Imaging Collagen in Scar Tissue: Developments in Second Harmonic Generation Microscopy for Biomedical Applications

Abstract
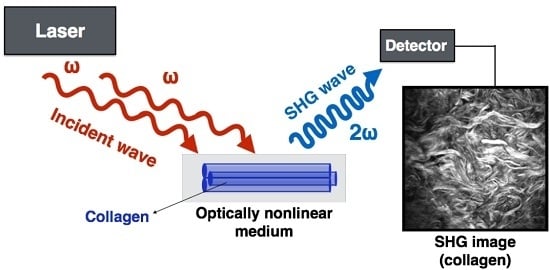
1. Introduction
2. The Extracellular Matrix
3. Fibrillar Collagen
4. Collagen Imaging Using Second Harmonic Generation (SHG) Microscopy
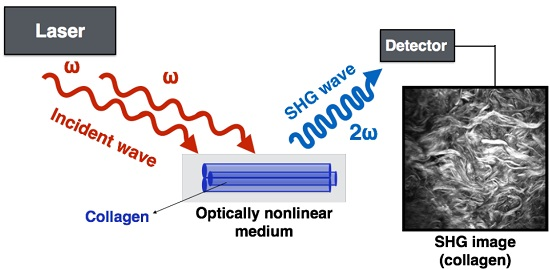
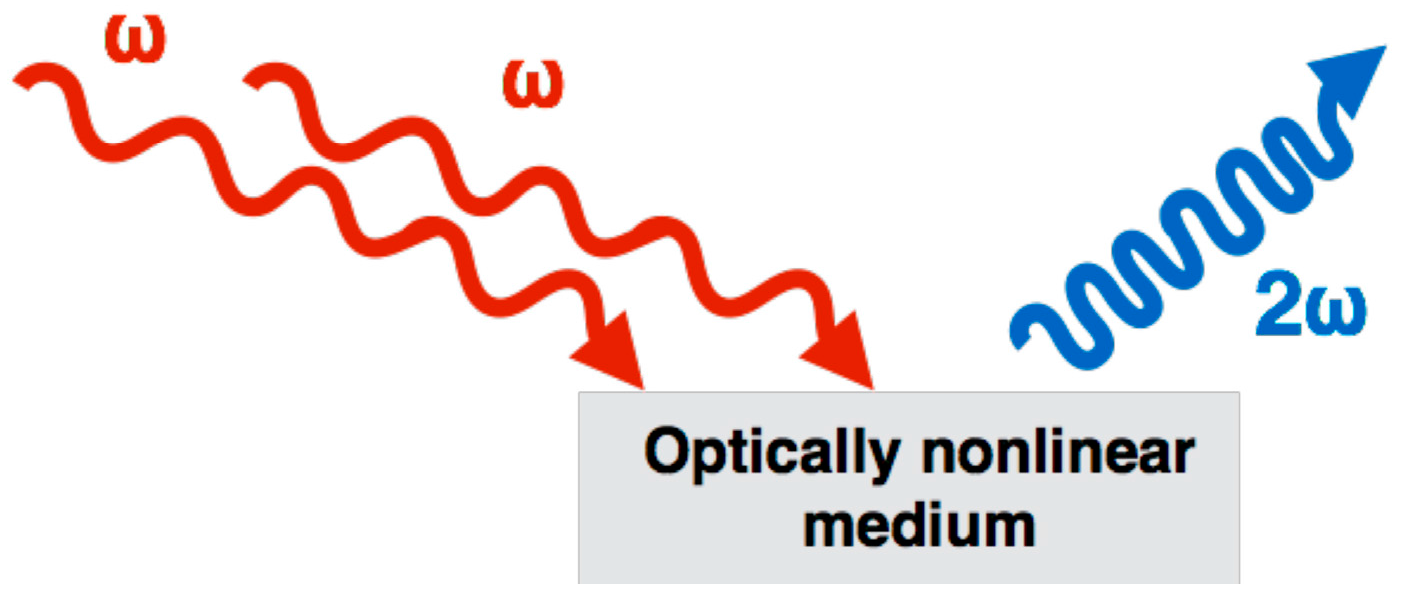
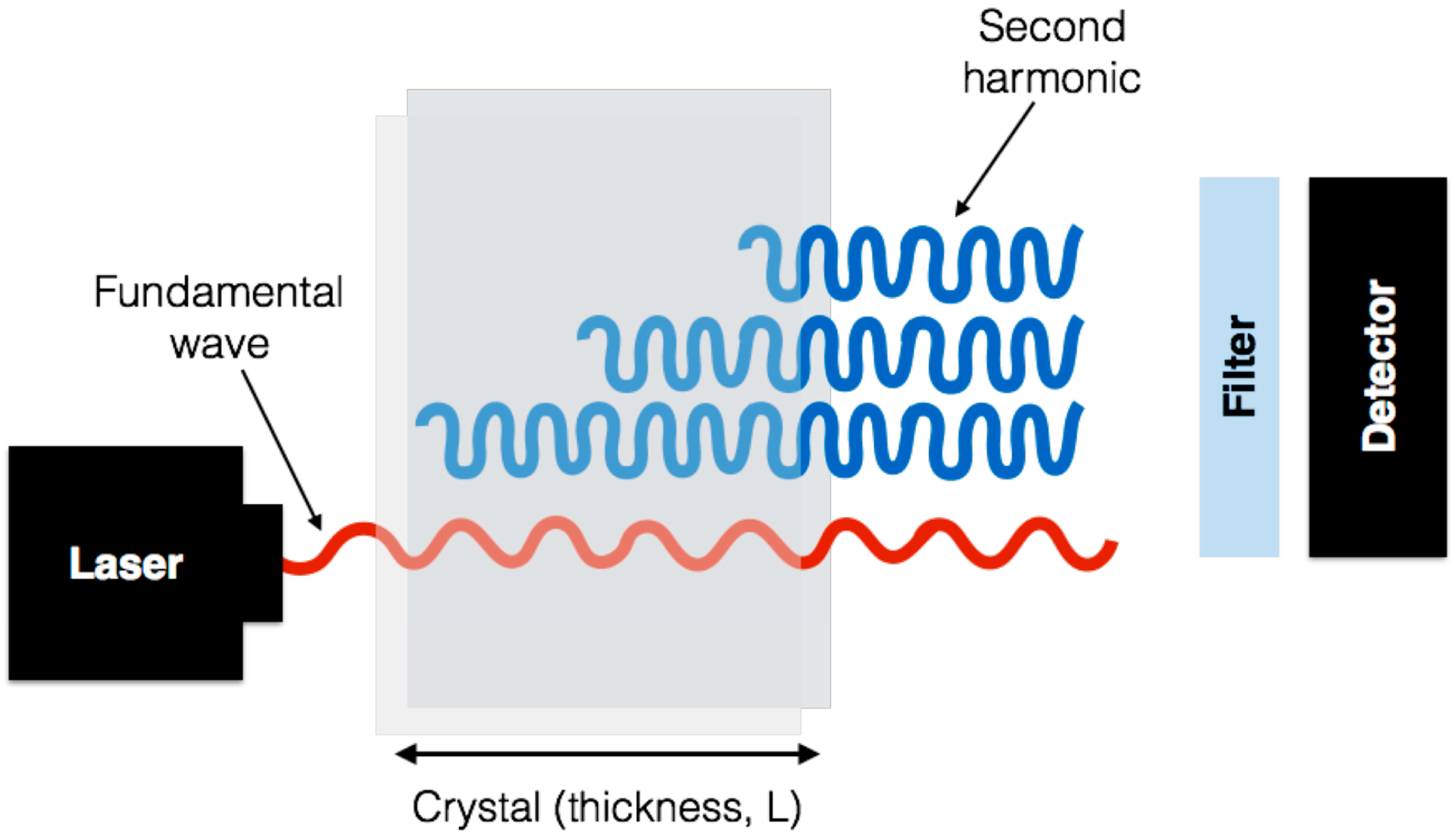
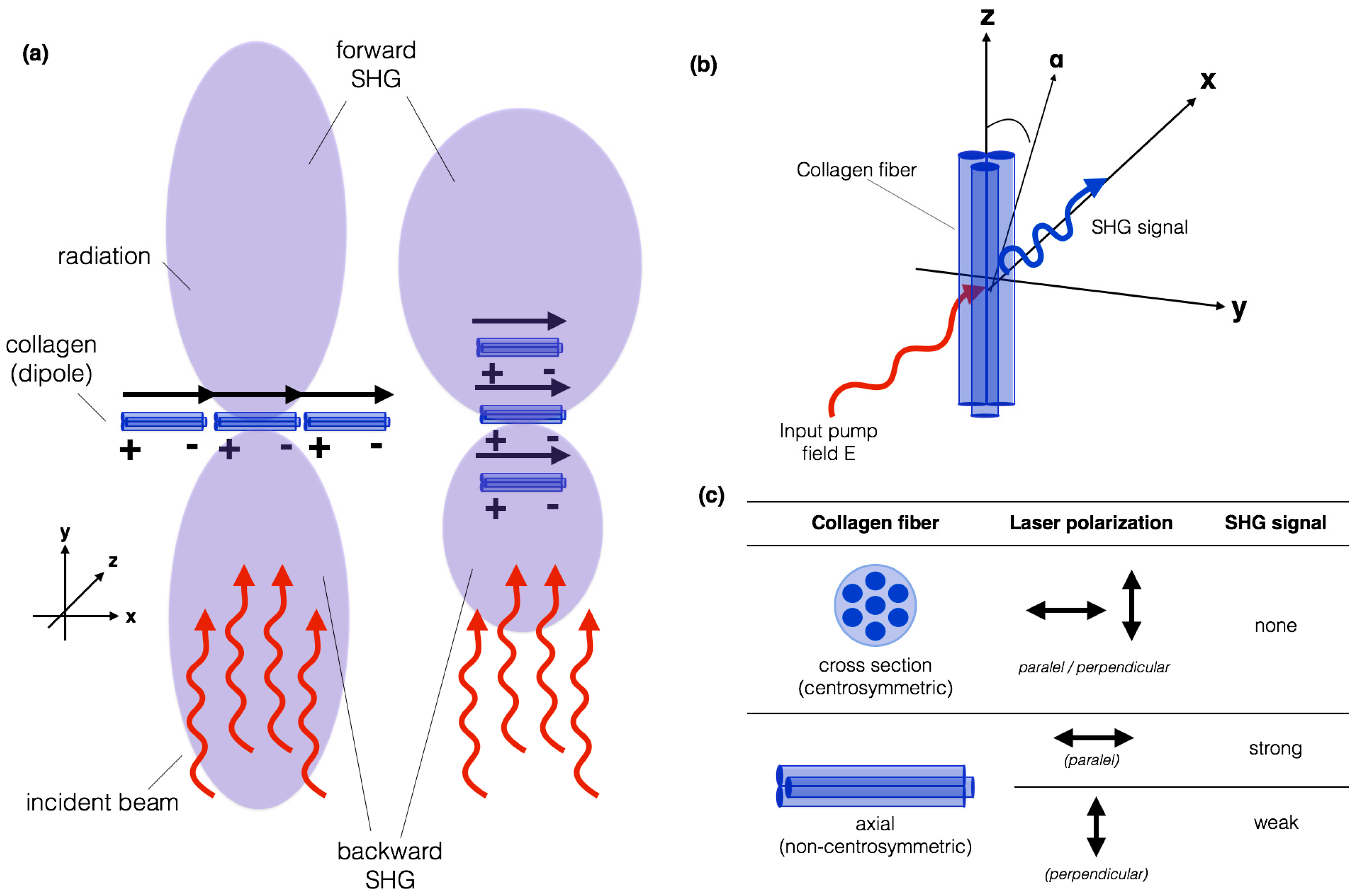
4.1. What Is SHG and How Does It Work?
4.2. How Does Fibrillar Collagen Generate SHG Signal
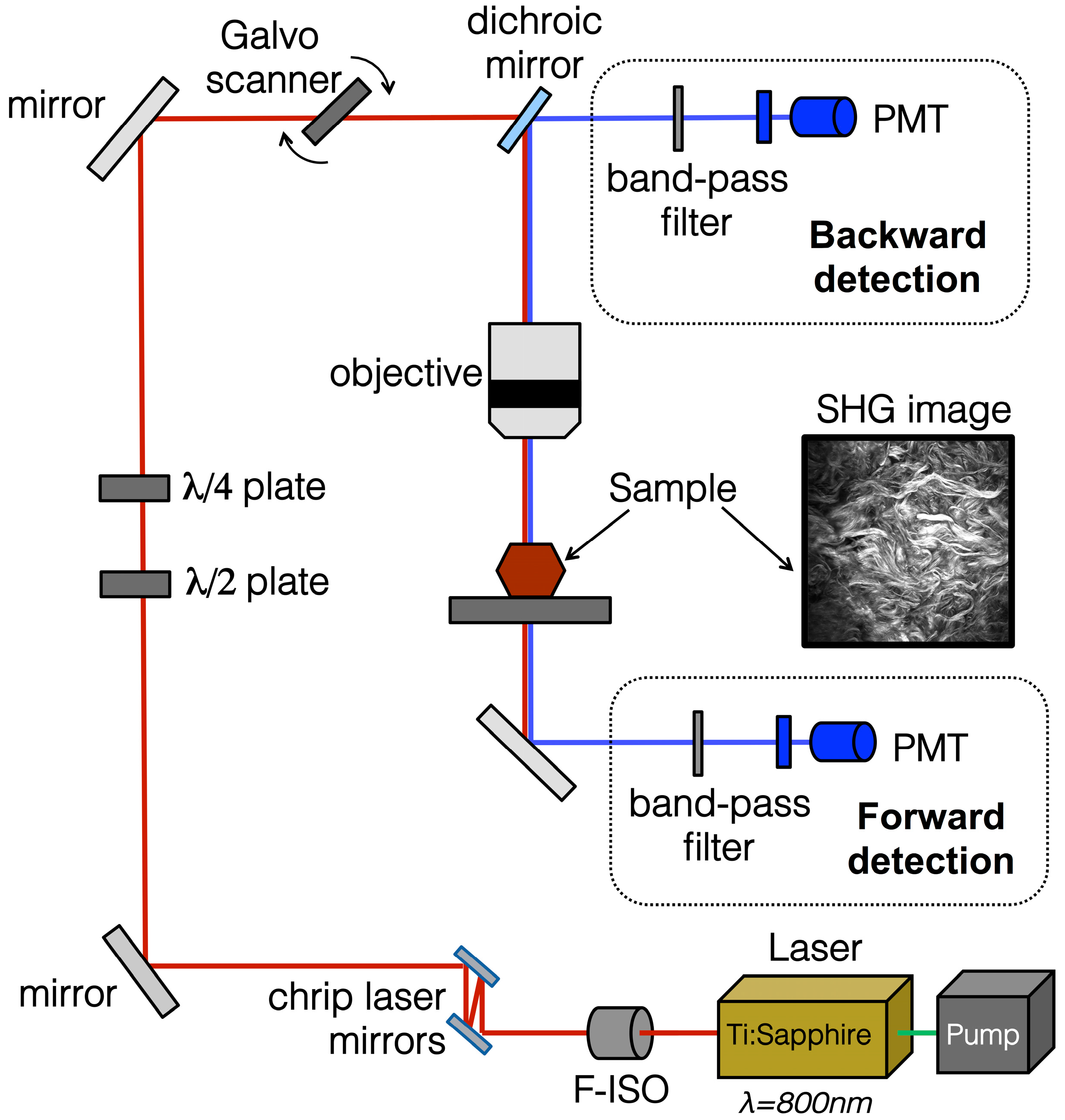
4.3. SHG Imaging Systems
4.4. Recent In Vivo Instrumentation Advances
5. Alterations in Fibrillar Collagens in the Disease State
5.1. Skin
5.2. The Lung
5.3. Cardiovascular System
5.4. Tendons and Ligaments
5.5. The Eye
5.6. In Vitro Models: Collagen Gels
6. Quantitative Image Analysis Methods
6.1. Intensity-Based Analysis
6.2. Forward–Backward SHG-Signal
6.3. Polarization
6.4. Transform-Based Methods
6.5. Texture Analysis
7. Limitations
8. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| SHG | Second harmonic generation |
| THG | Third harmonic generation |
| B-SHG | Backward second harmonic generation |
| F-SHG | Forward second harmonic generation |
| Col | Collagen |
| COPD | Chronic obstructive pulmonary disease |
| CARS | Coherent anti-Stokes raman spectroscopy |
| MEMS | Microelectromechanical system |
| GLCM | Gray level co-occurrence matrix |
| IDM | Inverse difference moment |
| RLM | Run length matrix |
| AMT | Angle Measure Technique |
| FFT | Fast Fourier Transform |
| ECM | Extracellular matrix |
| 3D | 3-dimension |
| LDL | Low density lipids |
| ASC | Apdipose stem cell |
| NLOM | Non-linear optical microscopy |
| TPEF | Two photon electron fluorescence |
| BPD | Bronchopulmonary dysplasia |
References
- Konigova, R.; Rychterova, V. Marjolin’s ulcer. Acta Chir. Plast. 2000, 42, 91–94. [Google Scholar] [PubMed]
- Trent, J.T.; Kirsner, R.S. Wounds and malignancy. Adv. Skin Wound Care 2003, 16, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Aarabi, S.; Longaker, M.T.; Gurtner, G.C. Hypertrophic scar formation following burns and trauma: New approaches to treatment. PLoS Med. 2007, 4, e234. [Google Scholar] [CrossRef] [PubMed]
- Singer, A.J.; Clark, R.A. Cutaneous wound healing. N. Engl. J. Med. 1999, 341, 738–746. [Google Scholar] [CrossRef] [PubMed]
- Colwell, A.S.; Longaker, M.T.; Lorenz, H.P. Fetal wound healing. Front. Biosci. 2003, 8, s1240–s1248. [Google Scholar] [PubMed]
- Brem, H.; Tomic-Canic, M. Cellular and molecular basis of wound healing in diabetes. J. Clin. Investig. 2007, 117, 1219–1222. [Google Scholar] [CrossRef] [PubMed]
- Sen, C.K.; Gordillo, G.M.; Roy, S.; Kirsner, R.; Lambert, L.; Hunt, T.K.; Gottrup, F.; Gurtner, G.C.; Longaker, M.T. Human skin wounds: A major and snowballing threat to public health and the economy. Wound Repair Regen. 2009, 17, 763–771. [Google Scholar] [CrossRef] [PubMed]
- Reish, R.G.; Eriksson, E. Scars: A review of emerging and currently available therapies. Plast. Reconstr. Surg. 2008, 122, 1068–1078. [Google Scholar] [CrossRef] [PubMed]
- Richmond, N.A.; Lamel, S.A.; Davidson, J.M.; Martins-Green, M.; Sen, C.K.; Tomic-Canic, M.; Vivas, A.C.; Braun, L.R.; Kirsner, R.S. US-National Institutes of Health-funded research for cutaneous wounds in 2012. Wound Repair Regen. 2013, 21, 789–792. [Google Scholar] [CrossRef] [PubMed]
- Mertz, J. Nonlinear microscopy: New techniques and applications. Curr. Opin. Neurobiol. 2004, 14, 610–616. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.M.; Zipfel, W.R.; Webb, W.W. Multiphoton microscopy in biological research. Curr. Opin. Chem. Biol. 2001, 5, 603–608. [Google Scholar] [CrossRef]
- Yue, S.; Slipchenko, M.N.; Cheng, J.X. Multimodal Nonlinear Optical Microscopy. Laser Photon. Rev. 2011, 5, 10. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, W.R.; Williams, R.M.; Webb, W.W. Nonlinear magic: Multiphoton microscopy in the biosciences. Nat. Biotechnol. 2003, 21, 1369–1377. [Google Scholar] [CrossRef] [PubMed]
- Campagnola, P. Second harmonic generation imaging microscopy: Applications to diseases diagnostics. Anal. Chem. 2011, 83, 3224–3231. [Google Scholar] [CrossRef] [PubMed]
- Campagnola, P.J.; Loew, L.M. Second-harmonic imaging microscopy for visualizing biomolecular arrays in cells, tissues and organisms. Nat. Biotechnol. 2003, 21, 1356–1360. [Google Scholar] [CrossRef] [PubMed]
- Campagnola, P.J.; Millard, A.C.; Terasaki, M.; Hoppe, P.E.; Malone, C.J.; Mohler, W.A. Three-dimensional high-resolution second-harmonic generation imaging of endogenous structural proteins in biological tissues. Biophys. J. 2002, 82, 493–508. [Google Scholar] [CrossRef]
- Zipfel, W.R.; Williams, R.M.; Christie, R.; Nikitin, A.Y.; Hyman, B.T.; Webb, W.W. Live tissue intrinsic emission microscopy using multiphoton-excited native fluorescence and second harmonic generation. Proc. Natl. Acad. Sci. USA 2003, 100, 7075–7080. [Google Scholar] [CrossRef] [PubMed]
- Masihzadeh, O.; Schlup, P.; Bartels, R.A. Label-free second harmonic generation holographic microscopy of biological specimens. Opt. Express 2010, 18, 9840–9851. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, C.P.; Olsen, B.R.; Ganikhanov, F.; Legare, F. Multimodal nonlinear optical imaging of collagen arrays. J. Struct. Biol. 2008, 164, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Ajeti, V.; Nadiarnykh, O.; Ponik, S.M.; Keely, P.J.; Eliceiri, K.W.; Campagnola, P.J. Structural changes in mixed Col I/Col V collagen gels probed by SHG microscopy: Implications for probing stromal alterations in human breast cancer. Biomed. Opt. Express 2011, 2, 2307–2316. [Google Scholar] [CrossRef] [PubMed]
- Cicchi, R.; Massi, D.; Sestini, S.; Carli, P.; De Giorgi, V.; Lotti, T.; Pavone, F.S. Multidimensional non-linear laser imaging of Basal Cell Carcinoma. Opt. Express 2007, 15, 10135–10148. [Google Scholar] [CrossRef] [PubMed]
- Conklin, M.W.; Eickhoff, J.C.; Riching, K.M.; Pehlke, C.A.; Eliceiri, K.W.; Provenzano, P.P.; Friedl, A.; Keely, P.J. Aligned collagen is a prognostic signature for survival in human breast carcinoma. Am. J. Pathol. 2011, 178, 1221–1232. [Google Scholar] [CrossRef] [PubMed]
- Dimitrow, E.; Ziemer, M.; Koehler, M.J.; Norgauer, J.; Konig, K.; Elsner, P.; Kaatz, M. Sensitivity and specificity of multiphoton laser tomography for in vivo and ex vivo diagnosis of malignant melanoma. J. Investig. Dermatol. 2009, 129, 1752–1758. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Giese, G.; Bille, J. Second harmonic generation imaging of collagen fibrils in cornea and sclera. Opt. Express 2005, 13, 5791–5797. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, N.D.; Brewer, M.A.; Utzinger, U. Endogenous optical biomarkers of ovarian cancer evaluated with multiphoton microscopy. Cancer Epidemiol. Biomark. Prev. 2007, 16, 2048–2057. [Google Scholar] [CrossRef] [PubMed]
- Kwon, G.P.; Schroeder, J.L.; Amar, M.J.; Remaley, A.T.; Balaban, R.S. Contribution of macromolecular structure to the retention of low-density lipoprotein at arterial branch points. Circulation 2008, 117, 2919–2927. [Google Scholar] [CrossRef] [PubMed]
- Lacomb, R.; Nadiarnykh, O.; Campagnola, P.J. Quantitative second harmonic generation imaging of the diseased state osteogenesis imperfecta: Experiment and simulation. Biophys. J. 2008, 94, 4504–4514. [Google Scholar] [CrossRef] [PubMed]
- Le, T.T.; Langohr, I.M.; Locker, M.J.; Sturek, M.; Cheng, J.X. Label-free molecular imaging of atherosclerotic lesions using multimodal nonlinear optical microscopy. J. Biomed. Opt. 2007, 12, 054007. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.J.; Jee, S.H.; Kuo, C.J.; Wu, R.J.; Lin, W.C.; Chen, J.S.; Liao, Y.H.; Hsu, C.J.; Tsai, T.F.; Chen, Y.F.; et al. Discrimination of basal cell carcinoma from normal dermal stroma by quantitative multiphoton imaging. Opt. Lett. 2006, 31, 2756–2758. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.J.; Wu, R., Jr.; Tan, H.Y.; Lo, W.; Lin, W.C.; Young, T.H.; Hsu, C.J.; Chen, J.S.; Jee, S.H.; Dong, C.Y. Evaluating cutaneous photoaging by use of multiphoton fluorescence and second-harmonic generation microscopy. Opt. Lett. 2005, 30, 2275–2277. [Google Scholar] [CrossRef] [PubMed]
- Lo, W.; Teng, S.W.; Tan, H.Y.; Kim, K.H.; Chen, H.C.; Lee, H.S.; Chen, Y.F.; So, P.T.; Dong, C.Y. Intact corneal stroma visualization of GFP mouse revealed by multiphoton imaging. Microsc. Res. Tech. 2006, 69, 973–975. [Google Scholar] [CrossRef] [PubMed]
- Nadiarnykh, O.; LaComb, R.B.; Brewer, M.A.; Campagnola, P.J. Alterations of the extracellular matrix in ovarian cancer studied by Second Harmonic Generation imaging microscopy. BMC Cancer 2010, 10, 94. [Google Scholar] [CrossRef] [PubMed]
- Provenzano, P.P.; Eliceiri, K.W.; Campbell, J.M.; Inman, D.R.; White, J.G.; Keely, P.J. Collagen reorganization at the tumor-stromal interface facilitates local invasion. BMC Med. 2006, 4, 38. [Google Scholar] [CrossRef] [PubMed]
- Raub, C.B.; Suresh, V.; Krasieva, T.; Lyubovitsky, J.; Mih, J.D.; Putnam, A.J.; Tromberg, B.J.; George, S.C. Noninvasive assessment of collagen gel microstructure and mechanics using multiphoton microscopy. Biophys. J. 2007, 92, 2212–2222. [Google Scholar] [CrossRef] [PubMed]
- Sahai, E.; Wyckoff, J.; Philippar, U.; Segall, J.E.; Gertler, F.; Condeelis, J. Simultaneous imaging of GFP, CFP and collagen in tumors in vivo using multiphoton microscopy. BMC Biotechnol. 2005, 5, 14. [Google Scholar] [CrossRef] [PubMed]
- Schenke-Layland, K.; Xie, J.; Angelis, E.; Starcher, B.; Wu, K.; Riemann, I.; MacLellan, W.R.; Hamm-Alvarez, S.F. Increased degradation of extracellular matrix structures of lacrimal glands implicated in the pathogenesis of Sjogren’s syndrome. Matrix Biol. 2008, 27, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Strupler, M.; Pena, A.M.; Hernest, M.; Tharaux, P.L.; Martin, J.L.; Beaurepaire, E.; Schanne-Klein, M.C. Second harmonic imaging and scoring of collagen in fibrotic tissues. Opt. Express 2007, 15, 4054–4065. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Chang, S.; Tai, D.C.; Tan, N.; Xiao, G.; Tang, H.; Yu, H. Nonlinear optical microscopy: Use of second harmonic generation and two-photon microscopy for automated quantitative liver fibrosis studies. J. Biomed. Opt. 2008, 13, 064010. [Google Scholar] [CrossRef] [PubMed]
- Ko, A.C.; Ridsdale, A.; Mostaço-Guidolin, L.B.; Major, A.; Stolow, A.; Sowa, M.G. Nonlinear optical microscopy in decoding arterial diseases. Biophys. Rev. 2012, 4, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Mostaço-Guidolin, L.B.; Ko, A.C.; Wang, F.; Xiang, B.; Hewko, M.; Tian, G.; Major, A.; Shiomi, M.; Sowa, M.G. Collagen morphology and texture analysis: From statistics to classification. Sci. Rep. 2013, 3, 2190. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.W.; Langohr, I.M.; Sturek, M.; Cheng, J.X. Imaging and quantitative analysis of atherosclerotic lesions by CARS-based multimodal nonlinear optical microscopy. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 1342–1348. [Google Scholar] [CrossRef] [PubMed]
- Zoumi, A.; Yeh, A.; Tromberg, B.J. Imaging cells and extracellular matrix in vivo by using second-harmonic generation and two-photon excited fluorescence. Proc. Natl. Acad. Sci. USA 2002, 99, 11014–11019. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Takai, K.; Weaver, V.M.; Werb, Z. Extracellular matrix degradation and remodeling in development and disease. Cold Spring Harb. Perspect. Biol. 2011, 3, a005058. [Google Scholar] [CrossRef] [PubMed]
- Mecham, R.P. Overview of extracellular matrix. Curr. Protoc. Cell Biol. 2012. [Google Scholar] [CrossRef]
- Mouw, J.K.; Ou, G.; Weaver, V.M. Extracellular matrix assembly: A multiscale deconstruction. Nat. Rev. Mol. Cell Biol. 2014, 15, 771–785. [Google Scholar] [CrossRef] [PubMed]
- Rozario, T.; DeSimone, D.W. The extracellular matrix in development and morphogenesis: A dynamic view. Dev. Biol. 2010, 341, 126–140. [Google Scholar] [CrossRef] [PubMed]
- Barczyk, M.M.; Olsen, L.H.; da Franca, P.; Loos, B.G.; Mustafa, K.; Gullberg, D.; Bolstad, A.I. A role for α11β1 integrin in the human periodontal ligament. J. Dent. Res. 2009, 88, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Dzamba, B.J.; Keene, D.R.; Isogai, Z.; Charbonneau, N.L.; Karaman-Jurukovska, N.; Simon, M.; Sakai, L.Y. Assembly of epithelial cell fibrillins. J. Investig. Dermatol. 2001, 117, 1612–1620. [Google Scholar] [PubMed]
- Ramirez, F.; Sakai, L.Y. Biogenesis and function of fibrillin assemblies. Cell Tissue Res. 2010, 339, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Durbeej, M. Laminins. Cell Tissue Res. 2010, 339, 259. [Google Scholar] [CrossRef] [PubMed]
- Gordon, M.K.; Hahn, R.A. Collagens. Cell Tissue Res. 2010, 339, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Brodsky, B.; Persikov, A.V. Molecular structure of the collagen triple helix. Adv. Protein Chem. 2005, 70, 301–339. [Google Scholar] [PubMed]
- Celerin, M.; Ray, J.M.; Schisler, N.J.; Day, A.W.; Stetler-Stevenson, W.G.; Laudenbach, D.E. Fungal fimbriae are composed of collagen. EMBO J. 1996, 15, 4445–4453. [Google Scholar] [PubMed]
- King, N.; Westbrook, M.J.; Young, S.L.; Kuo, A.; Abedin, M.; Chapman, J.; Fairclough, S.; Hellsten, U.; Isogai, Y.; Letunic, I.; et al. The genome of the choanoflagellate Monosiga brevicollis and the origin of metazoans. Nature 2008, 451, 783–788. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, M.; Jacobsson, M.; Bjorck, L. Genome-based identification and analysis of collagen-related structural motifs in bacterial and viral proteins. J. Biol. Chem. 2003, 278, 32313–32316. [Google Scholar] [CrossRef] [PubMed]
- Fratzl, P.; Misof, K.; Zizak, I.; Rapp, G.; Amenitsch, H.; Bernstorff, S. Fibrillar structure and mechanical properties of collagen. J. Struct. Biol. 1998, 122, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Hulmes, D.J. Building collagen molecules, fibrils, and suprafibrillar structures. J. Struct. Biol. 2002, 137, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Hulmes, D.J. Collagen Structure and Mechanics; Springer: Berlin, Germany, 2008. [Google Scholar]
- Myllyharju, J.; Kivirikko, K.I. Collagens, modifying enzymes and their mutations in humans, flies and worms. Trends Genet. 2004, 20, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Ricard-Blum, S.; Ruggiero, F. The collagen superfamily: From the extracellular matrix to the cell membrane. Pathol. Biol. 2005, 53, 430–442. [Google Scholar] [CrossRef] [PubMed]
- Cicchi, R.; Vogler, N.; Kapsokalyvas, D.; Dietzek, B.; Popp, J.; Pavone, F.S. From molecular structure to tissue architecture: Collagen organization probed by SHG microscopy. J. Biophotonics 2013, 6, 129–142. [Google Scholar] [CrossRef] [PubMed]
- Dunn, K.W.; Young, P.A. Principles of multiphoton microscopy. Nephron Exp. Nephrol. 2006, 103, e33–e40. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Zipfel, W.; Shear, J.B.; Williams, R.M.; Webb, W.W. Multiphoton fluorescence excitation: New spectral windows for biological nonlinear microscopy. Proc. Natl. Acad. Sci. USA 1996, 93, 10763–10768. [Google Scholar] [CrossRef] [PubMed]
- Bass, M.; van Strvland, E.W.; Williams, D.R.; Wolfe, W.L. Handbook of Optics; McGraw-Hill: New York, NY, USA, 2001; Volume 2. [Google Scholar]
- Sutherland, R.L. Handbook of Nonlinear Optics; CRC Press: Boca Raton, FL, USA, 2003. [Google Scholar]
- Svirko, Y.P.; Zheludev, N.I. Polarization of Light in Nonlineear Optics; Wiley-VCH: Weinheim, Germany, 2000. [Google Scholar]
- Boyd, G.D.; Kleinman, D.A. Parametric interation of focused Gaussian light beams. J. Appl. Phys. 1968, 39, 3597. [Google Scholar] [CrossRef]
- Brjorkholm, J.E. Opitcal second-harmonic generation using a focused Gaussian laser beam. Phys. Rev. 1966, 142, 126–136. [Google Scholar] [CrossRef]
- Dmitriev, V.G.; Gurzadyan, G.G.; Nikogosyan, D.N.; Lotsch, H.K.V. Handbook of Nonlinear Optical Crystals; Springer: Berlin, Germany, 1999. [Google Scholar]
- Sapaev, U.K.; Kulagin, I.A.; Usmanov, T. Theory of second-harmonic generation for limited laser beams in nonlinear crystals. J. Opt. B Quantum Semiclass. Opt. 2003, 5, 355–356. [Google Scholar] [CrossRef]
- Chen, X.; Nadiarynkh, O.; Plotnikov, S.; Campagnola, P.J. Second harmonic generation microscopy for quantitative analysis of collagen fibrillar structure. Nat. Protoc. 2012, 7, 654–669. [Google Scholar] [CrossRef] [PubMed]
- Masters, B.R.; So, P. Handbook of Biomedical Nonlinear Optical Microscopy; Oxford University Press: Oxford, UK, 2008. [Google Scholar]
- Roth, S.; Freund, I. Second harmonic generation in collagen. J. Chem. Phys. 1979, 70, 1637–1643. [Google Scholar] [CrossRef]
- Theodossiou, T.A.; Thrasivoulou, C.; Ekwobi, C.; Becker, D.L. Second harmonic generation confocal microscopy of collagen type I from rat tendon cryosections. Biophys. J. 2006, 91, 4665–4677. [Google Scholar] [CrossRef] [PubMed]
- Stoller, P.; Kim, B.M.; Rubenchik, A.M.; Reiser, K.M.; Da Silva, L.B. Polarization-dependent optical second-harmonic imaging of a rat-tail tendon. J. Biomed. Opt. 2002, 7, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Helmchen, F.; Fee, M.S.; Tank, D.W.; Denk, W. A miniature head-mounted two-photon microscope: High-resolution brain imaging in freely moving animals. Neuron 2001, 31, 903–912. [Google Scholar] [CrossRef]
- Llewellyn, M.E.; Barretto, R.P.; Delp, S.L.; Schnitzer, M.J. Minimally invasive high-speed imaging of sarcomere contractile dynamics in mice and humans. Nature 2008, 454, 784–788. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Leng, Y.; Xi, J.; Li, X. Scanning all-fibre-optic endomicroscopy system for 3D nonlinear optical imaging of biological tissues. Opt. Express 2009, 17, 7907–7915. [Google Scholar] [CrossRef] [PubMed]
- Crisafi, F.; Kumar, V.; Perri, A.; Marangoni, M.; Cerullo, G.; Polli, D. Multimodal nonlinear microscope based on a compact fibre-format laser source. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2017, 188, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Xu, Q.; Zhang, Y.; Zhan, Y.; Zheng, W.; Song, L. Fully integrated reflection-mode photoacoustic, two-photon, and second harmonic generation microscopy in vivo. Sci. Rep. 2016, 6, 32240. [Google Scholar] [CrossRef] [PubMed]
- Atsuta, K.; Ogura, Y.; Hase, E.; Minamikawa, T.; Yasui, T. In situ monitoring of collagen fibres in human skin using a photonic-crystal-fibre-coupled, hand-held, second-harmonic-generation microscope. Proc. SPIE 2017. [Google Scholar] [CrossRef]
- Sanchez, G.N.; Sinha, S.; Liske, H.; Chen, X.; Nguyen, V.; Delp, S.L.; Schnitzer, M.J. In Vivo Imaging of Human Sarcomere Twitch Dynamics in Individual Motor Units. Neuron 2015, 88, 1109–1120. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.C.; Campagnola, P.J. Wearable Second Harmonic Generation Imaging: The Sarcomeric Bridge to the Clinic. Neuron 2015, 88, 1067–1069. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Chen, J.; Zhuo, S.; Xiong, S.; Zeng, H.; Jiang, X.; Chen, R.; Xie, S. Nonlinear spectral imaging of human hypertrophic scar based on two-photon excited fluorescence and second-harmonic generation. Br. J. Dermatol. 2009, 161, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Rosin, N.L.; Agabalyan, N.; Olsen, K.; Martufi, G.; Gabriel, V.; Biernaskie, J.; Di Martino, E.S. Collagen structural alterations contribute to stiffening of tissue after split-thickness skin grafting. Wound Repair Regen. 2016, 24, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Su, P.J.; Chen, W.L.; Hong, J.B.; Li, T.H.; Wu, R.J.; Chou, C.K.; Chen, S.J.; Hu, C.; Lin, S.J.; Dong, C.Y. Discrimination of collagen in normal and pathological skin dermis through second-order susceptibility microscopy. Opt. Express 2009, 17, 11161–11671. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.H.; Yang, R.S.; al-Shaikh, R.; Lane, J.M. Collagen in tendon, ligament, and bone healing. A current review. Clin. Orthop. Relat. Res. 1995, 318, 265–278. [Google Scholar]
- Mansfield, J.C.; Winlove, C.P.; Moger, J.; Matcher, S.J. Collagen fibre arrangement in normal and diseased cartilage studied by polarization sensitive nonlinear microscopy. J. Biomed. Opt. 2008, 13, 044020. [Google Scholar] [CrossRef] [PubMed]
- Sivaguru, M.; Durgam, S.; Ambekar, R.; Luedtke, D.; Fried, G.; Stewart, A.; Toussaint, K.C., Jr. Quantitative analysis of collagen fibre organization in injured tendons using Fourier transform-second harmonic generation imaging. Opt. Express 2010, 18, 24983–24993. [Google Scholar] [CrossRef] [PubMed]
- Doras, C.; Taupier, G.; Barsella, A.; Mager, L.; Boeglin, A.; Bulou, H.; Bousquet, P.; Dorkenoo, K.D. Polarization state studies in second harmonic generation signals to trace atherosclerosis lesions. Opt. Express 2011, 19, 15062–15068. [Google Scholar] [CrossRef] [PubMed]
- Lilledahl, M.B.; Haugen, O.A.; de Lange Davies, C.; Svaasand, L.O. Characterization of vulnerable plaques by multiphoton microscopy. J. Biomed. Opt. 2007, 12, 044005. [Google Scholar] [CrossRef] [PubMed]
- Megens, R.T.; oude Egbrink, M.G.; Merkx, M.; Slaaf, D.W.; van Zandvoort, M.A. Two-photon microscopy on vital carotid arteries: Imaging the relationship between collagen and inflammatory cells in atherosclerotic plaques. J. Biomed. Opt. 2008, 13, 044022. [Google Scholar] [CrossRef] [PubMed]
- Mostaço-Guidolin, L.B.; Sowa, M.G.; Ridsdale, A.; Pegoraro, A.F.; Smith, M.S.; Hewko, M.D.; Kohlenberg, E.K.; Schattka, B.; Shiomi, M.; Stolow, A.; et al. Differentiating atherosclerotic plaque burden in arterial tissues using femtosecond CARS-based multimodal nonlinear optical imaging. Biomed. Opt. Express 2010, 1, 59–73. [Google Scholar] [CrossRef] [PubMed]
- Abraham, T.; Fong, G.; Scott, A. Second harmonic generation analysis of early Achilles tendinosis in response to in vivo mechanical loading. BMC Musculoskelet. Disord. 2011, 12, 26. [Google Scholar] [CrossRef] [PubMed]
- Kottmann, R.M.; Sharp, J.; Owens, K.; Salzman, P.; Xiao, G.Q.; Phipps, R.P.; Sime, P.J.; Brown, E.B.; Perry, S.W. Second harmonic generation microscopy reveals altered collagen microstructure in usual interstitial pneumonia versus healthy lung. Respir. Res. 2015, 16, 61. [Google Scholar] [CrossRef] [PubMed]
- Tjin, G.; Xu, P.; Kable, S.H.; Kable, E.P.; Burgess, J.K. Quantification of collagen I in airway tissues using second harmonic generation. J. Biomed. Opt. 2014, 19, 36005. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.Y.; Sun, Y.; Lo, W.; Teng, S.W.; Wu, R.J.; Jee, S.H.; Lin, W.C.; Hsiao, C.H.; Lin, H.C.; Chen, Y.F.; et al. Multiphoton fluorescence and second harmonic generation microscopy for imaging infectious keratitis. J. Biomed. Opt. 2007, 12, 024013. [Google Scholar] [CrossRef] [PubMed]
- Lutz, V.; Sattler, M.; Gallinat, S.; Wenck, H.; Poertner, R.; Fischer, F. Impact of collagen crosslinking on the second harmonic generation signal and the fluorescence lifetime of collagen autofluorescence. Skin Res. Technol. 2012, 18, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Pena, A.M.; Fagot, D.; Olive, C.; Michelet, J.F.; Galey, J.B.; Leroy, F.; Beaurepaire, E.; Martin, J.L.; Colonna, A.; Schanne-Klein, M.C. Multiphoton microscopy of engineered dermal substitutes: Assessment of 3-D collagen matrix remodeling induced by fibroblast contraction. J. Biomed. Opt. 2010, 15, 056018. [Google Scholar] [CrossRef] [PubMed]
- Wolf, K.A.S.; Schacht, V.; Coussens, L.M.; von Andrian, U.H.; van Rheenen, J.; Deryugina, E.; Friedl, P. Collagen-based cell migration models in vitro and in vivo. Semin. Cell Dev. Biol. 2009, 20, 931–941. [Google Scholar] [CrossRef] [PubMed]
- Gurtner, G.C.; Werner, S.; Barrandon, Y.; Longaker, M.T. Wound repair and regeneration. Nature 2008, 453, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Rahmani, W.; Abbasi, S.; Hagner, A.; Raharjo, E.; Kumar, R.; Hotta, A.; Magness, S.; Metzger, D.; Biernaskie, J. Hair follicle dermal stem cells regenerate the dermal sheath, repopulate the dermal papilla, and modulate hair type. Dev. Cell 2014, 31, 543–558. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhu, X.; Huang, Z.; Cai, J.; Chen, R.; Xiong, S.; Chen, G.; Zeng, H. Texture analysis of collagen second-harmonic generation images based on local difference local binary pattern and wavelets differentiates human skin abnormal scars from normal scars. J. Biomed. Opt. 2015, 20, 016021. [Google Scholar] [CrossRef] [PubMed]
- Junker, J.P.; Philip, J.; Kiwanuka, E.; Hackl, F.; Caterson, E.J.; Eriksson, E. Assessing quality of healing in skin: Review of available methods and devices. Wound Repair Regen. 2014, 22, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Kumar, P.; Nayak Badagabettu, S.; Prasad, K.; Kudva, R.; Vasudevarao, R.C. Surgical implications of asymmetric distribution of dermal collagen and elastic fibres in two orientations of skin samples from extremities. Plast. Surg. Int. 2014, 2014, 364573. [Google Scholar] [CrossRef] [PubMed]
- Verhaegen, P.D.; van Zuijlen, P.P.; Pennings, N.M.; van Marle, J.; Niessen, F.B.; van der Horst, C.M.; Middelkoop, E. Differences in collagen architecture between keloid, hypertrophic scar, normotrophic scar, and normal skin: An objective histopathological analysis. Wound Repair Regen. 2009, 17, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Hollinsky, C.; Sandberg, S. Measurement of the tensile strength of the ventral abdominal wall in comparison with scar tissue. Clin. Biomech. 2007, 22, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Bateman, E.D.; Turner-Warwick, M.; Adelmann-Grill, B.C. Immunohistochemical study of collagen types in human foetal lung and fibrotic lung disease. Thorax 1981, 36, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Thibeault, D.W.; Mabry, S.M.; Ekekezie, I.I.; Zhang, X.; Truog, W.E. Collagen scaffolding during development and its deformation with chronic lung disease. Pediatrics 2003, 111, 766–776. [Google Scholar] [CrossRef] [PubMed]
- Bradley, K.H.; McConnell, S.D.; Crystal, R.G. Lung collagen composition and synthesis. Characterization and changes with age. J. Biol. Chem. 1974, 249, 2674–2683. [Google Scholar] [PubMed]
- Mizikova, I.; Ruiz-Camp, J.; Steenbock, H.; Madurga, A.; Vadasz, I.; Herold, S.; Mayer, K.; Seeger, W.; Brinckmann, J.; Morty, R.E. Collagen and elastin cross-linking is altered during aberrant late lung development associated with hyperoxia. Am. J. Physiol. Lung Cell. Mol. Physiol. 2015, 308, L1145–L1158. [Google Scholar] [CrossRef] [PubMed]
- Pena, A.M.; Fabre, A.; Debarre, D.; Marchal-Somme, J.; Crestani, B.; Martin, J.L.; Beaurepaire, E.; Schanne-Klein, M.C. Three-dimensional investigation and scoring of extracellular matrix remodeling during lung fibrosis using multiphoton microscopy. Microsc. Res. Tech. 2007, 70, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Raub, C.B.; Mahon, S.; Narula, N.; Tromberg, B.J.; Brenner, M.; George, S.C. Linking optics and mechanics in an in vivo model of airway fibrosis and epithelial injury. J. Biomed. Opt. 2010, 15, 015004. [Google Scholar] [CrossRef] [PubMed]
- Abraham, T.; Hirota, J.A.; Wadsworth, S.; Knight, D.A. Minimally invasive multiphoton and harmonic generation imaging of extracellular matrix structures in lung airway and related diseases. Pulm. Pharmacol. Ther. 2011, 24, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Abraham, T.; Hogg, J. Extracellular matrix remodeling of lung alveolar walls in three dimensional space identified using second harmonic generation and multiphoton excitation fluorescence. J. Struct. Biol. 2010, 171, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Hackett, T.L.; Ferrante, S.C.; Hoptay, C.E.; Engelhardt, J.F.; Ingram, J.L.; Zhang, Y.; Alcala, S.E.; Shaheen, F.; Matz, E.; Pillai, D.K.; et al. A Heterotopic Xenograft Model of Human Airways for Investigating Fibrosis in Asthma. Am. J. Respir. Cell Mol. Biol. 2017, 56, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.D.; McDonough, J.E.; Zeskind, J.E.; Hackett, T.L.; Pechkovsky, D.V.; Brandsma, C.A.; Suzuki, M.; Gosselink, J.V.; Liu, G.; Alekseyev, Y.O.; et al. A gene expression signature of emphysema-related lung destruction and its reversal by the tripeptide GHK. Genome Med. 2012, 4, 67. [Google Scholar] [CrossRef] [PubMed]
- Mostaço-Guidolin, L.B.; Osei, E.T.; Hajimohammadi, S.; Ullah, J.; Hackett, T.-L. Novel non-linear optical imaging to understand the composition of firbrilar collagen and elastin in remodeled asthmatic airways. Am. J. Respir. Crit. Care Med. 2016, 193, A6173. [Google Scholar]
- Khan, M.G. Encyclopedia of Heart Disease; Academic Press: Cambridge, MA, USA, 2005. [Google Scholar]
- Furchgott, R.F. Role of endothelium in responses of vascular smooth muscle. Circ. Res. 1983, 53, 557–573. [Google Scholar] [CrossRef] [PubMed]
- Ross, R. Atherosclerosis is an inflammatory disease. Am. Heart J. 1999, 138, S419–S420. [Google Scholar] [CrossRef]
- Chen, X.; Huang, Z.; Xi, G.; Chen, Y.; Lin, D.; Wang, J.; Li, Z.; Sun, L.; Chen, J.; Chen, R. Quantitative analysis of collagen change between normal and cancerous thyroid tissues based on SHG method. Proc. SPIE 2011. [Google Scholar] [CrossRef]
- Han, X.; Burke, R.M.; Zettel, M.L.; Tang, P.; Brown, E.B. Second harmonic properties of tumor collagen: Determining the structural relationship between reactive stroma and healthy stroma. Opt. Express 2008, 16, 1846–1859. [Google Scholar] [CrossRef] [PubMed]
- Boulesteix, T.; Pena, A.M.; Pages, N.; Godeau, G.; Sauviat, M.P.; Beaurepaire, E.; Schanne-Klein, M.C. Micrometer scale ex vivo multiphoton imaging of unstained arterial wall structure. Cytometry A 2006, 69, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Van Zandvoort, M.; Engels, W.; Douma, K.; Beckers, L.; Oude Egbrink, M.; Daemen, M.; Slaaf, D.W. Two-photon microscopy for imaging of the (atherosclerotic) vascular wall: A proof of concept study. J. Vasc. Res. 2004, 41, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Fratzl, P. Collagen: Structure and Mechanics; Springer Science & Business Media: Berlin, Germany, 2008. [Google Scholar]
- Ottani, V.; Raspanti, M.; Ruggeri, A. Collagen structure and functional implications. Micron 2001, 32, 251–260. [Google Scholar] [CrossRef]
- Heybeli, N.K.B.; Yilmaz, B.; Guler, O. Musculoskeletal Research and Basic Science—“Tendons and ligaments”; Springer: Berlin, Germany, 2016; Volume 3. [Google Scholar]
- Kannus, P. Structure of the tendon connective tissue. Scand. J. Med. Sci. Sports 2000, 10, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Keene, D.R.; Sakai, L.Y.; Bachinger, H.P.; Burgeson, R.E. Type III collagen can be present on banded collagen fibrils regardless of fibril diameter. J. Cell Biol. 1987, 105, 2393–2402. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, T.; Jia, F.; Niyibizi, C.; Woo, S.L. Antisense oligonucleotides reduce synthesis of procollagen α1 (V) chain in human patellar tendon fibroblasts: Potential application in healing ligaments and tendons. Connect. Tissue Res. 2003, 44, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Sivaguru, M.D.S.; Ambekar, R.; Luedtke, D.; Fried, G.; Stewart, A.; Toussaint, K.C. Quantitative analysis of diseased horse tendons using Fourier-tranform-second-harmonic generation imaging. Proc. SPIE 2011. [Google Scholar] [CrossRef]
- Malik, R.A.; Kallinikos, P.; Abbott, C.A.; van Schie, C.H.; Morgan, P.; Efron, N.; Boulton, A.J. Corneal confocal microscopy: A non-invasive surrogate of nerve fibre damage and repair in diabetic patients. Diabetologia 2003, 46, 683–688. [Google Scholar] [CrossRef] [PubMed]
- Raub, C.B.; Putnam, A.J.; Tromberg, B.J.; George, S.C. Predicting bulk mechanical properties of cellularized collagen gels using multiphoton microscopy. Acta Biomater. 2010, 6, 4657–4665. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, N.D.; Hoying, J.B.; Botting, S.K.; Weiss, J.A.; Utzinger, U. In vitro model for endogenous optical signatures of collagen. J. Biomed. Opt. 2006, 11, 054021. [Google Scholar] [CrossRef] [PubMed]
- Turcotte, R.; Mattson, J.M.; Wu, J.W.; Zhang, Y.; Lin, C.P. Molecular Order of Arterial Collagen Using Circular Polarization Second-Harmonic Generation Imaging. Biophys. J. 2016, 110, 530–533. [Google Scholar] [CrossRef] [PubMed]
- Rao, R.A.; Mehta, M.R.; Leithem, S.; Toussaint, K.C., Jr. Quantitative analysis of forward and backward second-harmonic images of collagen fibres using Fourier transform second-harmonic-generation microscopy. Opt. Lett. 2009, 34, 3779–3781. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.M.; Zipfel, W.R.; Webb, W.W. Interpreting second-harmonic generation images of collagen I fibrils. Biophys. J. 2005, 88, 1377–1386. [Google Scholar] [CrossRef] [PubMed]
- Indahl, U.; Næs, T. Evaluation of alternative spectral feature extraction methods of textural images for multivariate modeling. J. Chemom. 1998, 12, 261–278. [Google Scholar] [CrossRef]
- Chang, T.; Kuo, C.J. Texture analysis and classification with tree-structured wavelet transform. IEEE Trans Image Process 1993, 2, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Tuceryan, M.J.; Jain, A.K. Texture Analysis. In Handbook of Pattern Recognition and Comuter Vision; Chen, C.H., Ed.; Word Scientific: Singapore, 1993; Volume 2, pp. 235–276. [Google Scholar]
- Unser, M. Texture classification and segmentation using wavelet frames. IEEE Trans. Image Process. 1995, 4, 1549–1560. [Google Scholar] [CrossRef] [PubMed]
- Arivazhagan, S.G.L. Texture classfication using wavelet transform. Pattern Recognit. Lett. 2003, 24, 1513–1521. [Google Scholar] [CrossRef]
- Mostaço-Guidolin, L.B.; Ko, A.C.; Popescu, D.P.; Smith, M.S.; Kohlenberg, E.K.; Shiomi, M.; Major, A.; Sowa, M.G. Evaluation of texture parameters for the quantitative description of multimodal nonlinear optical images from atherosclerotic rabbit arteries. Phys. Med. Biol. 2011, 56, 5319–5334. [Google Scholar] [CrossRef] [PubMed]
- Bharati, M.H.; Liu, J.J.; MacGregor, J.F. Image texture analysis: Methods and comparisons. Chemom. Intell. Lab. Syst. 2004, 72, 57–71. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organ/Tissue | Key Conclusions | Representative Reference |
|---|---|---|
| Skin |
| [23,30,84,85,86] |
| Tendon and Ligaments |
| [75,87,88,89] |
| Cardiovascular |
| [28,40,90,91,92,93] |
| Lung |
| [94,95,96] |
| Eye |
| [24,97] |
| In-vitro models |
| [34,98,99,100] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mostaço-Guidolin, L.; Rosin, N.L.; Hackett, T.-L. Imaging Collagen in Scar Tissue: Developments in Second Harmonic Generation Microscopy for Biomedical Applications. Int. J. Mol. Sci. 2017, 18, 1772. https://doi.org/10.3390/ijms18081772
Mostaço-Guidolin L, Rosin NL, Hackett T-L. Imaging Collagen in Scar Tissue: Developments in Second Harmonic Generation Microscopy for Biomedical Applications. International Journal of Molecular Sciences. 2017; 18(8):1772. https://doi.org/10.3390/ijms18081772
Chicago/Turabian StyleMostaço-Guidolin, Leila, Nicole L. Rosin, and Tillie-Louise Hackett. 2017. "Imaging Collagen in Scar Tissue: Developments in Second Harmonic Generation Microscopy for Biomedical Applications" International Journal of Molecular Sciences 18, no. 8: 1772. https://doi.org/10.3390/ijms18081772
APA StyleMostaço-Guidolin, L., Rosin, N. L., & Hackett, T.-L. (2017). Imaging Collagen in Scar Tissue: Developments in Second Harmonic Generation Microscopy for Biomedical Applications. International Journal of Molecular Sciences, 18(8), 1772. https://doi.org/10.3390/ijms18081772