
The Importance of Pericytes in Healing: Wounds and other Pathologies



Abstract
:1. An Introduction to Pericytes
1.1. Morphology, Location and Function
1.2. Pericyte Origin and MSC Properties
2. Pericytes in Wound Healing
2.1. Pericytes and Inflammation
2.2. Pericytes and Re-Epithelialisation
2.3. Pericytes and Angiogenesis
2.4. Pericytes and Matrix Deposition/Fibrosis
3. Pericytes in Other Pathologies
4. Pericytes as a Therapeutic Agent
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Sims, D.E. The pericyte—A review. Tissue Cell 1986, 18, 153–174. [Google Scholar] [CrossRef]
- Mills, S.J.; Cowin, A.J.; Kaur, P. Pericytes, mesenchymal stem cells and the wound healing process. Cells 2013, 2, 621–634. [Google Scholar] [CrossRef] [PubMed]
- Rucker, H.K.; Wynder, H.J.; Thomas, W.E. Cellular mechanisms of cns pericytes. Brain Res. Bull. 2000, 51, 363–369. [Google Scholar] [CrossRef]
- Stratman, A.N.; Malotte, K.M.; Mahan, R.D.; Davis, M.J.; Davis, G.E. Pericyte recruitment during vasculogenic tube assembly stimulates endothelial basement membrane matrix formation. Blood 2009, 114, 5091–5101. [Google Scholar] [CrossRef] [PubMed]
- Tilton, R.G.; Kilo, C.; Williamson, J.R. Pericyte-Endothelial relationships in cardiac and skeletal-muscle capillaries. Microvasc. Res. 1979, 18, 325–335. [Google Scholar] [CrossRef]
- Armulik, A.; Genové, G.; Betsholtz, C. Pericytes: Developmental, physiological, and pathological perspectives, problems, and promises. Dev. Cell 2011, 21, 193–215. [Google Scholar] [CrossRef] [PubMed]
- Geevarghese, A.; Herman, I.M. Pericyte-endothelial cross-talk: Implications and opportunities for advanced cellular therapies. Transl. Res. 2014, 163, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Gaengel, K.; Genové, G.; Armulik, A.; Betsholtz, C. Endothelial-Mural cell signaling in vascular development and angiogenesis. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 630. [Google Scholar] [CrossRef] [PubMed]
- Proebstl, D.; Voisin, M.-B.; Woodfin, A.; Whiteford, J.; D’Acquisto, F.; Jones, G.E.; Rowe, D.; Nourshargh, S. Pericytes support neutrophil subendothelial cell crawling and breaching of venular walls in vivo. J. Exp. Med. 2012, 209, 1219–1234. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Cao, C.; Chen, Z.; Bankaitis, V.; Tzima, E.; Sheibani, N.; Burridge, K. Pericytes regulate vascular basement membrane remodeling and govern neutrophil extravasation during inflammation. PLoS ONE 2012, 7, e45499. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-W.; Okada, M.; Proto, J.D.; Gao, X.; Sekiya, N.; Beckman, S.A.; Corselli, M.; Crisan, M.; Saparov, A.; Tobita, K.; et al. Human pericytes for ischemic heart repair. Stem Cells 2013, 31, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Wilm, B.; Ipenberg, A.; Hastie, N.D.; Burch, J.B.E.; Bader, D.M. The serosal mesothelium is a major source of smooth muscle cells of the gut vasculature. Development 2005, 132, 5317. [Google Scholar] [CrossRef] [PubMed]
- Que, J.; Wilm, B.; Hasegawa, H.; Wang, F.; Bader, D.; Hogan, B.L.M. Mesothelium contributes to vascular smooth muscle and mesenchyme during lung development. Proc. Natl. Acad. Sci. USA 2008, 105, 16626–16630. [Google Scholar] [CrossRef] [PubMed]
- Asahina, K.; Zhou, B.; Pu, W.T.; Tsukamoto, H. Septum transversum-derived mesothelium gives rise to hepatic stellate cells and perivascular mesenchymal cells in developing mouse liver. Hepatology 2011, 53, 983–995. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.L.; Martin, J.C.; Sun, Y.; Cui, L.; Wang, L.; Ouyang, K.; Yang, L.; Bu, L.; Liang, X.; Zhang, X.; et al. A myocardial lineage derives from tbx18 epicardial cells. Nature 2008, 454, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Wessels, A.; Pérez-Pomares, J.M. The epicardium and epicardially derived cells (EPDCs) as cardiac stem cells. Anat. Rec. A Discov. Mol. Cell Evol. Biol. 2004, 276A, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Etchevers, H.C.; Vincent, C.; Le Douarin, N.M.; Couly, G.F. The cephalic neural crest provides pericytes and smooth muscle cells to all blood vessels of the face and forebrain. Development 2001, 128, 1059. [Google Scholar] [PubMed]
- Díaz-Flores, L.; Gutiérrez, R.; Madrid, J.F.; Varela, H.; Valladares, F.; Acosta, E.; Martín-Vasallo, P.; Díaz-Flores, L. Pericytes. Morphofunction, interactions and pathology in a quiescent and activated mesenchymal cell niche. Histol. Histopathol. 2009, 24, 909–969. [Google Scholar] [PubMed]
- Bergers, G.; Song, S. The role of pericytes in blood-vessel formation and maintenance. Neuro Oncol. 2005, 7, 452–464. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, C.G.M.; Nieuweboer, F.E.; Pei, J.Y.; Xu, Y.J.; Burgisser, P.; van Mulligen, E.; el Azzouzi, H.; Duncker, D.J.; Verhaar, M.C.; Cheng, C. The complex mural cell: Pericyte function in health and disease. Int. J. Cardiol. 2015, 190, 75–89. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.-P.; Rowley, J.E.; Redpath, A.N.; Tilman, J.D.; Fellous, T.G.; Johnson, J.R. Pericytes, mesenchymal stem cells and their contributions to tissue repair. Pharmacol. Ther. 2015, 151, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Ansell, D.M.; Izeta, A. Pericytes in wound healing: Friend or foe? Exp. Dermatol. 2015, 24, 833–834. [Google Scholar] [CrossRef] [PubMed]
- Corselli, M.; Chen, C.-W.; Crisan, M.; Lazzari, L.; Péault, B. Perivascular ancestors of adult multipotent stem cells. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1104. [Google Scholar] [CrossRef] [PubMed]
- V Nehls, D.D. Heterogeneity of microvascular pericytes for smooth muscle type α-actin. J. Cell Biol. 1991, 113, 147–154. [Google Scholar] [CrossRef]
- Chen, W.C.W.; Baily, J.E.; Corselli, M.; Diaz, M.; Sun, B.; Xiang, G.; Gray, G.A.; Huard, J.; Péault, B. Human myocardial pericytes: Multipotent mesodermal precursors exhibiting cardiac specificity. Stem Cells 2015, 33, 557–573. [Google Scholar] [CrossRef] [PubMed]
- König, M.A.; Canepa, D.D.; Cadosch, D.; Casanova, E.; Heinzelmann, M.; Rittirsch, D.; Plecko, M.; Hemmi, S.; Simmen, H.-P.; Cinelli, P.; et al. Direct transplantation of native pericytes from adipose tissue: A new perspective to stimulate healing in critical size bone defects. Cytotherapy 2016, 18, 41–52. [Google Scholar]
- Herrmann, M.; Bara, J.J.; Sprecher, C.M.; Menzel, U.; Jalowiec, J.M.; Osinga, R.; Scherberich, A.; Alini, M.; Verrier, S. Pericyte plasticity—Comparative investigation of the angiogenic and multilineage potential of pericytes from different human tissues. Cells Mater. 2016, 31, 236–241. [Google Scholar]
- Caplan, A.I. All mscs are pericytes? Cell Stem Cell 2008, 3, 229–230. [Google Scholar] [CrossRef] [PubMed]
- Blocki, A.; Wang, Y.; Koch, M.; Peh, P.; Beyer, S.; Law, P.; Hui, J.; Raghunath, M. Not all MSCs can act as pericytes: Functional in vitro assays to distinguish pericytes from other mesenchymal stem cells in angiogenesis. Stem Cells Dev. 2013, 22, 2347–2355. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Meirelles, L.; Caplan, A.I.; Nardi, N.B. In search of the in vivo identity of mesenchymal stem cells. Stem Cells 2008, 26, 2287–2299. [Google Scholar] [CrossRef] [PubMed]
- Davis, G.E.; Saunders, W.B. Molecular balance of capillary tube formation versus regression in wound repair: Role of matrix metalloproteinases and their inhibitors. J. Investig. Dermatol. Symp. Proc. 2006, 11, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Antonelli-Orlidge, A.; Saunders, K.B.; Smith, S.R.; D’Amore, P.A. An activated form of transforming growth factor β is produced by cocultures of endothelial cells and pericytes. Proc. Natl. Acad. Sci. USA 1989, 86, 4544–4548. [Google Scholar] [CrossRef] [PubMed]
- Kutcher, M.E.; Herman, I.M. The pericyte: Cellular regulator of microvascular blood flow. Microvasc. Res. 2009, 77, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Miller, F.N.; Sims, D.E.; Schuschke, D.A.; Abney, D.L. Differentiation of light-dye effects in the microcirculation. Microvasc. Res. 1992, 44, 166–184. [Google Scholar] [CrossRef]
- Sims, D.E.; Miller, F.N.; Horne, M.M.; Edwards, M.J. Interleukin-2 alters the positions of capillary and venule pericytes in rat cremaster muscle. J Submicrosc. Cytol. Pathol. 1994, 26, 507–513. [Google Scholar] [PubMed]
- Neng, L.; Zhang, F.; Kachelmeier, A.; Shi, X. Endothelial cell, pericyte, and perivascular resident macrophage-type melanocyte interactions regulate cochlear intrastrial fluid–blood barrier permeability. J. Associ. Res. Otolaryngol. 2013, 14, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Ogura, S.; Kurata, K.; Hattori, Y.; Takase, H.; Ishiguro-Oonuma, T.; Hwang, Y.; Ahn, S.; Park, I.; Ikeda, W.; Kusuhara, S.; et al. Sustained inflammation after pericyte depletion induces irreversible blood-retina barrier breakdown. JCI Insight 2017, 2, e90905. [Google Scholar] [CrossRef] [PubMed]
- Stark, K.; Eckart, A.; Haidari, S.; Tirniceriu, A.; Lorenz, M.; von Bruhl, M.-L.; Gartner, F.; Khandoga, A.G.; Legate, K.R.; Pless, R.; et al. Capillary and arteriolar pericytes attract innate leukocytes exiting through venules and ‘instruct’ them with pattern-recognition and motility programs. Nat. Immunol. 2013, 14, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Balabanov, R.; Dore-Duffy, P. Role of the cns microvascular pericyte in the blood-brain barrier. J. Neurosci. Res. 1998, 53, 637–644. [Google Scholar] [CrossRef]
- Tu, Z.; Li, Y.; Smith, D.S.; Sheibani, N.; Huang, S.; Kern, T.; Lin, F. Retinal pericytes inhibit activated T cell proliferation. Invest. Ophthalmol. Vis. Sci. 2011, 52, 9005–9010. [Google Scholar] [CrossRef] [PubMed]
- Jansson, D.; Rustenhoven, J.; Feng, S.; Hurley, D.; Oldfield, R.L.; Bergin, P.S.; Mee, E.W.; Faull, R.L.; Dragunow, M. A role for human brain pericytes in neuroinflammation. J. Neuroinflam. 2014, 11, 104. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.F.; Mittelsteadt, K.L.; Brauer, R.; McKinney, B.L.; Hallstrand, T.S.; Parks, W.C.; Chen, P.; Schnapp, L.M.; Liles, W.C.; Duffield, J.S.; et al. Lung pericyte-like cells are functional interstitial immune sentinel cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2017, 312, L556. [Google Scholar] [CrossRef] [PubMed]
- Paquet-Fifield, S.; Schlüter, H.; Li, A.; Aitken, T.; Gangatirkar, P.; Blashki, D.; Koelmeyer, R.; Pouliot, N.; Palatsides, M.; Ellis, S.; et al. A role for pericytes as microenvironmental regulators of human skin tissue regeneration. J. Clin. Investig. 2009, 119, 2795–2806. [Google Scholar] [CrossRef] [PubMed]
- Popescu, F.C.; Busuioc, C.J.; Mogosanu, G.D.; Pop, O.T.; Parvanescu, H.; Lascar, I.; Nicolae, C.I.; Mogoanta, L. Pericytes and myofibroblasts reaction in experimental thermal third degree skin burns. Rom. J. Morphol. Embryol. 2011, 52, 1011–1017. [Google Scholar] [PubMed]
- Sundberg, C.; Ljungstrom, M.; Lindmark, G.; Gerdin, B.; Rubin, K. Microvascular pericytes express platelet-derived growth factor-beta receptors in human healing wounds and colorectal adenocarcinoma. Am. J. Pathol. 1993, 143, 1377–1388. [Google Scholar] [PubMed]
- Lin, S.L.; Kisseleva, T.; Brenner, D.A.; Duffield, J.S. Pericytes and perivascular fibroblasts are the primary source of collagen-producing cells in obstructive fibrosis of the kidney. Am. J. Pathol. 2008, 173, 1617–1627. [Google Scholar] [CrossRef] [PubMed]
- Clark, E.R.; Clark, E.L. The relation of ‘rouget’ cells to capillary contractility. Dev. Dyn. 1925, 35, 265–282. [Google Scholar] [CrossRef]
- Rajkumar, V.S.; Shiwen, X.; Bostrom, M.; Leoni, P.; Muddle, J.; Ivarsson, M.; Gerdin, B.; Denton, C.P.; Bou-Gharios, G.; Black, C.M.; et al. Platelet-derived growth factor-beta receptor activation is essential for fibroblast and pericyte recruitment during cutaneous wound healing. Am. J. Pathol. 2006, 169, 2254–2265. [Google Scholar] [CrossRef] [PubMed]
- Mohle, R.; Green, D.; Moore, M.A.; Nachman, R.L.; Rafii, S. Constitutive production and thrombin-induced release of vascular endothelial growth factor by human megakaryocytes and platelets. Proc. Natl. Acad. Sci. USA 1997, 94, 663–668. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, Y.P.; Kirsner, R.S. Angiogenesis in wound repair: Angiogenic growth factors and the extracellular matrix. Microsc. Res. Tech. 2003, 60, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Tonnesen, M.G.; Feng, X.; Clark, R.A. Angiogenesis in wound healing. J. Investig. Dermatol. Symp. Proc. 2000, 5, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, A.G.; Iruela-Arispe, M.L. Extracellular matrix, inflammation, and the angiogenic response. Cardiovasc. Res. 2010, 86, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Hellstrom, M.; Gerhardt, H.; Kalen, M.; Li, X.; Eriksson, U.; Wolburg, H.; Betsholtz, C. Lack of pericytes leads to endothelial hyperplasia and abnormal vascular morphogenesis. J. Cell Biol. 2001, 153, 543–553. [Google Scholar] [CrossRef] [PubMed]
- Maisonpierre, P.C.; Suri, C.; Jones, P.F.; Bartunkova, S.; Wiegand, S.J.; Radziejewski, C.; Compton, D.; McClain, J.; Aldrich, T.H.; Papadopoulos, N.; et al. Angiopoietin-2, a natural antagonist for tie2 that disrupts in vivo angiogenesis. Science 1997, 277, 55. [Google Scholar] [CrossRef] [PubMed]
- Lindblom, P.; Gerhardt, H.; Liebner, S.; Abramsson, A.; Enge, M.; Hellstrom, M.; Backstrom, G.; Fredriksson, S.; Landegren, U.; Nystrom, H.C.; et al. Endothelial PDGF-β retention is required for proper investment of pericytes in the microvessel wall. Genes Dev. 2003, 17, 1835–1840. [Google Scholar] [CrossRef] [PubMed]
- Genove, G.; Mollick, T.; Johansson, K. Photoreceptor degeneration, structural remodeling and glial activation: A morphological study on a genetic mouse model for pericyte deficiency. Neuroscience 2014, 279, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Raines, S.M.; Richards, O.C.; Schneider, L.R.; Schueler, K.L.; Rabaglia, M.E.; Oler, A.T.; Stapleton, D.S.; Genove, G.; Dawson, J.A.; Betsholtz, C.; et al. Loss of PDGF-β activity increases hepatic vascular permeability and enhances insulin sensitivity. Am. J. Physiol. Endocrinol. Metab. 2011, 301, E517–E526. [Google Scholar] [CrossRef] [PubMed]
- Villasenor, R.; Kuennecke, B.; Ozmen, L.; Ammann, M.; Kugler, C.; Gruninger, F.; Loetscher, H.; Freskgard, P.O.; Collin, L. Region-specific permeability of the blood-brain barrier upon pericyte loss. J. Cereb. Blood Flow. Metab. 2017. [Google Scholar] [CrossRef] [PubMed]
- Darby, I.; Skalli, O.; Gabbiani, G. Alpha-smooth muscle actin is transiently expressed by myofibroblasts during experimental wound healing. Lab. Investig. 1990, 63, 21–29. [Google Scholar] [PubMed]
- Dulauroy, S.; Di Carlo, S.E.; Langa, F.; Eberl, G.; Peduto, L. Lineage tracing and genetic ablation of adam12+ perivascular cells identify a major source of profibrotic cells during acute tissue injury. Nat. Med. 2012, 18, 1262–1270. [Google Scholar] [CrossRef] [PubMed]
- LeBleu, V.S.; Taduri, G.; O’Connell, J.; Teng, Y.; Cooke, V.G.; Woda, C.; Sugimoto, H.; Kalluri, R. Origin and function of myofibroblasts in kidney fibrosis. Nat. Med. 2013, 19, 1047–1053. [Google Scholar] [CrossRef] [PubMed]
- Birbrair, A.; Zhang, T.; Files, D.C.; Mannava, S.; Smith, T.; Wang, Z.M.; Messi, M.L.; Mintz, A.; Delbono, O. Type-1 pericytes accumulate after tissue injury and produce collagen in an organ-dependent manner. Stem Cell Res. Ther. 2014, 5, 122. [Google Scholar] [CrossRef] [PubMed]
- Birbrair, A.; Zhang, T.; Wang, Z.-M.; Messi, M.L.; Olson, J.D.; Mintz, A.; Delbono, O. Type-2 pericytes participate in normal and tumoral angiogenesis. Am. J. Physiol. Cell Physiol. 2014, 307, C25–C38. [Google Scholar] [CrossRef] [PubMed]
- Mederacke, I.; Hsu, C.C.; Troeger, J.S.; Huebener, P.; Mu, X.; Dapito, D.H.; Pradere, J.P.; Schwabe, R.F. Fate tracing reveals hepatic stellate cells as dominant contributors to liver fibrosis independent of its aetiology. Nat. Commun. 2013, 4, 2823. [Google Scholar] [CrossRef] [PubMed]
- Meng, M.-B.; Zaorsky, N.G.; Deng, L.; Wang, H.-H.; Chao, J.; Zhao, L.-J.; Yuan, Z.-Y.; Ping, W. Pericytes: A double-edged sword in cancer therapy. Future Oncol. 2014, 11, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Xian, X.; Håkansson, J.; Ståhlberg, A.; Lindblom, P.; Betsholtz, C.; Gerhardt, H.; Semb, H. Pericytes limit tumor cell metastasis. J. Clin. Investig. 2006, 116, 642–651. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.-J.; You, W.-K.; Bonaldo, P.; Seyfried, T.N.; Pasquale, E.B.; Stallcup, W.B. Pericyte deficiencies lead to aberrant tumor vascularization in the brain of the ng2 null mouse. Dev. Biol. 2010, 344, 1035–1046. [Google Scholar] [CrossRef] [PubMed]
- Gibby, K.; You, W.-K.; Kadoya, K.; Helgadottir, H.; Young, L.J.T.; Ellies, L.G.; Chang, Y.; Cardiff, R.D.; Stallcup, W.B. Early vascular deficits are correlated with delayed mammary tumorigenesis in the mmtv-pymt transgenic mouse following genetic ablation of the ng2 proteoglycan. Breast Cancer Res. 2012, 14, R67. [Google Scholar] [CrossRef] [PubMed]
- Stallcup, W.B.; You, W.K.; Kucharova, K.; Cejudo-Martin, P.; Yotsumoto, F. Ng2 proteoglycan-dependent contributions of pericytes and macrophages to brain tumor vascularization and progression. Microcirculation 2016, 23, 122–133. [Google Scholar] [CrossRef] [PubMed]
- You, W.K.; Yotsumoto, F.; Sakimura, K.; Adams, R.H.; Stallcup, W.B. Ng2 proteoglycan promotes tumor vascularization via integrin-dependent effects on pericyte function. Angiogenesis 2014, 17, 61–76. [Google Scholar] [CrossRef] [PubMed]
- Braverman, I.M.; Sibley, J.; Keh, A. Ultrastructural analysis of the endothelial-pericyte relationship in diabetic cutaneous vessels. J. Investig. Dermatol. 1990, 95, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Braverman, I.M.; Keh-Yen, A. Ultrastructural abnormalities of the microvasculature and elastic fibers in the skin of juvenile diabetics. J. Investig. Dermatol. 1984, 82, 270–274. [Google Scholar] [CrossRef] [PubMed]
- Shakya, S.; Wang, Y.; Mack, J.A.; Maytin, E.V. Hyperglycemia-induced changes in hyaluronan contribute to impaired skin wound healing in diabetes: Review and perspective. Int. J. Cell Biol. 2015, 2015, 701738. [Google Scholar] [CrossRef] [PubMed]
- Laaff, H.; Vandscheidt, W.; Weiss, J.M.; Schaefer, H.E.; Schoepf, E. Immunohistochemical investigation of pericytes in chronic venous insufficiency. Vasa 1991, 20, 323–328. [Google Scholar] [PubMed]
- Safi, S.Z.; Qvist, R.; Kumar, S.; Batumalaie, K.; Ismail, I.S.B. Molecular mechanisms of diabetic retinopathy, general preventive strategies, and novel therapeutic targets. BioMed Res. Int. 2014, 2014, 801269. [Google Scholar] [CrossRef] [PubMed]
- Geraldes, P.; Hiraoka-Yamamoto, J.; Matsumoto, M.; Clermont, A.; Leitges, M.; Marette, A.; Aiello, L.P.; Kern, T.S.; King, G.L. Activation of pkcδ and shp1 by hyperglycemia causes vascular cell apoptosis and diabetic retinopathy. Nat. Med. 2009, 15, 1298–1306. [Google Scholar] [CrossRef] [PubMed]
- Ejaz, S.; Chekarova, I.; Ejaz, A.; Sohail, A.; Lim, C.W. Importance of pericytes and mechanisms of pericyte loss during diabetic retinopathy. Diabetes Obes. Metab. 2008, 10, 53–63. [Google Scholar] [PubMed]
- Ricard, N.; Tu, L.; Le Hiress, M.; Huertas, A.; Phan, C.; Thuillet, R.; Sattler, C.; Fadel, E.; Seferian, A.; Montani, D.; et al. Increased pericyte coverage mediated by endothelial-derived fibroblast growth factor-2 and interleukin-6 is a source of smooth muscle–like cells in pulmonary hypertension. Circulation 2014, 129, 1586. [Google Scholar] [CrossRef] [PubMed]
- Sengillo, J.D.; Winkler, E.A.; Walker, C.T.; Sullivan, J.S.; Johnson, M.; Zlokovic, B.V. Deficiency in mural vascular cells coincides with blood-brain barrier disruption in alzheimer’s disease. Brain Pathol. 2013, 23, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Humphreys, B.D.; Lin, S.-L.; Kobayashi, A.; Hudson, T.E.; Nowlin, B.T.; Bonventre, J.V.; Valerius, M.T.; McMahon, A.P.; Duffield, J.S. Fate tracing reveals the pericyte and not epithelial origin of myofibroblasts in kidney fibrosis. Am. J. Pathol. 2010, 176, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Winkler, E.A.; Sagare, A.P.; Zlokovic, B.V. The pericyte: A forgotten cell type with important implications for Alzheimer’s disease? Brain Pathol. 2014, 24, 371–386. [Google Scholar] [CrossRef] [PubMed]
- Faulkner, J.L.; Szcykalski, L.M.; Springer, F.; Barnes, J.L. Origin of interstitial fibroblasts in an accelerated model of angiotensin ii-induced renal fibrosis. Am. J. Pathol. 2005, 167, 1193–1205. [Google Scholar] [CrossRef]
- Kramann, R.; Humphreys, B.D. Kidney pericytes: Roles in regeneration and fibrosis. Semin. Nephrol. 2014, 34, 374–383. [Google Scholar] [CrossRef] [PubMed]
- Crisan, M.; Yap, S.; Casteilla, L.; Chen, C.-W.; Corselli, M.; Park, T.S.; Andriolo, G.; Sun, B.; Zheng, B.; Zhang, L.; et al. A perivascular origin for mesenchymal stem cells in multiple human organs. Cell Stem Cell 2008, 3, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Dar, A.; Domev, H.; Ben-Yosef, O.; Tzukerman, M.; Zeevi-Levin, N.; Novak, A.; Germanguz, I.; Amit, M.; Itskovitz-Eldor, J. Multipotent vasculogenic pericytes from human pluripotent stem cells promote recovery of murine ischemic limb. Circulation 2012, 125, 87. [Google Scholar] [CrossRef] [PubMed]
- Park, T.S.; Gavina, M.; Chen, C.-W.; Sun, B.; Teng, P.-N.; Huard, J.; Deasy, B.M.; Zimmerlin, L.; Péault, B. Placental perivascular cells for human muscle regeneration. Stem Cells Dev. 2011, 20, 451–463. [Google Scholar] [CrossRef] [PubMed]
- Birbrair, A.; Zhang, T.; Wang, Z.-M.; Messi, M.L.; Mintz, A.; Delbono, O. Type-1 pericytes participate in fibrous tissue deposition in aged skeletal muscle. Am. J. Pathol. 2013, 305, C1098–C1113. [Google Scholar] [CrossRef] [PubMed]
- Greenhalgh, S.N.; Conroy, K.P.; Henderson, N.C. Healing scars: Targeting pericytes to treat fibrosis. Int. J. Med. 2015, 108, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Kida, Y.; Duffield, J.S. Frontiers in research: Chronic kidney diseases: The pivotal role of pericytes in kidney fibrosis. Clin. Exp. Pharmacol. Physiol. 2011, 38, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Tawonsawatruk, T.; West, C.C.; Murray, I.R.; Soo, C.; Péault, B.; Simpson, A.H.R.W. Adipose derived pericytes rescue fractures from a failure of healing—Non-union. Sci. Rep. 2016, 6, 22779. [Google Scholar] [CrossRef] [PubMed]
- Mendel, T.A.; Clabough, E.B.D.; Kao, D.S.; Demidova-Rice, T.N.; Durham, J.T.; Zotter, B.C.; Seaman, S.A.; Cronk, S.M.; Rakoczy, E.P.; Katz, A.J.; et al. Pericytes derived from adipose-derived stem cells protect against retinal vasculopathy. PLoS ONE 2013, 8, e65691. [Google Scholar] [CrossRef]
- Zebardast, N.; Lickorish, D.; Davies, J.E. Human umbilical cord perivascular cells (HUCPVC): A mesenchymal cell source for dermal wound healing. Organogenesis 2010, 6, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Zamora, D.O.; Natesan, S.; Becerra, S.; Wrice, N.; Chung, E.; Suggs, L.J.; Christy, R.J. Enhanced wound vascularization using a dsASCs seeded FPEG scaffold. Angiogenesis 2013, 16, 745–757. [Google Scholar] [CrossRef] [PubMed]
- Mills, S.J.; Zhuang, L.; Arandjelovic, P.; Cowin, A.J.; Kaur, P. Effects of human pericytes in a murine excision model of wound healing. Exp. Dermatol. 2015, 24, 881–882. [Google Scholar] [CrossRef] [PubMed]
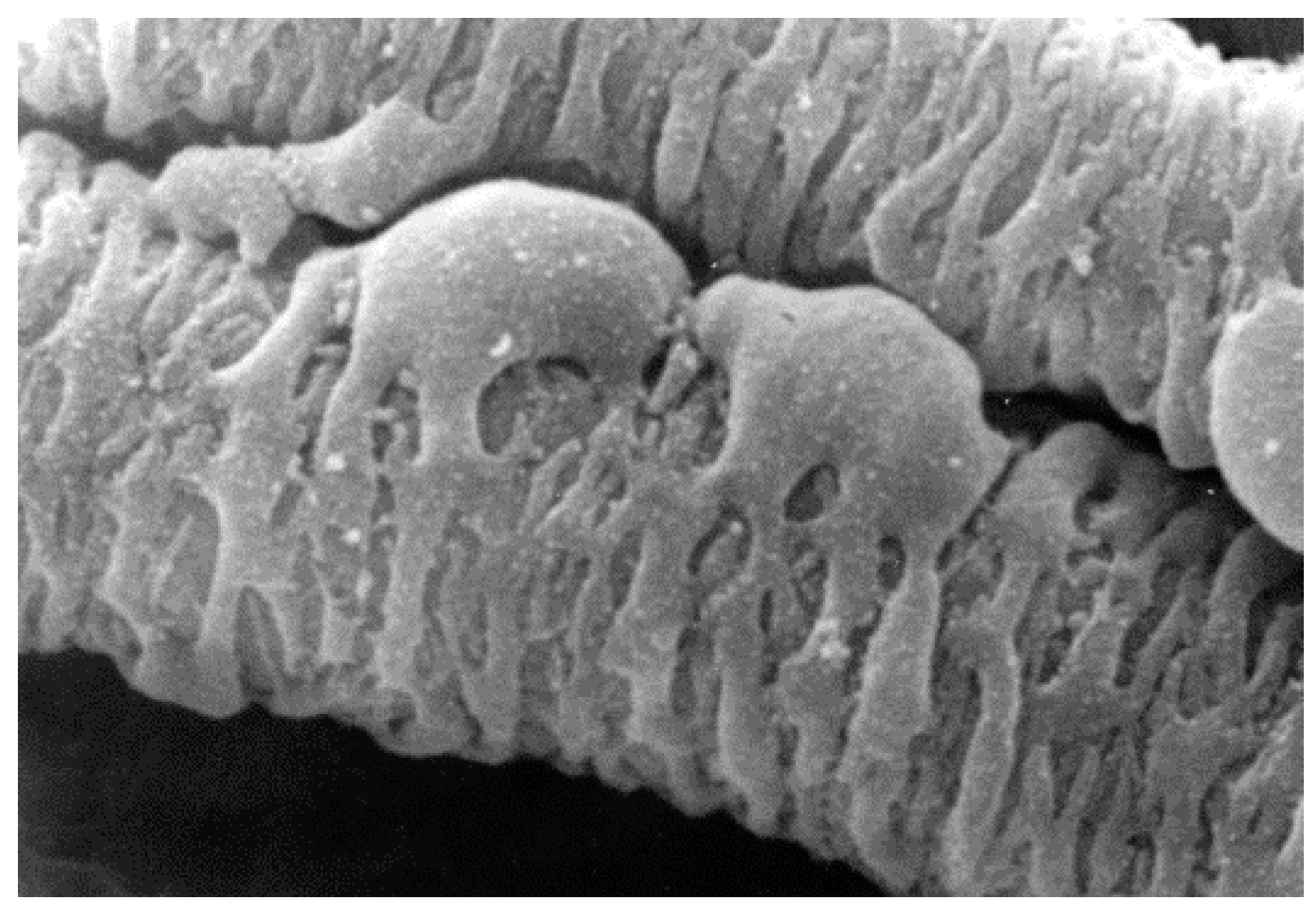

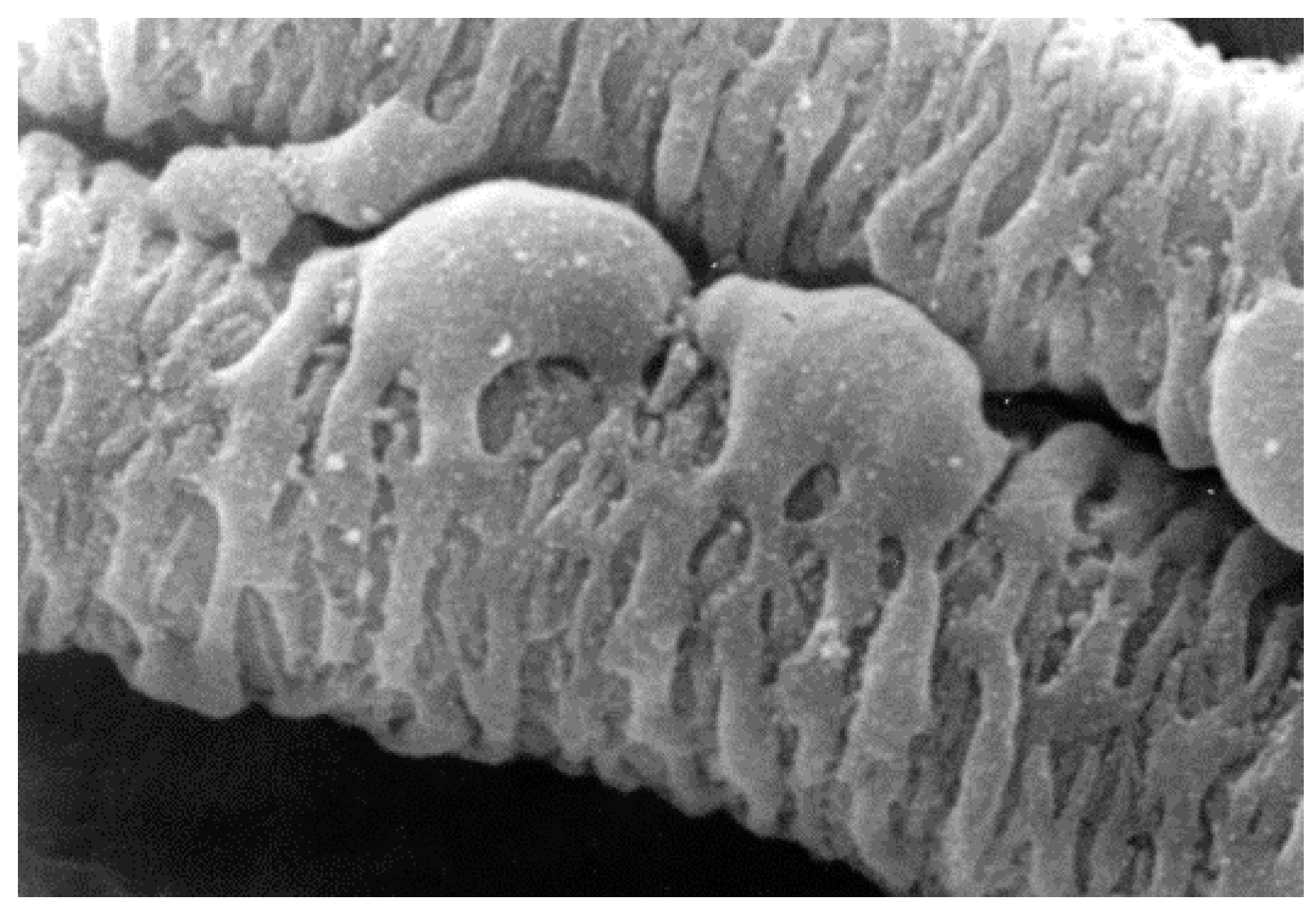{kind=link}
| Wound Healing Process | Pericyte Functions |
|---|---|
| Angiogenesis | Structural support of existing blood vessels [19] |
| Regulation of EC proliferation and migration to form new vessels [8] | |
| Prevention of capillary tube regression by TIMP-3 expression [31] | |
| Stabilisation of newly formed capillaries [32,33] | |
| Inflammation | Regulation of vessel permeability [34,35,36] |
| Regulation of neutrophil extravasion [9,10] | |
| Regulation of macrophage extravasion [11,37] | |
| Control of leukocyte trafficking [38] | |
| Control of T cell activation [39,40] | |
| Response to inflammatory signals [41,42] | |
| Re-epithelialisation | Regulation of keratinocyte migration [43] |
| Fibrosis | Production of collagen [44,45] |
| Differentiation into myofibroblasts [46] | |
| Tissue regeneration | MSC-like properties: differentiation potential includes adipocytes, osteoblasts, chondrocytes, phagocytes and granulocytes [2,25,26] |
| Disorder | Pericyte Aberrance Observed | Pericyte Functions Likely to Impact Disease |
|---|---|---|
| Diabetic chronic healing | Decreased pericyte numbers in dermis, pericytes exhibit altered morphology [72,73,74,75] | Angiogenesis-decreased vascularisation |
| Vessel permeability-leaky vessels lead to prolonged and uncontrolled inflammation | ||
| Fibrosis-pericytes promote fibrotic vessels | ||
| Stem cell properties-replacement of lost cell/tissue types | ||
| Diabetic retinopathy | Decreased pericyte numbers, increased pericyte apoptosis [76] | Angiogenesis-decreased control of endothelial proliferation |
| Vessel permeability-leakiness of vessels | ||
| Solid tumour | Unknown, however control of angiogenesis has long been recognised as an important target in treatment of solid tumours | Angiogenesis-tumour relies on new vasculature for blood supply |
| Endothelial control-metastasis of cancer | ||
| Vessel permeability-ability of chemotherapeutic agents to pass from bloodstream to tumour tissue | ||
| Pulmonary arterial hypertension (PAH) | Increased pericyte coverage on pulmonary arteries [78] | Angiogenesis-excessive remodelling of pulmonary vasculature and endothelial dysfunction |
| Alzheimers (AD) | Degeneration at blood brain barrier (BBB) [79] | Angiogenesis-break down of vessels causes decreased cereberal bloodflow leading to neurodegeneration |
| Vessel permeability-accumulation of damaging molecules in the brain | ||
| Chronic kidney disease (CKD) | Differentiation of pericytes into myofibroblasts [80] | Fibrosis-pericytes thought to be source of myofibroblasts contributing to excessive fibrotic activity |
| Angiogenesis-differentiation of pericytes into myofibroblasts leaves less pericytes to stabilise vasculature |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thomas, H.M.; Cowin, A.J.; Mills, S.J. The Importance of Pericytes in Healing: Wounds and other Pathologies. Int. J. Mol. Sci. 2017, 18, 1129. https://doi.org/10.3390/ijms18061129
Thomas HM, Cowin AJ, Mills SJ. The Importance of Pericytes in Healing: Wounds and other Pathologies. International Journal of Molecular Sciences. 2017; 18(6):1129. https://doi.org/10.3390/ijms18061129
Chicago/Turabian StyleThomas, Hannah M., Allison J. Cowin, and Stuart J. Mills. 2017. "The Importance of Pericytes in Healing: Wounds and other Pathologies" International Journal of Molecular Sciences 18, no. 6: 1129. https://doi.org/10.3390/ijms18061129
APA StyleThomas, H. M., Cowin, A. J., & Mills, S. J. (2017). The Importance of Pericytes in Healing: Wounds and other Pathologies. International Journal of Molecular Sciences, 18(6), 1129. https://doi.org/10.3390/ijms18061129