Sympathetic Overactivity in Chronic Kidney Disease: Consequences and Mechanisms

Abstract
: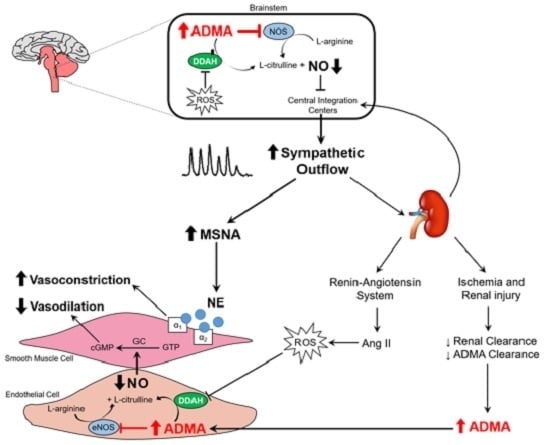
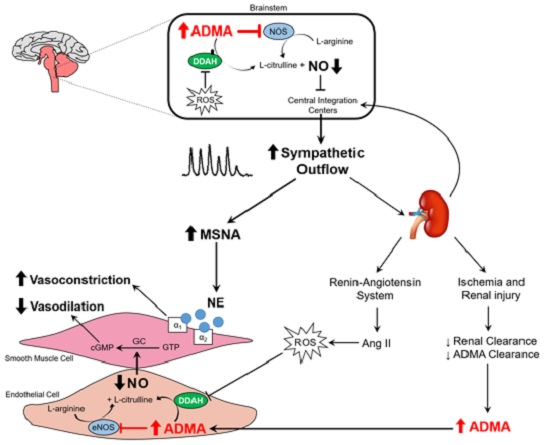{kind=link}
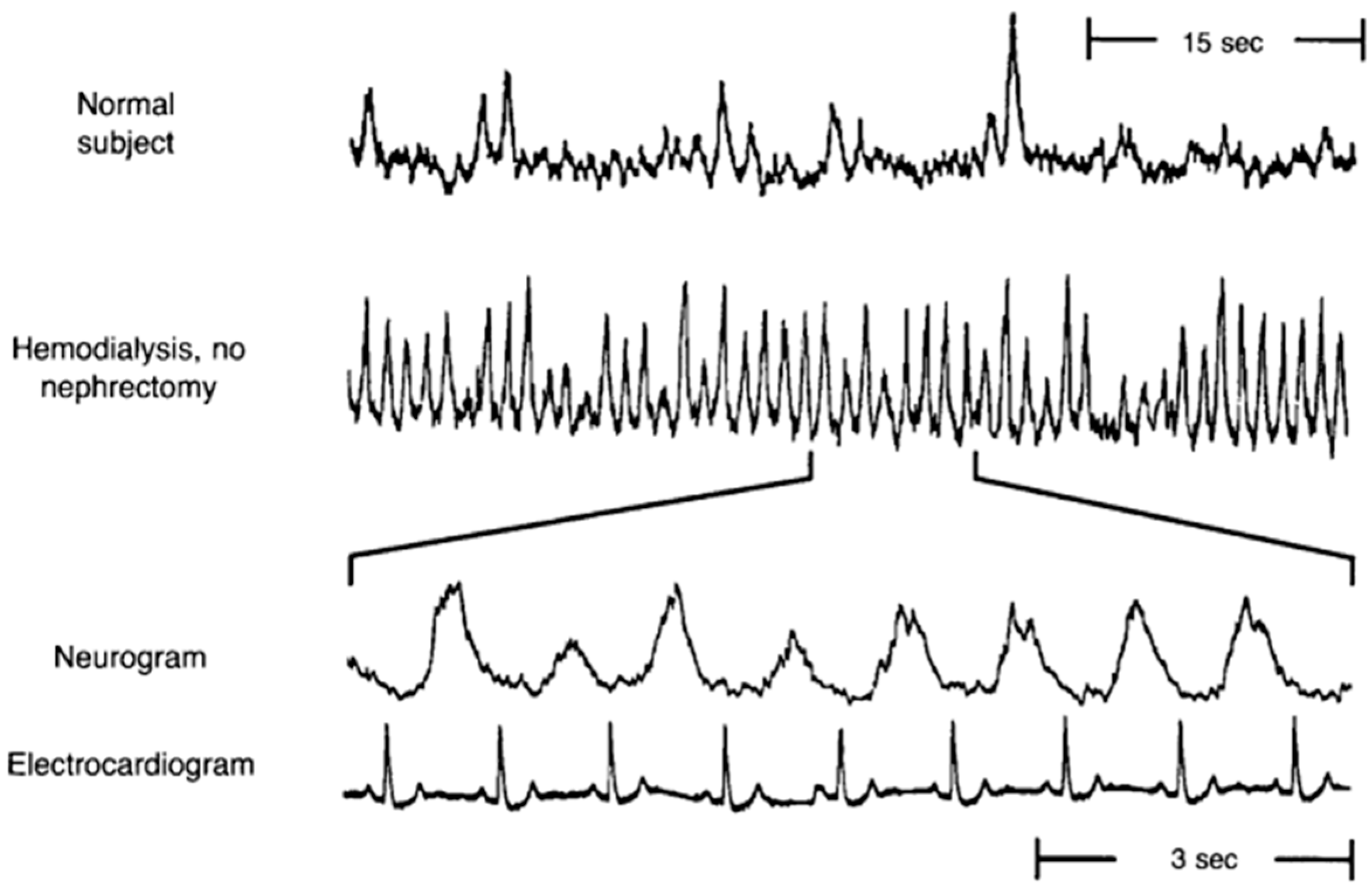{kind=link}
{kind=link}
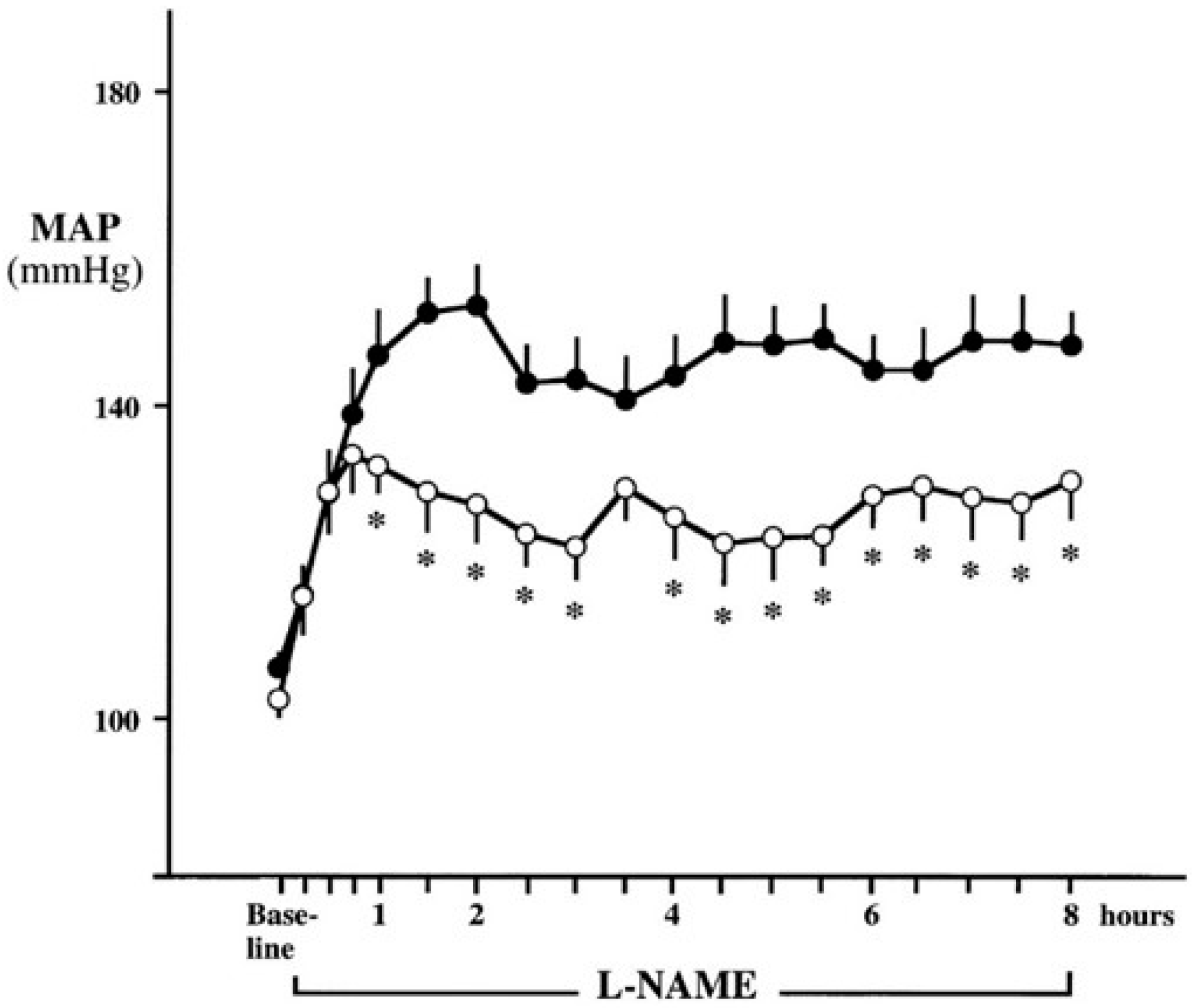{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Consequences of Sympathetic Overactivity
3. Mechanisms of Sympathetic Overactivity in Chronic Kidney Disease (CKD)
3.1. Renin–Angiotensin System
3.2. Renal Afferents
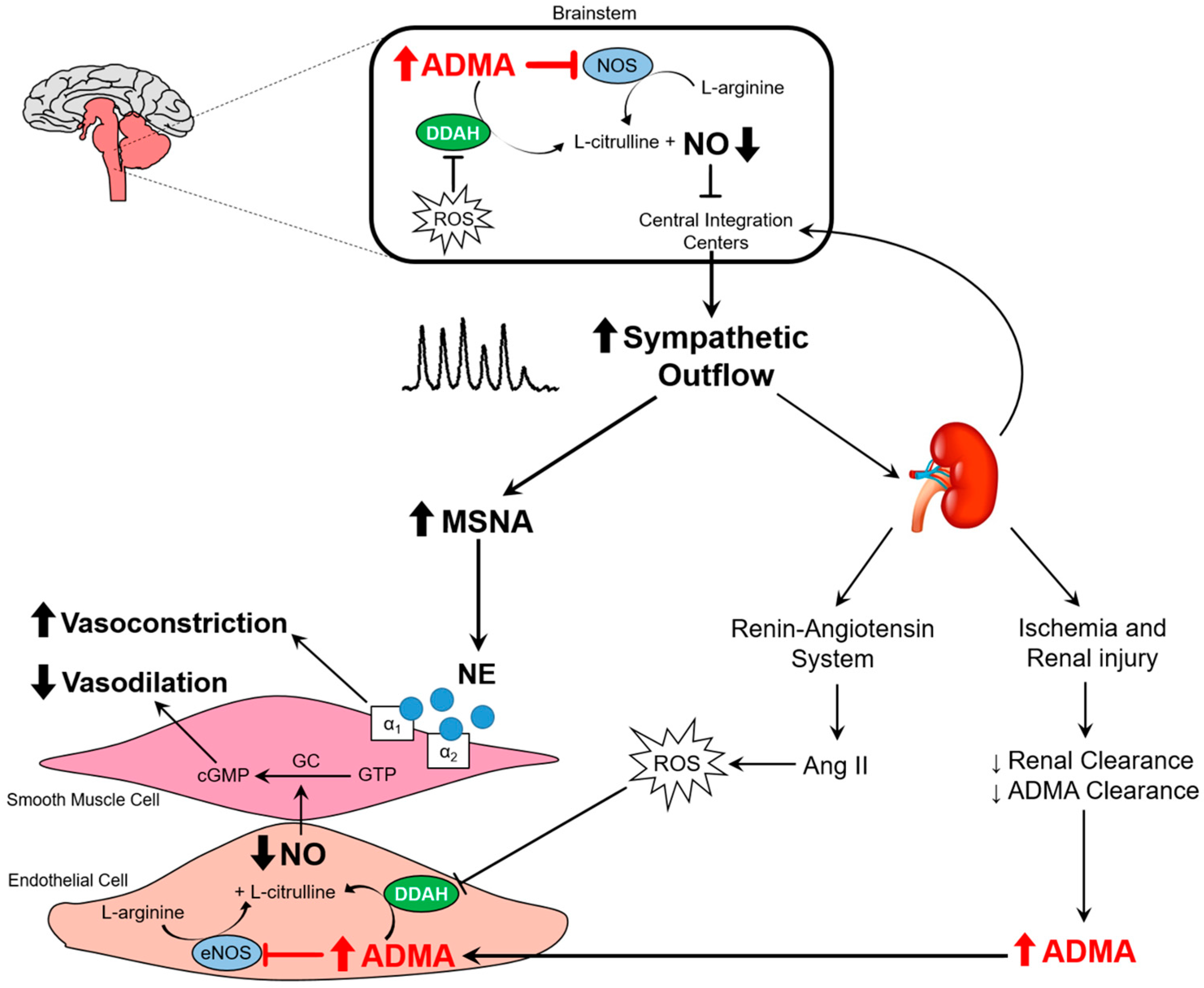
3.3. Nitric Oxide Pathway
3.4. Oxidative Stress
4. Therapeutic Strategies
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ACE | Angiotensin converting enzyme |
| ADMA | Asymmetric dimethylarginine |
| bs | Bursts |
| cGMP | Cellular guanosine monophosphate |
| CKD | Chronic kidney disease |
| CV | Cardiovascular |
| DDAH | Dimethylarginine dimethylaminohydrolase |
| eGFR | Estimated glomerular filtration rate |
| eNOS | Endothelial nitric oxide synthase |
| hb | Heartbeats |
| iNOS | Inducible nitric oxide synthase |
| l-NAME | l-NG-nitroarginine methyl ester |
| l-NMMA | NG-monomethyl l-arginine |
| MSNA | Muscle sympathetic nerve activity |
| NAD(P)H | Nicotinamide adenine dinucleotide phosphate |
| NE | Norepinephrine |
| NO | Nitric oxide |
| NOS | Nitric oxide synthase |
| nNOS | Neuronal nitric oxide synthase |
| NTS | Nucleus tractus solitarius |
| PVN | Paraventricular nucleus |
| RVLM | Rostral ventrolateral medulla |
| SNA | Sympathetic nerve activity |
| SOD | Superoxide dismutase |
References
- Coresh, J.; Selvin, E.; Stevens, L.A.; Manzi, J.; Kusek, J.W.; Eggers, P.; Van, L.F.; Levey, A.S. Prevalence of chronic kidney disease in the United States. JAMA 2007, 298, 2038–2047. [Google Scholar] [CrossRef] [PubMed]
- Saran, R.; Li, Y.; Robinson, B.; Abbott, K.C.; Agodoa, L.Y.; Ayanian, J.; Bragg-Gresham, J.; Balkrishnan, R.; Chen, J.L.; Cope, E.; et al. US Renal Data System 2015 Annual Data Report: Epidemiology of kidney disease in the United States. Am. J. Kidney Dis. 2016. [Google Scholar] [CrossRef] [PubMed]
- Honeycutt, A.A.; Segel, J.E.; Zhuo, X.; Hoerger, T.J.; Imai, K.; Williams, D. Medical costs of CKD in the Medicare population. J. Am. Soc. Nephrol. 2013, 24, 1478–1483. [Google Scholar] [CrossRef] [PubMed]
- Go, A.S.; Chertow, G.M.; Fan, D.; McCulloch, C.E.; Hsu, C.Y. Chronic kidney disease and the risks of death, cardiovascular events, and hospitalization. N. Engl. J. Med. 2004, 351, 1296–1305. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, K.; van der Velde, M.; Astor, B.C.; Woodward, M.; Levey, A.S.; de Jong, P.E.; Coresh, J.; Gansevoort, R.T. Association of estimated glomerular filtration rate and albuminuria with all-cause and cardiovascular mortality in general population cohorts: A collaborative meta-analysis. Lancet 2010, 375, 2073–2081. [Google Scholar] [PubMed]
- Herzog, C.A.; Asinger, R.W.; Berger, A.K.; Charytan, D.M.; Diez, J.; Hart, R.G.; Eckardt, K.U.; Kasiske, B.L.; McCullough, P.A.; Passman, R.S.; et al. Cardiovascular disease in chronic kidney disease. A clinical update from Kidney Disease: Improving Global Outcomes (KDIGO). Kidney Int. 2011, 80, 572–586. [Google Scholar] [CrossRef] [PubMed]
- Green, D.; Roberts, P.R.; New, D.I.; Kalra, P.A. Sudden cardiac death in hemodialysis patients: An in-depth review. Am. J. Kidney Dis. 2011, 57, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.J.; Foley, R.N.; Gilbertson, D.T.; Chen, S.C. United States Renal Data System public health surveillance of chronic kidney disease and end-stage renal disease. Kidney Int. Suppl. 2015, 5, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Zoccali, C.; Mallamaci, F.; Parlongo, S.; Cutrupi, S.; Benedetto, F.A.; Tripepi, G.; Bonanno, G.; Rapisarda, F.; Fatuzzo, P.; Seminara, G.; et al. Plasma norepinephrine predicts survival and incident cardiovascular events in patients with end-stage renal disease. Circulation 2002, 105, 1354–1359. [Google Scholar] [CrossRef] [PubMed]
- Mallamaci, F.; Tripepi, G.; Maas, R.; Malatino, L.; Boger, R.; Zoccali, C. Analysis of the relationship between norepinephrine and asymmetric dimethyl arginine levels among patients with end-stage renal disease. J. Am. Soc. Nephrol. 2004, 15, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Converse, R.L., Jr.; Jacobsen, T.N.; Toto, R.D.; Jost, C.M.; Cosentino, F.; Fouad-Tarazi, F.; Victor, R.G. Sympathetic overactivity in patients with chronic renal failure. N. Engl. J. Med. 1992, 327, 1912–1918. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Campese, V.M.; Nobakht, N.; Middlekauff, H.R. Differential distribution of muscle and skin sympathetic nerve activity in patients with end-stage renal disease. J. Appl. Physiol. 2008, 105, 1873–1876. [Google Scholar] [CrossRef] [PubMed]
- Grassi, G.; Quarti-Trevano, F.; Seravalle, G.; Arenare, F.; Volpe, M.; Furiani, S.; Dell’Oro, R.; Mancia, G. Early sympathetic activation in the initial clinical stages of chronic renal failure. Hypertension 2011, 57, 846–851. [Google Scholar] [CrossRef] [PubMed]
- Grassi, G.; Seravalle, G.; Ghiadoni, L.; Tripepi, G.; Bruno, R.M.; Mancia, G.; Zoccali, C. Sympathetic nerve traffic and asymmetric dimethylarginine in chronic kidney disease. Clin. J. Am. Soc. Nephrol. 2011, 6, 2620–2627. [Google Scholar] [CrossRef] [PubMed]
- Klein, I.H.; Ligtenberg, G.; Oey, P.L.; Koomans, H.A.; Blankestijn, P.J. Enalapril and losartan reduce sympathetic hyperactivity in patients with chronic renal failure. J. Am. Soc. Nephrol. 2003, 14, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Ligtenberg, G.; Blankestijn, P.J.; Oey, P.L.; Klein, I.H.; Dijkhorst-Oei, L.T.; Boomsma, F.; Wieneke, G.H.; van Huffelen, A.C.; Koomans, H.A. Reduction of sympathetic hyperactivity by enalapril in patients with chronic renal failure. N. Engl. J. Med. 1999, 340, 1321–1328. [Google Scholar] [CrossRef] [PubMed]
- Elias, A.N.; Vaziri, N.D.; Maksy, M. Plasma norepinephrine, epinephrine, and dopamine levels in end-stage renal disease. Effect of hemodialysis. Arch. Intern. Med. 1985, 145, 1013–1015. [Google Scholar] [CrossRef] [PubMed]
- Klein, I.H.; Ligtenberg, G.; Neumann, J.; Oey, P.L.; Koomans, H.A.; Blankestijn, P.J. Sympathetic nerve activity is inappropriately increased in chronic renal disease. J. Am. Soc. Nephrol. 2003, 14, 3239–3244. [Google Scholar] [CrossRef] [PubMed]
- Klein, I.H.; Ligtenberg, G.; Oey, P.L.; Koomans, H.A.; Blankestijn, P.J. Sympathetic activity is increased in polycystic kidney disease and is associated with hypertension. J. Am. Soc. Nephrol. 2001, 12, 2427–2433. [Google Scholar] [PubMed]
- Amann, K.; Rump, L.C.; Simonaviciene, A.; Oberhauser, V.; Wessels, S.; Orth, S.R.; Gross, M.L.; Koch, A.; Bielenberg, G.W.; van Kats, J.P.; et al. Effects of low dose sympathetic inhibition on glomerulosclerosis and albuminuria in subtotally nephrectomized rats. J. Am. Soc. Nephrol. 2000, 11, 1469–1478. [Google Scholar] [PubMed]
- Fisher, J.P.; Young, C.N.; Fadel, P.J. Central sympathetic overactivity: Maladies and mechanisms. Auton. Neurosci. 2009, 148, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Grassi, G.; Seravalle, G.; Dell’Oro, R.; Mancia, G. Sympathetic mechanisms, organ damage, and antihypertensive treatment. Curr. Hypertens. Rep. 2011, 13, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Abboud, F.M. The sympathetic system in hypertension. State-of-the-art review. Hypertension 1982, 4, 208–225. [Google Scholar] [PubMed]
- Smith, P.A.; Graham, L.N.; Mackintosh, A.F.; Stoker, J.B.; Mary, D.A. Relationship between central sympathetic activity and stages of human hypertension. Am. J. Hypertens. 2004, 17, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Grassi, G.; Cattaneo, B.M.; Seravalle, G.; Lanfranchi, A.; Mancia, G. Baroreflex control of sympathetic nerve activity in essential and secondary hypertension. Hypertension 1998, 31, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Levey, A.S. Controlling the epidemic of cardiovascular disease in chronic renal disease: Where do we start? Am. J. Kidney Dis. 1998, 32, 5–13. [Google Scholar] [CrossRef]
- Coresh, J.; Wei, G.L.; McQuillan, G.; Brancati, F.L.; Levey, A.S.; Jones, C.; Klag, M.J. Prevalence of high blood pressure and elevated serum creatinine level in the United States: Findings from the third National Health and Nutrition Examination Survey (1988–1994). Arch. Intern. Med. 2001, 161, 1207–1216. [Google Scholar] [CrossRef] [PubMed]
- Feihl, F.; Liaudet, L.; Levy, B.I.; Waeber, B. Hypertension and microvascular remodelling. Cardiovasc. Res. 2008, 78, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Folkow, B. Physiological aspects of primary hypertension. Physiol. Rev. 1982, 62, 347–504. [Google Scholar] [PubMed]
- Katholi, R.E.; Couri, D.M. Left ventricular hypertrophy: Major risk factor in patients with hypertension: Update and practical clinical applications. Int. J. Hypertens. 2011. [Google Scholar] [CrossRef] [PubMed]
- Simpson, P. Norepinephrine-stimulated hypertrophy of cultured rat myocardial cells is an alpha 1 adrenergic response. J. Clin. Investig. 1983, 72, 732–738. [Google Scholar] [CrossRef] [PubMed]
- Raum, W.J.; Laks, M.M.; Garner, D.; Ikuhara, M.H.; Swerdloff, R.S. Norepinephrine increases β-receptors and adenylate cyclase in canine myocardium. Am. J. Physiol. 1984, 246, H31–H36. [Google Scholar] [PubMed]
- Laks, M.M.; Morady, F.; Swan, H.J. Myocardial hypertrophy produced by chronic infusion of subhypertensive doses of norepinephrine in the dog. Chest 1973, 64, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Bevan, R.D. Effect of sympathetic denervation on smooth muscle cell proliferation in the growing rabbit ear artery. Circ. Res. 1975, 37, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Bevan, R.D. Trophic effects of peripheral adrenergic nerves on vascular structure. Hypertension 1984, 6, 19–26. [Google Scholar] [CrossRef]
- Coutsos, M.; Sala-Mercado, J.A.; Ichinose, M.; Li, Z.; Dawe, E.J.; O’Leary, D.S. Muscle metaboreflex-induced coronary vasoconstriction limits ventricular contractility during dynamic exercise in heart failure. Am. J. Physiol. Heart Circ. Physiol. 2013, 304, 1029–1037. [Google Scholar] [CrossRef] [PubMed]
- Volders, P.G. Novel insights into the role of the sympathetic nervous system in cardiac arrhythmogenesis. Heart Rhythm 2010, 7, 1900–1906. [Google Scholar] [CrossRef] [PubMed]
- DiBona, G.F. Sympathetic nervous system and the kidney in hypertension. Curr. Opin. Nephrol. Hypertens. 2002, 11, 197–200. [Google Scholar] [CrossRef] [PubMed]
- Rafiq, K.; Noma, T.; Fujisawa, Y.; Ishihara, Y.; Arai, Y.; Nabi, A.H.; Suzuki, F.; Nagai, Y.; Nakano, D.; Hitomi, H.; et al. Renal sympathetic denervation suppresses de novo podocyte injury and albuminuria in rats with aortic regurgitation. Circulation 2012, 125, 1402–1413. [Google Scholar] [CrossRef] [PubMed]
- Jamerson, K.A.; Julius, S.; Gudbrandsson, T.; Andersson, O.; Brant, D.O. Reflex sympathetic activation induces acute insulin resistance in the human forearm. Hypertension 1993, 21, 618–623. [Google Scholar] [CrossRef] [PubMed]
- Salzer, D.A.; Medeiros, P.J.; Craen, R.; Shoemaker, J.K. Neurogenic-nitric oxide interactions affecting brachial artery mechanics in humans: Roles of vessel distensibility vs. diameter. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 295, R1181–R1187. [Google Scholar] [CrossRef] [PubMed]
- Boutouyrie, P.; Lacolley, P.; Girerd, X.; Beck, L.; Safar, M.; Laurent, S. Sympathetic activation decreases medium-sized arterial compliance in humans. Am. J. Physiol. 1994, 267, H1368–H1376. [Google Scholar] [PubMed]
- Failla, M.; Grappiolo, A.; Emanuelli, G.; Vitale, G.; Fraschini, N.; Bigoni, M.; Grieco, N.; Denti, M.; Giannattasio, C.; Mancia, G. Sympathetic tone restrains arterial distensibility of healthy and atherosclerotic subjects. J. Hypertens. 1999, 17, 1117–1123. [Google Scholar] [CrossRef] [PubMed]
- Thijssen, D.H.; de, G.P.; Kooijman, M.; Smits, P.; Hopman, M.T. Sympathetic nervous system contributes to the age-related impairment of flow-mediated dilation of the superficial femoral artery. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, 3122–3129. [Google Scholar] [CrossRef] [PubMed]
- Hijmering, M.L.; Stroes, E.S.; Olijhoek, J.; Hutten, B.A.; Blankestijn, P.J.; Rabelink, T.J. Sympathetic activation markedly reduces endothelium-dependent, flow-mediated vasodilation. J. Am. Coll. Cardiol. 2002, 39, 683–688. [Google Scholar] [CrossRef]
- Santos, A.C.; Alves, M.J.; Rondon, M.U.; Barretto, A.C.; Middlekauff, H.R.; Negrao, C.E. Sympathetic activation restrains endothelium-mediated muscle vasodilatation in heart failure patients. Am. J. Physiol. Heart Circ. Physiol. 2005, 289, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, J.R.; Manuck, S.B. Using ethological principles to study psychosocial influences on coronary atherosclerosis in monkeys. Acta Physiol. Scand. Suppl. 1997, 640, 96–99. [Google Scholar] [PubMed]
- Kaplan, J.R.; Manuck, S.B.; Clarkson, T.B.; Lusso, F.M.; Taub, D.M.; Miller, E.W. Social stress and atherosclerosis in normocholesterolemic monkeys. Science 1983, 220, 733–735. [Google Scholar] [CrossRef] [PubMed]
- Strojek, K.; Grzeszczak, W.; Gorska, J.; Leschinger, M.I.; Ritz, E. Lowering of microalbuminuria in diabetic patients by a sympathicoplegic agent: Novel approach to prevent progression of diabetic nephropathy? J. Am. Soc. Nephrol. 2001, 12, 602–605. [Google Scholar] [PubMed]
- Anguiano, L.; Riera, M.; Pascual, J.; Valdivielso, J.M.; Barrios, C.; Betriu, A.; Clotet, S.; Mojal, S.; Fernandez, E.; Soler, M.J. Circulating angiotensin converting enzyme 2 activity as a biomarker of silent atherosclerosis in patients with chronic kidney disease. Atherosclerosis 2016, 253, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Medina, A.; Bell, P.R.; Briggs, J.D.; Brown, J.J.; Fine, A.; Lever, A.F.; Morton, J.J.; Paton, A.M.; Robertson, J.I.; Tree, M.; et al. Changes of blood pressure, renin, and angiotensin after bilateral nephrectomy in patients with chronic renal failure. Br. Med. J. 1972, 4, 694–696. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, H.N.; Rosenberg, M.E.; Hostetter, T.H. Role of the renin-angiotensin-aldosterone system in the progression of renal disease: A critical review. Semin. Nephrol. 1997, 17, 431–440. [Google Scholar] [PubMed]
- Reid, I.A. Interactions between ANG II, sympathetic nervous system, and baroreceptor reflexes in regulation of blood pressure. Am. J. Physiol. 1992, 262, E763–E778. [Google Scholar] [PubMed]
- Siddiqi, L.; Joles, J.A.; Grassi, G.; Blankestijn, P.J. Is kidney ischemia the central mechanism in parallel activation of the renin and sympathetic system? J. Hypertens. 2009, 27, 1341–1349. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Zhu, D.N.; Yu, Z.; Wang, J.Q.; Sun, Z.J.; Yao, T. Expression of angiotensin II type 1 (AT1) receptor in the rostral ventrolateral medulla in rats. J. Appl. Physiol. 2002, 92, 2153–2161. [Google Scholar] [CrossRef] [PubMed]
- Muratani, H.; Averill, D.B.; Ferrario, C.M. Effect of angiotensin II in ventrolateral medulla of spontaneously hypertensive rats. Am. J. Physiol. 1991, 260, 977–984. [Google Scholar]
- Hirooka, Y.; Potts, P.D.; Dampney, R.A. Role of angiotensin II receptor subtypes in mediating the sympathoexcitatory effects of exogenous and endogenous angiotensin peptides in the rostral ventrolateral medulla of the rabbit. Brain Res. 1997, 772, 107–114. [Google Scholar] [CrossRef]
- Allen, A.M. Blockade of angiotensin AT1-receptors in the rostral ventrolateral medulla of spontaneously hypertensive rats reduces blood pressure and sympathetic nerve discharge. J. Renin Angiotensin Aldosterone Syst. 2001, 2, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Allen, A.M.; Moeller, I.; Jenkins, T.A.; Zhuo, J.; Aldred, G.P.; Chai, S.Y.; Mendelsohn, F.A. Angiotensin receptors in the nervous system. Brain Res. Bull. 1998, 47, 17–28. [Google Scholar] [CrossRef]
- Davisson, R.L. Physiological genomic analysis of the brain renin-angiotensin system. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 285, 498–511. [Google Scholar] [CrossRef] [PubMed]
- Campese, V.M.; Kogosov, E. Renal afferent denervation prevents hypertension in rats with chronic renal failure. Hypertension 1995, 25, 878–882. [Google Scholar] [CrossRef] [PubMed]
- Ye, S.; Zhong, H.; Yanamadala, V.; Campese, V.M. Renal injury caused by intrarenal injection of phenol increases afferent and efferent renal sympathetic nerve activity. Am. J. Hypertens. 2002, 15, 717–724. [Google Scholar] [CrossRef]
- Ye, S.; Ozgur, B.; Campese, V.M. Renal afferent impulses, the posterior hypothalamus, and hypertension in rats with chronic renal failure. Kidney Int. 1997, 51, 722–727. [Google Scholar] [CrossRef] [PubMed]
- Ye, S.; Zhong, H.; Yanamadala, S.; Campese, V.M. Oxidative stress mediates the stimulation of sympathetic nerve activity in the phenol renal injury model of hypertension. Hypertension 2006, 48, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Katholi, R.E.; Whitlow, P.L.; Hageman, G.R.; Woods, W.T. Intrarenal adenosine produces hypertension by activating the sympathetic nervous system via the renal nerves in the dog. J. Hypertens. 1984, 2, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Recordati, G.; Moss, N.G.; Genovesi, S.; Rogenes, P. Renal chemoreceptors. J. Auton. Nerv. Syst. 1981, 3, 237–251. [Google Scholar] [CrossRef]
- Vallon, V.; Muhlbauer, B.; Osswald, H. Adenosine and kidney function. Physiol. Rev. 2006, 86, 901–940. [Google Scholar] [CrossRef] [PubMed]
- Hausberg, M.; Kosch, M.; Harmelink, P.; Barenbrock, M.; Hohage, H.; Kisters, K.; Dietl, K.H.; Rahn, K.H. Sympathetic nerve activity in end-stage renal disease. Circulation 2002, 106, 1974–1979. [Google Scholar] [CrossRef] [PubMed]
- Desir, G.V.; Peixoto, A.J. Renalase in hypertension and kidney disease. Nephrol. Dial. Transplant. 2014, 29, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Li, G.; Wang, P.; Velazquez, H.; Yao, X.; Li, Y.; Wu, Y.; Peixoto, A.; Crowley, S.; Desir, G.V. Renalase is a novel, soluble monoamine oxidase that regulates cardiac function and blood pressure. J. Clin. Investig. 2005, 115, 1275–1280. [Google Scholar] [CrossRef] [PubMed]
- Desir, G.V. Renalase deficiency in chronic kidney disease, and its contribution to hypertension and cardiovascular disease. Curr. Opin. Nephrol. Hypertens. 2008, 17, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Xu, J.; Wang, P.; Velazquez, H.; Li, Y.; Wu, Y.; Desir, G.V. Catecholamines regulate the activity, secretion, and synthesis of renalase. Circulation 2008, 117, 1277–1282. [Google Scholar] [CrossRef] [PubMed]
- Mungrue, I.N.; Bredt, D.S.; Stewart, D.J.; Husain, M. From molecules to mammals: What’s NOS got to do with it? Acta Physiol. Scand. 2003, 179, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, I.; Togashi, H.; Yoshioka, M.; Saito, H.; Yanagida, M.; Tamura, M.; Kobayashi, T.; Yasuda, H.; Gross, S.S.; Levi, R. NG-methyl-l-arginine, an inhibitor of l-arginine-derived nitric oxide synthesis, stimulates renal sympathetic nerve activity in vivo. A role for nitric oxide in the central regulation of sympathetic tone? Circ. Res. 1992, 70, 607–611. [Google Scholar] [CrossRef] [PubMed]
- Sander, M.; Hansen, J.; Victor, R.G. The sympathetic nervous system is involved in the maintenance but not initiation of the hypertension induced by N(omega)-nitro-l-arginine methyl ester. Hypertension 1997, 30, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Sander, M.; Chavoshan, B.; Victor, R.G. A large blood pressure-raising effect of nitric oxide synthase inhibition in humans. Hypertension 1999, 33, 937–942. [Google Scholar] [CrossRef] [PubMed]
- Cannon, R.O., III. Role of nitric oxide in cardiovascular disease: Focus on the endothelium. Clin. Chem. 1998, 44, 1809–1819. [Google Scholar] [PubMed]
- Cooke, J.P.; Dzau, V.J. Nitric oxide synthase: Role in the genesis of vascular disease. Annu. Rev. Med. 1997, 48, 489–509. [Google Scholar] [CrossRef] [PubMed]
- Shapoval, L.N.; Sagach, V.F.; Pobegailo, L.S. Nitric oxide influences ventrolateral medullary mechanisms of vasomotor control in the cat. Neurosci. Lett. 1991, 132, 47–50. [Google Scholar] [CrossRef]
- Harada, S.; Tokunaga, S.; Momohara, M.; Masaki, H.; Tagawa, T.; Imaizumi, T.; Takeshita, A. Inhibition of nitric oxide formation in the nucleus tractus solitarius increases renal sympathetic nerve activity in rabbits. Circ. Res. 1993, 72, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Tagawa, T.; Imaizumi, T.; Harada, S.; Endo, T.; Shiramoto, M.; Hirooka, Y.; Takeshita, A. Nitric oxide influences neuronal activity in the nucleus tractus solitarius of rat brainstem slices. Circ. Res. 1994, 75, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Togashi, H.; Sakuma, I.; Yoshioka, M.; Kobayashi, T.; Yasuda, H.; Kitabatake, A.; Saito, H.; Gross, S.S.; Levi, R. A central nervous system action of nitric oxide in blood pressure regulation. J. Pharmacol. Exp. Ther. 1992, 262, 343–347. [Google Scholar] [PubMed]
- Kishi, T.; Hirooka, Y.; Sakai, K.; Shigematsu, H.; Shimokawa, H.; Takeshita, A. Overexpression of eNOS in the RVLM causes hypotension and bradycardia via GABA release. Hypertension 2001, 38, 896–901. [Google Scholar] [PubMed]
- Kishi, T.; Hirooka, Y.; Ito, K.; Sakai, K.; Shimokawa, H.; Takeshita, A. Cardiovascular effects of overexpression of endothelial nitric oxide synthase in the rostral ventrolateral medulla in stroke-prone spontaneously hypertensive rats. Hypertension 2002, 39, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, H.; Averill, D.B.; Khosla, M.C.; Ferrario, C.M. Role of nitric oxide and angiotensin II in the regulation of sympathetic nerve activity in spontaneously hypertensive rats. Hypertension 1993, 21, 476–484. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J.; Jacobsen, T.N.; Victor, R.G. Is nitric oxide involved in the tonic inhibition of central sympathetic outflow in humans? Hypertension 1994, 24, 439–444. [Google Scholar] [CrossRef] [PubMed]
- Augustyniak, R.A.; Victor, R.G.; Morgan, D.A.; Zhang, W. l-NAME and ADMA-induced sympathetic neural activation in conscious rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 290, R726–R732. [Google Scholar] [CrossRef] [PubMed]
- Young, C.N.; Fisher, J.P.; Gallagher, K.M.; Whaley-Connell, A.; Chaudhary, K.; Victor, R.G.; Thomas, G.D.; Fadel, P.J. Inhibition of nitric oxide synthase evokes central sympatho-excitation in healthy humans. J. Physiol. 2009, 587, 4977–4986. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Wagner, L.; Schmidt, R.J.; Baylis, C. Circulating endothelial nitric oxide synthase inhibitory factor in some patients with chronic renal disease. Kidney Int. 2001, 59, 1466–1472. [Google Scholar] [CrossRef] [PubMed]
- Kielstein, J.T.; Boger, R.H.; Bode-Boger, S.M.; Frolich, J.C.; Haller, H.; Ritz, E.; Fliser, D. Marked increase of asymmetric dimethylarginine in patients with incipient primary chronic renal disease. J. Am. Soc. Nephrol. 2002, 13, 170–176. [Google Scholar] [PubMed]
- Ravani, P.; Tripepi, G.; Malberti, F.; Testa, S.; Mallamaci, F.; Zoccali, C. Asymmetrical dimethylarginine predicts progression to dialysis and death in patients with chronic kidney disease: A competing risks modeling approach. J. Am. Soc. Nephrol. 2005, 16, 2449–2455. [Google Scholar] [CrossRef] [PubMed]
- Ueda, S.; Yamagishi, S.; Kaida, Y.; Okuda, S. Asymmetric dimethylarginine may be a missing link between cardiovascular disease and chronic kidney disease. Nephrology 2007, 12, 582–590. [Google Scholar] [CrossRef] [PubMed]
- Zoccali, C. Asymmetric dimethylarginine (ADMA): A cardiovascular and renal risk factor on the move. J. Hypertens. 2006, 24, 611–619. [Google Scholar] [CrossRef] [PubMed]
- Zoccali, C.; Bode-Boger, S.; Mallamaci, F.; Benedetto, F.; Tripepi, G.; Malatino, L.; Cataliotti, A.; Bellanuova, I.; Fermo, I.; Frolich, J.; et al. Plasma concentration of asymmetrical dimethylarginine and mortality in patients with end-stage renal disease: A prospective study. Lancet 2001, 358, 2113–2117. [Google Scholar] [CrossRef]
- Fliser, D.; Kronenberg, F.; Kielstein, J.T.; Morath, C.; Bode-Boger, S.M.; Haller, H.; Ritz, E. Asymmetric dimethylarginine and progression of chronic kidney disease: The mild to moderate kidney disease study. J. Am. Soc. Nephrol. 2005, 16, 2456–2461. [Google Scholar] [CrossRef] [PubMed]
- Boger, R.H.; Bode-Boger, S.M.; Szuba, A.; Tsao, P.S.; Chan, J.R.; Tangphao, O.; Blaschke, T.F.; Cooke, J.P. Asymmetric dimethylarginine (ADMA): A novel risk factor for endothelial dysfunction: Its role in hypercholesterolemia. Circulation 1998, 98, 1842–1847. [Google Scholar] [CrossRef] [PubMed]
- Young, J.M.; Terrin, N.; Wang, X.; Greene, T.; Beck, G.J.; Kusek, J.W.; Collins, A.J.; Sarnak, M.J.; Menon, V. Asymmetric dimethylarginine and mortality in stages 3 to 4 chronic kidney disease. Clin. J. Am. Soc. Nephrol. 2009, 4, 1115–1120. [Google Scholar] [CrossRef] [PubMed]
- Watson, C.P.; Pazarentzos, E.; Fidanboylu, M.; Padilla, B.; Brown, R.; Thomas, S.A. The transporter and permeability interactions of asymmetric dimethylarginine (ADMA) and l-arginine with the human blood-brain barrier in vitro. Brain Res. 2016, 1648, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Vallance, P.; Leiper, J. Cardiovascular biology of the asymmetric dimethylarginine:dimethylarginine dimethylaminohydrolase pathway. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 1023–1030. [Google Scholar] [CrossRef] [PubMed]
- Tran, C.T.; Leiper, J.M.; Vallance, P. The DDAH/ADMA/NOS pathway. Atheroscler. Suppl. 2003, 4, 33–40. [Google Scholar] [CrossRef]
- Achan, V.; Broadhead, M.; Malaki, M.; Whitley, G.; Leiper, J.; MacAllister, R.; Vallance, P. Asymmetric dimethylarginine causes hypertension and cardiac dysfunction in humans and is actively metabolized by dimethylarginine dimethylaminohydrolase. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 1455–1459. [Google Scholar] [CrossRef] [PubMed]
- Dounousi, E.; Papavasiliou, E.; Makedou, A.; Ioannou, K.; Katopodis, K.P.; Tselepis, A.; Siamopoulos, K.C.; Tsakiris, D. Oxidative stress is progressively enhanced with advancing stages of CKD. Am. J. Kidney Dis. 2006, 48, 752–760. [Google Scholar] [CrossRef] [PubMed]
- Oberg, B.P.; McMenamin, E.; Lucas, F.L.; McMonagle, E.; Morrow, J.; Ikizler, T.A.; Himmelfarb, J. Increased prevalence of oxidant stress and inflammation in patients with moderate to severe chronic kidney disease. Kidney Int. 2004, 65, 1009–1016. [Google Scholar] [CrossRef] [PubMed]
- Himmelfarb, J.; Stenvinkel, P.; Ikizler, T.A.; Hakim, R.M. The elephant in uremia: Oxidant stress as a unifying concept of cardiovascular disease in uremia. Kidney Int. 2002, 62, 1524–1538. [Google Scholar] [CrossRef] [PubMed]
- Martinez Cantarin, M.P.; Whitaker-Menezes, D.; Lin, Z.; Falkner, B. Uremia induces adipose tissue inflammation and muscle mitochondrial dysfunction. Nephrol. Dial. Transplant. 2017, 32, 943–951. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Morimoto, S.; Okigaki, M.; Seo, M.; Someya, K.; Morita, T.; Matsubara, H.; Sugiura, T.; Iwasaka, T. Decreased plasma level of vitamin C in chronic kidney disease: Comparison between diabetic and non-diabetic patients. Nephrol. Dial. Transplant. 2011, 26, 1252–1257. [Google Scholar] [CrossRef] [PubMed]
- Hirooka, Y.; Kishi, T.; Sakai, K.; Takeshita, A.; Sunagawa, K. Imbalance of central nitric oxide and reactive oxygen species in the regulation of sympathetic activity and neural mechanisms of hypertension. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, R818–R826. [Google Scholar] [CrossRef] [PubMed]
- Oliveira-Sales, E.B.; Nishi, E.E.; Carillo, B.A.; Boim, M.A.; Dolnikoff, M.S.; Bergamaschi, C.T.; Campos, R.R. Oxidative stress in the sympathetic premotor neurons contributes to sympathetic activation in renovascular hypertension. Am. J. Hypertens. 2009, 22, 484–492. [Google Scholar] [CrossRef] [PubMed]
- Nishihara, M.; Hirooka, Y.; Matsukawa, R.; Kishi, T.; Sunagawa, K. Oxidative stress in the rostral ventrolateral medulla modulates excitatory and inhibitory inputs in spontaneously hypertensive rats. J. Hypertens. 2012, 30, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Kishi, T.; Hirooka, Y.; Kimura, Y.; Ito, K.; Shimokawa, H.; Takeshita, A. Increased reactive oxygen species in rostral ventrolateral medulla contribute to neural mechanisms of hypertension in stroke-prone spontaneously hypertensive rats. Circulation 2004, 109, 2357–2362. [Google Scholar] [CrossRef] [PubMed]
- Campese, V.M.; Ye, S.; Zhong, H.; Yanamadala, V.; Ye, Z.; Chiu, J. Reactive oxygen species stimulate central and peripheral sympathetic nervous system activity. Am. J. Physiol. Heart Circ. Physiol. 2004, 287, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Griendling, K.K.; Minieri, C.A.; Ollerenshaw, J.D.; Alexander, R.W. Angiotensin II stimulates NADH and NADPH oxidase activity in cultured vascular smooth muscle cells. Circ. Res. 1994, 74, 1141–1148. [Google Scholar] [CrossRef] [PubMed]
- Campese, V.M.; Shaohua, Y.; Huiquin, Z. Oxidative stress mediates angiotensin II-dependent stimulation of sympathetic nerve activity. Hypertension 2005, 46, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.H.; Hsu, K.S.; Huang, C.C.; Wang, L.L.; Ou, C.C.; Chan, J.Y. NADPH oxidase-derived superoxide anion mediates angiotensin II-induced pressor effect via activation of p38 mitogen-activated protein kinase in the rostral ventrolateral medulla. Circ. Res. 2005, 97, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.P.; Mayhan, W.G.; Bidasee, K.R.; Zheng, H. Enhanced angiotensin II-mediated central sympathoexcitation in streptozotocin-induced diabetes: Role of superoxide anion. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, R311–R320. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, M.C.; Lazartigues, E.; Lang, J.A.; Sinnayah, P.; Ahmad, I.M.; Spitz, D.R.; Davisson, R.L. Superoxide mediates the actions of angiotensin II in the central nervous system. Circ. Res. 2002, 91, 1038–1045. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Chen, A.D.; Xu, Y.; Chen, Q.; Gao, X.Y.; Wang, W.; Zhu, G.Q. Long-term administration of tempol attenuates postinfarct ventricular dysfunction and sympathetic activity in rats. Pflug. Arch. 2009, 458, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.G.; Zhang, Z.H.; Yu, Y.; Felder, R.B. Systemically administered tempol reduces neuronal activity in paraventricular nucleus of hypothalamus and rostral ventrolateral medulla in rats. J. Hypertens. 2009, 27, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Ito, A.; Tsao, P.S.; Adimoolam, S.; Kimoto, M.; Ogawa, T.; Cooke, J.P. Novel mechanism for endothelial dysfunction: Dysregulation of dimethylarginine dimethylaminohydrolase. Circulation 1999, 99, 3092–3095. [Google Scholar] [CrossRef] [PubMed]
- Cohn, J.N.; Pfeffer, M.A.; Rouleau, J.; Sharpe, N.; Swedberg, K.; Straub, M.; Wiltse, C.; Wright, T.J. Adverse mortality effect of central sympathetic inhibition with sustained-release moxonidine in patients with heart failure (MOXCON). Eur. J. Heart Fail. 2003, 5, 659–667. [Google Scholar] [CrossRef]
- Planitz, V. Crossover comparison of moxonidine and clonidine in mild to moderate hypertension. Eur. J. Clin. Pharmacol. 1984, 27, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Swedberg, K.; Bristow, M.R.; Cohn, J.N.; Dargie, H.; Straub, M.; Wiltse, C.; Wright, T.J. Effects of sustained-release moxonidine, an imidazoline agonist, on plasma norepinephrine in patients with chronic heart failure. Circulation 2002, 105, 1797–1803. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Wang, W.; Li, Y.L.; Schultz, H.D.; Liu, D.; Cornish, K.G.; Zucker, I.H. Simvastatin therapy normalizes sympathetic neural control in experimental heart failure: Roles of angiotensin II type 1 receptors and NAD(P)H oxidase. Circulation 2005, 112, 1763–1770. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Wang, W.; Zucker, I.H. Simvastatin inhibits central sympathetic outflow in heart failure by a nitric-oxide synthase mechanism. J. Pharmacol. Exp. Ther. 2008, 326, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Deo, S.H.; Fisher, J.P.; Vianna, L.C.; Kim, A.; Chockalingam, A.; Zimmerman, M.C.; Zucker, I.H.; Fadel, P.J. Statin therapy lowers muscle sympathetic nerve activity and oxidative stress in patients with heart failure. Am. J. Physiol. Heart Circ. Physiol. 2012, 303, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Lewandowski, J.; Symonides, B.; Gaciong, Z.; Sinski, M. The effect of statins on sympathetic activity: A meta-analysis. Clin. Auton. Res. 2015, 25, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Siddiqi, L.; Joles, J.A.; Oey, P.L.; Blankestijn, P.J. Atorvastatin reduces sympathetic activity in patients with chronic kidney disease. J. Hypertens. 2011, 29, 2176–2180. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.M.; Lin, M.S.; Hsu, J.T.; Hsiao, J.F.; Chang, S.T.; Pan, K.L.; Lin, C.L.; Lin, Y.S. Effects of statin therapy on cerebrovascular and renal outcomes in patients with predialysis advanced chronic kidney disease and dyslipidemia. J. Clin. Lipidol. 2017, 11, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Major, R.W.; Cheung, C.K.; Gray, L.J.; Brunskill, N.J. Statins and Cardiovascular Primary Prevention in CKD: A Meta-Analysis. Clin. J. Am. Soc. Nephrol. 2015, 10, 732–739. [Google Scholar] [CrossRef] [PubMed]
- Messow, C.M.; Isles, C. Meta-analysis of statins in chronic kidney disease: Who benefits? QJM 2017. [Google Scholar] [CrossRef] [PubMed]
- Boaz, M.; Smetana, S.; Weinstein, T.; Matas, Z.; Gafter, U.; Iaina, A.; Knecht, A.; Weissgarten, Y.; Brunner, D.; Fainaru, M.; et al. Secondary prevention with antioxidants of cardiovascular disease in endstage renal disease (SPACE): Randomised placebo-controlled trial. Lancet 2000, 356, 1213–1218. [Google Scholar] [CrossRef]
- Bruno, R.M.; Daghini, E.; Ghiadoni, L.; Sudano, I.; Rugani, I.; Varanini, M.; Passino, C.; Emdin, M.; Taddei, S. Effect of acute administration of vitamin C on muscle sympathetic activity, cardiac sympathovagal balance, and baroreflex sensitivity in hypertensive patients. Am. J. Clin. Nutr. 2012, 96, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.E.; El, M.S.; Lenders, J.W.; Bellersen, L.; Verheugt, F.W.; Smits, P.; Tack, C.J. High dose ascorbic acid does not reverse central sympathetic overactivity in chronic heart failure. J. Clin. Pharm. Ther. 2011, 36, 546–552. [Google Scholar] [CrossRef] [PubMed]
- Singer, R.F. Vitamin C supplementation in kidney failure: Effect on uraemic symptoms. Nephrol. Dial. Transplant. 2011, 26, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Tahara, N.; Yamagishi, S.; Mizoguchi, M.; Tahara, A.; Imaizumi, T. Pioglitazone decreases asymmetric dimethylarginine levels in patients with impaired glucose tolerance or type 2 diabetes. Rejuvenation Res. 2013, 16, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Wakino, S.; Hayashi, K.; Tatematsu, S.; Hasegawa, K.; Takamatsu, I.; Kanda, T.; Homma, K.; Yoshioka, K.; Sugano, N.; Saruta, T. Pioglitazone lowers systemic asymmetric dimethylarginine by inducing dimethylarginine dimethylaminohydrolase in rats. Hypertens Res. 2005, 28, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Esler, M.D.; Krum, H.; Sobotka, P.A.; Schlaich, M.P.; Schmieder, R.E.; Bohm, M. Renal sympathetic denervation in patients with treatment-resistant hypertension (The symplicity HTN-2 trial): A randomised controlled trial. Lancet 2010, 376, 1903–1909. [Google Scholar] [PubMed]
- Krum, H.; Schlaich, M.; Whitbourn, R.; Sobotka, P.A.; Sadowski, J.; Bartus, K.; Kapelak, B.; Walton, A.; Sievert, H.; Thambar, S.; et al. Catheter-based renal sympathetic denervation for resistant hypertension: A multicentre safety and proof-of-principle cohort study. Lancet 2009, 373, 1275–1281. [Google Scholar] [CrossRef]
- Jacob, F.; Ariza, P.; Osborn, J.W. Renal denervation chronically lowers arterial pressure independent of dietary sodium intake in normal rats. Am. J. Physiol. Heart Circ. Physiol. 2003, 284, H2302–H2310. [Google Scholar] [CrossRef] [PubMed]
- Ojeda, N.B.; Johnson, W.R.; Dwyer, T.M.; Alexander, B.T. Early renal denervation prevents development of hypertension in growth-restricted offspring. Clin. Exp. Pharmacol. Physiol. 2007, 34, 1212–1216. [Google Scholar] [CrossRef] [PubMed]
- Katholi, R.E.; Rocha-Singh, K.J. The role of renal sympathetic nerves in hypertension: Has percutaneous renal denervation refocused attention on their clinical significance? Prog. Cardiovasc. Dis. 2009, 52, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Hendel, M.D.; Collister, J.P. Renal denervation attenuates long-term hypertensive effects of Angiotensin II in the rat. Clin. Exp. Pharmacol. Physiol. 2006, 33, 1225–1230. [Google Scholar] [CrossRef] [PubMed]
- Lohmeier, T.E.; Hildebrandt, D.A.; Dwyer, T.M.; Barrett, A.M.; Irwin, E.D.; Rossing, M.A.; Kieval, R.S. Renal denervation does not abolish sustained baroreflex-mediated reductions in arterial pressure. Hypertension 2007, 49, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, D.L.; Kandzari, D.E.; O’Neill, W.W.; D’Agostino, R.; Flack, J.M.; Katzen, B.T.; Leon, M.B.; Liu, M.; Mauri, L.; Negoita, M.; et al. A controlled trial of renal denervation for resistant hypertension. N. Engl. J. Med. 2014, 370, 1393–1401. [Google Scholar] [CrossRef] [PubMed]
- Hering, D.; Mahfoud, F.; Walton, A.S.; Krum, H.; Lambert, G.W.; Lambert, E.A.; Sobotka, P.A.; Bohm, M.; Cremers, B.; Esler, M.D.; et al. Renal denervation in moderate to severe CKD. J. Am. Soc. Nephrol. 2012, 23, 1250–1257. [Google Scholar] [CrossRef] [PubMed]
- Papademetriou, V.; Doumas, M.; Anyfanti, P.; Faselis, C.; Kokkinos, P.; Tsioufis, C. Renal nerve ablation for hypertensive patients with chronic kidney disease. Curr. Vasc. Pharmacol. 2014, 12, 47–54. [Google Scholar] [PubMed]
- Gattone, V.H.; Siqueira, T.M., Jr.; Powell, C.R.; Trambaugh, C.M.; Lingeman, J.E.; Shalhav, A.L. Contribution of renal innervation to hypertension in rat autosomal dominant polycystic kidney disease. Exp. Biol. Med. 2008, 233, 952–957. [Google Scholar] [CrossRef] [PubMed]
- Kiuchi, M.G.; Maia, G.L.; de Queiroz Carreira, M.A.; Kiuchi, T.; Chen, S.; Andrea, B.R.; Graciano, M.L.; Lugon, J.R. Effects of renal denervation with a standard irrigated cardiac ablation catheter on blood pressure and renal function in patients with chronic kidney disease and resistant hypertension. Eur. Heart J. 2013, 34, 2114–2121. [Google Scholar] [CrossRef] [PubMed]
- Schlaich, M.P.; Bart, B.; Hering, D.; Walton, A.; Marusic, P.; Mahfoud, F.; Bohm, M.; Lambert, E.A.; Krum, H.; Sobotka, P.A.; et al. Feasibility of catheter-based renal nerve ablation and effects on sympathetic nerve activity and blood pressure in patients with end-stage renal disease. Int. J. Cardiol. 2013, 168, 2214–2220. [Google Scholar] [CrossRef] [PubMed]
- Lohmeier, T.E.; Iliescu, R.; Dwyer, T.M.; Irwin, E.D.; Cates, A.W.; Rossing, M.A. Sustained suppression of sympathetic activity and arterial pressure during chronic activation of the carotid baroreflex. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Heusser, K.; Tank, J.; Engeli, S.; Diedrich, A.; Menne, J.; Eckert, S.; Peters, T.; Sweep, F.C.; Haller, H.; Pichlmaier, A.M.; et al. Carotid baroreceptor stimulation, sympathetic activity, baroreflex function, and blood pressure in hypertensive patients. Hypertension 2010, 55, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Bisognano, J.D.; Bakris, G.; Nadim, M.K.; Sanchez, L.; Kroon, A.A.; Schafer, J.; de Leeuw, P.W.; Sica, D.A. Baroreflex activation therapy lowers blood pressure in patients with resistant hypertension: Results from the double-blind, randomized, placebo-controlled rheos pivotal trial. J. Am. Coll. Cardiol. 2011, 58, 765–773. [Google Scholar] [CrossRef] [PubMed]

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaur, J.; Young, B.E.; Fadel, P.J. Sympathetic Overactivity in Chronic Kidney Disease: Consequences and Mechanisms. Int. J. Mol. Sci. 2017, 18, 1682. https://doi.org/10.3390/ijms18081682
Kaur J, Young BE, Fadel PJ. Sympathetic Overactivity in Chronic Kidney Disease: Consequences and Mechanisms. International Journal of Molecular Sciences. 2017; 18(8):1682. https://doi.org/10.3390/ijms18081682
Chicago/Turabian StyleKaur, Jasdeep, Benjamin E. Young, and Paul J. Fadel. 2017. "Sympathetic Overactivity in Chronic Kidney Disease: Consequences and Mechanisms" International Journal of Molecular Sciences 18, no. 8: 1682. https://doi.org/10.3390/ijms18081682
APA StyleKaur, J., Young, B. E., & Fadel, P. J. (2017). Sympathetic Overactivity in Chronic Kidney Disease: Consequences and Mechanisms. International Journal of Molecular Sciences, 18(8), 1682. https://doi.org/10.3390/ijms18081682