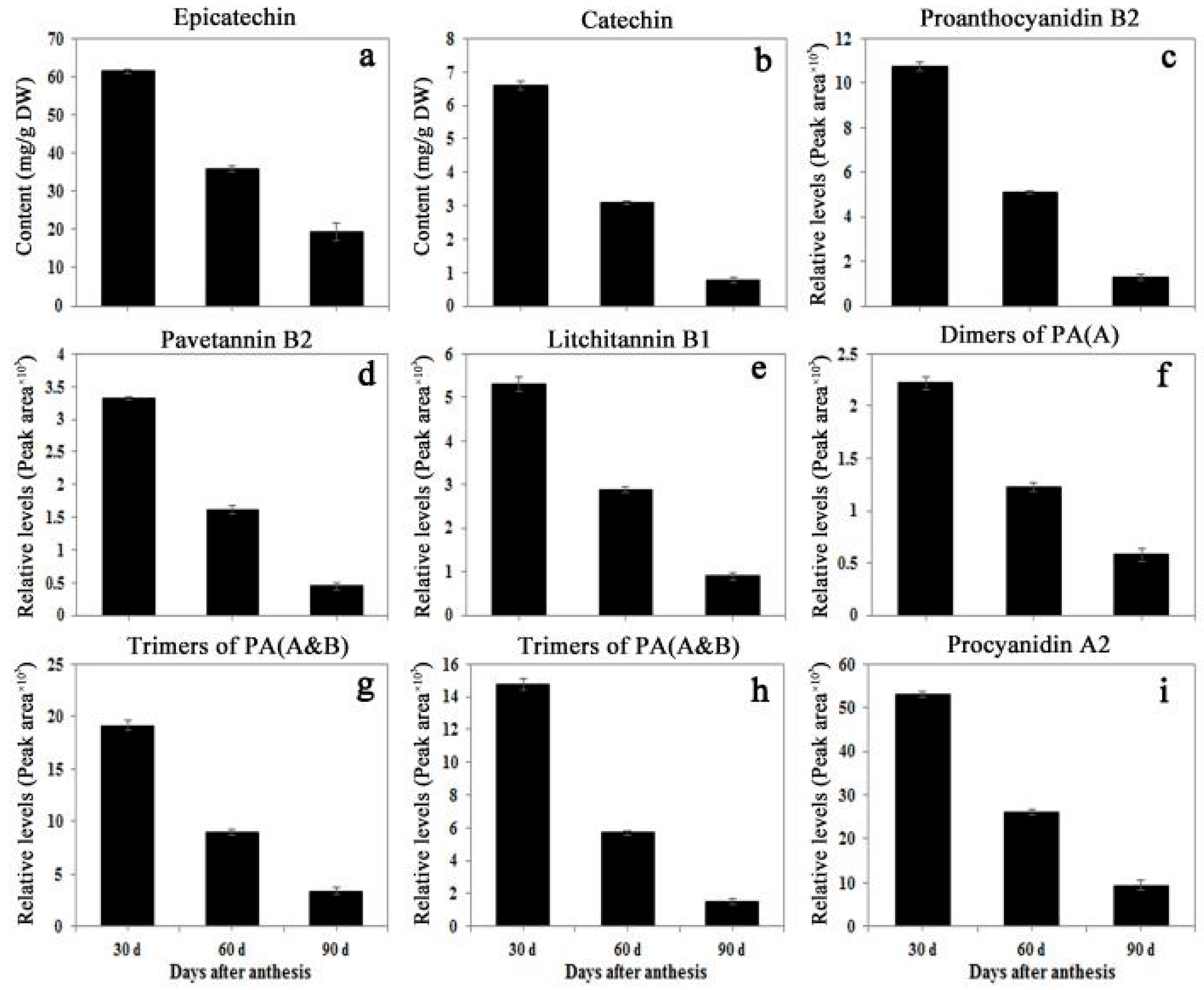
2.4. Contents of Catechins and Proanthocyanidins in Litchi pericarp during Fruit Development
2.5. Expression of Catechin and Anthocyanin Biosynthesis Related Genes in Litchi pericarp during Fruit Development
3. Materials and Methods
3.1. Plant Materials
3.2. Extraction and Purification of Litchi pericarp Phenolics
3.3. Analysis for Total Phenolic Compounds
3.4. Determination of Total Flavonoid Content
3.5. Ferric Reducing Antioxidant Power (FRAP) Assays
3.6. DPPH Radical Scavenging Activity
3.7. Inhibition Activity Assay on Cancer Cells
3.8. HPLC-ESI-MS/MS Analysis of Fractions P1–P4
3.9. Determination of the Contents of Catechin-Type Compounds and Proanthocyanidins by HPLC
3.10. Gene Expression Analysis
3.11. Statistics
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| GAE | gallic acid equivalents |
| RE | rutin equivalents |
| CCK-8 | cell counting Kit-8 |
| EC | (−)-epicatechin |
| C | (+)-catechin |
| EGCG: | (−)-epigallocatechin gallate; |
| ECG | (−)-epicatechin-3-gallate |
| CG | (−)-catechin gallate |
| FRAP | Ferric reducing antioxidant power |
| DAA | day after anthesis |
| CHI | chalcone isomerase |
| ANS | anthocyanin synthase |
| UFGT | UDP-glucose: flavonoid 3-O-glucosyltransferase |
| ANR | anthocyanidin reductase |
| LAR | leuacoanthocyanidin reductase |
| SEM | standard error of mean |
| BHT | butylated hydroxytoluene |
References
- Bhoopat, L.; Srichairatanakool, S.; Kanjanapothi, D.; Taesotikul, T.; Thananchai, H.; Bhoopat, T. Hepatoprotective effects of lychee (Litchi chinensis sonn.): A combination of antioxidant and anti-apoptotic activities. J. Ethnopharmacol. 2011, 136, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Brat, P.; Georgé, S.; Bellamy, A.; Chaffaut, L.D. Daily polyphenol intake in France from fruit and vegetables1. J. Nutr. 2006, 136, 2368–2373. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.R.M.; Mohamed, G.A. Litchi chinensis: Medicinal uses, phytochemistry, and pharmacology. J. Ethnopharmacol. 2015, 174, 492–513. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Xie, H.; Hao, J.; Jiang, Y.; Wei, X. Flavonoid glycosides from the seeds of litchi chinensis. J. Agric. Food Chem. 2011, 59, 1205–1209. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.B.; Wu, K.S.; Gu, Y.; Chen, J.Z. Research progress in the chemical constituents and pharmacological effects of lychee seeds. J. Infect. Tradit. Chin. Med. 2007, 14, 97–98. [Google Scholar]
- Chen, H.B.; Wang, H.C.; Zhuang, L.J. The litchi research and development in China in 2010. In Proceedings of the 2011 China National Workship on Litchi and Longan, Guangzhou, China, 28–30 June 2011; pp. 1–9. [Google Scholar]
- Sarni-Manchado, P.; Le Roux, E.; Le Guernevé, C.; Lozano, Y.; Cheynier, V. Phenolic composition of litchi fruit pericarp. J. Agric. Food Chem. 2000, 48, 5995–6002. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Jiang, Y.; Su, X.; Zhang, Z.; Shi, J. Antioxidant properties of anthocyanins extracted from litchi (Litchi chinenesis sonn.) fruit pericarp tissues in relation to their role in the pericarp browning. Food Chem. 2007, 101, 1365–1371. [Google Scholar] [CrossRef]
- Zhao, M.; Yang, B.; Wang, J.; Liu, Y.; Yu, L.; Jiang, Y. Immunomodulatory and anticancer activities of flavonoids extracted from litchi (Litchi chinensis sonn.) pericarp. Int. Immunopharmacol. 2007, 7, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Ogasawara, J.; Kitadate, K.; Nishioka, H.; Fujii, H.; Sakurai, T.; Kizaki, T.; Izawa, T.; Ishida, H.; Ohno, H. Oligonol, a new lychee fruit-derived low-molecular form of polyphenol, enhances lipolysis in primary rat adipocytes through activation of the ERK1/2 pathway. Phytother. Res. 2009, 23, 1626–1633. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Noh, J.S.; Fujii, H.; Roh, S.-S.; Song, Y.-O.; Choi, J.S.; Chung, H.Y.; Yokozawa, T. Oligonol, a low-molecular-weight polyphenol derived from lychee fruit, attenuates gluco-lipotoxicity-mediated renal disorder in type 2 diabetic db/db mice. Drug Discov. Ther. 2015, 9, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Kessy, H.; Hu, Z.; Zhao, L.; Zhou, M. Effect of steam blanching and drying on phenolic compounds of Litchi pericarp. Molecules 2016, 21, 729. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Sharma, B.; Kanwar, S.S.; Kumar, A. Lead phytochemicals for anticancer drug development. Front. Plant Sci. 2016, 7, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Pang, X.; Yang, C.; Ji, Z.; Jiang, Y. Purification and structural analysis of anthocyanins from Litchi pericarp. Food Chem. 2004, 84, 601–604. [Google Scholar] [CrossRef]
- Hu, Y.; Deng, L.; Chen, J.; Zhou, S.; Liu, S.; Fu, Y.; Yang, C.; Liao, Z.; Chen, M. An analytical pipeline to compare and characterise the anthocyanin antioxidant activities of purple sweet potato cultivars. Food Chem. 2016, 194, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Sommella, E.; Pepe, G.; Pagano, F.; Conte, G.; Carimi, F.; Tenore, G.C.; Novellino, E.; Manfra, M.; Russo, M.; Campiglia, P. Rapid screening of antioxidant anthocyanins in autochthonous nero d’avola grape clones by pre-column DPPH reaction coupled to UHPLC-UV/Vis-IT-Tof: A strategy to combine chemical data and genetic diversity. Food Anal. Methods 2016, 9, 2780–2790. [Google Scholar] [CrossRef]
- Faramarzi, S.; Pacifico, S.; Yadollahi, A.; Lettieri, A.; Nocera, P.; Piccolella, S. Red-fleshed apples: Old autochthonous fruits as a novel source of anthocyanin antioxidants. Plant Food Hum. Nutr. 2015, 70, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Faria, A.; Pestana, D.; Teixeira, D.; de Freitas, V.; Mateus, N.; Calhau, C. Blueberry anthocyanins and pyruvic acid adducts: Anticancer properties in breast cancer cell lines. Phytother. Res. 2010, 24, 1862–1869. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.Z.; Hu, F.C.; Hu, G.B.; Li, X.J.; Huang, X.M.; Wang, H.C. Differential expression of anthocyanin biosynthetic genes in relation to anthocyanin accumulation in the pericarp of litchi chinensis sonn. PLoS ONE 2011, 6, e19455. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Quantick, P.C.; Grigor, J.M. Changes in phenolic compounds in litchi (Litchi chinensis sonn.) fruit during postharvest storage. Postharvest Biol. Technol. 2000, 19, 165–172. [Google Scholar] [CrossRef]
- Jing, G.; Huang, H.; Yang, B.; Li, J.; Zheng, X.; Jiang, Y. Effect of pyrogallol on the physiology and biochemistry of litchi fruit during storage. Chem. Cent. J. 2013, 7, 19. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Yang, B.; Wang, J.; Li, B.; Jiang, Y. Identification of the major flavonoids from pericarp tissues of lychee fruit in relation to their antioxidant activities. Food Chem. 2006, 98, 539–544. [Google Scholar] [CrossRef]
- Xu, X.; Xie, H.; Wang, Y.; Wei, X. A-type proanthocyanidins from lychee seeds and their antioxidant and antiviral activities. J. Agric. Food Chem. 2010, 58, 11667–11672. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Wu, D.; Jiang, Y.; Prasad, K.N.; Lin, S.; Jiang, G.; He, J.; Zhao, M.; Luo, W.; Yang, B. Identification of flavonoids in litchi (Litchi chinensis sonn.) leaf and evaluation of anticancer activities. J. Funct. Foods 2014, 6, 555–563. [Google Scholar]
- Liu, L.; Xie, B.; Cao, S.; Yang, E.; Xu, X.; Guo, S. A-type procyanidins from litchi chinensis pericarp with antioxidant activity. Food Chem. 2007, 105, 1446–1451. [Google Scholar] [CrossRef]
- Takanashi, K.; Suda, M.; Matsumoto, K.; Ishihara, C.; Toda, K.; Kawaguchi, K.; Senga, S.; Kobayashi, N.; Ichikawa, M.; Katoh, M.; et al. Epicatechin oligomers longer than trimers have anti-cancer activities, but not the catechin counterparts. Sci. Rep. 2017, 7, 7791. [Google Scholar]
- Chen, Q.; Zhang, X.N.; Yu, H.W.; Yan, W.; Tang, H.R. Changes of total anthocyanins and proanthocyanidins in the developing blackberry fruits. J. Int. ChemTech Res. 2012, 4, 129–137. [Google Scholar]
- Carbone, F.; Preuss, A.; DeVos, R.C.H.; D’Amico, E.; Perrotta, G.; Bovy, A.G.; Martens, S.; Rosati, C. Developmental, genetic and environmental factors affect the expression of flavonoid genes, enzymes and metabolites in strawberry fruits *. Plant Cell Environ. 2009, 32, 1117–1131. [Google Scholar] [CrossRef] [PubMed]
- Bogs, J.; Jaffé, F.W.; Takos, A.M.; Walker, A.R.; Robinson, S.P. The grapevine transcription factor vvmybpa1 regulates proanthocyanidin synthesis during fruit development. Plant Physiol. 2007, 143, 1347–1361. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, J.; Xia, R. Expression of chalcone synthase and chalcone isomerase genes and accumulation of corresponding flavonoids during fruit maturation of guoqing no. 4 satsuma mandarin (Citrus unshiu Marcow). Sci. Horticult. 2010, 125, 110–116. [Google Scholar]
- Guan, C.; Song, X.; Ji, J.; Li, X.; Jin, C.; Guan, W.; Li, J.; Wang, G. Salicylic acid treatment enhances expression of chalcone isomerase gene and accumulation of corresponding flavonoids during fruit maturation of lycium chinense. Eur. Food Res. Technol. 2014, 239, 857–865. [Google Scholar] [CrossRef]
- Wang, H.; Wang, W.; Li, H.; Zhang, P.; Zhan, J.; Huang, W. Expression and tissue and subcellular localization of anthocyanidin synthase (ANS) in grapevine. Protoplasma 2011, 248, 267–279. [Google Scholar] [CrossRef] [PubMed]
- Szankowski, I.; Flachowsky, H.; Li, H.; Halbwirth, H.; Treutter, D.; Regos, I.; Hanke, M.-V.; Stich, K.; Fischer, T.C. Shift in polyphenol profile and sublethal phenotype caused by silencing of anthocyanidin synthase in apple (Malus sp.). Planta 2009, 229, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Han, Z.Y.; Tian, J.; Zhang, X.; Song, T.T.; Yao, Y.C. The expression level of anthocyanidin synthase determines the anthocyanin content of crabapple (Malus sp.) petals. Acta Physiol. Plant. 2015, 37, 109. [Google Scholar]
- Yoshihara, N.; Imayama, T.; Fukuchi-Mizutani, M.; Okuhara, H.; Tanaka, Y.; Ino, I.; Yabuya, T. cDNA cloning and characterization of UDP-glucose: Anthocyanidin 3-O-glucosyltransferase in iris hollandica. Plant Sci. 2005, 169, 496–501. [Google Scholar] [CrossRef]
- Lai, B.; Li, X.J.; Hu, B.; Qin, Y.H.; Huang, X.M.; Wang, H.C.; Hu, G.B. LcMYB1 is a key determinant of differential anthocyanin accumulation among genotypes, tissues, developmental phases and aba and light stimuli in litchi chinensis. PLoS ONE 2014, 9, e86293. [Google Scholar] [CrossRef] [PubMed]
- Tanner, G.J.; Francki, K.T.; Abrahams, S.; Watson, J.M.; Philip, J.; Larkin, P.J.; Ashton, A.R. Proanthocyanidin biosynthesis in plants. Purification of legume leucoanthocyanidin reductase and molecular cloning of its cDNA. J. Biol. Chem. 2003, 278, 31647–31656. [Google Scholar] [CrossRef] [PubMed]
- Xie, D.Y.; Sharma, S.B.; Paiva, N.L.; Ferreira, D.; Dixon, R.A. Role of anthocyanidin reductase, encoded by BANYULS in plant flavonoid biosynthesis. Science 2003, 299, 396–399. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, J.; Kühnel, C.; Brandt, J.; Duy, D.; Punyasiri, P.A.N.; Forkmann, G.; Fischer, T.C. Biosynthesis of flavan 3-ols by leucoanthocyanidin 4-reductases and anthocyanidin reductases in leaves of grape (Vitis vinifera L.), apple (Malus x domestica Borkh.) and other crops. Plant Physiol. Biochem. 2006, 44, 323–334. [Google Scholar] [PubMed]
- Zhang, X.N.; Chen, Q.; Yu, D.Q.; Zhou, S.L.; Tang, H.R. Expression analysis of the ANR and LAR gene in Fragaria × ananassa cv. Toyonaka. J. Agric. Sci. 2013, 5, 1916–9752. [Google Scholar] [CrossRef]
- Gagné, S.; Lacampagne, S.; Claisse, O.; Gény, L. Leucoanthocyanidin reductase and anthocyanidin reductase gene expression and activity in flowers, young berries and skins of Vitis vinifera L. Cv. Cabernet-sauvignon during development. Plant Physiol. Biochem. 2009, 47, 282–290. [Google Scholar]
- Dewanto, V.; Wu, X.; Adom, K.K.; Liu, R.H. Thermal processing enhances the nutritional value of tomatoes by increasing total antioxidant activity. J. Agric. Food Chem. 2002, 50, 3010–3014. [Google Scholar] [CrossRef] [PubMed]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The frap assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Zheng, W.J.; Feng, Q.; Liu, J.; Guo, Y.K.; Gao, L.F.; Li, R.M.; Xu, M.; Yan, G.Z.; Yin, Z.N.; Zhang, S.; et al. Inhibition of 6-phosphogluconate dehydrogenase reverses cisplatin resistance in ovarian and lung cancer. Front. Pharmacol. 2017, 8, 421. [Google Scholar] [CrossRef] [PubMed]
- Rzeppa, S.; Von Bargen, C.; Bittner, K.; Humpf, H.U. Analysis of flavan-3-ols and procyanidins in food samples by reversed phase high-performance liquid chromatography coupled to electrospray ionization tandem mass spectrometry (RP-HPLC-ESI-MS/MS). J. Agric. Food Chem. 2011, 59, 10594–10603. [Google Scholar] [CrossRef] [PubMed]
- Ponchel, F.; Toomes, C.; Bransfield, K.; Leong, F.T.; Douglas, S.H.; Field, S.L.; Bell, S.M.; Combaret, V.; Puisieux, A.; Mighell, A.J.; et al. Real-time PCR based on SYBR-green I fluorescence: An alternative to the taqman assay for a relative quantification of gene rearrangements, gene amplifications and micro gene deletions. BMC Biotechnol. 2003, 3, 18. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | Compound Number | Mode | Mass | m/z | * Molecular Formula | Auto MS-MS Fragment Ions | * Tentative Identification |
|---|---|---|---|---|---|---|---|
| P1 | 1 | pos | 290.0792 | 291.0869 | C15H14O6 | 139.0388; 123.044 | EC (Epicatechin) [20,21] |
| neg | 290.1537 | 289.1464 | 245.0816; 125.0243 | ||||
| P2 | 2 | pos | 578.1425 | 579.1498 | C30H26O12 | 127.0391; 291.0866; 409.0920 | PA (Proanthocyanidin) B2 [22] |
| neg | 578.1418 | 577.1345 | 125.0244; 289.0713; 407.0778 | ||||
| P3 | 3 | pos | 864.1899 | 887.1792 | C45H36O18 | 289.0714; 425.0902; 577.1345; 713.1506 | Pavetannin B2 (Trimers of complex Proanthocyanidin A and B) [23] |
| neg | 864.1899 | 863.1867 | 125.0244; 289.0717; 423.0712; 575.1186; 711.1357 | ||||
| 4 | pos | 866.2053 | 889.1945 | C45H38O18 | 289.0706; 601.1310; 717.1510 | Litchitannin B1 (Trimers of Proanthocyanidin B) | |
| neg | - | - | - | ||||
| P4 | 5 | pos | - | - | C30H24O12 | - | 2 × PA(A) (Dimers of Proanthocyanidin A) [24] |
| neg | 576.1257 | 575.1184 | - | ||||
| 6 | pos | 866.2056 | 867.2129 | C45H38O18 | 247.0601; 289.0705; 427.1025; 579.1492; 715.1647 | Litchitannin B2 (Trimers of Proanthocyanidin B) | |
| neg | 866.2053 | 865.1981 | 125.0243; 287.0556; 407.0755; 575.1187; 713.1531 | ||||
| 7 | pos | 864.1902 | 865.1974 | C45H36O18 | 287.0550; 411.1073; 533.1076; 575.1179; 713.1497 | 3 × PA(A&B) (Trimers of complex Proanthocyanidin A and B) [23] | |
| neg | 864.1899 | 863.1826 | 125.0247; 289.0716; 411.0720; 573.1025; 711.1346 | ||||
| 8 | pos | 848.1951 | 849.2023 | C45H36O17 | 287.0557; 517.1125; 713.1504 | Trimeric PA (with an afzelechin or epiafzelechin) [23] | |
| neg | 848.1948 | 847.1875 | 289.0710; 411.0723; 557.1095 | ||||
| 9 | pos | 864.1905 | 865.1979 | C45H36O18 | 287.0551; 425.0869; 577.1336; 713.1497 | 3 × PA (A&B) [23] | |
| neg | 864.1899 | 863.1827 | 125.0243; 285.0404; 575.1201; 711.1362 | ||||
| 10 | pos | 576.127 | 577.1342 | C30H24O12 | 123.0440; 287.0549; 425.0864 | Procyanidin A2 [25] |
| Genes | GenBank ID | Forward Primer (5′ to 3′) | Reverse Primer (5′ to 3′) |
|---|---|---|---|
| LcCHI | HQ402910 | CGGAGTTTACTTGGAGGATGT | CAGTGACCTTCTCAGAGTATTG |
| LcANS | HQ402913 | AGGAAGTTGGTGGTCTGGAAG | CCGTTGCTGAGGATTTCAATGGTG |
| LcUFGT | HQ402914 | GCCACCAGCGGTTCCTAATA | ATGCCTCTGCTACTGCTACAATCT |
| LcLAR | Litchi_GLEAN_10054331 | AGATTGGACGAACTCTACC | TTGAAACAATCATCCAACG |
| LcANR | Litchi_GLEAN_10049142 | ATACATTTGTGCTGCTTTC | TCATAGGTTTCTTCAAGCC |
| LcActin | HQ615689 | ACCGTATGAGCAAGGAAATCACTG | TCGTCGTACTCACCCTTTGAAATC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, Y.; Fang, F.; Zhang, X.; Liu, B.; Luo, H.; Li, Z.; Zhang, X.; Zhang, Z.; Pang, X. B Type and Complex A/B Type Epicatechin Trimers Isolated from Litchi pericarp Aqueous Extract Show High Antioxidant and Anticancer Activity. Int. J. Mol. Sci. 2018, 19, 301. https://doi.org/10.3390/ijms19010301
Gong Y, Fang F, Zhang X, Liu B, Luo H, Li Z, Zhang X, Zhang Z, Pang X. B Type and Complex A/B Type Epicatechin Trimers Isolated from Litchi pericarp Aqueous Extract Show High Antioxidant and Anticancer Activity. International Journal of Molecular Sciences. 2018; 19(1):301. https://doi.org/10.3390/ijms19010301
Chicago/Turabian StyleGong, Yihui, Fang Fang, Xin Zhang, Bin Liu, Honghui Luo, Zhen Li, Xuelian Zhang, Zhaoqi Zhang, and Xuequn Pang. 2018. "B Type and Complex A/B Type Epicatechin Trimers Isolated from Litchi pericarp Aqueous Extract Show High Antioxidant and Anticancer Activity" International Journal of Molecular Sciences 19, no. 1: 301. https://doi.org/10.3390/ijms19010301
APA StyleGong, Y., Fang, F., Zhang, X., Liu, B., Luo, H., Li, Z., Zhang, X., Zhang, Z., & Pang, X. (2018). B Type and Complex A/B Type Epicatechin Trimers Isolated from Litchi pericarp Aqueous Extract Show High Antioxidant and Anticancer Activity. International Journal of Molecular Sciences, 19(1), 301. https://doi.org/10.3390/ijms19010301