Relationship of Triamine-Biocide Tolerance of Salmonella enterica Serovar Senftenberg to Antimicrobial Susceptibility, Serum Resistance and Outer Membrane Proteins

 , , ,
, , ,
Abstract
1. Introduction
2. Results
2.1. Salmonella enterica Tolerance to the Biocide Formulation
2.2. Antibiotic Susceptibility Profiling
2.3. Bactericidal Activity of Human Serum against S. Senftenberg Variants Tolerant to the Disinfectant
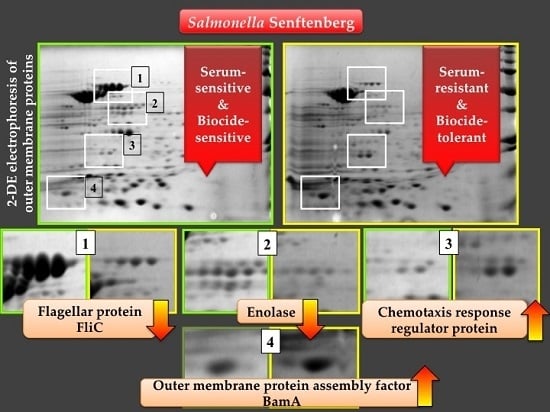
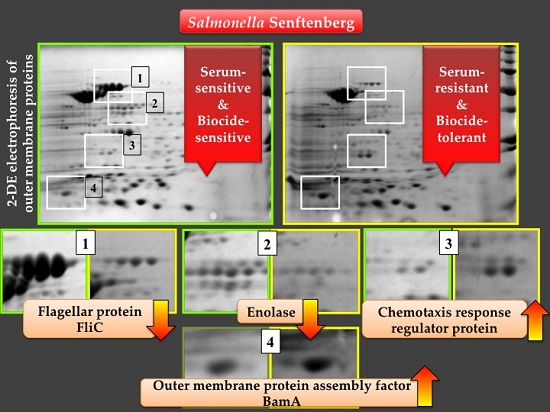
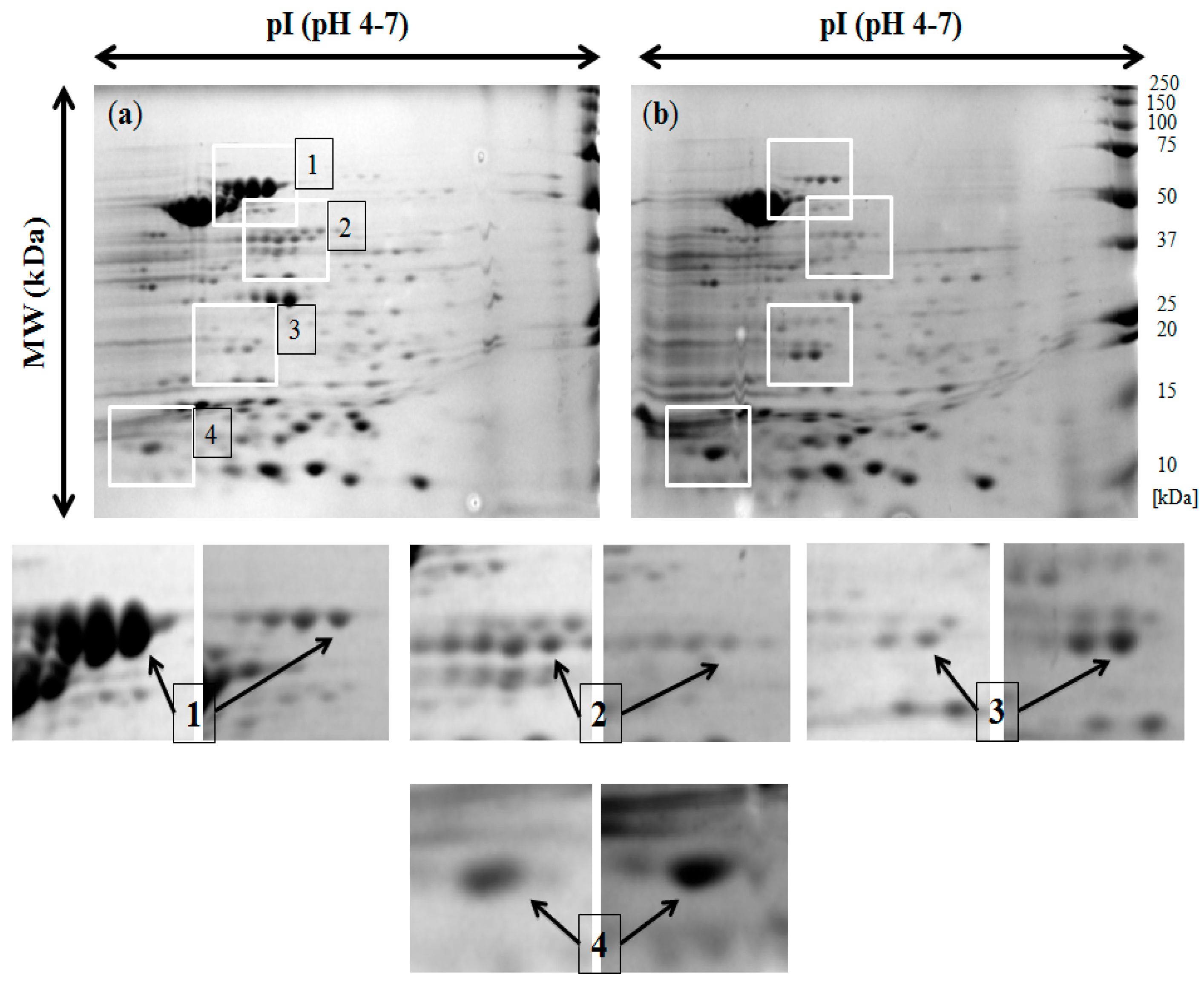
2.4. Analysis of the Two-Dimensional (2-DE) Profiles of Isolated Membrane Proteins
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Disinfectants and Antibiotics
4.3. Antimicrobial Susceptibility
4.4. Isolation of Biocide Tolerant Variants and the Stability of Their Phenotypes
4.5. Serum
4.6. Serum C3 Concentration
4.7. Serum Susceptibility Assay
4.8. Outer Membrane Proteins Isolation and Preparation
4.9. Two-Dimensional Gel Electrophoresis
4.10. In-Gel Protein Digestion and MS Protein Identification
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| aST | After the test of stability |
| bST | Before the test of stability |
| CFU | Colony-forming units |
| h | Hours |
| HIS | Heat-inactivated normal human serum |
| MDR | Multidrug-resistant |
| MIC | Minimal inhibitory concentration |
| MBC | Minimal bactericidal concentration |
| NHS | Normal human serum |
| OMPs | Outer membrane proteins |
| RP | Resistance pattern |
| QAC | Quaternary ammonium salts |
| R | Resistant |
| S | Sensitive |
| 2-DE | Two-dimensional gel electrophoresis |
References
- Ortega Morente, E.; Fernández-Fuentes, M.A.; Grande Burgos, M.J.; Abriouel, H.; Pérez Pulido, R.; Gálvez, A. Biocide tolerance in bacteria. Int. J. Food Microbiol. 2013, 162, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Shengzhi, Y.; Guoyan, W.; Mei, L.; Wenwen, D.; Hongning, W.; Likou, Z. Antibiotic and disinfectant resistance of Salmonella isolated from egg production chains. Yi Chuan 2016, 38, 948–956. [Google Scholar] [PubMed]
- Whitehead, R.N.; Overton, T.W.; Kemp, C.L.; Webber, M.A. Exposure of Salmonella enterica serovar Typhimurium to high level biocide challenge can select multidrug resistant mutants in a single step. PLoS ONE 2011, 6, e22833. [Google Scholar] [CrossRef] [PubMed]
- Gadea, R.; Glibota, N.; Pérez Pulido, R.; Gálvez, A.; Ortega, E. Adaptation to biocides cetrimide and chlorhexidine in bacteria from organic foods: Association with tolerance to other antimicrobials and physical stresses. J. Agric. Food Chem. 2017, 65, 1758–1770. [Google Scholar] [CrossRef] [PubMed]
- Silha, D.; Silhová, L.; Vytrasová, J. Survival of selected bacteria of Arcobacter genus in disinfectants and possibility of acquired secondary resistance to disinfectants. J. Microb. Biotech. Food Sci. 2016, 5, 326–329. [Google Scholar] [CrossRef]
- Curiao, T.; Marchi, E.; Grandgirard, D.; León-Sampedro, R.; Viti, C.; Leib, S.L.; Baquero, F.; Oggioni, M.R.; Martinez, J.L.; Coque, T.M. Multiple adaptive routes of Salmonella enterica Typhimurium to biocide and antibiotic exposure. BMC Genom. 2016, 17, 491–507. [Google Scholar] [CrossRef] [PubMed]
- Majtánová, L.; Majtán, V. Effect of disinfectants on surface hydrophobicity and mobility in Salmonella enterica serovar Typhimurium DT104. Ceska Slov. Farm. 2003, 52, 141–147. [Google Scholar] [PubMed]
- Futoma-Kołoch, B.; Ksiażczyk, M.; Korzekwa, K.; Migdał, I.; Pawlak, A.; Jankowska, M.; Kędziora, A.; Dorotkiewicz-Jach, A.; Bugla-Płoskońska, G. Selection and electrophoretic characterization of Salmonella enterica subsp. enterica biocide variants resistant to antibiotics. Pol. J. Vet. Sci. 2015, 18, 725–732. [Google Scholar]
- Futoma-Kołoch, B.; Ksiażczyk, M. The risk of Salmonella resistance following exposure to common disinfectants: An emerging problem. Biol. Int. 2013, 53, 54–66. [Google Scholar]
- Majtán, V.; Majtánová, L. Effect of disinfectants on the metabolism of Salmonella enterica serovar enteritidis. Folia Microbiol. 2003, 48, 643–648. [Google Scholar] [CrossRef]
- Uzer Celik, E.; Tunac, A.T.; Ates, M.; Sen, B.H. Antimicrobial activity of different disinfectants against cariogenic microorganisms. Braz. Oral Res. 2016, 30, e125. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chapman, J.S. Biocide resistance mechanisms. Int. Biod. Biodegr. 2003, 51, 133–138. [Google Scholar] [CrossRef]
- Randall, L.P.; Coles, S.W.; Coldham, N.G.; Penuela, L.G.; Mott, A.C.; Woodward, M.J.; Piddock, L.J.V.; Webber, M. Commonly used farm disinfectants can select for mutant Salmonella enterica serovar Typhimurium with decreased susceptibility to biocides and antibiotics without compromising virulence. J. Antimicrob. Chemother. 2007, 60, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- Son, M.S.; Del Castilho, C.; Duncalf, K.A.; Carney, D.; Weiner, J.H.; Turner, R.J. Mutagenesis of SugE, a small multidrug resistance protein. Biochem. Biophys. Res. Commun. 2003, 312, 914–921. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.; Meng, J.; McDermott, P.F.; Wang, F.; Yang, Q.; Cao, G.; Hoffmann, M.; Zhao, S. Presence of disinfectant resistance genes in Escherichia coli isolated from retail meats in the USA. J. Antimicrob. Chemother. 2014, 69, 2644–2649. [Google Scholar] [CrossRef] [PubMed]
- Kariuki, S.; Gordon, M.A.; Feasey, N.; Parry, C.M. Antimicrobial resistance and management of invasive Salmonella disease. Vaccine 2015, 33, C21–C29. [Google Scholar] [CrossRef] [PubMed]
- Futoma-Kołoch, B.; Bugla-Płoskońska, G.; Sarowska, J. Searching for outer membrane proteins typical of serum-sensitive and serum-resistant phenotypes of Salmonella. In Salmonella-Distribution, Adaptation, Control Measures, and Molecular Technologies; Annous, B.A., Ed.; InTech: Rijeka, Croatia, 2012; pp. 265–290. [Google Scholar]
- Riva, R.; Korhonen, T.K.; Meri, S. The outer membrane protease PgtE of Salmonella enterica interferes with the alternative complement pathway by cleaving factors B and H. Front. Microbiol. 2015, 6, 63. [Google Scholar] [CrossRef] [PubMed]
- Futoma-Kołoch, B.; Godlewska, U.; Guz-Regner, K.; Dorotkiewicz-Jach, A.; Klausa, E.; Rybka, J.; Bugla-Płoskońska, G. Presumable role of outer membrane proteins of Salmonella containing sialylated lipopolysaccharides serovar Ngozi, sv. Isaszeg and subspecies arizonae in determining susceptibility to human serum. Gut Pathog. 2015, 7, 18. [Google Scholar] [CrossRef] [PubMed]
- Dudek, B.; Krzyżewska, E.; Kapczyńska, K.; Rybka, J.; Pawlak, A.; Korzekwa, K.; Klausa, E.; Bugla-Płoskońska, G. Proteomic analysis of outer membrane proteins from Salmonella Enteritidis strains with different sensitivity to human serum. PLoS ONE 2016, 11, e0164069. [Google Scholar] [CrossRef] [PubMed]
- McDonnell, G.; Russell, A.D. Antiseptics and disinfectants: Activity, action, and resistance. Clin. Microbiol. Rev. 1999, 12, 147–179. [Google Scholar] [PubMed]
- Karatzas, K.A.G.; Randall, L.P.; Webber, M.; Piddock, L.J.V.; Humphrey, T.J.; Woodward, M.J.; Coldham, N.G. Phenotypic and proteomic characterization of MAR variants of Salmonella enterica serovar Typhimurium selected following exposure to disinfectants. Appl. Environ. Microbiol. 2007, 74, 1508–1516. [Google Scholar] [CrossRef] [PubMed]
- Condell, O.; Sheridan, Á.; Power, K.A.; Bonilla-Santiago, R.; Sergeant, K.; Renault, J.; Burgess, C.; Fanning, S.; Nally, J.E. Comparative proteomic analysis of Salmonella tolerance to the biocide active agent triclosan. J. Proteom. 2012, 75, 4505–4519. [Google Scholar] [CrossRef] [PubMed]
- Roy, V.; Adams, B.L.; Bentley, W.E. Developing next generation antimicrobials by intercepting AI-2 mediated quorum sensing. Enz. Microb. Technol. 2011, 49, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Müderris, T.; Ürkmez, F.Y.; Küçüker, Ş.A.; Sağlam, M.F.; Yılmaz, G.R.; Güner, R.; Güleşen, R.; Açıkgöz, Z.C. Bacteremia caused by ciprofloxacin-resistant Salmonella serotype Kentucky: A case report and the review of literature. Mikrobiyol. Bull. 2016, 50, 598–605. [Google Scholar]
- Bravo, D.; Silva, C.; Carter, J.A.; Hoare, A.; Alvarez, S.A.; Blondel, C.J.; Zaldívar, M.; Valvano, M.A.; Contreras, I. Growth-phase regulation of lipopolysaccharide O-antigen chain length influences serum resistance in serovars of Salmonella. J. Med. Microbiol. 2008, 57, 938–946. [Google Scholar] [CrossRef] [PubMed]
- Marshall, J.M.; Gunn, J.S. The O-antigen capsule of Salmonella enterica serovar Typhimurium facilitates serum resistance and surface expression of FliC. Infect. Immun. 2015, 83, 3946–3959. [Google Scholar] [CrossRef] [PubMed]
- Hart, P.J.; O’Shaughnessy, C.M.; Siggins, M.K.; Bobat, S.; Kingsley, R.A.; Goulding, D.A.; Crump, J.A.; Reyburn, H.; Micoli, F.; Dougan, G.; et al. Differential killing of Salmonella enterica serovar Typhi by antibodies targeting Vi and lipopolysaccharide O:9 antigen. PLoS ONE 2016, 11, e0145945. [Google Scholar] [CrossRef] [PubMed]
- Wanga, W.; Jeffery, C.J. An analysis of surface proteomics results reveals novel candidates for intracellular/surface moonlighting proteins in bacteria. Mol. BioSyst. 2016, 12, 1420–1431. [Google Scholar] [CrossRef] [PubMed]
- Seweryn, E.; Pietkiewicz, J.; Szamborska, A.; Gamian, A. Enolase on the surface of prockaryotic and eukaryotic cells is a receptor for human plasminogen. Post. Hig. Med. Dośw. 2007, 61, 672–682. [Google Scholar]
- Andrews, J.M. Determination of minimum inhibitory concentrations. J. Antimicrob. Chemother. 2001, 48 (Suppl. 1), 5–16. [Google Scholar] [CrossRef] [PubMed]
- European Committee on Antimicrobial Susceptibility Testing. 2015, pp. 1–78. Available online: http://www.eucast.org/.
- Ricci, V.; Tzakas, P.; Buckley, A.; Piddock, L.J.V. Ciprofloxacin-resistant Salmonella enterica serovar Typhimurium strains are difficult to select in the absence of AcrB and TolC. Antimicrob. Agents Chemother. 2006, 50, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Futoma-Kołoch, B.; Godlewska, U.; Bugla-Płoskońska, G.; Pawlak, A. Bacterial cell surface—Place where C3 complement activation occurs. 13th Conference Molecular biology in diagnostics of infectious diseases and biotechnology. Diag. Mol. 2012, 120–123. [Google Scholar]
- Bugla-Płoskońska, G.; Rybka, J.; Futoma-Kołoch, B.; Cisowska, A.; Gamian, A.; Doroszkiewicz, W. Sialic acid-containing lipopolysaccharides of Salmonella O48 strains-potential role in camouflage and susceptibility to the bactericidal effect of normal human serum. Microb. Ecol. 2010, 59, 601–613. [Google Scholar] [CrossRef] [PubMed]
- Doroszkiewicz, W. Mechanism of antigenic variation in Shigella flexneri bacilli. IV. Role of lipopolysaccharides and their components in the sensitivity of Shigella flexneri 1b and its Lac+ recombinant to killing action of serum. Arch. Immunol. Ther. Exp. 1997, 45, 235–242. [Google Scholar]
- Murphy, T.F.; Bartos, L.C. Surface-exposed and antigenically conserved determinants of outer membrane proteins of Branhamella catarrhalis. Infect. Immun. 1989, 57, 2938–2941. [Google Scholar] [PubMed]
- Bugla-Płoskońska, G.; Korzeniowska-Kowal, A.; Guz-Regner, K. Reptiles as a source of Salmonella O48-clinically important bacteria for children: The relationship between resistance to normal cord serum and outer membrane protein patterns. Microb. Ecol. 2011, 61, 41–51. [Google Scholar] [CrossRef] [PubMed]
- O’Farrell, P.H. High resolution two-dimensional electrophoresis of proteins. J. Biol. Chem. 1975, 250, 4007–4021. [Google Scholar] [PubMed]
- Bednarz-Misa, I.; Serek, P.; Dudek, B.; Pawlak, A.; Bugla-Płoskońska, G.; Gamian, A. Application of zwitterionic detergent to the solubilization of Klebsiella pneumoniae outer membrane proteins for two-dimensional gel electrophoresis. J. Microbiol. Methods 2014, 107, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Bugla-Płoskońska, G.; Futoma-Kołoch, B.; Skwara, A.; Doroszkiewicz, W. Use of zwitterionic type of detergent in isolation of Escherichia coli O56 outer membrane proteins improves their two-dimensional electrophoresis (2-DE). Pol. J. Microbiol. 2009, 58, 205–209. [Google Scholar] [PubMed]
- Shevchenko, A.; Tomas, H.; Havlis, J.; Olsen, J.V.; Mann, M. In-gel digestion for mass spectrometric characterization of proteins and proteomes. Nat. Protoc. 2006, 1, 2856–2860. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Time of Incubation | Concentration of Biocide | S. Senftenberg Strain | ||||
|---|---|---|---|---|---|---|
| 131 | 132 | 133 | 134 | 135 | ||
| 1-day preculture in LB broth | none | + | + | + | + | + |
| 7 days in Luria-Bertani (LB) broth | 0.5 × MIC | + 0.05 | + 0.2 | + 0.05 | + 0.05 | + 0.1 |
| Gradient 4 × 4 days in LB broth | 0.75 × MIC | + 0.075 | + 0.3 | + 0.075 | + 0.075 | + 0.15 |
| 1.0 × MIC | + 0.1 | + 0.4 | + 0.1 | + 0.1 | + 0.2 | |
| 1.25 × MIC | + 0.125 | + 0.5 | + 0.125 | + 0.125 | + 0.25 | |
| 1.5 × MIC | + 0.15 | + 0.6 | + 0.15 | + 0.15 | + 0.3 | |
| 1 day in LB broth | 2 × MIC | + 0.2 | + 0.8 | + 0.2 | + 0.2 | + 0.4 |
| 4 × MIC | + 0.4 | + 1.6 | + 0.4 | + 0.4 | + 0.8 | |
| 6 × MIC | − 0.6 | − 2.4 | + 0.6 | − 0.6 | + 1.2 | |
| 8 × MIC | − 0.8 | − 3.2 | − 0.8 | − 0.8 | − 1.6 | |
| Identification on Brilliant Green | from the highest MIC (where growth was observed) | + | + | + | + | + |
| Stability test 10 days in LB broth | none | + | + | + | + | + |
| Identification on Brilliant Green | none | + | + | + | + | + |
| Test | S. Senftenberg Strains | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 131 | 131 bST | 131 aST | 132 | 132 bST | 132 aST | 133 | 133 bST | 133 aST | 134 | 134 bST | 134 aST | 135 | 135 bST | 135 aST | |
| MIC (µL/mL) | 0.1 | 0.4 | 0.4 | 0.4 | 1.6 | 0.2 | 0.1 | 0.6 | 0.4 | 0.1 | 0.4 | 0.2 | 0.2 | 0.8 | 0.2 |
| MBC (µL/mL) | 0.1 | nt | 0.4 | 0.4 | nt | 0.4 | 0.1 | nt | 0.8 | 0.1 | nt | 0.2 | 0.4 | nt | 0.8 |
| Spots | Identified Proteins | Gene Symbols | Molecular Weight (kDa) | pI | Expression |
|---|---|---|---|---|---|
| 1 | Flagellin (FliC) | fliC | 52.081 | 4.85 | downregulated |
| 2 | Enolase | eno | 45.628 | 5.25 | downregulated |
| 3 | Chemotaxis response regulator protein-glutamate methylesterase | cheB | 37.498 | 8.48 | upregulated |
| 4 | Outer membrane protein assembly factor BamA | bamA | 89.525 | 4.92 | upregulated |
| No. | Strain | Maximal Tolerance to Biocide (See Table 1) | MIC (See Table 2) | RP in 22.5% NH (See Table S1) | RP in 45% NHS (See Table S2) | Comments |
|---|---|---|---|---|---|---|
| 131 | S. Senftenberg | 0.1 | R in T1 | S | Resistance of the variants is maintained | |
| 131bST | 4 × MIC | higher | R in T1 | R in T1 | ||
| 131aST | higher | R in T1 and T2 | R in T1 and T2 | |||
| 133 | S. Senftenberg | 0.1 | S | S | Resistance of the variant is maintained | |
| 133bST | 6 × MIC | higher | S | S | ||
| 133aST | higher | R in T1 | R in T1 | |||
| 135 | S. Senftenberg | R in T1 and T2 | R in T1 and T2 | Resistance of the variants to NHS is lost in both serum concentrations | ||
| 135bST | 6 × MIC | 0.2 | S | S | ||
| 135aST | the same | S | S | |||
| 132 | S. Senftenberg | 0.4 | R in T1 | R in T1 | Resistance of the variants to NHS is lost in higher serum concentration | |
| 132bST | 4 × MIC | higher | R in T2 | S | ||
| 132aST | lower | R in T1 | S | |||
| 134 | S. Senftenberg | 0.1 | R in T1 | R in T1 | Resistance of the variants is maintained | |
| 134bST | 4 × MIC | higher | R in T1 and T2 | R in T1 | ||
| 134aST | higher | R in T1 and T2 | R in T1 and T2 |
| Active Substances | Recommended Contact Time | Experimental Contact Time (See Table 1) | Recommended Working Concentration | Experimental Working Concentration | Mechanisms of Action Against Bacteria |
|---|---|---|---|---|---|
| triamine, 2-aminoethanol, cationic surfactants, nonionic surfactants | 5–10 min | 24 days | (2.5%) 2.5 mL/100 mL | From 5 µL/100 mL (0.005%) to 320 µL/100 mL (0.32%) | penetration of outer membrane of bacterial cell disrupting of RNA of the microorganism preventing of replication of DNA |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Futoma-Kołoch, B.; Dudek, B.; Kapczyńska, K.; Krzyżewska, E.; Wańczyk, M.; Korzekwa, K.; Rybka, J.; Klausa, E.; Bugla-Płoskońska, G. Relationship of Triamine-Biocide Tolerance of Salmonella enterica Serovar Senftenberg to Antimicrobial Susceptibility, Serum Resistance and Outer Membrane Proteins. Int. J. Mol. Sci. 2017, 18, 1459. https://doi.org/10.3390/ijms18071459
Futoma-Kołoch B, Dudek B, Kapczyńska K, Krzyżewska E, Wańczyk M, Korzekwa K, Rybka J, Klausa E, Bugla-Płoskońska G. Relationship of Triamine-Biocide Tolerance of Salmonella enterica Serovar Senftenberg to Antimicrobial Susceptibility, Serum Resistance and Outer Membrane Proteins. International Journal of Molecular Sciences. 2017; 18(7):1459. https://doi.org/10.3390/ijms18071459
Chicago/Turabian StyleFutoma-Kołoch, Bożena, Bartłomiej Dudek, Katarzyna Kapczyńska, Eva Krzyżewska, Martyna Wańczyk, Kamila Korzekwa, Jacek Rybka, Elżbieta Klausa, and Gabriela Bugla-Płoskońska. 2017. "Relationship of Triamine-Biocide Tolerance of Salmonella enterica Serovar Senftenberg to Antimicrobial Susceptibility, Serum Resistance and Outer Membrane Proteins" International Journal of Molecular Sciences 18, no. 7: 1459. https://doi.org/10.3390/ijms18071459
APA StyleFutoma-Kołoch, B., Dudek, B., Kapczyńska, K., Krzyżewska, E., Wańczyk, M., Korzekwa, K., Rybka, J., Klausa, E., & Bugla-Płoskońska, G. (2017). Relationship of Triamine-Biocide Tolerance of Salmonella enterica Serovar Senftenberg to Antimicrobial Susceptibility, Serum Resistance and Outer Membrane Proteins. International Journal of Molecular Sciences, 18(7), 1459. https://doi.org/10.3390/ijms18071459