In Situ Forming Gelatin Hydrogels-Directed Angiogenic Differentiation and Activity of Patient-Derived Human Mesenchymal Stem Cells

 ,
, 
Abstract
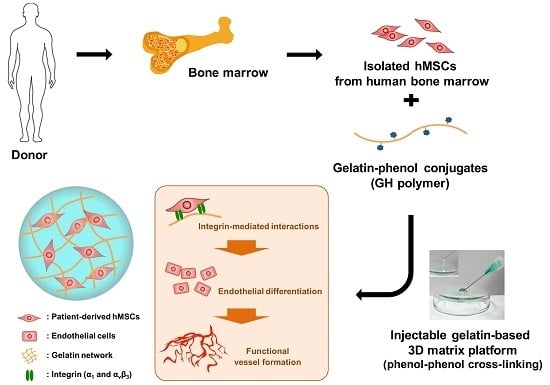
:
1. Introduction
2. Results and Discussion
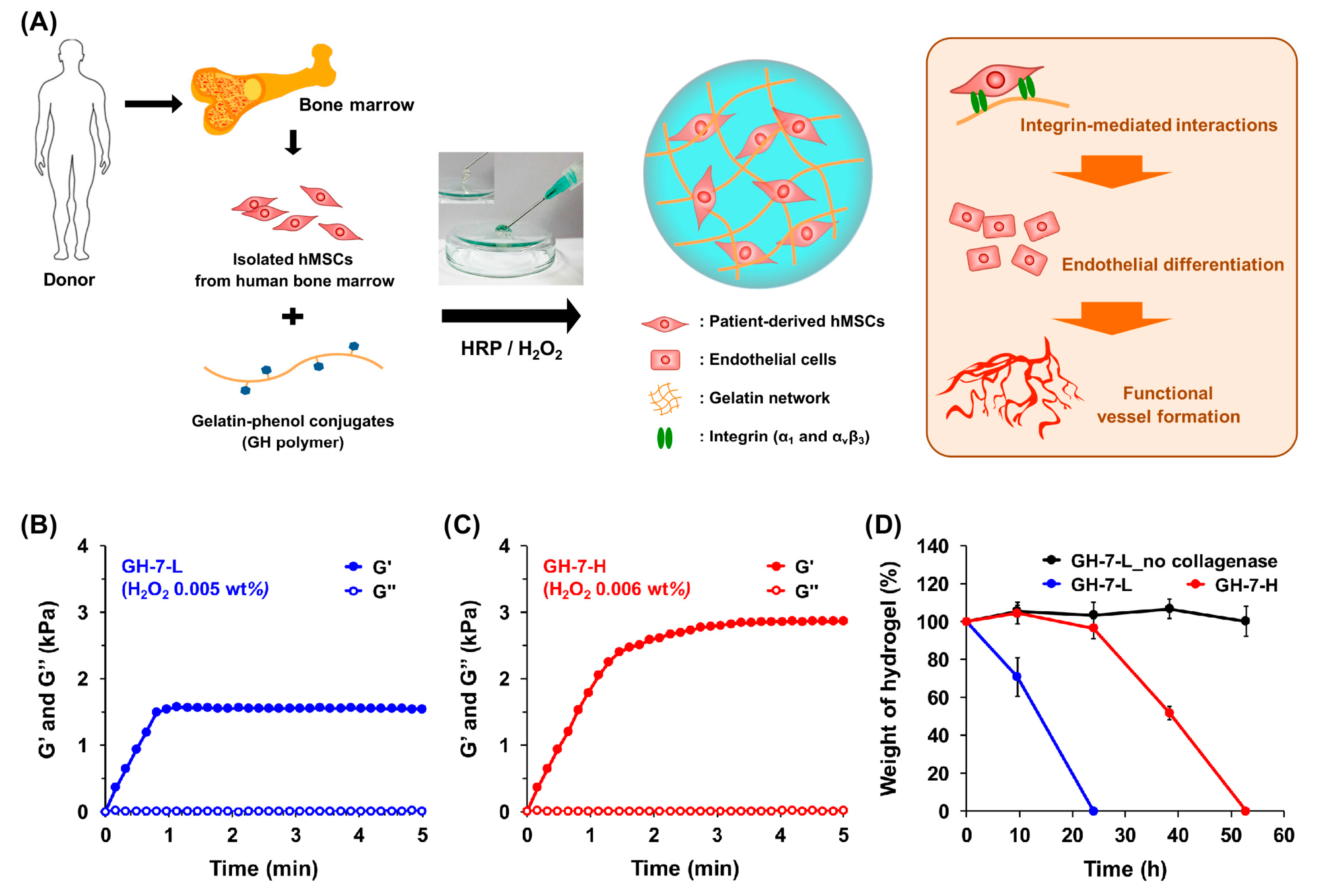
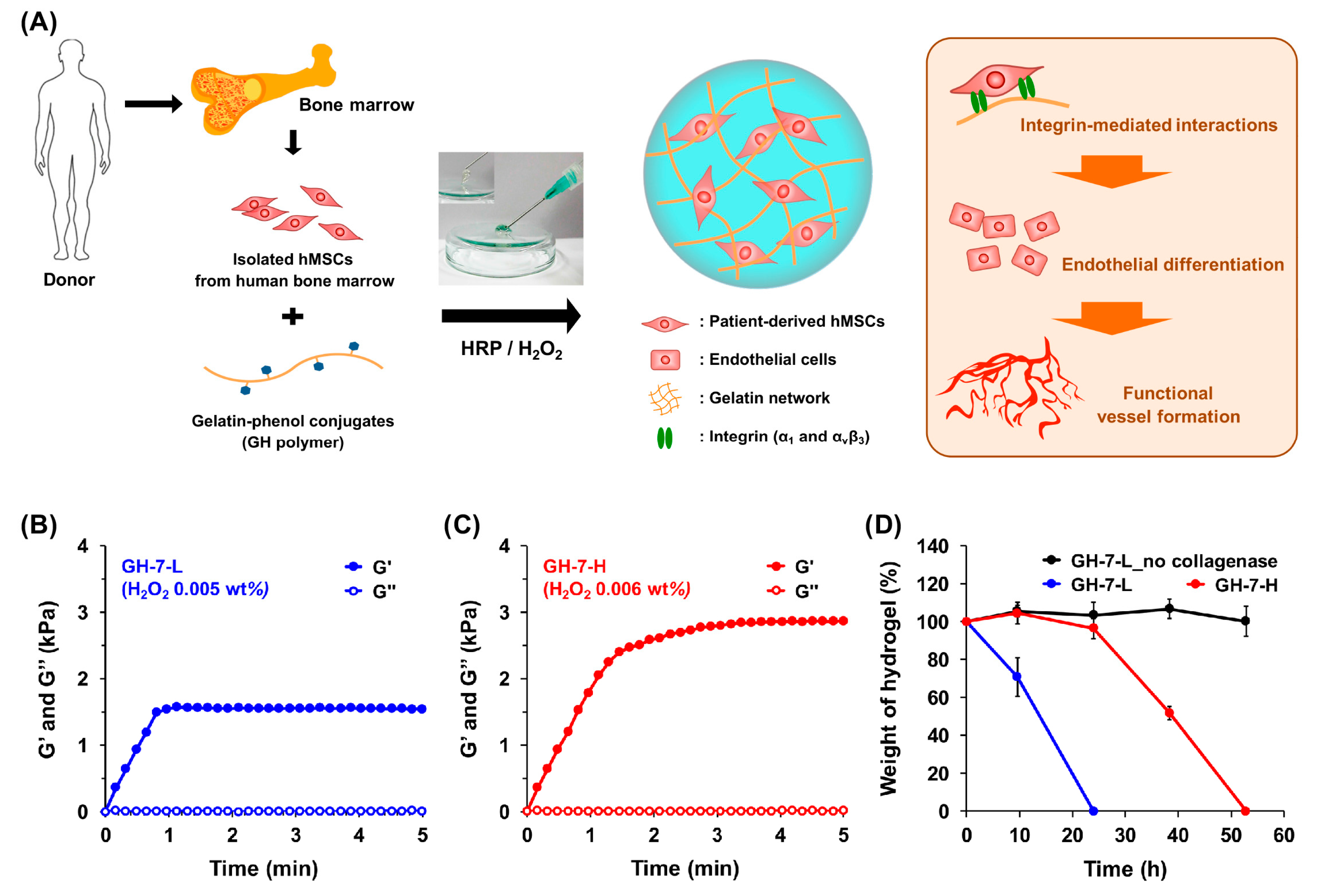
2.1. Characterization of In Situ Forming Gelatin−Hydroxyphenyl Propionic Acid (GH) Hydrogels
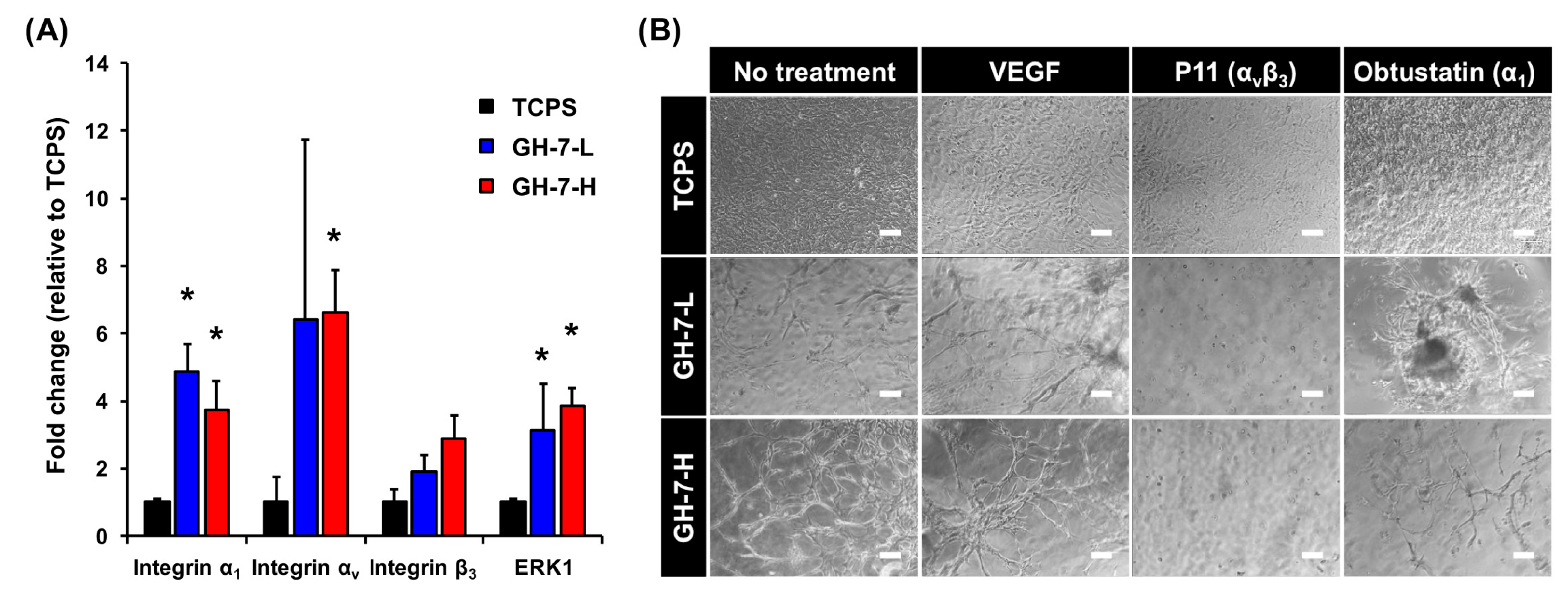
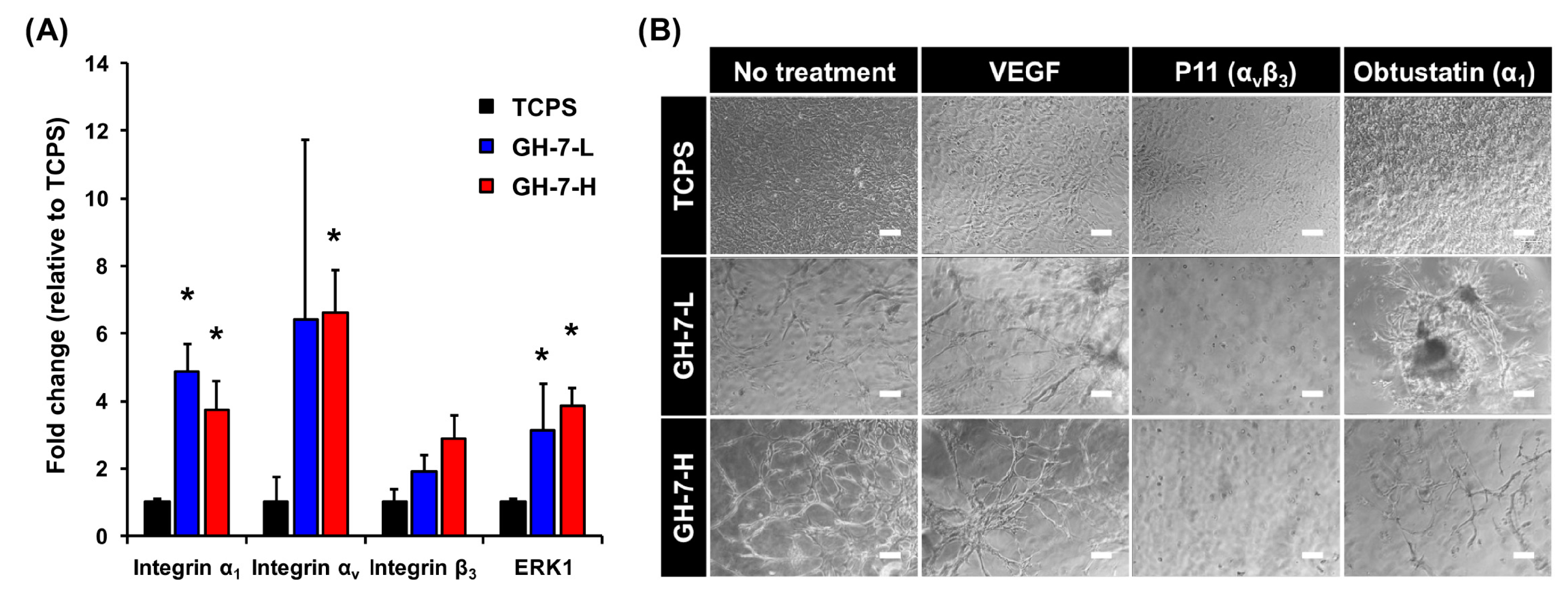
2.2. Gelatin Hydrogel Material-Driven Differentiation of MSCs (Mesenchymal Stem Cells) into Endothelial Cells
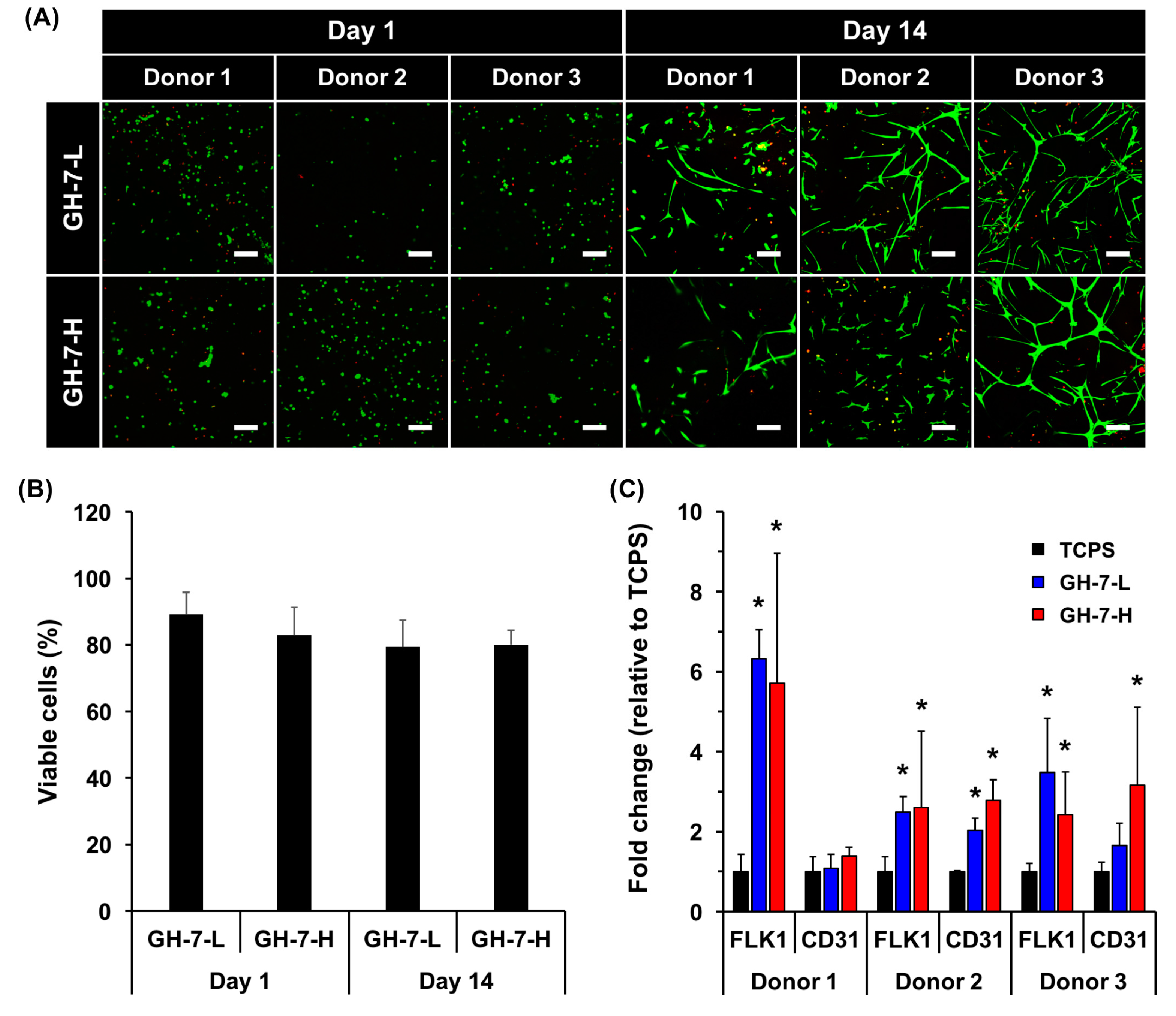
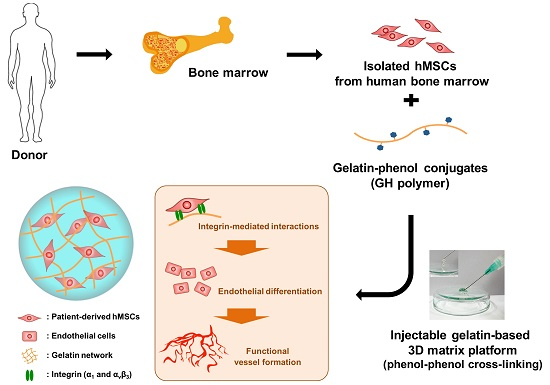
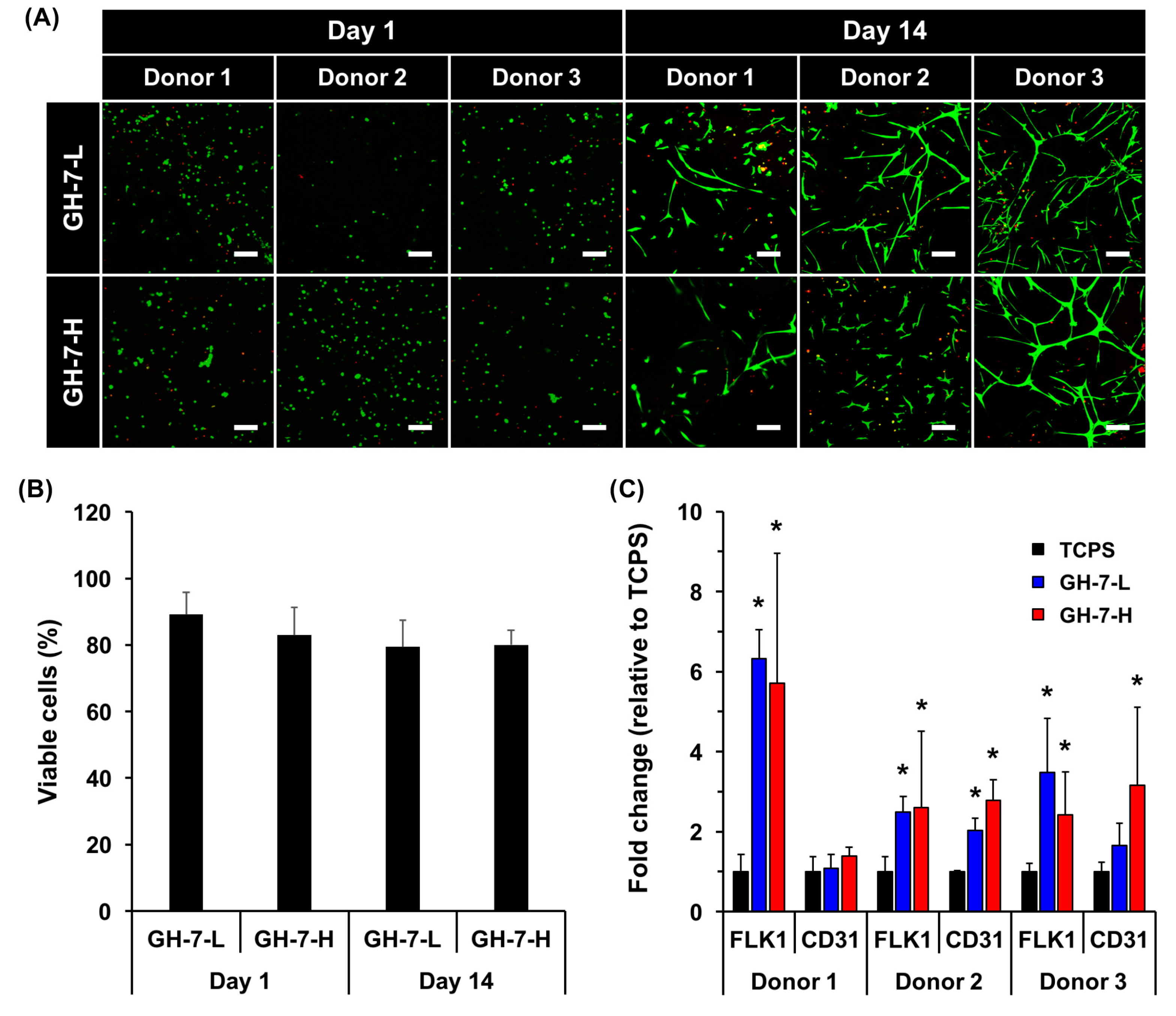
2.3. In Vitro Endothelial Differentiation of Patient-Derived MSCs in GH Hydrogels
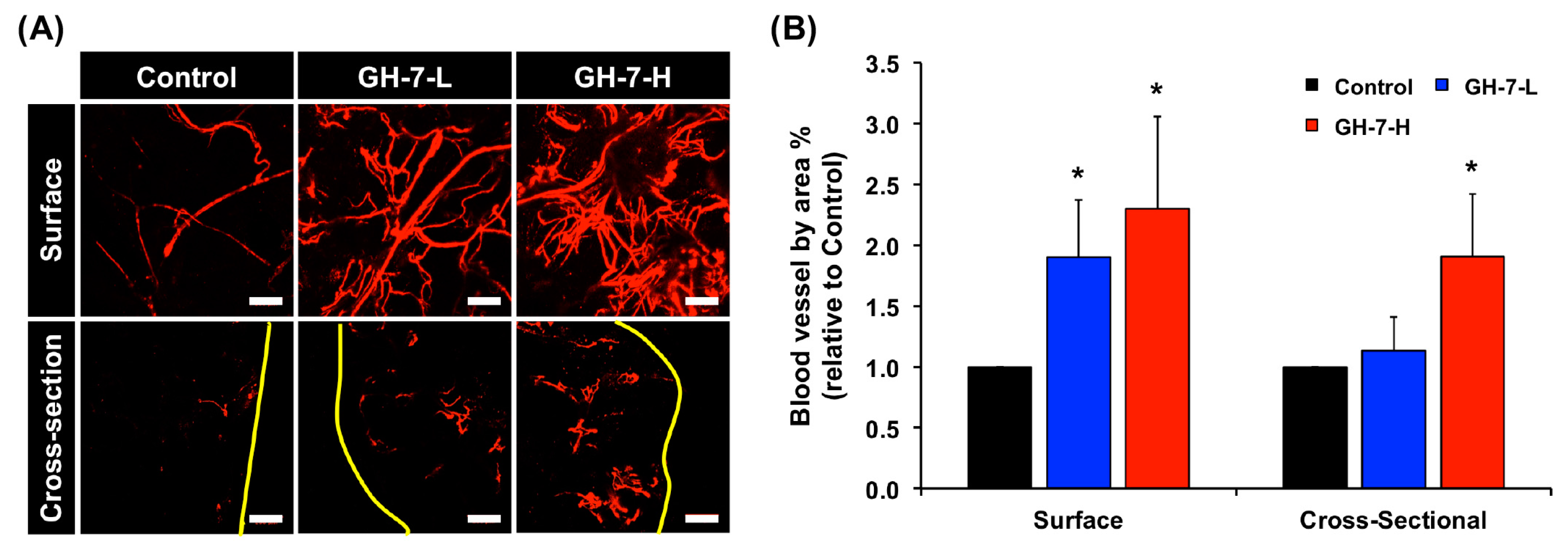
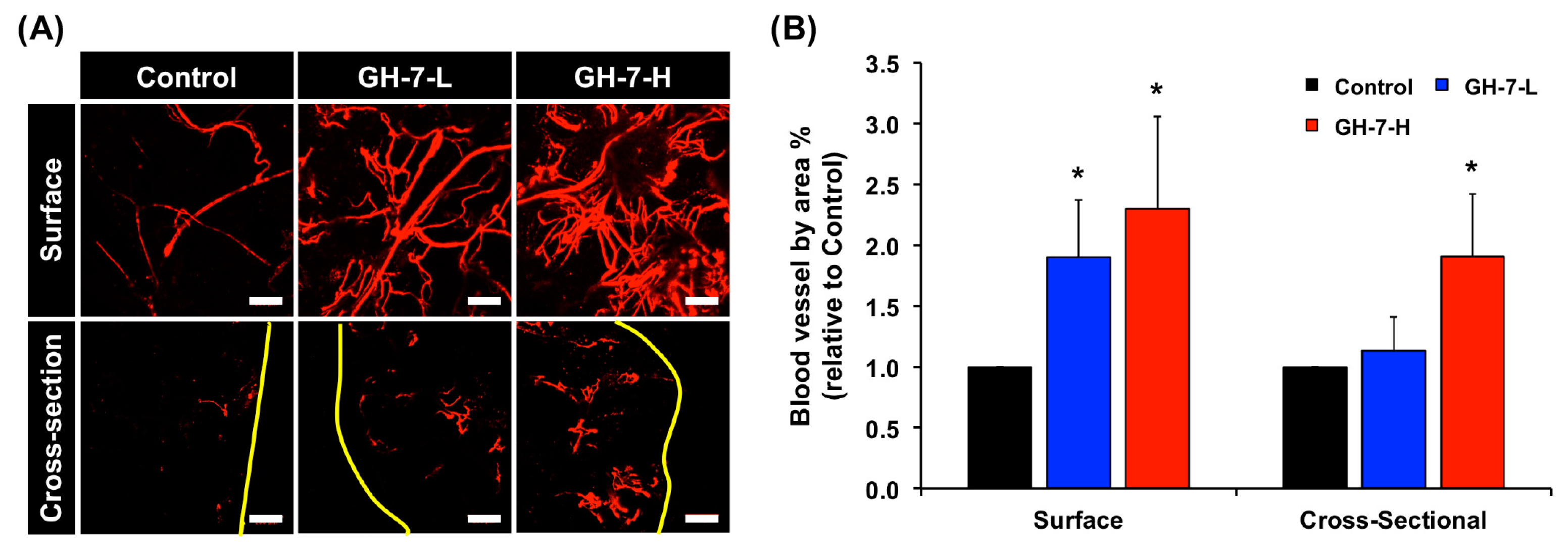
2.4. In Vivo Vascularization of hMSCs Delivered with GH Hydrogel
3. Materials and Methods
3.1. Materials
3.2. Preparation and Characterization of GH Hydrogels with Different Mechanical Stiffness
3.3. In Vitro 3D Culture of hMSC in GH Hydrogels
3.4. hMSC Delivery in GH Hydrogels on Polyvinyl Alcohol (PVA) Scaffolds In Vivo
3.5. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Han, H.; Ning, H.; Liu, S.; Lu, Q.; Fan, Z.; Lu, H.; Lu, G.; Kaplan, D.L. Silk Biomaterials with Vascularization Capacity. Adv. Funct. Mater. 2016, 26, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Chuang, C.H.; Lin, R.Z.; Tien, H.W.; Chu, Y.C.; Li, Y.C.; Melero-Martin, J.M.; Chen, Y.C. Enzymatic regulation of functional vascular networks using gelatin hydrogels. Acta Biomater. 2015, 19, 85–99. [Google Scholar] [CrossRef] [PubMed]
- Levenberg, S.; Rouwkema, J.; Macdonald, M.; Garfein, E.S.; Kohane, D.S.; Darland, D.C.; Marini, R.; van Blitterswijk, C.A.; Mulligan, R.C.; D’Amore, P.A.; Langer, R. Engineering vascularized skeletal muscle tissue. Nat. Biotechnol. 2005, 23, 879–884. [Google Scholar] [CrossRef] [PubMed]
- Griffith, L.G.; Naughton, G. Tissue engineering—current challenges and expanding opportunities. Science 2002, 295, 1009–1014. [Google Scholar] [CrossRef] [PubMed]
- Lovett, M.; Lee, K.; Edwards, A.; Kaplan, D.L. Vascularization strategies for tissue engineering. Tissue Eng. Part B 2009, 15, 353–370. [Google Scholar] [CrossRef] [PubMed]
- Linh, N.T.; Abueva, C.D.; Lee, B.T. Enzymatic in situ formed hydrogel from gelatin-tyramine and chitosan-4-hydroxylphenyl acetamide for the co-delivery of human adipose-derived stem cells and platelet-derived growth factor towards vascularization. Biomed. Mater. 2017, 12, 015026. [Google Scholar] [CrossRef] [PubMed]
- Eke, G.; Mangir, N.; Hasirci, N.; MacNeil, S.; Hasirci, V. Development of a UV crosslinked biodegradable hydrogel containing adipose derived stem cells to promote vascularization for skin wounds and tissue engineering. Biomaterials 2017, 129, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Martino, M.M.; Briquez, P.S.; Guc, E.; Tortelli, F.; Kilarski, W.W.; Metzger, S.; Rice, J.J.; Kuhn, G.A.; Muller, R.; Swartz, M.A.; et al. Growth factors engineered for super-affinity to the extracellular matrix enhance tissue healing. Science 2014, 343, 885–888. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Ogasawara, A.K.; Yang, R.; Wei, W.; He, G.W.; Zioncheck, T.F.; Bunting, S.; de Vos, A.M.; Jin, H. KDR (VEGF Receptor 2) Is the Major Mediator for the Hypotensive Effect of VEGF. Hypertension 2002, 39, 1095–1100. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.; Komada, M.R.; Sane, D.C. Abnormal angiogenesis in diabetes mellitus. Med. Res. Rev. 2003, 23, 117–145. [Google Scholar] [CrossRef] [PubMed]
- Petreaca, M.; Martins-Green, M. The Dynamics of Cell-ECM Interactions, with Implications for Tissue Engineering. Princ. of Tissue Eng. 2014, 161–187. [Google Scholar]
- Eming, S.A.; Hubbell, J.A. Extracellular matrix in angiogenesis: Dynamic structures with translational potential. Exp. Dermatol. 2011, 20, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Rico, P.; Mnatsakanyan, H.; Dalby, M.J.; Salmerón-Sánchez, M. Material-Driven Fibronectin Assembly Promotes Maintenance of Mesenchymal Stem Cell Phenotypes. Adv. Funct. Mater. 2016, 26, 6563–6573. [Google Scholar] [CrossRef]
- Sun, H.; Zhu, F.; Hu, Q.; Krebsbach, P.H. Controlling stem cell-mediated bone regeneration through tailored mechanical properties of collagen scaffolds. Biomaterials 2014, 35, 1176–1184. [Google Scholar] [CrossRef] [PubMed]
- Park, K.M.; Gerecht, S. Hypoxia-inducible hydrogels. Nat. Commun. 2014, 5, 4075. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Wiraja, C.; Muhammad, H.B.; Xu, C.; Wang, D.A. Improvement of endothelial progenitor outgrowth cell (EPOC)-mediated vascularization in gelatin-based hydrogels through pore size manipulation. Acta Biomater. 2017, 58, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Dalby, M.J.; Gadegaard, N.; Oreffo, R.O.C. Harnessing nanotopography and integrin–matrix interactions to influence stem cell fate. Nat. Mater. 2014, 13, 558–569. [Google Scholar] [CrossRef] [PubMed]
- Ko, D.Y.; Shinde, U.P.; Yeon, B.; Jeong, B. Recent progress of in situ formed gels for biomedical applications. Prog. Polym. Sci. 2013, 38, 672–701. [Google Scholar] [CrossRef]
- Sivashanmugam, A.; Arun Kumar, R.; Vishnu Priya, M.; Nair, S.V.; Jayakumar, R. An overview of injectable polymeric hydrogels for tissue engineering. Eur. Polym. J. 2015, 72, 543–565. [Google Scholar] [CrossRef]
- Lee, Y.; Bae, J.W.; Lee, J.W.; Suh, W.; Park, K.D. Enzyme-catalyzed in situ forming gelatin hydrogels as bioactive wound dressings: Effects of fibroblast delivery on wound healing efficacy. J. Mater. Chem. B 2014, 2, 7712–7718. [Google Scholar] [CrossRef]
- Lee, Y.; Bae, J.W.; Oh, D.H.; Park, K.M.; Chun, Y.W.; Sung, H.-J.; Park, K.D. In situ forming gelatin-based tissue adhesives and their phenolic content-driven properties. J. Mater. Chem. B 2013, 1, 2407–2414. [Google Scholar] [CrossRef]
- Knopf-Marques, H.; Barthes, J.; Wolfova, L.; Vidal, B.; Koenig, G.; Bacharouche, J.; Francius, G.; Sadam, H.; Liivas, U.; Lavalle, P.; Vrana, N.E. Auxiliary Biomembranes as a Directional Delivery System To Control Biological Events in Cell-Laden Tissue-Engineering Scaffolds. ACS Omega 2017, 2, 918–929. [Google Scholar] [CrossRef]
- Lee, S.H.; Lee, Y.; Chun, Y.W.; Crowder, S.W.; Young, P.P.; Park, K.D.; Sung, H.J. In Situ Crosslinkable Gelatin Hydrogels for Vasculogenic Induction and Delivery of Mesenchymal Stem Cells. Adv. Funct. Mater. 2014, 24, 6771–6781. [Google Scholar] [CrossRef] [PubMed]
- Waterston, R.H.; Lindblad-Toh, K.; Birney, E.; Rogers, J.; Abril, J.F.; Agarwal, P.; Agarwala, R.; Ainscough, R.; Alexandersson, M.; An, P.; et al. Initial sequencing and comparative analysis of the mouse genome. Nature 2002, 420, 520–562. [Google Scholar] [PubMed]
- Choi, B.; Loh, X.J.; Tan, A.; Loh, C.K.; Ye, E.; Joo, M.K.; Jeong, B. Introduction to In Situ Forming Hydrogels for Biomedical Applications. In In-Situ Gelling Polymers; Springer: Berlin, Germany, 2015; pp. 5–35. [Google Scholar]
- Trappmann, B.; Gautrot, J.E.; Connelly, J.T.; Strange, D.G.; Li, Y.; Oyen, M.L.; Cohen Stuart, M.A.; Boehm, H.; Li, B.; Vogel, V.; et al. Extracellular-matrix tethering regulates stem-cell fate. Nat. Mater. 2012, 11, 642–649. [Google Scholar] [CrossRef] [PubMed]
- Engler, A.J.; Sen, S.; Sweeney, H.L.; Discher, D.E. Matrix elasticity directs stem cell lineage specification. Cell 2006, 126, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Oswald, J.; Boxberger, S.; Jørgensen, B.; Feldmann, S.; Ehninger, G.; Bornhäuser, M.; Werner, C. Mesenchymal Stem Cells Can Be Differentiated Into Endothelial Cells In Vitro. Stem Cells 2004, 22, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Kubota, Y.; Kleinman, H.K.; Martin, G.R.; Lawley, T.J. Role of laminin and basement membrane in the morphological differentiation of human endothelial cells into capillary-like structures. J. Cell. Biol. 1988, 107, 1589–1598. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.H.; Wang, T.M.; Young, T.H.; Lai, Y.K.; Yen, M.L. The critical role of ECM proteins within the human MSC niche in endothelial differentiation. Biomaterials 2013, 34, 4223–4234. [Google Scholar] [CrossRef] [PubMed]
- Nichol, J.W.; Koshy, S.T.; Bae, H.; Hwang, C.M.; Yamanlar, S.; Khademhosseini, A. Cell-laden microengineered gelatin methacrylate hydrogels. Biomaterials 2010, 31, 5536–5544. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.-Y.; Kim, J.-T.; Lee, W.-J.; Lee, Y.; Park, K.M.; Yang, Y.-I.; Park, K.D. Engineered extracellular microenvironment with a tunable mechanical property for controlling cell behavior and cardiomyogenic fate of cardiac stem cells. Acta Biomater. 2017, 50, 234–248. [Google Scholar] [CrossRef] [PubMed]
- Marcinkiewicz, C.; Weinreb, P.H.; Calvete, J.J.; Kisiel, D.G.; Mousa, S.A.; Tuszynski, G.P.; Lobb, R.R. Obtustatin: A potent selective inhibitor of alpha1beta1 integrin in vitro and angiogenesis in vivo. Cancer Res. 2003, 63, 2020–2023. [Google Scholar] [PubMed]
- Choi, Y.; Kim, E.; Lee, Y.; Han, M.H.; Kang, I.C. Site-specific inhibition of integrin alpha v beta 3-vitronectin association by a ser-asp-val sequence through an Arg-Gly-Asp-binding site of the integrin. Proteomics 2010, 10, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Zachman, A.L.; Crowder, S.W.; Ortiz, O.; Zienkiewicz, K.J.; Bronikowski, C.M.; Yu, S.S.; Giorgio, T.D.; Guelcher, S.A.; Kohn, J.; Sung, H.J. Pro-angiogenic and anti-inflammatory regulation by functional peptides loaded in polymeric implants for soft tissue regeneration. Tissue Eng. Part A 2013, 19, 437–447. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession Number | Forward Primer (5′–3′) | Reverse Primer (3′–5′) | Species |
|---|---|---|---|---|
| Integrin α1 | NM_001033228.3 | TCAGTGGAGAGCAGATCGGA | CCCACAGGGCTCATTCTTGT | Mouse |
| Integrin αv | NM_008402.3 | GTGCCAGCCCATTGAGTTTG | TGGAGCACAGGCCAAGATTT | Mouse |
| Integrin β3 | NM_016780.2 | GCCTGGTGCTCAGATGAGACT | GATCTTCGAATCATCTGGCCG | Mouse |
| ERK1 | NM_011952.2 | CAACCCAAACAAGCGCATCA | AGGAGCAGGACCAGATCCAA | Mouse |
| GAPDH | NM_001289726 | TGAAGCAGGCATCTGAGGG | CGAAGGTGGAAGAGTGGGAG | Mouse |
| FLK1 | NM_002253.2 | GAGGGGAACTGAAGACAGGC | GGCCAAGAGGCTTACCTAGC | Human |
| CD31 | NM_000442.4 | CCAAGCCCGAACTGGAATCT | CACTGTCCGACTTTGAGGCT | Human |
| GAPDH | NM_002046.4 | GCACCGTCAAGGCTGAGAAC | TGGTGAAGACGCCAGTGGA | Human |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.; Balikov, D.A.; Lee, J.B.; Lee, S.H.; Lee, S.H.; Lee, J.H.; Park, K.D.; Sung, H.-J. In Situ Forming Gelatin Hydrogels-Directed Angiogenic Differentiation and Activity of Patient-Derived Human Mesenchymal Stem Cells. Int. J. Mol. Sci. 2017, 18, 1705. https://doi.org/10.3390/ijms18081705
Lee Y, Balikov DA, Lee JB, Lee SH, Lee SH, Lee JH, Park KD, Sung H-J. In Situ Forming Gelatin Hydrogels-Directed Angiogenic Differentiation and Activity of Patient-Derived Human Mesenchymal Stem Cells. International Journal of Molecular Sciences. 2017; 18(8):1705. https://doi.org/10.3390/ijms18081705
Chicago/Turabian StyleLee, Yunki, Daniel A. Balikov, Jung Bok Lee, Sue Hyun Lee, Seung Hwan Lee, Jong Hun Lee, Ki Dong Park, and Hak-Joon Sung. 2017. "In Situ Forming Gelatin Hydrogels-Directed Angiogenic Differentiation and Activity of Patient-Derived Human Mesenchymal Stem Cells" International Journal of Molecular Sciences 18, no. 8: 1705. https://doi.org/10.3390/ijms18081705
APA StyleLee, Y., Balikov, D. A., Lee, J. B., Lee, S. H., Lee, S. H., Lee, J. H., Park, K. D., & Sung, H.-J. (2017). In Situ Forming Gelatin Hydrogels-Directed Angiogenic Differentiation and Activity of Patient-Derived Human Mesenchymal Stem Cells. International Journal of Molecular Sciences, 18(8), 1705. https://doi.org/10.3390/ijms18081705