Functional Analysis of Promoters from Three Subtypes of the PI3K Family and Their Roles in the Regulation of Lipid Metabolism by Insulin in Yellow Catfish Pelteobagrus fulvidraco



Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
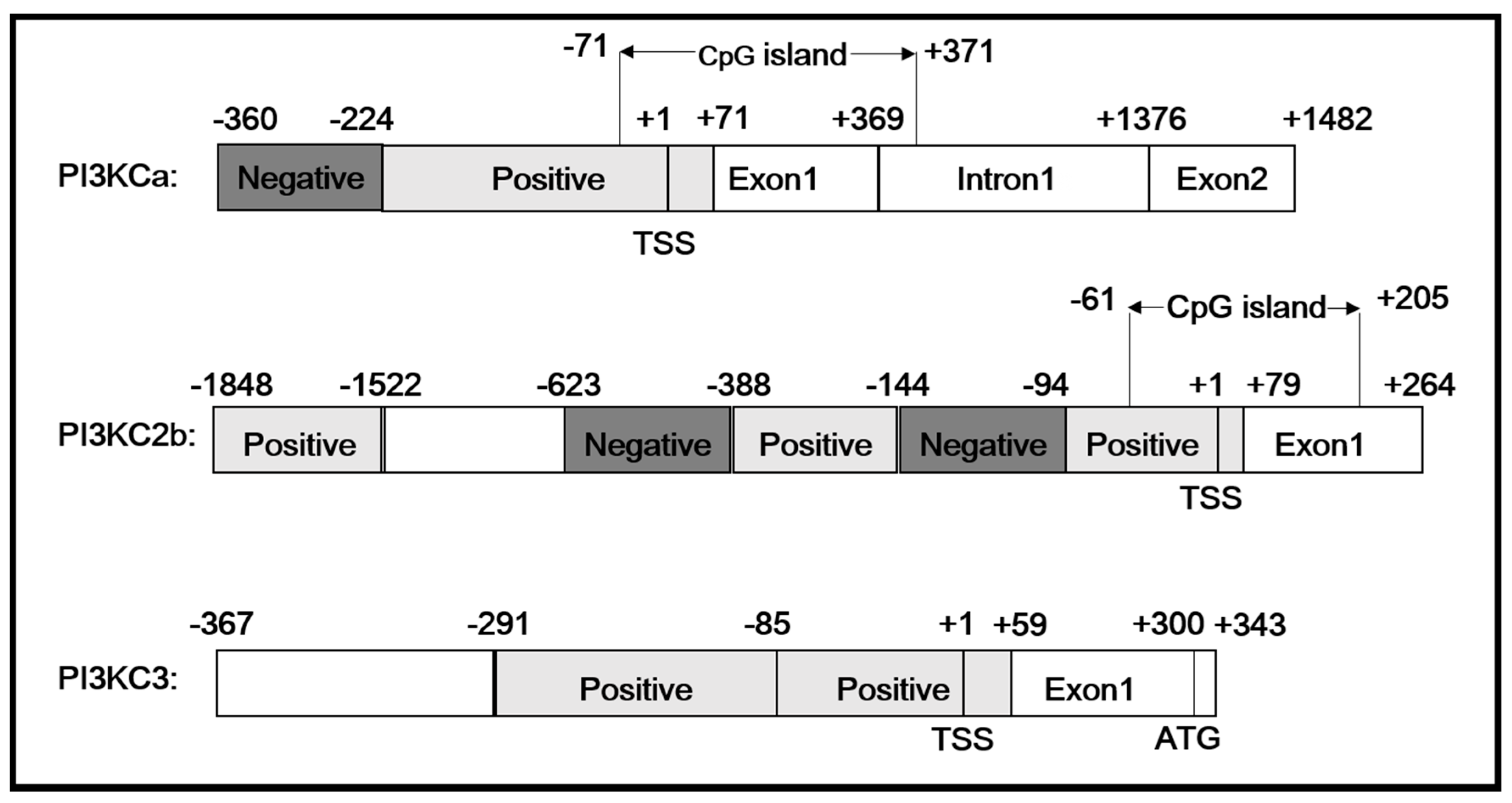
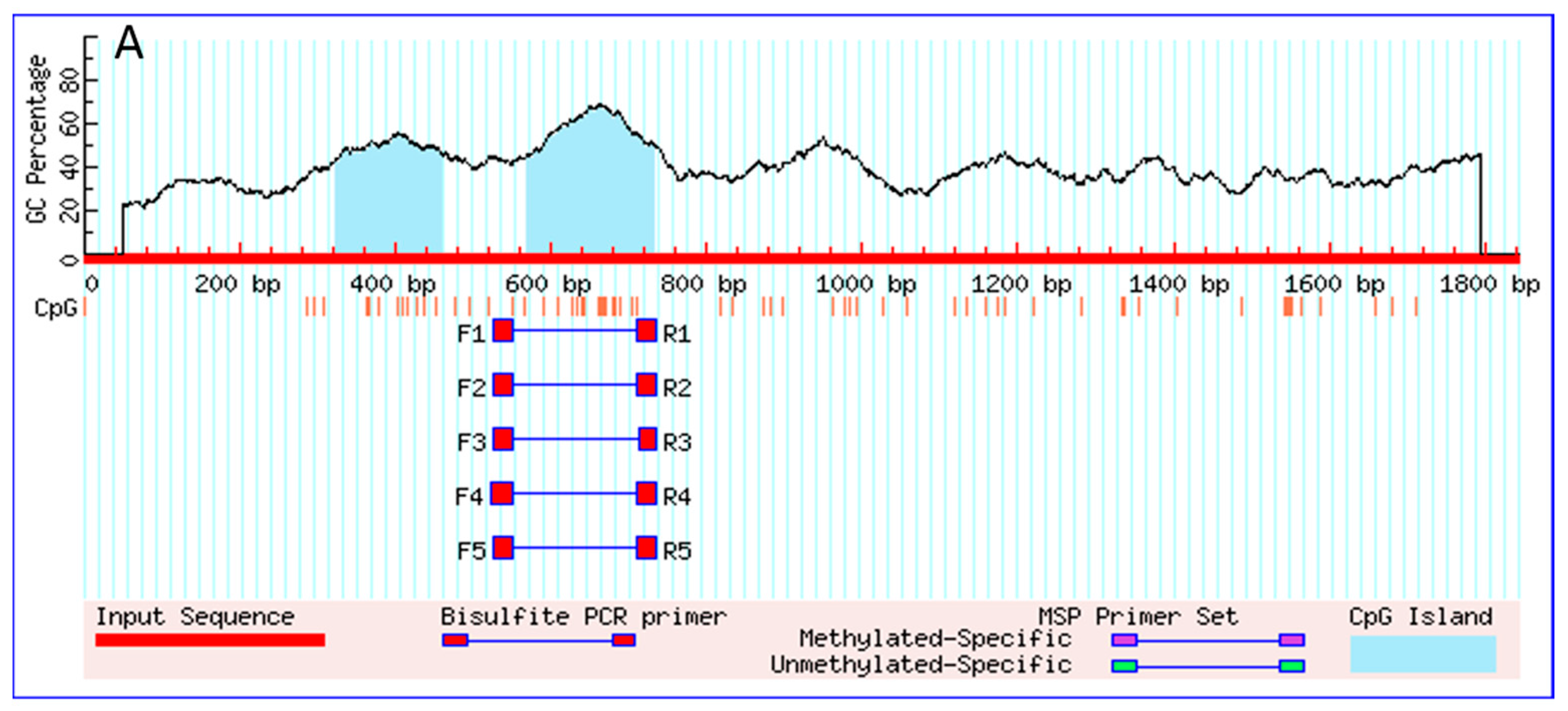
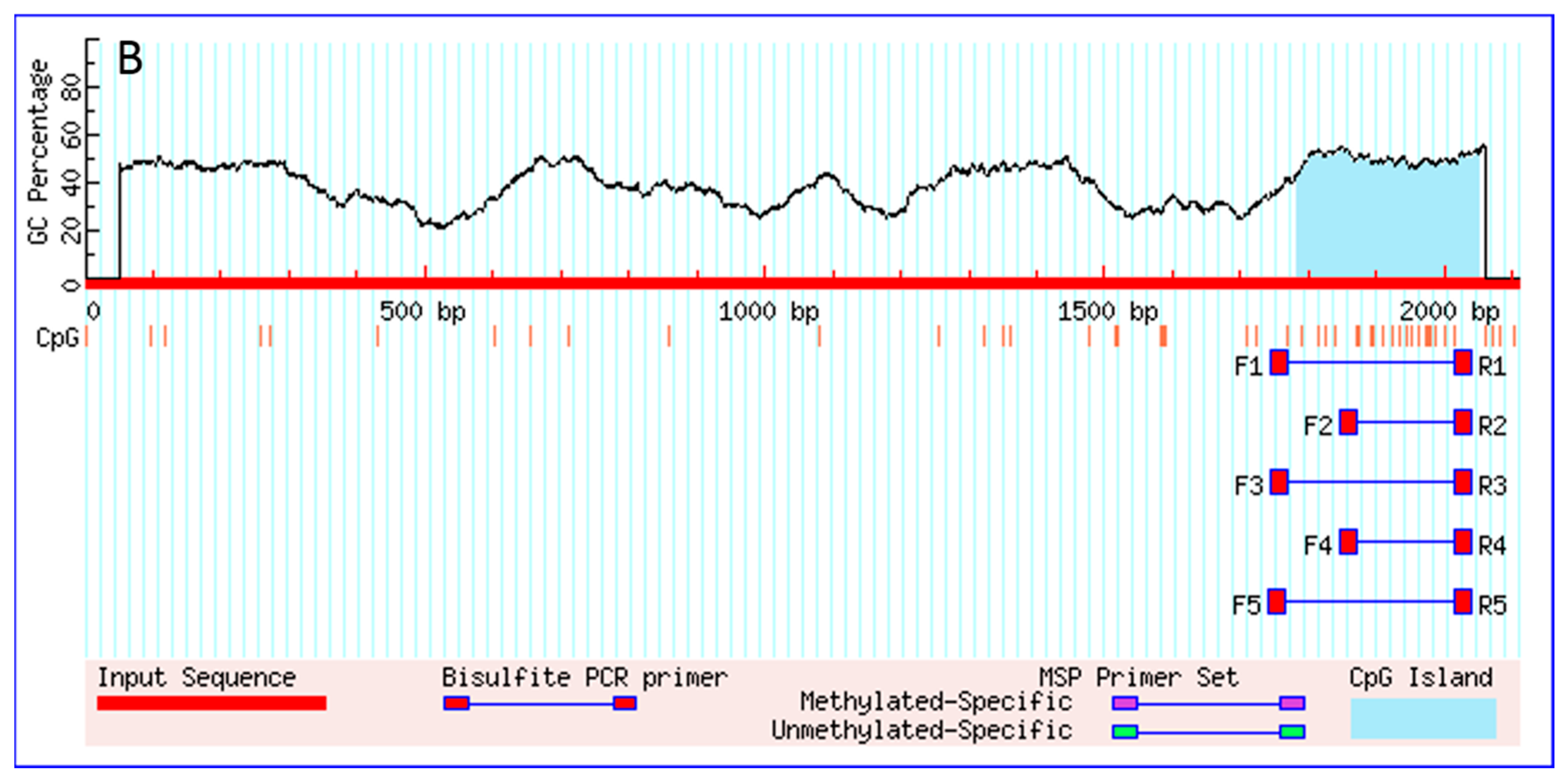
2.1. Cloning and Sequences Analysis of Promoters of PI3KCa, PI3KC2b and PI3KC3
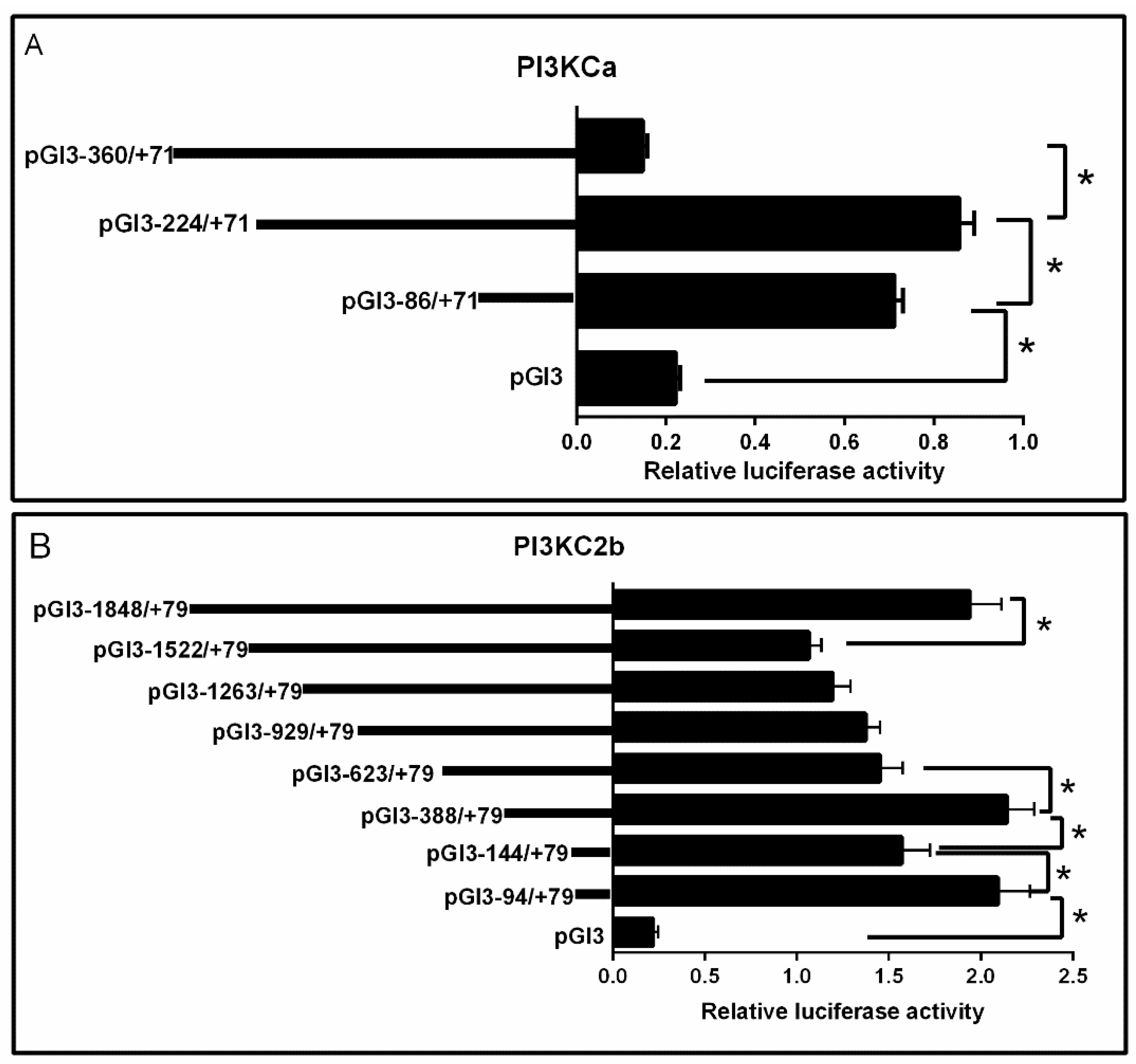
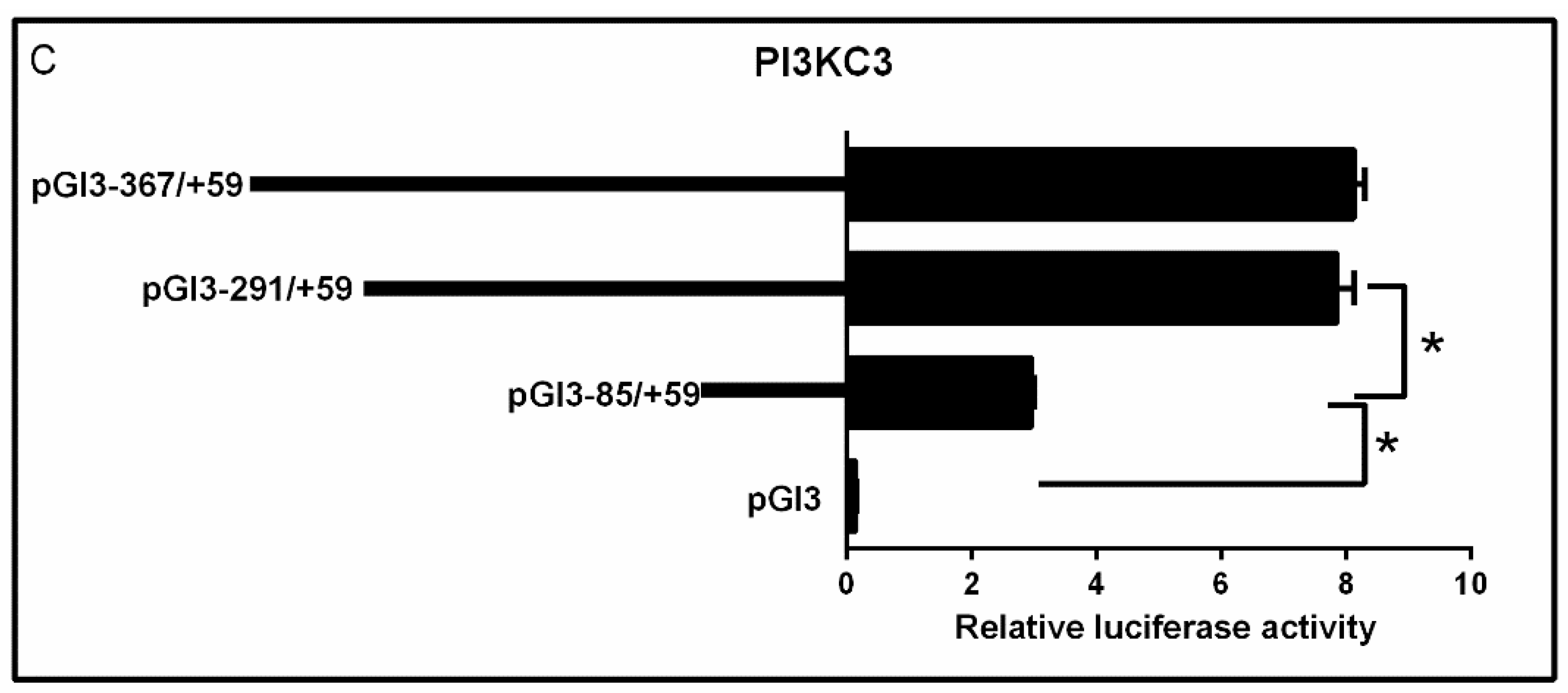
2.2. 5′-Deletion Assay of the PI3KCa, PI3KC2b and PI3KC3 Promoter
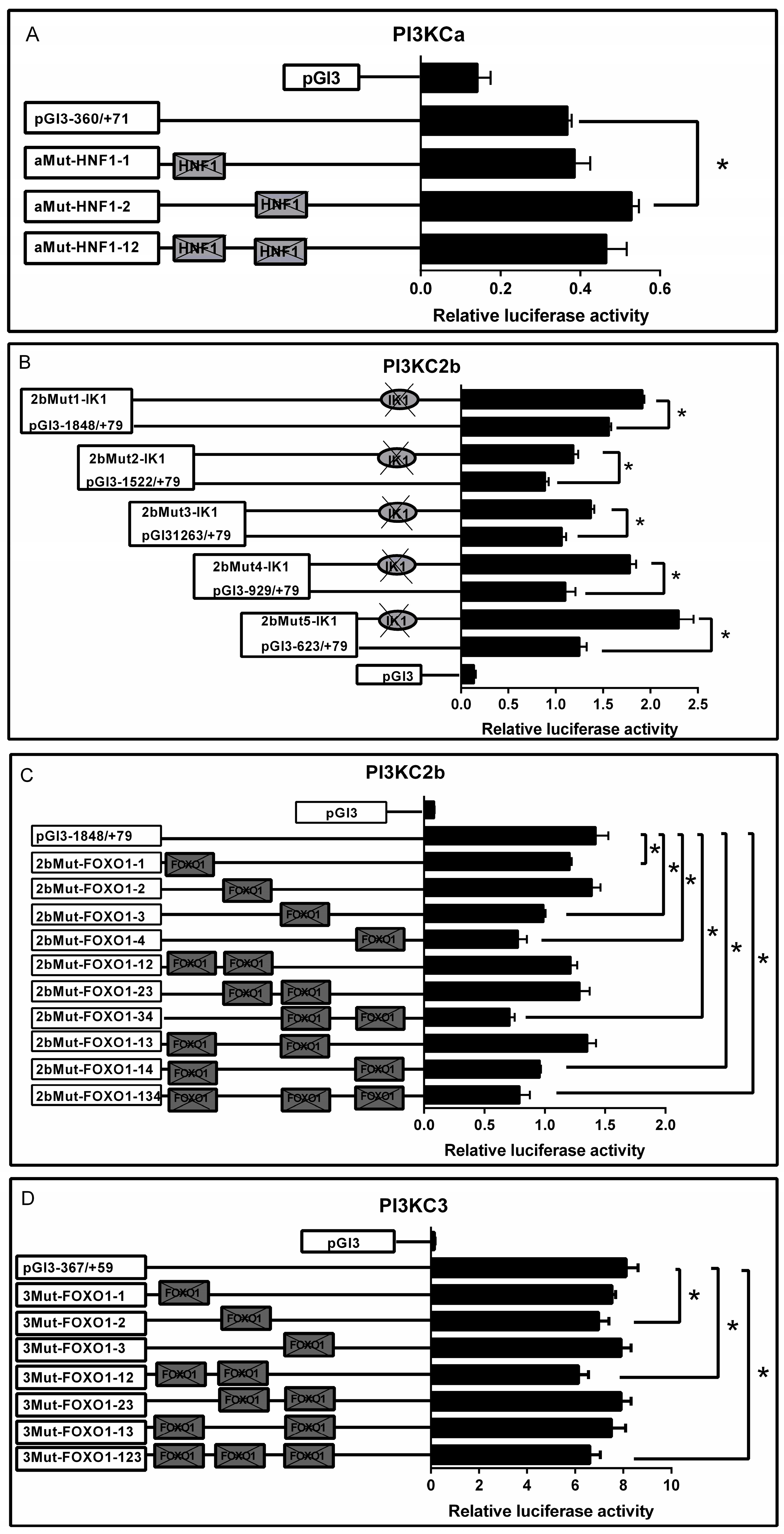
2.3. Site-Mutation Analysis
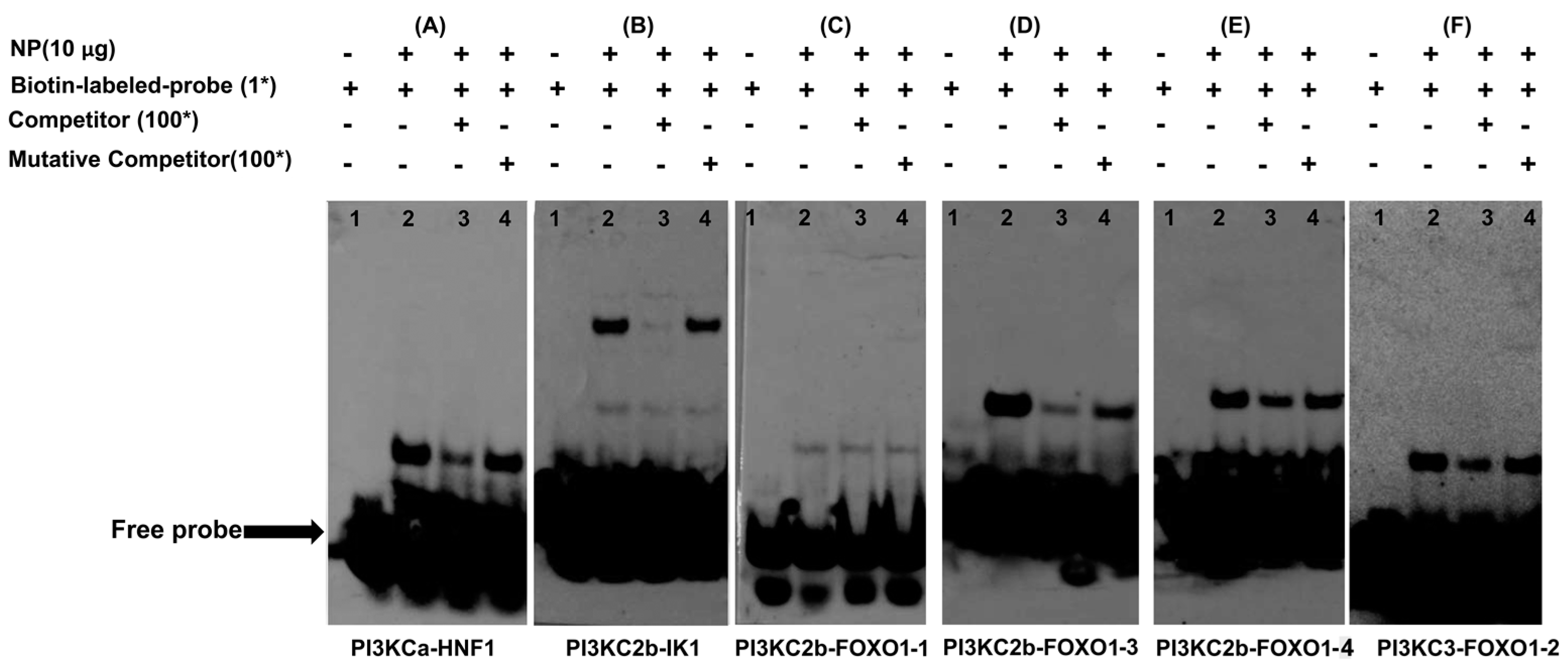
2.4. HNF1, FOXO1 and IK1 Bind with the Promoters
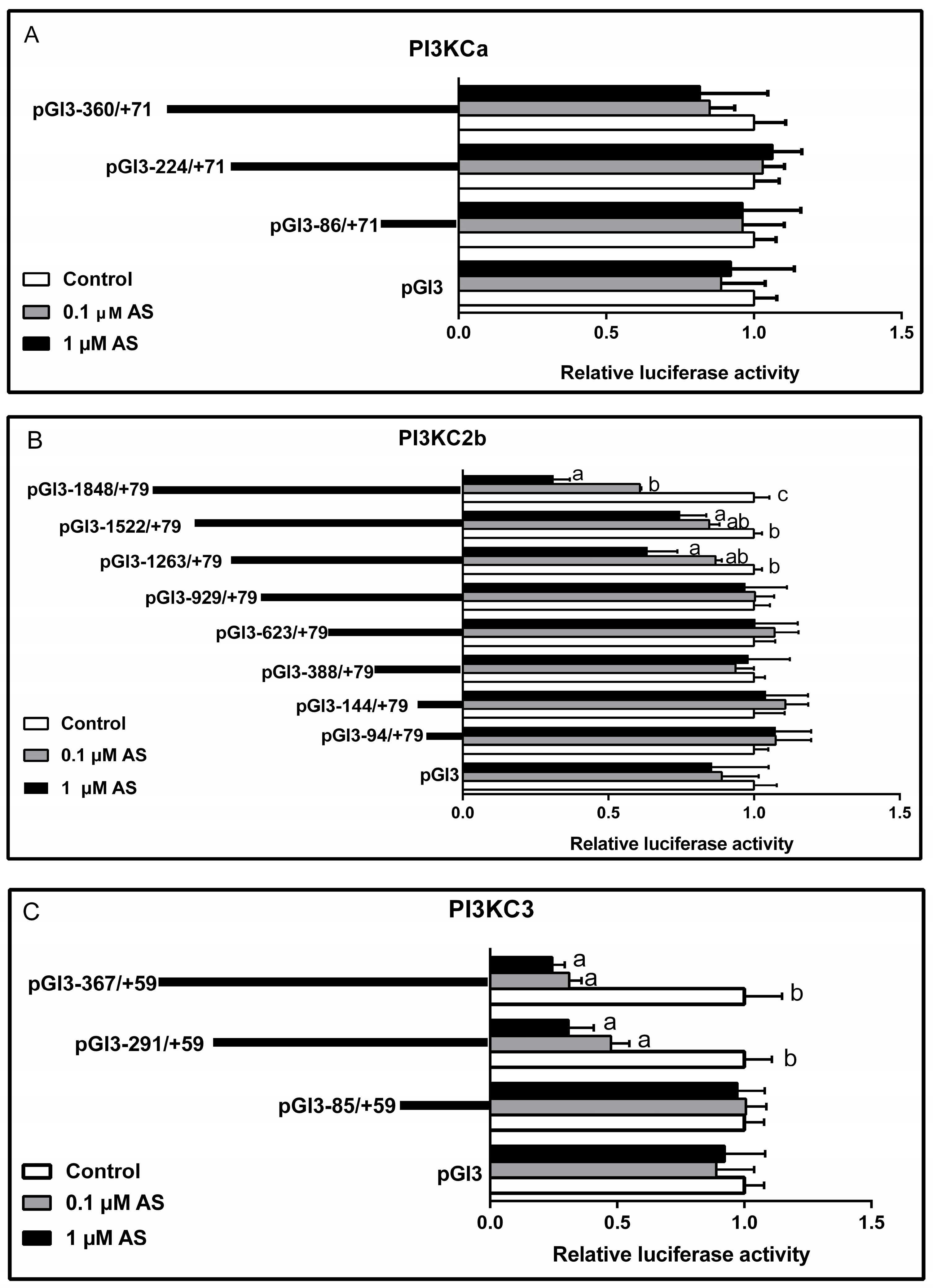
2.5. Effect of AS on Plasmid Luciferase Activity
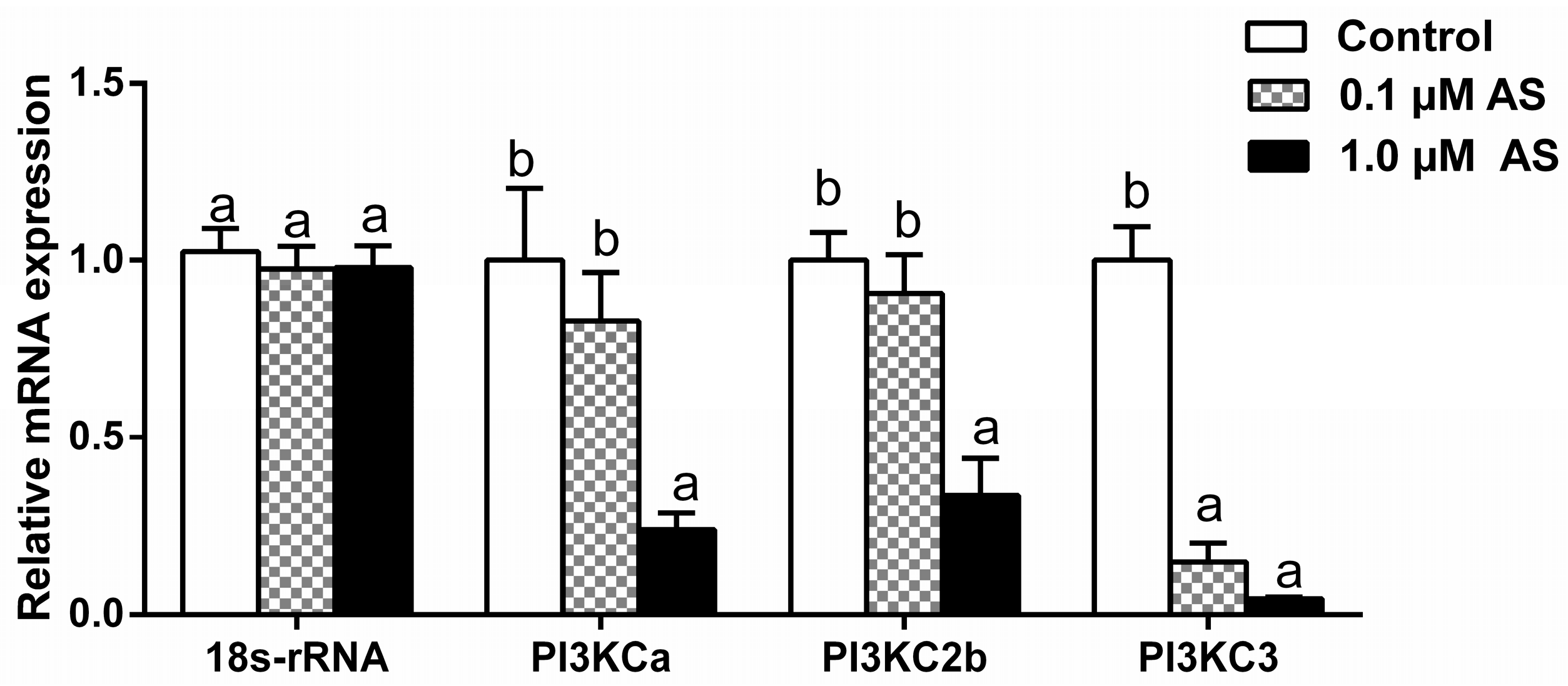
2.6. Effect of AS on PI3Ks Gene Expressions
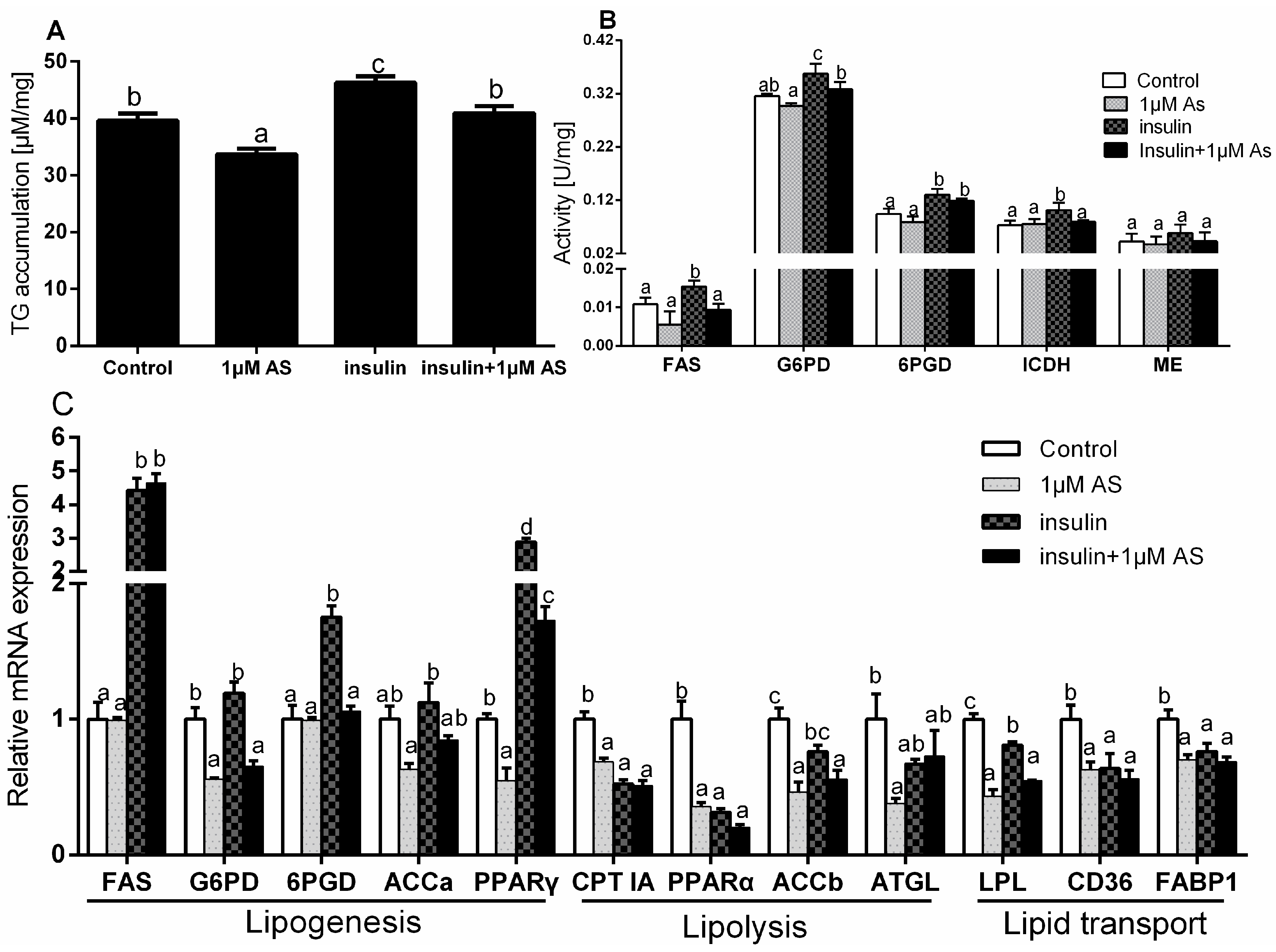
2.7. Effect of Insulin and/or AS on TG Accumulation, Enzymatic Activity and Gene Expressions
3. Discussion
4. Materials and Methods
4.1. Experimental Animals and Reagents
4.2. Experimental Treatment
4.2.1. Exp. 1: Cloning and Functional Analysis of PI3KCa, PI3KC2b and PI3KC3 Promoters
Cloning of Promoters and 5′-Deletion Plasmids Construction
Transfections and Luciferase Assays
Sequence Analysis
Site-Mutation Analysis of Several Binding Sites on the Promoters of PI3KCa, PI3KC2b and PI3KC3
Electrophoretic Mobility-Shift Assay (EMSA)
4.2.2. Exp2. Insulin and/or AS Incubation with Hepatocytes of Yellow Catfish
TG Accumulation and Enzyme Activity Analysis
mRNA Level Determination by Quantitative Real-Time PCR
4.2.3. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| 6PGD | 6-phosphogluconate dehydrogenase |
| ACC | acetyl-CoA carboxylase |
| Akt | protein kinase B |
| AP1 | activator protein-1 |
| AS | AS1842856 |
| ATGL | adipose triacylglyceride lipase |
| BRE | B recognition element |
| CD36 | fatty acid translocase |
| CPT IA | carnitine palmitoyltransferase IA |
| DCE | downstream core element |
| DRE | DNA recognition element |
| eEF | translation elongation factor |
| FAS | fatty acid synthase |
| FABP1 | fatty acid binding protein 1 |
| FBP | fatty acid binding protein |
| FOXO1 | forkhead box O |
| G6PD | glucose 6-phosphate dehydrogenase |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase |
| HNF1 | hepatocyte nuclear factor 1 |
| HPRT | hypoxanthineguanine phosphoribosyltransferase |
| ICDH | isocitrate dehydrogenase |
| IK1 | ikaros zinc finger 1 |
| LPL | lipoprteinlipase |
| ME | malic enzyme |
| MS-222 | tricaine methanesulfonate |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazoliumbromide |
| NF-κB | nuclear factor-kappa B |
| PBS | phosphate-buffered saline |
| PI3K | Phosphatidylinositol-3 kinase |
| PPAR | peroxisome proliferators-activated receptors |
| RPL7 | ribosomal protein L7 |
| STAT | Signal transducers and activators of transcription protein |
| TFBS | transcription factor binding site |
| TG | triacylglyceride |
| TUBA | tubulin α chain |
| UBCE | ubiquitin-conjugating enzyme |
References
- Liu, L.; Song, X.; He, D.; Komma, C.; Kita, A.; Virbasius, J.V.; Zhou, G.W. Crystal structure of the C2 domain of class II phosphatidylinositide 3-kinase C2α. J. Biol. Chem. 2006, 281, 4254–4260. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.E.; Jackson, S.P. Class I phosphoinositide 3-kinases. Int. J. Biochem. Cell Biol. 2003, 35, 1028–1033. [Google Scholar] [CrossRef]
- Zhuo, M.Q.; Pan, Y.X.; Wu, K.; Xu, Y.H.; Luo, Z. Characterization and mechanism of phosphoinositide 3-kinases (PI3Ks) members in insulin-induced changes of protein metabolism in yellow catfish Pelteobagrus fulvidraco. Gen. Comp. Endocrinol. 2017, 247, 34–45. [Google Scholar] [CrossRef] [PubMed]
- O’Malley, B.W.; Towle, H.C.; Schwartz, R.J. Regulation of gene expression in eucaryotes. Annu. Rev. Genet. 1977, 11, 239–275. [Google Scholar] [CrossRef] [PubMed]
- Lenhard, B.; Sandelin, A.; Carninci, P. Metazoan promoters: Emerging characteristics and insights into transcriptional regulation. Nat. Rev. Genet. 2012, 13, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Kok, K.; Geering, B.; Vanhaesebroeck, B. Regulation of phosphoinositide 3-kinase expression in health and disease. Trends Biochem. Sci. 2009, 34, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Hui, R.C.Y.; Gomes, A.R.; Constantinidou, D.; Costa, J.R.; Karadedou, C.T.; DeMattos, S.F.; Lam, E.W.F. The forkhead transcription factor FOXO3a increases PI3K/Akt activity in drug-resistant leukaemic cells through induction of PIK3CA expression. Mol. Cell. Biol. 2008, 28, 5886–5898. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Huang, J.; Greshock, J.; Liang, S.; Barchetti, A.; Hasegawa, K.; Katsaros, D. Transcriptional regulation of PIK3CA oncogene by NF-κB in ovarian cancer microenvironment. PLoS ONE 2008, 3, e1758. [Google Scholar] [CrossRef] [PubMed]
- Astanehe, A.; Arenillas, D.; Wasserman, W.W.; Leung, P.C.; Dunn, S.E.; Davies, B.R.; Auersperg, N. Mechanisms underlying p53 regulation of PIK3Ca transcription in ovarian surface epithelium and in ovarian cancer. J. Cell Sci. 2008, 121, 664–674. [Google Scholar] [CrossRef] [PubMed]
- Gaikwad, S.M.; Gunjal, L.; Junutula, A.R.; Astanehe, A.; Gambhir, S.S.; Ray, P. Non-invasive imaging of phosphoinositide-3-kinase-catalytic-subunit-alpha (PIK3CA) promoter modulation in small animal models. PLoS ONE 2013, 8, e55971. [Google Scholar] [CrossRef] [PubMed]
- Gowda, C.S.; Song, C.; Ding, Y.; Muthusami, S.; Payne, K.; Ge, Z.; Dovat, S. Targeting casein kinase II exerts a therapeutic effect in pediatric high risk leukemia via inhibition of cell cycle progression and the PI3K pathway. Blood 2015, 126, 4800. [Google Scholar]
- Wu, K.; Tan, X.Y.; Wei, C.C.; You, W.J.; Zhuo, M.Q.; Song, Y.F. Isolation and expression analysis of STAT members from Synechogobius hasta and their roles in leptin affecting lipid metabolism. Int. J. Mol. Sci. 2016, 17, 406. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Tan, X.Y.; Xu, Y.H.; Chen, Q.L.; Pan, Y.X. JAK and STAT members of yellow catfish Pelteobagrus fulvidraco and their roles in leptin affecting lipid metabolism. Gen. Comp. Endocrinol. 2016, 226, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Saltiel, A.R.; Kahn, C.R. Insulin signaling and the regulation of glucose and lipid metabolism. Nature 2001, 414, 799–806. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, M.Q.; Luo, Z.; Pan, Y.X.; Wu, K.; Fan, Y.F.; Zhang, L.H.; Song, Y.F. Effects of insulin and its related signaling pathways on lipid metabolism in the yellow catfish Pelteobagrus fulvidraco. J. Exp. Biol. 2015, 218, 3083–3090. [Google Scholar] [CrossRef] [PubMed]
- Barthel, A.; Schmoll, D.; Unterman, T.G. FoxO proteins in insulin action and metabolism. Trends Endocrinol. Metab. 2005, 16, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Webb, A.E.; Brunet, A. FOXO transcription factors: Key regulators of cellular quality control. Trends Biochem. Sci. 2014, 39, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Dong, H.H. FoxO integration of insulin signaling with glucose and lipid metabolism. J. Endocrinol. 2017, 233, R67–R79. [Google Scholar] [CrossRef] [PubMed]
- Nagashima, T.; Shigematsu, N.; Maruki, R.; Urano, Y.; Tanaka, H.; Shimaya, A.; Shimokawa, T.; Shibasaki, M. Discovery of novel forkhead box O1 inhibitors for treating type 2 diabetes: Improvement of fasting glycemia in diabetic db/db mice. Mol. Pharmacol. 2010, 78, 961–970. [Google Scholar] [CrossRef] [PubMed]
- Elliott, W.H.; Elliott, D.C. Biochemistry and Molecular Biology, 4th ed.; Academic Press: Oxford, UK, 2009. [Google Scholar]
- Yahagi, N.; Shimano, H.; Hasty, A.H.; Amemiya-Kudo, M.; Okazaki, H.; Tamura, Y.; Harada, K. A crucial role of sterol regulatory element-binding protein-1 in the regulation of lipogenic gene expression by polyunsaturated fatty acids. J. Biol. Chem. 1999, 274, 35840–35844. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Olson, P.; Evans, R.M. Minireview: Lipid metabolism, metabolic diseases and peroxisome proliferator-activated receptors. Endocrinology 2003, 144, 2201–2207. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.L.; Zhuo, M.Q.; Luo, Z.; Pan, Y.X.; Song, Y.F.; Huang, C.; Zhu, Q.L.; Hu, W.; Chen, Q.L. Peroxisome proliferator-activated receptor gamma (PPARG) in yellow catfish Pelteobagrus fulvidraco: Molecular characterization, mRNA expression and transcriptional regulation by insulin in vivo and in vitro. Gen. Comp. Endocrinol. 2015, 212, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Gurmaches, J.; Østbye, T.K.; Navarro, I.; Torgersen, J.; Hevrøy, E.M.; Ruyter, B.; Torstensen, B.E. In vivo and in vitro insulin and fasting control of the transmembrane fatty acid transport proteins in Atlantic salmon (Salmo salar). Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R947–R957. [Google Scholar] [CrossRef] [PubMed]
- Valen, E.; Sandelin, A. Genomic and chromatin signals underlying transcription start-site selection. Trends Genet. 2011, 27, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Bird, A. DNA methylation patterns and epigenetic memory. Genes Dev. 2002, 16, 6–21. [Google Scholar] [CrossRef] [PubMed]
- Chan, T.M.; Leung, K.S.; Lee, K.H. TFBS identification based on genetic algorithm with combined representations and adaptive post-processing. Bioinformatics 2008, 24, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Abell, K.; Bilancio, A.; Clarkson, R.W.; Tiffen, P.G.; Altaparmakov, A.I.; Burdon, T.G.; Watson, C.J. Stat3-induced apoptosis requires a molecular switch in PI(3)K subunit composition. Nat. Cell Biol. 2005, 7, 392–398. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Zhou, Y.; Shen, P. NF-kappa B and its regulation on the immune system. Cell. Mol. Immunol. 2004, 1, 343–350. [Google Scholar] [PubMed]
- Hayden, M.S.; Ghosh, S. Shared principles in NF-κB signaling. Cell 2008, 132, 344–362. [Google Scholar] [CrossRef] [PubMed]
- You, W.J.; Fan, Y.F.; Xu, Y.H.; Wu, K.; Tan, X.Y. Molecular characterization and functional analysis of PPARα promoter in yellow catfish Pelteobagrus fulvidraco. Gene 2017, 627, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.H.; Luo, Z.; Wu, K.; Fan, Y.F.; You, W.J.; Zhang, L.H. Structure and functional analysis of promoters from two liver isoforms of CPT I in grass carp Ctenopharyngodon idella. Int. J. Mol. Sci. 2017, 18, 2405. [Google Scholar] [CrossRef] [PubMed]
- Rieusset, J.; Auwerx, J.; Vidal, H. Regulation of gene expression by activation of the peroxisome proliferator-activated receptor g with Rosiglitazone (BRL 49653) in human adipocytes. Biochem. Biophys. Res. Commun. 1999, 265, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Rieusset, J.; Chambrier, C.; Bouzakri, K.; Dusserre, E.; Auwerx, J.; Riou, J.P.; Vidal, H. The expression of the p85asubunit of phosphatidylinositol 3-kinase is induced by activation of the peroxisome proliferator-activated receptor g in human adipocytes. Diabetologia 2001, 44, 544–554. [Google Scholar] [CrossRef] [PubMed]
- Rieusset, J.; Roques, M.; Bouzakri, K.; Chevillotte, E.; Vidal, H. Regulation of p85aphosphatidylinositol-3-kinase expression by peroxisome proliferator-activated receptors (PPARs) in human muscle cells. FEBS Lett. 2001, 502, 98–100. [Google Scholar] [CrossRef]
- Ktistaki, E.; Talianidis, I. Modulation of hepatic gene expression by hepatocyte nuclear factor 1. Science 1997, 277, 109–112. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, Y.; Gao, Y.; Cui, Y.; Liu, H.; Li, M.; Tian, Y. Downregulation of HNF1 homeobox B is associated with drug resistance in ovarian cancer. Oncol. Rep. 2014, 32, 979–988. [Google Scholar] [CrossRef] [PubMed]
- Kolasa, I.K.; Rembiszewska, A.; Felisiak, A.; Ziolkowska-Seta, I.; Murawska, M.; Moes, J.; Kupryjanczyk, J. PIK3CA amplification associates with resistance to chemotherapy in ovarian cancer patients. Cancer Biol. Ther. 2009, 8, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Sellars, M.; Kastner, P.; Chan, S. Ikaros in B cell development and function. World J. Biol. Chem. 2011, 2, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Balzarotti, G.; Tibolla, G.; Bonacina, F.; D’Alonzo, C.; Dhyani, A.; Falasca, M.; Catapano, A. PI3K-C2B plays a key role in the activation and the proliferation of t lymphocytes: Impact on vascular diseases. Atherosclerosis 2015, 241, e35. [Google Scholar] [CrossRef]
- Obsil, T.; Obsilova, V. Structure/function relationships underlying regulation of FOXO transcription factors. Oncogene 2008, 27, 2263–2275. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, M.; Han, S.; Kitamura, T.; Accili, D. Dual role of transcription factor FoxO1 in controlling hepatic insulin sensitivity and lipid metabolism. J. Clin. Investig. 2006, 116, 2464–2472. [Google Scholar] [CrossRef] [PubMed]
- Kamagate, A.; Dong, H.H. FoxO1 integrates insulin signaling to VLDL production. Cell Cycle 2008, 7, 3162–3170. [Google Scholar] [CrossRef] [PubMed]
- Zou, P.; Liu, L.; Zheng, L.; Liu, L.; Stoneman, R.E.; Cho, A.; Cheng, Z. Targeting FoxO1 with AS1842856 suppresses adipogenesis. Cell Cycle 2014, 13, 3759–3767. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zheng, L.D.; Zou, P.; Brooke, J.; Smith, C.; Long, Y.C.; Cheng, Z. FoxO1 antagonist suppresses autophagy and lipid droplet growth in adipocytes. Cell Cycle 2016, 15, 2033–2041. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, M.Q.; Luo, Z.; Wu, K.; Zhu, Q.L.; Zheng, J.L.; Zhang, L.H.; Chen, Q.L. Regulation of insulin on lipid metabolism in freshly isolated hepatocytes from yellow catfish (Pelteobagrus fulvidraco). Comp. Biochem. Physiol. 2014, 177B, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.L.; Luo, Z.; Wu, K.; Huang, C.; Zhuo, M.Q.; Song, Y.F.; Hu, W. Differential effects of dietary copper deficiency and excess on lipid metabolism in yellow catfish Pelteobagrus fulvidraco. Comp. Biochem. Physiol. 2015, 184B, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Makoveichuk, E.; Vorrsjö, E.; Olivecrona, T.; Olivecrona, G. TNF-α decreases lipoprotein lipase activity in 3T3-L1 adipocytes by up-regulation of angiopoietin-like protein 4. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2017, 1862, 533–540. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Xia, Z. FOXO1 Overactivation via Enhancing CD36 Exacerbates Myocardial Ischemia Reperfusion Injury in Diabetic Rats. FASEB J. 2017, 31, 673. [Google Scholar]
- Chakrabarti, P.; Kandror, K.V. FoxO1 controls insulin-dependent adipose triglyceride lipase (ATGL) expression and lipolysis in adipocytes. J. Biol. Chem. 2009, 284, 13296–13300. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, E.; Flier, E.; Molle, D.; Accili, D.; McGraw, T.E. Hyperinsulinemia leads to uncoupled insulin regulation of the GLUT4 glucose transporter and the FoxO1 transcription factor. Proc. Natl. Acad. Sci. USA 2011, 108, 10162–10167. [Google Scholar] [CrossRef] [PubMed]
- Primers, P. High-efficiency thermal asymmetric interlaced PCR for amplification of unknown flanking sequences. Biotechniques 2007, 43, 649–656. [Google Scholar]
- Read, M.A.; Cordle, S.R.; Veach, R.A.; Carlisle, C.D.; Hawiger, J. Cell-free pool of CD14 mediates activation of transcription factor NF-kappa B by lipopolysaccharide in human endothelial cells. Proc. Natl. Acad. Sci. USA 1993, 90, 9887–9891. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Chakrabarty, K.; Leveille, G.A. Acetyl CoA carboxylase and fatty acid synthetase activities in liver and adipose tissue of meal-fed rats. Proc. Soc. Exp. Biol. Med. 1969, 131, 1051–1054. [Google Scholar] [CrossRef] [PubMed]
- Barroso, J.B.; Peragon, J.; Garcıa-Salguero, L.; Higuera, M.; Lupianez, J.A. Variations in the kinetic behaviour of the NADPH-production systems in different tissues of the trout when fed on an amino-acid-based diet at different frequencies. Int. J. Biochem. Cell Biol. 1999, 31, 277–290. [Google Scholar] [CrossRef]
- Wise, E.M.; Ball, E.G. Malic enzyme and lipogenesis. Proc. Natl. Acad. Sci. USA 1964, 52, 1255–1263. [Google Scholar] [CrossRef] [PubMed]
- Bernt, E.; Bergmeyer, H.U. Isocitrate dehydrogenase. In Methods of Enzymatic Analysis; Bergmeyer, H.U., Ed.; Academic Press: New York, NY, USA, 1974; Volume 2, pp. 624–627. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-type binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Vandesompele, J.; De-Preter, K.; Pattyn, F.; Poppe, B.; Van-Roy, N.; De-Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, R0034. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Li, Y.; Cheng, J.; Chen, L.; Zeng, M.; Wu, Y.; Wang, J.; Zhang, J.; Chu, W. Transcriptome analysis and postprandial expression of amino acid transporter genes in the fast muscles and gut of Chinese perch (Siniperca chuatsi). PLoS ONE 2016, 11, e0159533. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Li, Y.; Cheng, J.; Chen, L.; Zhu, X.; Feng, Z.; Zhang, J.; Chu, W. Daily rhythmicity of the clock gene transcripts in fast and slow muscle fibers from Chinese perch (Siniperca chuatsi). BMC Genom. 2016, 17, 1008. [Google Scholar] [CrossRef] [PubMed]











© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhuo, M.-Q.; Luo, Z.; Xu, Y.-H.; Li, D.-D.; Pan, Y.-X.; Wu, K. Functional Analysis of Promoters from Three Subtypes of the PI3K Family and Their Roles in the Regulation of Lipid Metabolism by Insulin in Yellow Catfish Pelteobagrus fulvidraco. Int. J. Mol. Sci. 2018, 19, 265. https://doi.org/10.3390/ijms19010265
Zhuo M-Q, Luo Z, Xu Y-H, Li D-D, Pan Y-X, Wu K. Functional Analysis of Promoters from Three Subtypes of the PI3K Family and Their Roles in the Regulation of Lipid Metabolism by Insulin in Yellow Catfish Pelteobagrus fulvidraco. International Journal of Molecular Sciences. 2018; 19(1):265. https://doi.org/10.3390/ijms19010265
Chicago/Turabian StyleZhuo, Mei-Qin, Zhi Luo, Yi-Huan Xu, Dan-Dan Li, Ya-Xiong Pan, and Kun Wu. 2018. "Functional Analysis of Promoters from Three Subtypes of the PI3K Family and Their Roles in the Regulation of Lipid Metabolism by Insulin in Yellow Catfish Pelteobagrus fulvidraco" International Journal of Molecular Sciences 19, no. 1: 265. https://doi.org/10.3390/ijms19010265
APA StyleZhuo, M.-Q., Luo, Z., Xu, Y.-H., Li, D.-D., Pan, Y.-X., & Wu, K. (2018). Functional Analysis of Promoters from Three Subtypes of the PI3K Family and Their Roles in the Regulation of Lipid Metabolism by Insulin in Yellow Catfish Pelteobagrus fulvidraco. International Journal of Molecular Sciences, 19(1), 265. https://doi.org/10.3390/ijms19010265