
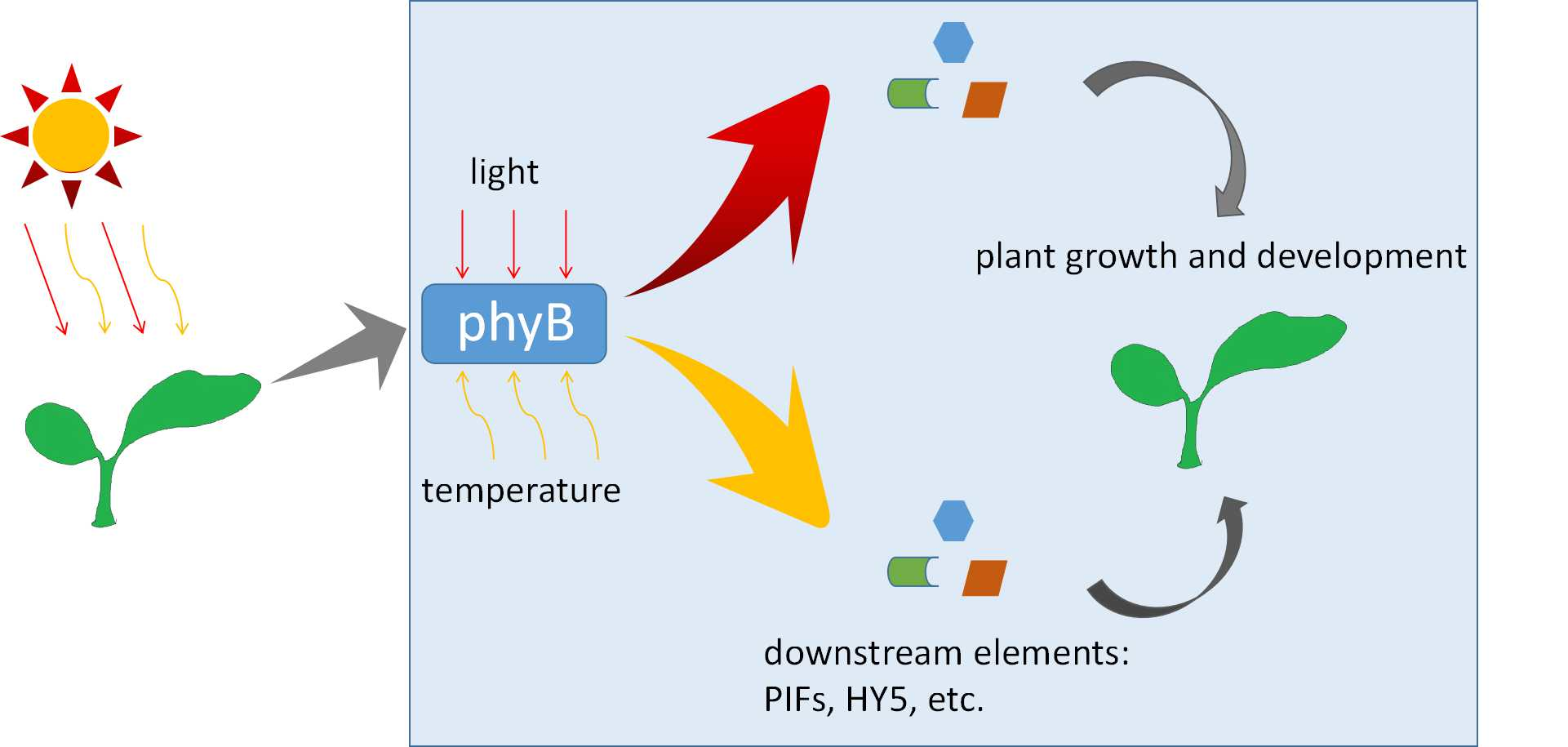
Photoreceptor PhyB Involved in Arabidopsis Temperature Perception and Heat-Tolerance Formation

Abstract
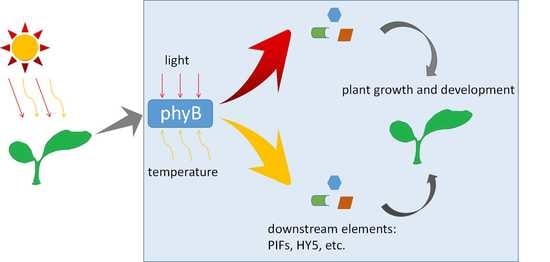
:
1. Introduction
2. Results and Discussion
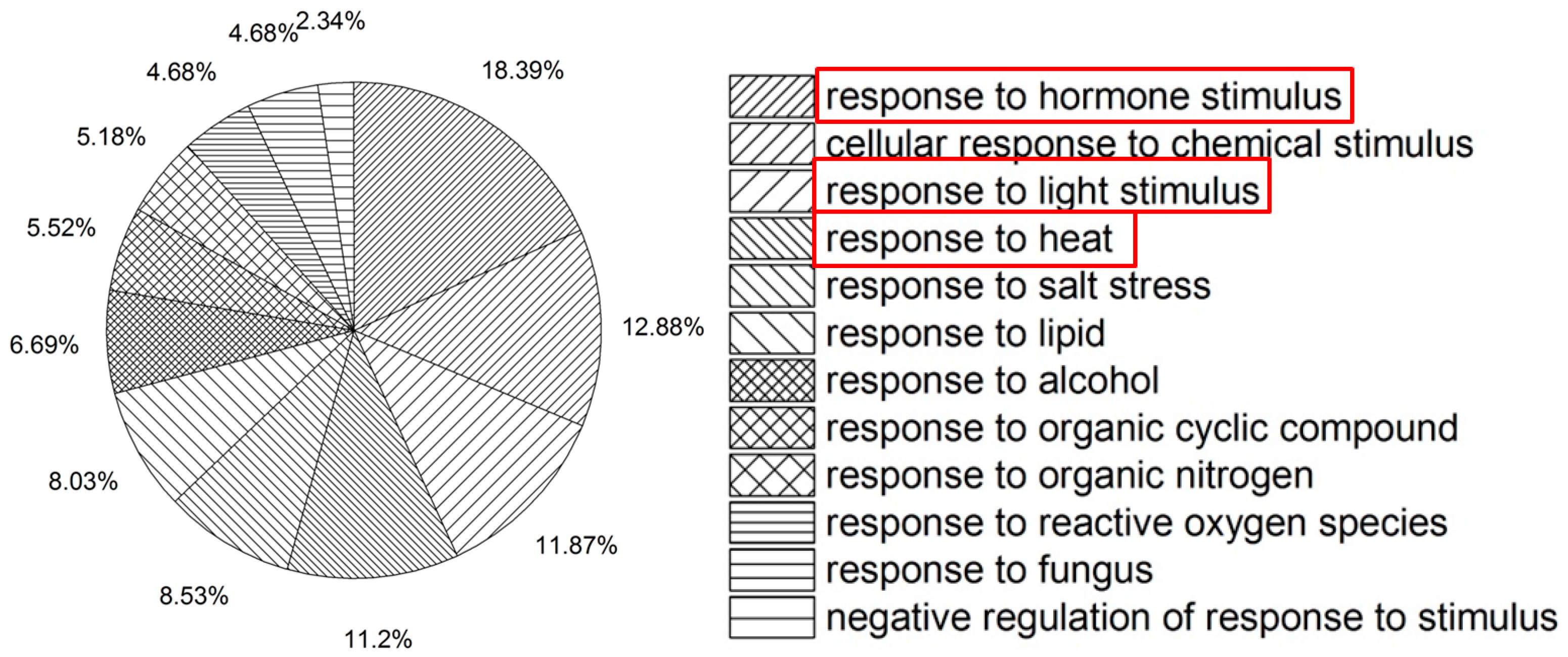
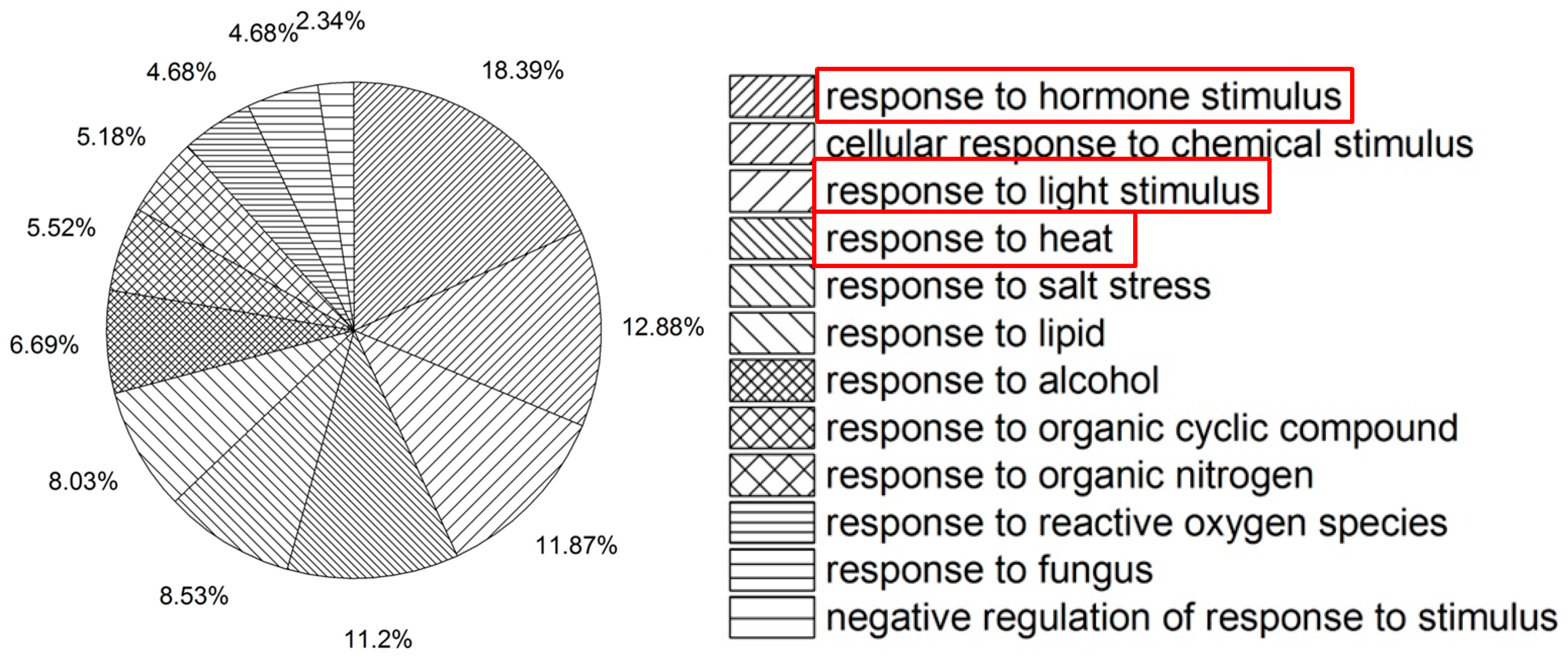
2.1. Microarray Analysis of col-0 under Heat Stress
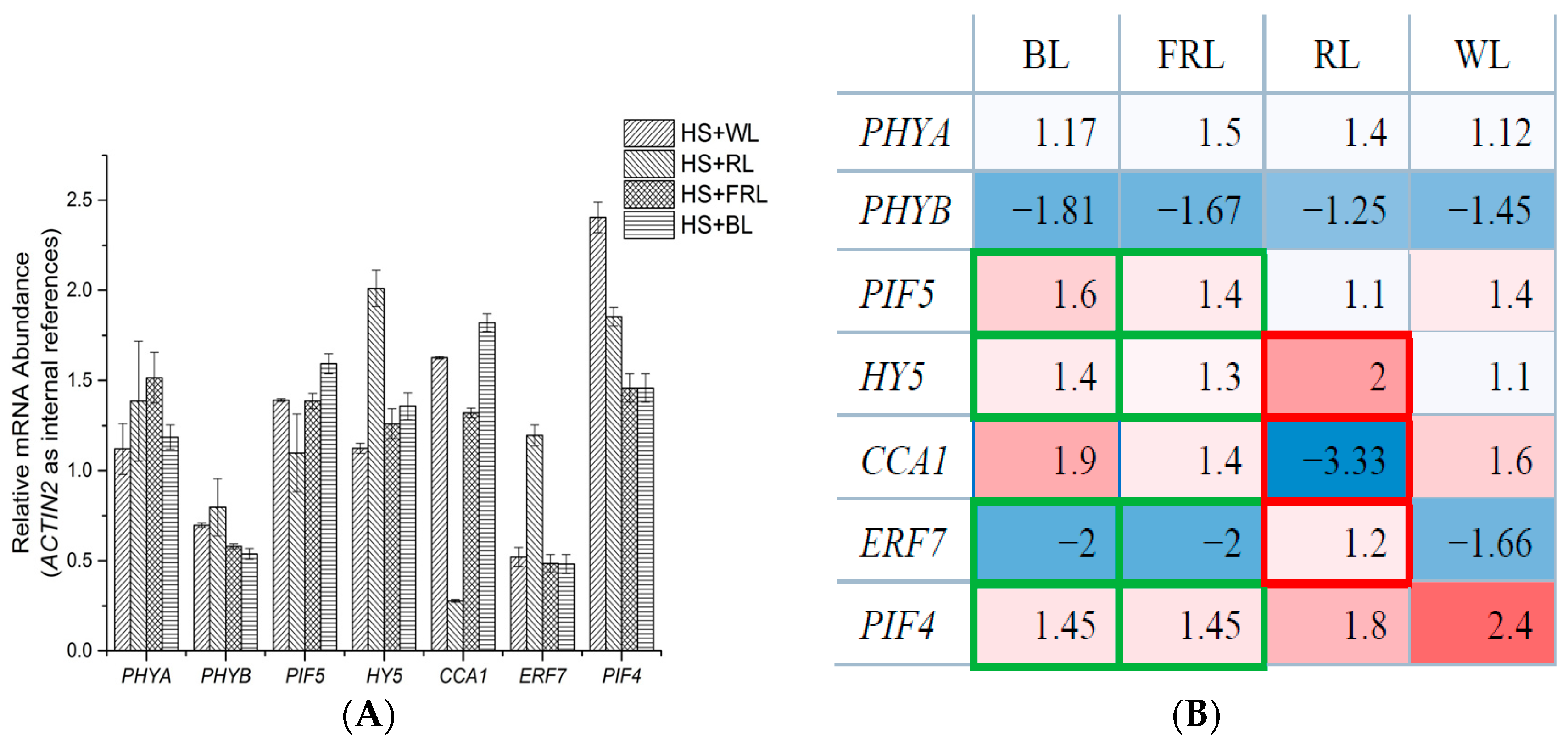
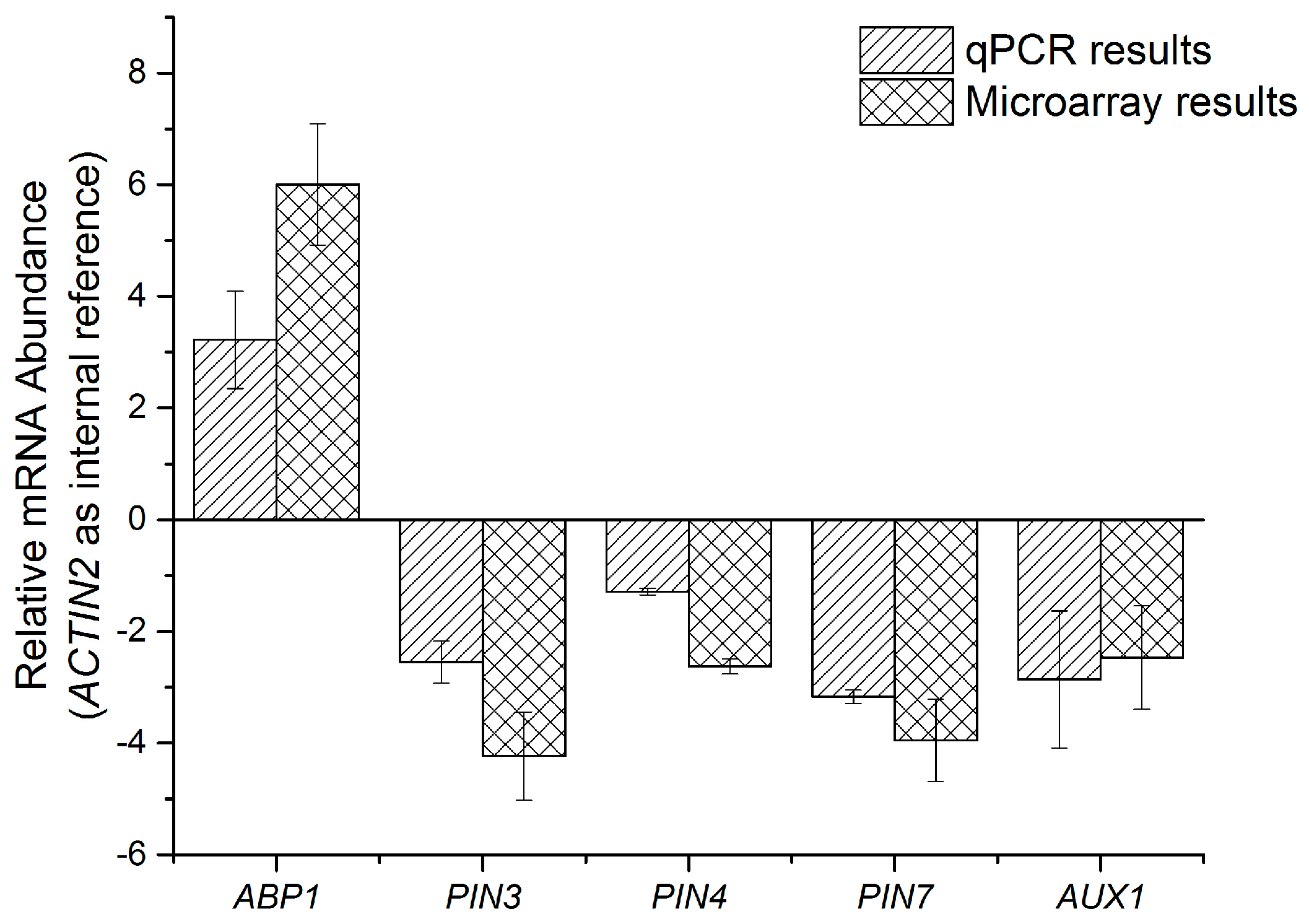
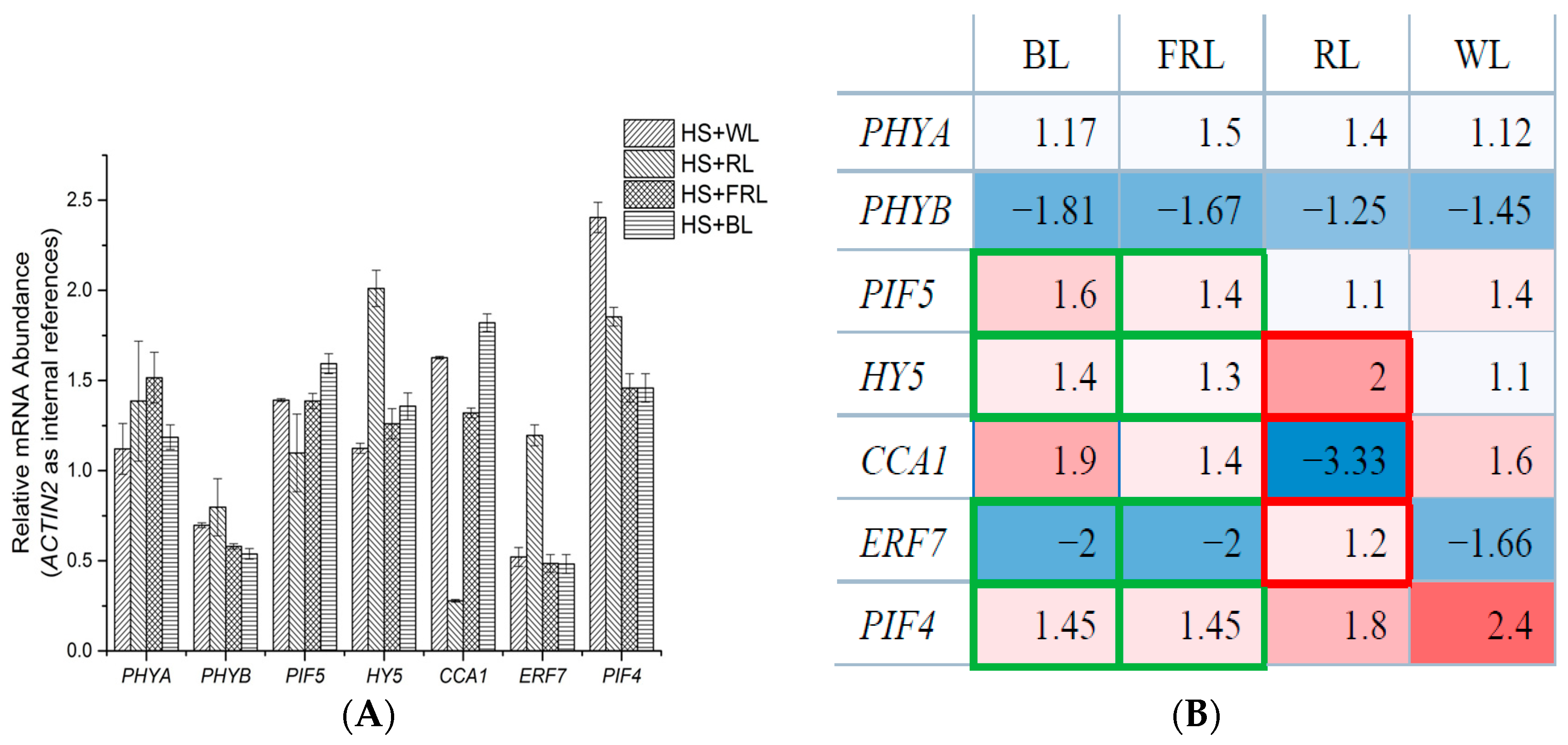
2.2. qPCR Analysis of Transcriptomic Response to HS under Light Conditions
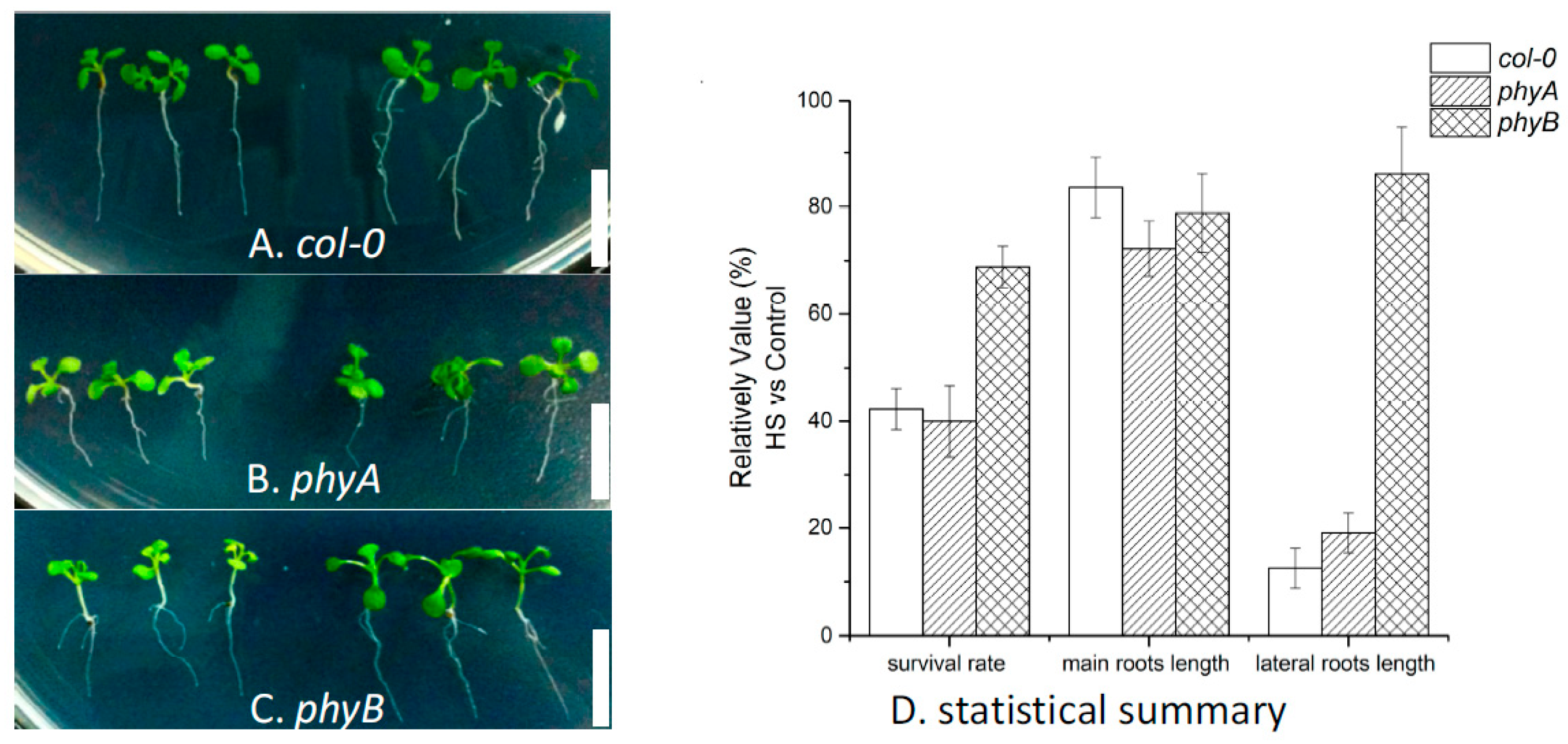
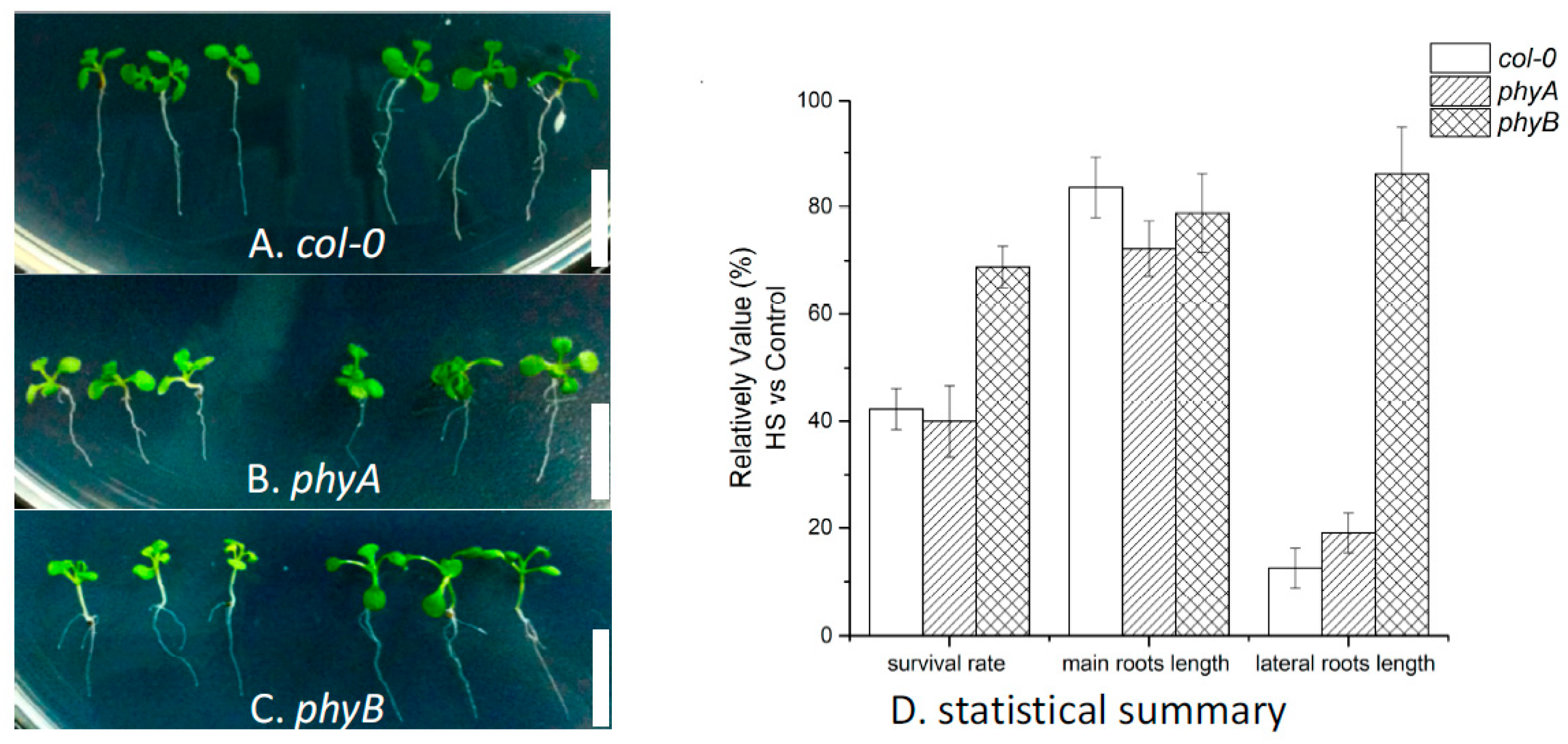
2.3. Effects of HS on Different Arabidopsis Genotypes
3. Materials and Methods
3.1. Plant Materials and Growth Conditions
3.2. Heat Stress Treatment
3.3. Light Conditions Adopted during HS Treatment
3.4. Microarray Hybridization and Analysis
3.5. qPCR Analysis
4. Conclusions
Supplementary Materials
Acknowledgment
Author Contributions
Conflicts of Interest
References
- Nakashima, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K. The transcriptional regulatory network in the drought response and its crosstalk in abiotic stress responses including drought, cold, and heat. Front. Plant Sci. 2014, 5, 170. [Google Scholar] [CrossRef] [PubMed]
- Maibam, P.; Nawkar, G.M.; Park, J.H.; Sahi, V.P.; Lee, S.Y.; Kang, C.H. The influence of light quality, circadian rhythm, and photoperiod on the CBF-mediated freezing tolerance. Int. J. Mol. Sci. 2013, 14, 11527–11543. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Huang, L.; Hong, Y.; Zhang, H.; Song, F.; Li, D. Comprehensive analysis suggests overlapping expression of rice onac transcription factors in abiotic and biotic stress responses. Int. J. Mol. Sci. 2015, 16, 4306–4326. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.-Y.; Lin, L.-F.; Jheng, J.-L.; Wang, C.-C.; Yang, J.-H.; Chou, M.-L. Identification of heat shock transcription factor genes involved in thermotolerance of octoploid cultivated strawberry. Int. J. Mol. Sci. 2016, 17, 2130. [Google Scholar] [CrossRef] [PubMed]
- Vinterhalter, D.; Vinterhalter, B. Phototropic responses of potato under conditions of continuous light and subsequent darkness. Plant Growth Regul. 2015, 75, 725–732. [Google Scholar] [CrossRef]
- McWatters, H.G.; Devlin, P.F. Timing in plants—A rhythmic arrangement. FEBS Lett. 2011, 585, 1474–1484. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, N. Cryptochromes and seed dormancy: The molecular mechanism of blue light inhibition of grain germination. Plant Cell 2014, 26, 846. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, Y.; Sumitomo, K.; Oda, A.; Shimizu, H.; Hisamatsu, T. Day light quality affects the night-break response in the short-day plant chrysanthemum, suggesting differential phytochrome-mediated regulation of flowering. J. Plant Physiol. 2012, 169, 1789–1796. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.W.; Huang, X.H.; Wang, Y.R. Hormonal and temperature regulation of seed dormancy and germination in leymus chinensis. Plant Growth Regul. 2012, 67, 199–207. [Google Scholar] [CrossRef]
- Xin, C.; Wang, X.; Cai, J.; Zhou, Q.; Liu, F.; Dai, T.; Cao, W.; Jiang, D. Changes of transcriptome and proteome are associated with the enhanced post-anthesis high temperature tolerance induced by pre-anthesis heat priming in wheat. Plant Growth Regul. 2015, 1–11. [Google Scholar] [CrossRef]
- Mittler, R.; Finka, A.; Goloubinoff, P. How do plants feel the heat? Trends Biochem. Sci. 2012, 37, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef] [PubMed]
- Zinn, K.E.; Tunc-Ozdemir, M.; Harper, J.F. Temperature stress and plant sexual reproduction: Uncovering the weakest links. J. Exp. Bot. 2010, 61, 1959–1968. [Google Scholar] [CrossRef] [PubMed]
- Kosova, K.; Vitamvas, P.; Urban, M.O.; Klima, M.; Roy, A.; Prasil, I.T. Biological networks underlying abiotic stress tolerance in temperate crops—A proteomic perspective. Int. J. Mol. Sci. 2015, 16, 20913–20942. [Google Scholar] [CrossRef] [PubMed]
- Miura, K.; Furumoto, T. Cold signaling and cold response in plants. Int. J. Mol. Sci. 2013, 14, 5312–5337. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.; Barah, P.; Suarez-Rodriguez, M.C.; Bressendorff, S.; Friis, P.; Costantino, P.; Bones, A.M.; Nielsen, H.B.; Mundy, J. Transcriptome responses to combinations of stresses in Arabidopsis. Plant Physiol. 2013, 161, 1783–1794. [Google Scholar] [CrossRef] [PubMed]
- Bita, C.E.; Gerats, T. Plant tolerance to high temperature in a changing environment: Scientific fundamentals and production of heat stress-tolerant crops. Front. Plant Sci. 2013, 4, 273. [Google Scholar] [CrossRef] [PubMed]
- Hahn, A.; Kilian, J.; Mohrholz, A.; Ladwig, F.; Peschke, F.; Dautel, R.; Harter, K.; Berendzen, K.W.; Wanke, D. Plant core environmental stress response genes are systemically coordinated during abiotic stresses. Int. J. Mol. Sci. 2013, 14, 7617–7641. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Liu, Q.; Hu, B.; Wu, W. Comparative transcriptome profiling of Arabidopsis Col-0 in responses to heat stress under different light conditions. Plant Growth Regul. 2015, 79, 209–218. [Google Scholar] [CrossRef]
- Liu, H.C.; Charng, Y.Y. Common and distinct functions of Arabidopsis class A1 and A2 heat shock factors in diverse abiotic stress responses and development. Plant Physiol. 2013, 163, 276–290. [Google Scholar] [CrossRef] [PubMed]
- Joseph, M.P.; Papdi, C.; Kozma-Bognár, L.; Nagy, I.; López-Carbonell, M.; Rigó, G.; Koncz, C.; Szabados, L. The Arabidopsis zinc finger protein3 interferes with abscisic acid and light signaling in seed germination and plant development. Plant Physiol. 2014, 165, 1203–1220. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Gao, Z.; Luan, W. Interaction between temperature and photoperiod in regulation of flowering time in rice. Sci. China Life Sci. 2012, 55, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Thines, B.C.; Youn, Y.; Duarte, M.I.; Harmon, F.G. The time of day effects of warm temperature on flowering time involve PIF4 and PIF5. J. Exp. Bot. 2014, 65, 1141–1151. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Chory, J. Phytochrome signaling mechanisms and the control of plant development. Trends Cell Biol. 2011, 21, 664–671. [Google Scholar] [CrossRef] [PubMed]
- Franklin, K.A.; Quail, P.H. Phytochrome functions in Arabidopsis development. J. Exp. Bot. 2010, 61, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Legris, M.; Klose, C.; Burgie, E.S.; Rojas, C.C.R.; Neme, M.; Hiltbrunner, A.; Wigge, P.A.; Schäfer, E.; Vierstra, R.D.; Casal, J.J. Phytochrome B integrates light and temperature signals in Arabidopsis. Science 2016, 354, 897–900. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.-H.; Domijan, M.; Klose, C.; Biswas, S.; Ezer, D.; Gao, M.; Khattak, A.K.; Box, M.S.; Charoensawan, V.; Cortijo, S. Phytochromes function as thermosensors in Arabidopsis. Science 2016. [Google Scholar] [CrossRef] [PubMed]
- Song, A.; Zhu, X.; Chen, F.; Gao, H.; Jiang, J.; Chen, S. A chrysanthemum heat shock protein confers tolerance to abiotic stress. Int. J. Mol. Sci. 2014, 15, 5063–5078. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ji, X.; Zheng, L.; Nie, X.; Wang, Y. Microarray analysis of transcriptional responses to abscisic acid and salt stress in Arabidopsis thaliana. Int. J. Mol. Sci. 2013, 14, 9979–9998. [Google Scholar] [CrossRef] [PubMed]
- Montero-Barrientos, M.; Hermosa, R.; Cardoza, R.E.; Gutierrez, S.; Nicolas, C.; Monte, E. Transgenic expression of the Trichoderma harzianum hsp70 gene increases Arabidopsis resistance to heat and other abiotic stresses. J Plant Physiol. 2010, 167, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.A.; Bianchetti, R.E.; Freschi, L. Shedding light on ethylene metabolism in higher plants. Front. Plant Sci. 2014, 5, 665. [Google Scholar] [CrossRef] [PubMed]
- Jumtee, K.; Okazawa, A.; Harada, K.; Fukusaki, E.; Takano, M.; Kobayashi, A. Comprehensive metabolite profiling of phya phyb phyc triple mutants to reveal their associated metabolic phenotype in rice leaves. J. Biosci. Bioeng. 2009, 108, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Lucas, S.; Durmaz, E.; Akpinar, B.A.; Budak, H. The drought response displayed by a dre-binding protein from triticum dicoccoides. Plant Physiol. Biochem. 2011, 49, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Tavakol, E.; Sardaro, M.L.S.; Shariati, V.; Rossini, L.; Porceddu, E. Isolation, promoter analysis and expression profile of Dreb2 in response to drought stress in wheat ancestors. Gene 2014, 549, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Ergen, N.Z.; Thimmapuram, J.; Bohnert, H.J.; Budak, H. Transcriptome pathways unique to dehydration tolerant relatives of modern wheat. Funct. Integr. Genom. 2009, 9, 377–396. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Rashotte, A.M.; Dane, F. Citrullus colocynthis NAC transcription factors CcNAC1 and CcNAC2 are involved in light and auxin signaling. Plant Cell Rep. 2014, 33, 1673–1686. [Google Scholar] [CrossRef] [PubMed]
- Budak, H.; Hussain, B.; Khan, Z.; Ozturk, N.Z.; Ullah, N. From genetics to functional genomics: Improvement in drought signaling and tolerance in wheat. Front. Plant Sci. 2015, 6, 1012. [Google Scholar] [CrossRef] [PubMed]
- Qu, A.L.; Ding, Y.F.; Jiang, Q.; Zhu, C. Molecular mechanisms of the plant heat stress response. Biochem. Biophys. Res. Commun. 2013, 432, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Wabnik, K.; Govaerts, W.; Friml, J.; Kleine-Vehn, J. Feedback models for polarized auxin transport: An emerging trend. Mol. Biosyst. 2011, 7, 2352–2359. [Google Scholar] [CrossRef] [PubMed]
- Vandenbussche, F.; Petrášek, J.; Žádníková, P.; Hoyerová, K.; Pešek, B.; Raz, V.; Swarup, R.; Bennett, M.; Zažímalová, E.; Benková, E. The auxin influx carriers AUX1 and LAX3 are involved in auxin-ethylene interactions during apical hook development in Arabidopsis thaliana seedlings. Development 2010, 137, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Grandont, L.; Li, H.; Hauschild, R.; Paque, S.; Abuzeineh, A.; Rakusova, H.; Benkova, E.; Perrot-Rechenmann, C.; Friml, J. Inhibition of cell expansion by rapid abp1-mediated auxin effect on microtubules. Nature 2014, 516, 90–93. [Google Scholar] [CrossRef] [PubMed]
- Sasaki-Sekimoto, Y.; Jikumaru, Y.; Obayashi, T.; Saito, H.; Masuda, S.; Kamiya, Y.; Ohta, H.; Shirasu, K. Bhlh transcription factors ja-associated MYC2-like 1, JAM2 and JAM3 are negative regulators of jasmonate responses in Arabidopsis. Plant Physiol. 2013. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Huang, R. Enhanced tolerance to freezing in tobacco and tomato overexpressing transcription factor terf2/leerf2 is modulated by ethylene biosynthesis. Plant Mol. Biol. 2010, 73, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, R.F.; Campos, M.L.; Azevedo, R.A. The role of phytochrome in stress tolerance. J. Integr. Plant Biol. 2011, 53, 920–929. [Google Scholar] [CrossRef] [PubMed]
- Leivar, P.; Monte, E.; Al-Sady, B.; Carle, C.; Storer, A.; Alonso, J.M.; Ecker, J.R.; Quail, P.H. The Arabidopsis phytochrome-interacting factor PIF7, together with PIF3 and PIF4, regulates responses to prolonged red light by modulating phyb levels. Plant Cell Online 2008, 20, 337–352. [Google Scholar] [CrossRef] [PubMed]
- Pedmale, U.V.; Huang, S.-S.C.; Zander, M.; Cole, B.J.; Hetzel, J.; Ljung, K.; Reis, P.A.; Sridevi, P.; Nito, K.; Nery, J.R. Cryptochromes interact directly with PIFs to control plant growth in limiting blue light. Cell 2016, 164, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Karayekov, E.; Sellaro, R.; Legris, M.; Yanovsky, M.J.; Casal, J.J. Heat shock-induced fluctuations in clock and light signaling enhance phytochrome B-mediated Arabidopsis deetiolation. Plant Cell 2013, 25, 2892–2906. [Google Scholar] [CrossRef] [PubMed]
- Shikata, H.; Hanada, K.; Ushijima, T.; Nakashima, M.; Suzuki, Y.; Matsushita, T. Phytochrome controls alternative splicing to mediate light responses in Arabidopsis. Proc. Natl. Acad. Sci. USA 2014, 111, 18781–18786. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.B.; Lian, H.L.; Wang, W.X.; Xu, F.; Yang, H.Q. Pivotal roles of the phytochrome-interacting factors in cryptochrome signaling. Mol. Plant 2016, 9, 496–497. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Guo, Z.; Li, H.; Wang, M.; Onac, E.; Zhou, J.; Xia, X.; Shi, K.; Yu, J.; Zhou, Y. Phytochrome A and B function antagonistically to regulate cold tolerance via abscisic acid-dependent jasmonate signaling. Plant Physiol. 2016, 170, 459–471. [Google Scholar] [CrossRef] [PubMed]
- Koini, M.A.; Alvey, L.; Allen, T.; Tilley, C.A.; Harberd, N.P.; Whitelam, G.C.; Franklin, K.A. High temperature-mediated adaptations in plant architecture require the bhlh transcription factor PIF4. Curr. Biol. 2009, 19, 408–413. [Google Scholar] [CrossRef] [PubMed]
- Stavang, J.A.; Gallego-Bartolome, J.; Gomez, M.D.; Yoshida, S.; Asami, T.; Olsen, J.E.; Garcia-Martinez, J.L.; Alabadi, D.; Blazquez, M.A. Hormonal regulation of temperature-induced growth in Arabidopsis. Plant J. Cell Mol. Biol. 2009, 60, 589–601. [Google Scholar] [CrossRef] [PubMed]
- Legris, M.; Nieto, C.; Sellaro, R.; Prat, S.; Casal, J.J. Perception and signalling of light and temperature cues in plants. Plant J. Cell Mol. Biol. 2017, 90, 683–697. [Google Scholar] [CrossRef] [PubMed]
- Gangappa, S.N.; Kumar, S.V. DET1 and HY5 control PIF4-mediated thermosensory elongation growth through distinct mechanisms. Cell Rep. 2017, 18, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.K.; Senapati, D.; Srivastava, A.; Chakraborty, M.; Gangappa, S.N.; Chattopadhyay, S. Short hypocotyl in white light1 interacts with elongated hypocotyl5 (HY5) and constitutive photomorphogenic1 (COP1) and promotes COP1-mediated degradation of HY5 during Arabidopsis seedling development. Plant Physiol. 2015, 169, 2922–2934. [Google Scholar] [PubMed]
- Nawkar, G.M.; Kang, C.H.; Maibam, P.; Park, J.H.; Jung, Y.J.; Chae, H.B.; Chi, Y.H.; Jung, I.J.; Kim, W.Y.; Yun, D.-J. HY5, a positive regulator of light signaling, negatively controls the unfolded protein response in Arabidopsis. Proc. Natl. Acad. Sci. USA 2017, 201609844. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Gene Title | Fold Change |
|---|---|---|
| 1 | TEM1 (TEMPRANILLO 1); transcription factor | −4.88839 |
| 2 | RGA1 (REPRESSOR OF GA1-3 1); protein binding/transcription factor | −2.82099 |
| 3 | GAI (GIBBERELLIC ACID INSENSITIVE); transcription factor | −2.27883 |
| 4 | PAT1 (phytochrome a signal transduction 1); signal transducer/transcription factor | −2.02137 |
| 5 | HY5 (ELONGATED HYPOCOTYL 5); double-stranded DNA binding/transcription factor | 2.29793 |
| 6 | NAC2; transcription factor | 3.5822 |
| 7 | MYB4; DNA binding/transcription factor | 3.60975 |
| 8 | ANAC13 (Arabidopsis thaliana NAC domain protein 13); transcription factor | 3.86913 |
| 9 | GA3OX1 (GIBBERELLIN 3-OXIDASE 1); gibberellin 3-β-dioxygenase/transcription factor | 4.73567 |
| 10 | ATNFXL1 (Arabidopsis thaliana NF-X-LIKE 1); protein binding/transcription factor/zin | 5.01115 |
| 11 | DREB2A; DNA binding/transcription activator/transcription factor | 16.786 |
| 12 | AT-HSFA7A; DNA binding/transcription factor | 97.0821 |
| 13 | AT-HSFA7B; DNA binding/transcription factor | 121.056 |
| 14 | ATHSFA2; DNA binding/transcription factor | 248.051 |
| No. | Gene Title | Fold Change |
|---|---|---|
| 1 | MYC2; DNA binding/transcription activator/transcription factor | −11.9908 |
| 2 | BT2 (BTB AND TAZ DOMAIN PROTEIN 2); protein binding/transcription factor | −8.87107 |
| 3 | ERF11 (ERF DOMAIN PROTEIN 11); DNA binding/transcription factor/transcription repressor | −8.04081 |
| 4 | MYB73 (MYB DOMAIN PROTEIN 73); DNA binding/transcription factor | −7.82007 |
| 5 | TEM1 (TEMPRANILLO 1); transcription factor | −4.88839 |
| 6 | RAV1; DNA binding/transcription factor/transcription repressor | −4.52717 |
| 7 | ATAIB (ABA-INDUCIBLE BHLH-TYPE TRANSCRIPTION FACTOR); DNA binding/transcription factor | −4.26839 |
| 8 | AS1 (ASYMMETRIC LEAVES 1); DNA binding/protein homodimerization/transcription factor | −3.37803 |
| 9 | MYBL2 (Arabidopsis MYB-LIKE 2); DNA binding/transcription factor | −3.15927 |
| 10 | MYB28 (MYB DOMAIN PROTEIN 28); DNA binding/transcription factor | −3.15868 |
| 11 | ERF8; DNA binding/transcription factor/transcription repressor | −3.03177 |
| 12 | RGA1 (REPRESSOR OF GA1-3 1); protein binding/transcription factor | −2.82099 |
| 13 | AtTCP14 (TEOSINTE BRANCHED1, CYCLOIDEA and PCF (TCP) 14); transcription factor | −2.73614 |
| 14 | ERF7 (ETHYLINE RESPONSE FACTOR 7); DNA binding/protein binding/transcription factor | −2.47281 |
| 15 | EIL1 (ETHYLENE-INSENSITIVE3-LIKE 1); transcription factor/transcription regulator | −2.37976 |
| 16 | GAI (GIBBERELLIC ACID INSENSITIVE); transcription factor | -2.27883 |
| 17 | AtMYB47 (MYB DOMAIN PROTEIN 47); DNA binding/transcription factor | -2.14196 |
| 18 | RAP2.7 (RELATED TO AP2.7); DNA binding/transcription factor | -2.10654 |
| 19 | IAA2 (INDOLE-3-ACETIC ACID INDUCIBLE 2); transcription factor | -2.09408 |
| 20 | HY5 (ELONGATED HYPOCOTYL 5); double-stranded DNA binding/transcription factor | 2.29793 |
| 21 | CRF6 (CYTOKININ RESPONSE FACTOR 6); DNA binding/transcription factor | 2.87763 |
| 22 | MYB4; DNA binding/transcription factor | 3.60975 |
| 23 | GA3OX1 (GIBBERELLIN 3-OXIDASE 1); gibberellin 3-β-dioxygenase/transcription factor | 4.73567 |
| 24 | MYB43 (MYB DOMAIN PROTEIN 43); DNA binding/transcription factor | 4.94925 |
| 25 | EPR1 (EARLY-PHYTOCHROME-RESPONSIVE1); DNA binding/transcription factor | 7.39638 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, J.; Liu, Q.; Hu, B.; Wu, W. Photoreceptor PhyB Involved in Arabidopsis Temperature Perception and Heat-Tolerance Formation. Int. J. Mol. Sci. 2017, 18, 1194. https://doi.org/10.3390/ijms18061194
Song J, Liu Q, Hu B, Wu W. Photoreceptor PhyB Involved in Arabidopsis Temperature Perception and Heat-Tolerance Formation. International Journal of Molecular Sciences. 2017; 18(6):1194. https://doi.org/10.3390/ijms18061194
Chicago/Turabian StyleSong, Junyi, Qijun Liu, Biru Hu, and Wenjian Wu. 2017. "Photoreceptor PhyB Involved in Arabidopsis Temperature Perception and Heat-Tolerance Formation" International Journal of Molecular Sciences 18, no. 6: 1194. https://doi.org/10.3390/ijms18061194
APA StyleSong, J., Liu, Q., Hu, B., & Wu, W. (2017). Photoreceptor PhyB Involved in Arabidopsis Temperature Perception and Heat-Tolerance Formation. International Journal of Molecular Sciences, 18(6), 1194. https://doi.org/10.3390/ijms18061194