
From Belly to Brain: Targeting the Ghrelin Receptor in Appetite and Food Intake Regulation

Abstract
:
1. Introduction
1.1. Ghrelin and Appetite-Related Disorders
1.2. Homeostatic Feeding
1.3. Non-Homeostatic Feeding
1.4. Stress, Impulsivity and Cognition
1.5. Current Status and Implications
2. Growth Hormone Secretagogue Receptor (GHSR-1a) Receptor—Biodistribution and Signalling
2.1. Pleiotropic Pharmacodynamics
2.2. Central GHSR-1a Signalling
2.3. Peripheral GHSR-1a Signalling
2.4. Complementary Signalling: Gastrointestinal Motility, Glucose Homeostasis and Visceral Pain
2.5. Heterogenous Action—GHSR-1a as a Promiscuous Target
3. Ghrelin and Ghrelin Ligands: Pharmacokinetic Perspectives
3.1. Blood Brain Barrier Penetration
3.2. Vagus Nerve Signalling
3.3. Ghrelin Human Studies
3.4. Acyl and Desacyl-Ghrelin—Implications for Therapeutic Approaches
3.5. Synthetic Ghrelin Ligands
3.6. Hunger Is the Best Sauce—Targeting the Mesolimbic Reward Circuitry
3.7. Homeostatic “Gating” of the Reward System
4. Conclusions and Future Directions
Conflicts of Interest
Abbreviations
| GHSR-1a | Growth Hormone Secretagogue Receptor-Type 1a |
| BBB | Blood Brain Barrier |
| GPCR | G-Protein Coupled Receptor |
| CNS | Central Nervous System |
| CSF | Cerebrospinal Fluid |
| NTS | Nucleus Tractus Solitarius |
| CCK | Cholecystokinin |
| GH | Growth Hormone |
| GHRP | Growth Hormone Releasing Peptide |
| DIO | Diet Induced Obesity |
| VTA | Ventral Tegmental Area |
| NAcc | Nucleus Accumbens |
| LH | Lateral Hypothalamus |
| PBN | Parabrachial Nucleus |
References
- Schwartz, M.W.; Woods, S.C.; Porte, D.; Seeley, R.J.; Baskin, D.G. Central nervous system control of food intake. Nature 2000, 404, 661–671. [Google Scholar] [PubMed]
- Kojima, M.; Hosoda, H.; Date, Y.; Nakazato, M.; Matsuo, H.; Kangawa, K. Ghrelin is a growth-hormone-releasing acylated peptide from stomach. Nature 1999, 402, 656–660. [Google Scholar] [CrossRef] [PubMed]
- Cummings, D.E.; Purnell, J.Q.; Frayo, R.S.; Schmidova, K.; Wisse, B.E.; Weigle, D.S. A preprandial rise in plasma ghrelin levels suggests a role in meal initiation in humans. Diabetes 2001, 50, 1714–1719. [Google Scholar] [CrossRef] [PubMed]
- McFarlane, M.R.; Brown, M.S.; Goldstein, J.L.; Zhao, T.J. Induced ablation of ghrelin cells in adult mice does not decrease food intake, body weight, or response to high-fat diet. Cell Metab. 2014, 20, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Nakazato, M.; Murakami, N.; Date, Y.; Kojima, M.; Matsuo, H.; Kangawa, K.; Matsukura, S. A role for ghrelin in the central regulation of feeding. Nature 2001, 409, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Mason, B.L.; Wang, Q.; Zigman, J.M. The central nervous system sites mediating the orexigenic actions of ghrelin. Annu. Rev. Physiol. 2014, 76, 519–533. [Google Scholar] [CrossRef] [PubMed]
- Müller, T.D.; Nogueiras, R.; Andermann, M.L.; Andrews, Z.B.; Anker, S.D.; Argente, J.; Batterham, R.L.; Benoit, S.C.; Bowers, C.Y.; Broglio, F.; et al. Ghrelin. Mol. Metab. 2015, 4, 437–460. [Google Scholar] [CrossRef] [PubMed]
- Seim, I.; Collet, C.; Herington, A.C.; Chopin, L.K. Revised genomic structure of the human ghrelin gene and identification of novel exons, alternative splice variants and natural antisense transcripts. BMC Genom. 2007, 8, 298. [Google Scholar] [CrossRef] [PubMed]
- Asakawa, A.; Inui, A.; Fujimiya, M.; Sakamaki, R.; Shinfuku, N.; Ueta, Y.; Meguid, M.M.; Kasuga, M. Stomach regulates energy balance via acylated ghrelin and desacyl ghrelin. Gut 2005, 54, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Tschop, M.; Smiley, D.L.; Heiman, M.L. Ghrelin induces adiposity in rodents. Nature 2000, 407, 908–913. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Brown, M.S.; Liang, G.; Grishin, N.V.; Goldstein, J.L. Identification of the acyltransferase that octanoylates ghrelin, an appetite-stimulating peptide hormone. Cell 2008, 132, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Zigman, J.M.; Jones, J.E.; Lee, C.E.; Saper, C.B.; Elmquist, J.K. Expression of ghrelin receptor mRNA in the rat and the mouse brain. J. Comp. Neurol. 2006, 494, 528–548. [Google Scholar] [CrossRef] [PubMed]
- Gnanapavan, S.; Kola, B.; Bustin, S.A.; Morris, D.G.; McGee, P.; Fairclough, P.; Bhattacharya, S.; Carpenter, R.; Grossman, A.B.; Korbonits, M. The tissue distribution of the mRNA of ghrelin and subtypes of its receptor, GHS-R, in humans. J. Clin. Endocrinol. Metab. 2002, 87, 2988–2991. [Google Scholar] [CrossRef] [PubMed]
- Cowley, M.A.; Smith, R.G.; Diano, S.; Tschöp, M.; Pronchuk, N.; Grove, K.L.; Strasburger, C.J.; Bidlingmaier, M.; Esterman, M.; Heiman, M.L.; et al. The distribution and mechanism of action of ghrelin in the CNS demonstrates a novel hypothalamic circuit regulating energy homeostasis. Neuron 2003, 37, 649–661. [Google Scholar] [CrossRef]
- Takaya, K.; Ariyasu, H.; Kanamoto, N.; Iwakura, H.; Yoshimoto, A.; Harada, M.; Mori, K.; Komatsu, Y.; Usui, T.; Shimatsu, A.; et al. Ghrelin strongly stimulates growth hormone release in humans. J. Clin. Endocrinol. Metab. 2000, 85, 4908–4911. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, J.L.; Zhao, T.-J.; Li, R.; Sherbet, D.; Liang, G.; Brown, M. Surviving starvation: Essential role of the ghrelin-growth hormone axis. Cold Spring Harb. Symp. Quant. Biol. 2011, 76, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Abizaid, A.; Liu, Z.W.; Andrews, Z.B.; Shanabrough, M.; Borok, E.; Elsworth, J.D.; Roth, R.H.; Sleeman, M.W.; Picciotto, M.R.; Tschop, M.H.; et al. Ghrelin modulates the activity and synaptic input organization of midbrain dopamine neurons while promoting appetite. J. Clin. Investig. 2006, 116, 3229–3239. [Google Scholar] [CrossRef] [PubMed]
- Chuang, J.-C.; Perello, M.; Sakata, I.; Osborne-Lawrence, S.; Savitt, J.M.; Lutter, M.; Zigman, J.M. Ghrelin mediates stress-induced food-reward behavior in mice. J. Clin. Investig. 2011, 121, 2684–2692. [Google Scholar] [CrossRef] [PubMed]
- Carlini, V.P.; Monzon, M.E.; Varas, M.M.; Cragnolini, A.B.; Schioth, H.B.; Scimonelli, T.N.; de Barioglio, S.R. Ghrelin increases anxiety-like behavior and memory retention in rats. Biochem. Biophys. Res. Commun. 2002, 299, 739–743. [Google Scholar] [CrossRef]
- Dixit, V.D.; Schaffer, E.M.; Pyle, R.S.; Collins, G.D.; Sakthivel, S.K.; Palaniappan, R.; Lillard, J.W., Jr.; Taub, D.D. Ghrelin inhibits leptin- and activation-induced proinflammatory cytokine expression by human monocytes and t cells. J. Clin. Investig. 2004, 114, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Fukue, Y.; Teranishi, H.; Yoshida, Y.; Kojima, M. Molecular forms of hypothalamic ghrelin and its regulation by fasting and 2-deoxy-d-glucose administration. Endocrinology 2005, 146, 2510–2516. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liu, X.; Shu, Q.; Li, S.; Luo, F. Ghrelin attenuates lipopolysaccharide-induced acute lung injury through NO pathway. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2008, 14, Br141–Br146. [Google Scholar]
- Li, G.; Li, J.; Zhou, Q.; Song, X.; Liang, H.; Huang, L. Growth hormone releasing peptide-2, a ghrelin agonist, attenuates lipopolysaccharide-induced acute lung injury in rats. Tohoku J. Exp. Med. 2010, 222, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Baldanzi, G.; Filigheddu, N.; Cutrupi, S.; Catapano, F.; Bonissoni, S.; Fubini, A.; Malan, D.; Baj, G.; Granata, R.; Broglio, F.; et al. Ghrelin and des-acyl ghrelin inhibit cell death in cardiomyocytes and endothelial cells through ERK1/2 and PI 3-kinase/AKT. J. Cell Biol. 2002, 159, 1029–1037. [Google Scholar] [CrossRef] [PubMed]
- Muccioli, G.; Lorenzi, T.; Lorenzi, M.; Ghè, C.; Arnoletti, E.; Raso, G.M.; Castellucci, M.; Gualillo, O.; Meli, R. Beyond the metabolic role of ghrelin: A new player in the regulation of reproductive function. Peptides 2011, 32, 2514–2521. [Google Scholar] [CrossRef] [PubMed]
- Schellekens, H.; Dinan, T.G.; Cryan, J.F. Chapter thirteen—Ghrelin at the interface of obesity and reward. Vitam. Horm. 2013, 91, 285–323. [Google Scholar] [PubMed]
- Jerlhag, E.; Egecioglu, E.; Dickson, S.L.; Andersson, M.; Svensson, L.; Engel, J.A. Ghrelin stimulates locomotor activity and accumbal dopamine-overflow via central cholinergic systems in mice: Implications for its involvement in brain reward. Addict. Biol. 2006, 11, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Egecioglu, E.; Skibicka, K.P.; Hansson, C.; Alvarez-Crespo, M.; Friberg, P.A.; Jerlhag, E.; Engel, J.A.; Dickson, S.L. Hedonic and incentive signals for body weight control. Rev. Endocr. Metab. Disord. 2011, 12, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Dickson, S.L.; Egecioglu, E.; Landgren, S.; Skibicka, K.P.; Engel, J.A.; Jerlhag, E. The role of the central ghrelin system in reward from food and chemical drugs. Mol. Cell. Endocrinol. 2011, 340, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Berthoud, H.R. Homeostatic and non-homeostatic pathways involved in the control of food intake and energy balance. Obesity 2006, 14, 197S–200S. [Google Scholar] [CrossRef] [PubMed]
- Cummings, D.E. Ghrelin and the short- and long-term regulation of appetite and body weight. Physiol. Behav. 2006, 89, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Argilés, J.M.; López-Soriano, F.J.; Busquets, S. Novel approaches to the treatment of cachexia. Drug Discov. Today 2008, 13, 73–78. [Google Scholar] [CrossRef] [PubMed]
- De Vriese, C.; Perret, J.; Delporte, C. Focus on the short- and long-term effects of ghrelin on energy homeostasis. Nutrition 2010, 26, 579–584. [Google Scholar] [CrossRef] [PubMed]
- Malafarina, V.; Uriz-Otano, F.; Gil-Guerrero, L.; Iniesta, R. The anorexia of ageing: Physiopathology, prevalence, associated comorbidity and mortality. A systematic review. Maturitas 2013, 74, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Chapman, I.M. Endocrinology of anorexia of ageing. Best Pract. Res. Clin. Endocrinol. Metab. 2004, 18, 437–452. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). World Health Organisation Global Health and Ageing; World Health Organization: Geneva, Switzerland, 2011. [Google Scholar]
- DeBoer, M.D. Emergence of ghrelin as a treatment for cachexia syndromes. Nutrition 2008, 24, 806–814. [Google Scholar] [CrossRef] [PubMed]
- DeBoer, M.D. Ghrelin and cachexia: Will treatment with GHSR-1a agonists make a difference for patients suffering from chronic wasting syndromes? Mol. Cell. Endocrinol. 2011, 340, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Nass, R.; Gaylinn, B.D.; Thorner, M.O. The ghrelin axis in disease: Potential therapeutic indications. Mol. Cell. Endocrinol. 2011, 340, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Hickson, M. Malnutrition and ageing. Postgrad. Med. J. 2006, 82, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Wren, A.M.; Small, C.J.; Abbott, C.R.; Dhillo, W.S.; Seal, L.J.; Cohen, M.A.; Batterham, R.L.; Taheri, S.; Stanley, S.A.; Ghatei, M.A.; et al. Ghrelin causes hyperphagia and obesity in rats. Diabetes 2001, 50, 2540–2547. [Google Scholar] [CrossRef] [PubMed]
- Horvath, T.L.; Castaneda, T.; Tang-Christensen, M.; Pagotto, U.; Tschop, M.H. Ghrelin as a potential anti-obesity target. Curr. Pharm. Des. 2003, 9, 1383–1395. [Google Scholar] [CrossRef] [PubMed]
- World Health Organisation (WHO). 2016. Available online: http://www.Who.Int/mediacentre/factsheets/fs311/en/ (accessed on 15 June 2016).
- Isomaa, B.; Almgren, P.; Tuomi, T.; Forsen, B.; Lahti, K.; Nissen, M.; Taskinen, M.R.; Groop, L. Cardiovascular morbidity and mortality associated with the metabolic syndrome. Diabetes Care 2001, 24, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Martin, K.A.; Mani, M.V.; Mani, A. New targets to treat obesity and the metabolic syndrome. Eur. J. Pharmacol. 2015, 763, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Kenny, P.J. Reward mechanisms in obesity: New insights and future directions. Neuron 2011, 69, 664–679. [Google Scholar] [CrossRef] [PubMed]
- Volkow, N.D.; Wang, G.J.; Fowler, J.S.; Tomasi, D.; Baler, R. Food and drug reward: Overlapping circuits in human obesity and addiction. Curr. Top. Behav. Neurosci. 2012, 11, 1–24. [Google Scholar] [PubMed]
- Spanagel, R.; Weiss, F. The dopamine hypothesis of reward: Past and current status. Trends Neurosci. 1999, 22, 521–527. [Google Scholar] [CrossRef]
- Pierce, R.C.; Kumaresan, V. The mesolimbic dopamine system: The final common pathway for the reinforcing effect of drugs of abuse? Neurosci. Biobehav. Rev. 2006, 30, 215–238. [Google Scholar] [CrossRef] [PubMed]
- Stoeckel, L.E.; Weller, R.E.; Cook, E.W., 3rd; Twieg, D.B.; Knowlton, R.C.; Cox, J.E. Widespread reward-system activation in obese women in response to pictures of high-calorie foods. NeuroImage 2008, 41, 636–647. [Google Scholar] [CrossRef] [PubMed]
- Stice, E.; Yokum, S.; Blum, K.; Bohon, C. Weight gain is associated with reduced striatal response to palatable food. J. Neurosci. 2010, 30, 13105–13109. [Google Scholar] [CrossRef] [PubMed]
- Konturek, S.J.; Konturek, J.W.; Pawlik, T.; Brzozowski, T. Brain-gut axis and its role in the control of food intake. J. Physiol. Pharmacol. 2004, 55, 137–154. [Google Scholar] [PubMed]
- Avena, N.M.; Rada, P.; Hoebel, B.G. Evidence for sugar addiction: Behavioral and neurochemical effects of intermittent, excessive sugar intake. Neurosci. Biobehav. Rev. 2008, 32, 20–39. [Google Scholar] [CrossRef] [PubMed]
- Tanda, G.; di Chiara, G. A dopamine-μ1 opioid link in the rat ventral tegmentum shared by palatable food (fonzies) and non-psychostimulant drugs of abuse. Eur. J. Neurosci. 1998, 10, 1179–1187. [Google Scholar] [CrossRef] [PubMed]
- Volkow, N.D.; Wang, G.J.; Tomasi, D.; Baler, R.D. The addictive dimensionality of obesity. Biol. Psychiatry 2013, 73, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Hebebrand, J.; Albayrak, O.; Adan, R.; Antel, J.; Dieguez, C.; de Jong, J.; Leng, G.; Menzies, J.; Mercer, J.G.; Murphy, M.; et al. “Eating addiction”, rather than “food addiction”, better captures addictive-like eating behavior. Neurosci. Biobehav. Rev. 2014, 47, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Perello, M.; Dickson, S.L. Ghrelin signalling on food reward: A salient link between the gut and the mesolimbic system. J. Neuroendocrinol. 2015, 27, 424–434. [Google Scholar] [CrossRef] [PubMed]
- Druce, M.R.; Wren, A.M.; Park, A.J.; Milton, J.E.; Patterson, M.; Frost, G.; Ghatei, M.A.; Small, C.; Bloom, S.R. Ghrelin increases food intake in obese as well as lean subjects. Int. J. Obes. 2005, 29, 1130–1136. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, Y.; Kaneko, F.; Yamada, M.; Kishikawa, Y.; Kawahara, H.; Nishi, A. Food reward-sensitive interaction of ghrelin and opioid receptor pathways in mesolimbic dopamine system. Neuropharmacology 2013, 67, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Goldstone, A.P.; Prechtl, C.G.; Scholtz, S.; Miras, A.D.; Chhina, N.; Durighel, G.; Deliran, S.S.; Beckmann, C.; Ghatei, M.A.; Ashby, D.R.; et al. Ghrelin mimics fasting to enhance human hedonic, orbitofrontal cortex, and hippocampal responses to food. Am. J. Clin. Nutr. 2014, 99, 1319–1330. [Google Scholar] [CrossRef] [PubMed]
- Malik, S.; McGlone, F.; Bedrossian, D.; Dagher, A. Ghrelin modulates brain activity in areas that control appetitive behavior. Cell Metab. 2008, 7, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Andrews, Z.B. The extra-hypothalamic actions of ghrelin on neuronal function. Trends Neurosci. 2011, 34, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Pattij, T.; Janssen, M.C.; Vanderschuren, L.J.; Schoffelmeer, A.N.; van Gaalen, M.M. Involvement of dopamine D1 and D2 receptors in the nucleus accumbens core and shell in inhibitory response control. Psychopharmacology 2007, 191, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Dawe, S.; Loxton, N.J. The role of impulsivity in the development of substance use and eating disorders. Neurosci. Biobehav. Rev. 2004, 28, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Velazquez-Sanchez, C.; Ferragud, A.; Moore, C.F.; Everitt, B.J.; Sabino, V.; Cottone, P. High trait impulsivity predicts food addiction-like behavior in the rat. Neuropsychopharmacology 2014, 39, 2463–2472. [Google Scholar] [CrossRef] [PubMed]
- Schag, K.; Schonleber, J.; Teufel, M.; Zipfel, S.; Giel, K.E. Food-related impulsivity in obesity and binge eating disorder—A systematic review. Obes. Rev. 2013, 14, 477–495. [Google Scholar] [CrossRef] [PubMed]
- Anderberg, R.H.; Hansson, C.; Fenander, M.; Richard, J.E.; Dickson, S.L.; Nissbrandt, H.; Bergquist, F.; Skibicka, K.P. The stomach-derived hormone ghrelin increases impulsive behavior. Neuropsychopharmacology 2016, 41, 1199–1209. [Google Scholar] [CrossRef] [PubMed]
- Gibson, E.L. Emotional influences on food choice: Sensory, physiological and psychological pathways. Physiol. Behav. 2006, 89, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Oliver, G.; Wardle, J. Perceived effects of stress on food choice. Physiol. Behav. 1999, 66, 511–515. [Google Scholar] [CrossRef]
- Dallman, M.F. Stress-induced obesity and the emotional nervous system. Trends Endocrinol. Metab. TEM 2010, 21, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Swinburn, B.A.; Sacks, G.; Hall, K.D.; McPherson, K.; Finegood, D.T.; Moodie, M.L.; Gortmaker, S.L. The global obesity pandemic: Shaped by global drivers and local environments. Lancet 2011, 378, 804–814. [Google Scholar] [CrossRef]
- Marti, A.; Moreno-Aliaga, M.J.; Hebebrand, J.; Martinez, J.A. Genes, lifestyles and obesity. Int. J. Obes. Relat. Metab. Disord. 2004, 28, S29–S36. [Google Scholar] [CrossRef] [PubMed]
- Schellekens, H.; Finger, B.C.; Dinan, T.G.; Cryan, J.F. Ghrelin signalling and obesity: At the interface of stress, mood and food reward. Pharmacol. Ther. 2012, 135, 316–326. [Google Scholar] [CrossRef] [PubMed]
- Perello, M.; Sakata, I.; Birnbaum, S.; Chuang, J.-C.; Osborne-Lawrence, S.; Rovinsky, S.A.; Woloszyn, J.; Yanagisawa, M.; Lutter, M.; Zigman, J.M. Ghrelin increases the rewarding value of high-fat diet in an orexin-dependent manner. Biol. Psychiatry 2010, 67, 880–886. [Google Scholar] [CrossRef] [PubMed]
- Carr, K.A.; Daniel, T.O.; Lin, H.; Epstein, L.H. Reinforcement pathology and obesity. Curr. Drug Abuse Rev. 2011, 4, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Von Haehling, S.; Anker, S.D. Prevalence, incidence and clinical impact of cachexia: Facts and numbers—Update 2014. J. Cachexia Sarcopenia Muscle 2014, 5, 261–263. [Google Scholar] [CrossRef] [PubMed]
- Schellekens, H.; Dinan, T.G.; Cryan, J.F. Lean mean fat reducing “ghrelin” machine: Hypothalamic ghrelin and ghrelin receptors as therapeutic targets in obesity. Neuropharmacology 2010, 58, 2–16. [Google Scholar] [CrossRef] [PubMed]
- Andrews, Z.B. Central mechanisms involved in the orexigenic actions of ghrelin. Peptides 2011, 32, 2248–2255. [Google Scholar] [CrossRef] [PubMed]
- Stengel, A.; Goebel, M.; Wang, L.; Tache, Y. Ghrelin, des-acyl ghrelin and nesfatin-1 in gastric X/A-like cells: Role as regulators of food intake and body weight. Peptides 2010, 31, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Theander-Carrillo, C.; Wiedmer, P.; Cettour-Rose, P.; Nogueiras, R.; Perez-Tilve, D.; Pfluger, P.; Castaneda, T.R.; Muzzin, P.; Schurmann, A.; Szanto, I.; et al. Ghrelin action in the brain controls adipocyte metabolism. J. Clin. Investig. 2006, 116, 1983–1993. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.S.; Kotokorpi, P.; Eccles, S.R.; Barnes, S.K.; Tokarczuk, P.F.; Allen, S.K.; Whitworth, H.S.; Guschina, I.A.; Evans, B.A.; Mode, A.; et al. Ghrelin induces abdominal obesity via GHS-R-dependent lipid retention. Mol. Endocrinol. 2009, 23, 914–924. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.; Gomez-Ambrosi, J.; Catalan, V.; Gil, M.J.; Becerril, S.; Sainz, N.; Silva, C.; Salvador, J.; Colina, I.; Fruhbeck, G. Acylated and desacyl ghrelin stimulate lipid accumulation in human visceral adipocytes. Int. J. Obes. 2009, 33, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Wren, A.M.; Small, C.J.; Ward, H.L.; Murphy, K.G.; Dakin, C.L.; Taheri, S.; Kennedy, A.R.; Roberts, G.H.; Morgan, D.G.; Ghatei, M.A.; et al. The novel hypothalamic peptide ghrelin stimulates food intake and growth hormone secretion. Endocrinology 2000, 141, 4325–4328. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.Y.; Trumbauer, M.E.; Chen, A.S.; Weingarth, D.T.; Adams, J.R.; Frazier, E.G.; Shen, Z.; Marsh, D.J.; Feighner, S.D.; Guan, X.-M.; et al. Orexigenic action of peripheral ghrelin is mediated by neuropeptide y and agouti-related protein. Endocrinology 2004, 145, 2607–2612. [Google Scholar] [CrossRef] [PubMed]
- Nagaya, N.; Uematsu, M.; Kojima, M.; Ikeda, Y.; Yoshihara, F.; Shimizu, W.; Hosoda, H.; Hirota, Y.; Ishida, H.; Mori, H.; et al. Chronic administration of ghrelin improves left ventricular dysfunction and attenuates development of cardiac cachexia in rats with heart failure. Circulation 2001, 104, 1430–1435. [Google Scholar] [CrossRef] [PubMed]
- Mericq, V.; Cassorla, F.; Bowers, C.Y.; Avila, A.; Gonen, B.; Merriam, G.R. Changes in appetite and body weight in response to long-term oral administration of the ghrelin agonist GHRP-2 in growth hormone deficient children. J. Pediatr. Endocrinol. Metab. JPEM 2003, 16, 981–985. [Google Scholar] [CrossRef] [PubMed]
- Schmid, D.A.; Held, K.; Ising, M.; Uhr, M.; Weikel, J.C.; Steiger, A. Ghrelin stimulates appetite, imagination of food, GH, ACTH, and cortisol, but does not affect leptin in normal controls. Neuropsychopharmacology 2005, 30, 1187–1192. [Google Scholar] [CrossRef] [PubMed]
- Wynne, K.; Giannitsopoulou, K.; Small, C.J.; Patterson, M.; Frost, G.; Ghatei, M.A.; Brown, E.A.; Bloom, S.R.; Choi, P. Subcutaneous ghrelin enhances acute food intake in malnourished patients who receive maintenance peritoneal dialysis: A randomized, placebo-controlled trial. J. Am. Soc. Nephrol. JASN 2005, 16, 2111–2118. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Ahmed, S.; Smith, R.G. Deletion of ghrelin impairs neither growth nor appetite. Mol. Cell. Biol. 2003, 23, 7973–7981. [Google Scholar] [CrossRef] [PubMed]
- Petersen, P.S.; Woldbye, D.P.; Madsen, A.N.; Egerod, K.L.; Jin, C.; Lang, M.; Rasmussen, M.; Beck-Sickinger, A.G.; Holst, B. In vivo characterization of high basal signaling from the ghrelin receptor. Endocrinology 2009, 150, 4920–4930. [Google Scholar] [CrossRef] [PubMed]
- Holst, B.; Schwartz, T.W. Constitutive ghrelin receptor activity as a signaling set-point in appetite regulation. Trends Pharmacol. Sci. 2004, 25, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Zigman, J.M.; Nakano, Y.; Coppari, R.; Balthasar, N.; Marcus, J.N.; Lee, C.E.; Jones, J.E.; Deysher, A.E.; Waxman, A.R.; White, R.D.; et al. Mice lacking ghrelin receptors resist the development of diet-induced obesity. J. Clin. Investig. 2005, 115, 3564–3572. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Lin, L.; Qin, G.; Lu, X.; Fiorotto, M.; Dixit, V.D.; Sun, Y. Ablations of ghrelin and ghrelin receptor exhibit differential metabolic phenotypes and thermogenic capacity during aging. PLoS ONE 2011, 6, e16391. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Lee, J.H.; Bongmba, O.Y.; Ma, X.; Zhu, X.; Sheikh-Hamad, D.; Sun, Y. The suppression of ghrelin signaling mitigates age-associated thermogenic impairment. Aging 2014, 6, 1019–1032. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Lin, L.; Xu, P.; Saito, K.; Wei, Q.; Meadows, A.G.; Bongmba, O.Y.N.; Pradhan, G.; Zheng, H.; Xu, Y.; et al. Neuronal deletion of ghrelin receptor almost completely prevents diet-induced obesity. Diabetes 2016, 65, 2169–2178. [Google Scholar] [CrossRef] [PubMed]
- Pantel, J.; Legendre, M.; Cabrol, S.; Hilal, L.; Hajaji, Y.; Morisset, S.; Nivot, S.; Vie-Luton, M.-P.; Grouselle, D.; de Kerdanet, M. Loss of constitutive activity of the growth hormone secretagogue receptor in familial short stature. J. Clin. Investig. 2006, 116, 760–768. [Google Scholar] [CrossRef] [PubMed]
- Landgren, S.; Simms, J.A.; Thelle, D.S.; Strandhagen, E.; Bartlett, S.E.; Engel, J.A.; Jerlhag, E. The ghrelin signalling system is involved in the consumption of sweets. PLoS ONE 2011, 6, e18170. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Borgland, S.L. Regulation of the mesolimbic dopamine circuit by feeding peptides. Neuroscience 2015, 289, 19–42. [Google Scholar] [CrossRef] [PubMed]
- Swanson, L.W. The projections of the ventral tegmental area and adjacent regions: A combined fluorescent retrograde tracer and immunofluorescence study in the rat. Brain Res. Bull. 1982, 9, 321–353. [Google Scholar] [CrossRef]
- Tzschentke, T.M. The medial prefrontal cortex as a part of the brain reward system. Amino Acids 2000, 19, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Landgren, S.; Engel, J.A.; Hyytia, P.; Zetterberg, H.; Blennow, K.; Jerlhag, E. Expression of the gene encoding the ghrelin receptor in rats selected for differential alcohol preference. Behav. Brain Res. 2011, 221, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Diano, S.; Farr, S.A.; Benoit, S.C.; McNay, E.C.; da Silva, I.; Horvath, B.; Gaskin, F.S.; Nonaka, N.; Jaeger, L.B.; Banks, W.A.; et al. Ghrelin controls hippocampal spine synapse density and memory performance. Nat. Neurosci. 2006, 9, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Kern, A.; Mavrikaki, M.; Ullrich, C.; Albarran-Zeckler, R.; Brantley, A.F.; Smith, R.G. Hippocampal dopamine/DRD1 signaling dependent on the ghrelin receptor. Cell 2015, 163, 1176–1190. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Crespo, M.; Skibicka, K.P.; Farkas, I.; Molnár, C.S.; Egecioglu, E.; Hrabovszky, E.; Liposits, Z.; Dickson, S.L. The amygdala as a neurobiological target for ghrelin in rats: Neuroanatomical, electrophysiological and behavioral evidence. PLoS ONE 2012, 7, e46321. [Google Scholar] [CrossRef] [PubMed]
- Sakata, I.; Nakano, Y.; Osborne-Lawrence, S.; Rovinsky, S.A.; Lee, C.E.; Perello, M.; Anderson, J.G.; Coppari, R.; Xiao, G.; Lowell, B.B.; et al. Characterization of a novel ghrelin cell reporter mouse. Regul. Pept. 2009, 155, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Furness, J.; Hunne, B.; Matsuda, N.; Yin, L.; Russo, D.; Kato, I.; Fujimiya, M.; Patterson, M.; McLeod, J.; Andrews, Z. Investigation of the presence of ghrelin in the central nervous system of the rat and mouse. Neuroscience 2011, 193, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lear, P.V.; Iglesias, M.J.; Feijoo-Bandin, S.; Rodriguez-Penas, D.; Mosquera-Leal, A.; Garcia-Rua, V.; Gualillo, O.; Ghe, C.; Arnoletti, E.; Muccioli, G.; et al. Des-acyl ghrelin has specific binding sites and different metabolic effects from ghrelin in cardiomyocytes. Endocrinology 2010, 151, 3286–3298. [Google Scholar] [CrossRef] [PubMed]
- Pang, J.J.; Xu, R.K.; Xu, X.B.; Cao, J.M.; Ni, C.; Zhu, W.L.; Asotra, K.; Chen, M.C.; Chen, C. Hexarelin protects rat cardiomyocytes from angiotensin II-induced apoptosis in vitro. Am. J. Physiol. Heart Circ. Physiol. 2004, 286, H1063–H1069. [Google Scholar] [CrossRef] [PubMed]
- Bisi, G.; Podio, V.; Valetto, M.R.; Broglio, F.; Bertuccio, G.; Del Rio, G.; Arvat, E.; Boghen, M.F.; Deghenghi, R.; Muccioli, G.; et al. Acute cardiovascular and hormonal effects of GH and hexarelin, a synthetic GH-releasing peptide, in humans. J. Endocrinol. Investig. 1999, 22, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Barreiro, M.L.; Gaytan, F.; Caminos, J.E.; Pinilla, L.; Casanueva, F.F.; Aguilar, E.; Dieguez, C.; Tena-Sempere, M. Cellular location and hormonal regulation of ghrelin expression in rat testis. Biol. Reprod. 2002, 67, 1768–1776. [Google Scholar] [CrossRef] [PubMed]
- Gaytan, F.; Barreiro, M.L.; Caminos, J.E.; Chopin, L.K.; Herington, A.C.; Morales, C.; Pinilla, L.; Paniagua, R.; Nistal, M.; Casanueva, F.F.; et al. Expression of ghrelin and its functional receptor, the type 1a growth hormone secretagogue receptor, in normal human testis and testicular tumors. J. Clin. Endocrinol. Metab. 2004, 89, 400–409. [Google Scholar] [CrossRef] [PubMed]
- De Smet, B.; Mitselos, A.; Depoortere, I. Motilin and ghrelin as prokinetic drug targets. Pharmacol. Ther. 2009, 123, 207–223. [Google Scholar] [CrossRef] [PubMed]
- Date, Y.; Murakami, N.; Toshinai, K.; Matsukura, S.; Niijima, A.; Matsuo, H.; Kangawa, K.; Nakazato, M. The role of the gastric afferent vagal nerve in ghrelin-induced feeding and growth hormone secretion in rats. Gastroenterology 2002, 123, 1120–1128. [Google Scholar] [CrossRef] [PubMed]
- Dass, N.B.; Munonyara, M.; Bassil, A.K.; Hervieu, G.J.; Osbourne, S.; Corcoran, S.; Morgan, M.; Sanger, G.J. Growth hormone secretagogue receptors in rat and human gastrointestinal tract and the effects of ghrelin. Neuroscience 2003, 120, 443–453. [Google Scholar] [CrossRef]
- Takeshita, E.; Matsuura, B.; Dong, M.; Miller, L.J.; Matsui, H.; Onji, M. Molecular characterization and distribution of motilin family receptors in the human gastrointestinal tract. J. Gastroenterol. 2006, 41, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, H.; Mizuta, Y.; Isomoto, H.; Takeshima, F.; Ohnita, K.; Ohba, K.; Omagari, K.; Taniyama, K.; Kohno, S. Ghrelin enhances gastric motility through direct stimulation of intrinsic neural pathways and capsaicin-sensitive afferent neurones in rats. Scand. J. Gastroenterol. 2004, 39, 1209–1214. [Google Scholar] [CrossRef] [PubMed]
- Depoortere, I.; De Winter, B.; Thijs, T.; De Man, J.; Pelckmans, P.; Peeters, T. Comparison of the gastroprokinetic effects of ghrelin, GHRP-6 and motilin in rats in vivo and in vitro. Eur. J. Pharmacol. 2005, 515, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Trudel, L.; Tomasetto, C.; Rio, M.C.; Bouin, M.; Plourde, V.; Eberling, P.; Poitras, P. Ghrelin/motilin-related peptide is a potent prokinetic to reverse gastric postoperative ileus in rat. Am. J. Physiol. Gastrointest. Liver Physiol. 2002, 282, G948–G952. [Google Scholar] [CrossRef] [PubMed]
- Kitazawa, T.; De Smet, B.; Verbeke, K.; Depoortere, I.; Peeters, T.L. Gastric motor effects of peptide and non-peptide ghrelin agonists in mice in vivo and in vitro. Gut 2005, 54, 1078–1084. [Google Scholar] [CrossRef] [PubMed]
- Dornonville de la Cour, C.; Lindqvist, A.; Egecioglu, E.; Tung, Y.C.; Surve, V.; Ohlsson, C.; Jansson, J.O.; Erlanson-Albertsson, C.; Dickson, S.L.; Hakanson, R. Ghrelin treatment reverses the reduction in weight gain and body fat in gastrectomised mice. Gut 2005, 54, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Levin, F.; Edholm, T.; Schmidt, P.T.; Gryback, P.; Jacobsson, H.; Degerblad, M.; Hoybye, C.; Holst, J.J.; Rehfeld, J.F.; Hellstrom, P.M.; et al. Ghrelin stimulates gastric emptying and hunger in normal-weight humans. J. Clin. Endocrinol. Metab. 2006, 91, 3296–3302. [Google Scholar] [CrossRef] [PubMed]
- Fujino, K.; Inui, A.; Asakawa, A.; Kihara, N.; Fujimura, M.; Fujimiya, M. Ghrelin induces fasted motor activity of the gastrointestinal tract in conscious fed rats. J. Physiol. 2003, 550, 227–240. [Google Scholar] [CrossRef] [PubMed]
- Asakawa, A.; Inui, A.; Kaga, O.; Yuzuriha, H.; Nagata, T.; Ueno, N.; Makino, S.; Fujimiya, M.; Niijima, A.; Fujino, M.A.; et al. Ghrelin is an appetite-stimulatory signal from stomach with structural resemblance to motilin. Gastroenterology 2001, 120, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Masuda, Y.; Tanaka, T.; Inomata, N.; Ohnuma, N.; Tanaka, S.; Itoh, Z.; Hosoda, H.; Kojima, M.; Kangawa, K. Ghrelin stimulates gastric acid secretion and motility in rats. Biochem. Biophys. Res. Commun. 2000, 276, 905–908. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.G.; Qiu, W.C.; Wang, Z.G.; Yu, S.; Yan, J.; Zheng, Q. Down-regulation of ghrelin receptors in the small intestine delays small intestinal transit in vagotomized rats. Mol. Med. Rep. 2011, 4, 1061–1065. [Google Scholar] [PubMed]
- Feighner, S.D.; Tan, C.P.; McKee, K.K.; Palyha, O.C.; Hreniuk, D.L.; Pong, S.S.; Austin, C.P.; Figueroa, D.; MacNeil, D.; Cascieri, M.A.; et al. Receptor for motilin identified in the human gastrointestinal system. Science 1999, 284, 2184–2188. [Google Scholar] [CrossRef] [PubMed]
- Nunoi, H.; Matsuura, B.; Utsunomiya, S.; Ueda, T.; Miyake, T.; Furukawa, S.; Kumagi, T.; Ikeda, Y.; Abe, M.; Hiasa, Y.; et al. A relationship between motilin and growth hormone secretagogue receptors. Regul. Pept. 2012, 176, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Mighiu, P.I.; Yue, J.T.Y.; Filippi, B.M.; Abraham, M.A.; Chari, M.; Lam, C.K.L.; Yang, C.S.; Christian, N.R.; Charron, M.J.; Lam, T.K.T. Hypothalamic glucagon signaling inhibits hepatic glucose production. Nat. Med. 2013, 19, 766–772. [Google Scholar] [CrossRef] [PubMed]
- Morton, G.J.; Schwartz, M.W. Leptin and the central nervous system control of glucose metabolism. Physiol. Rev. 2011, 91, 389–411. [Google Scholar] [CrossRef] [PubMed]
- Obici, S.; Zhang, B.B.; Karkanias, G.; Rossetti, L. Hypothalamic insulin signaling is required for inhibition of glucose production. Nat. Med. 2002, 8, 1376–1382. [Google Scholar] [CrossRef] [PubMed]
- Sandoval, D.A.; Bagnol, D.; Woods, S.C.; D’Alessio, D.A.; Seeley, R.J. Arcuate glucagon-like peptide 1 receptors regulate glucose homeostasis but not food intake. Diabetes 2008, 57, 2046–2054. [Google Scholar] [CrossRef] [PubMed]
- Dezaki, K.; Hosoda, H.; Kakei, M.; Hashiguchi, S.; Watanabe, M.; Kangawa, K.; Yada, T. Endogenous ghrelin in pancreatic islets restricts insulin release by attenuating Ca2+ signaling in β-cells: Implication in the glycemic control in rodents. Diabetes 2004, 53, 3142–3151. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, H.; Funahashi, H.; Hirayama, M.; Takenoya, F.; Kita, T.; Kato, S.; Sakurai, J.; Lee, E.Y.; Inoue, S.; Date, Y.; et al. Morphological analysis of ghrelin and its receptor distribution in the rat pancreas. Regul. Pept. 2005, 126, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Date, Y.; Nakazato, M.; Hashiguchi, S.; Dezaki, K.; Mondal, M.S.; Hosoda, H.; Kojima, M.; Kangawa, K.; Arima, T.; Matsuo, H.; et al. Ghrelin is present in pancreatic α-cells of humans and rats and stimulates insulin secretion. Diabetes 2002, 51, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Tong, J.; Prigeon, R.L.; Davis, H.W.; Bidlingmaier, M.; Kahn, S.E.; Cummings, D.E.; Tschop, M.H.; D’Alessio, D. Ghrelin suppresses glucose-stimulated insulin secretion and deteriorates glucose tolerance in healthy humans. Diabetes 2010, 59, 2145–2151. [Google Scholar] [CrossRef] [PubMed]
- Reimer, M.K.; Pacini, G.; Ahren, B. Dose-dependent inhibition by ghrelin of insulin secretion in the mouse. Endocrinology 2003, 144, 916–921. [Google Scholar] [CrossRef] [PubMed]
- Broglio, F.; Arvat, E.; Benso, A.; Gottero, C.; Muccioli, G.; Papotti, M.; van der Lely, A.J.; Deghenghi, R.; Ghigo, E. Ghrelin, a natural GH secretagogue produced by the stomach, induces hyperglycemia and reduces insulin secretion in humans. J. Clin. Endocrinol. Metab. 2001, 86, 5083–5086. [Google Scholar] [CrossRef] [PubMed]
- Broglio, F.; Prodam, F.; Riganti, F.; Gottero, C.; Destefanis, S.; Granata, R.; Muccioli, G.; Abribat, T.; van der Lely, A.J.; Ghigo, E. The continuous infusion of acylated ghrelin enhances growth hormone secretion and worsens glucose metabolism in humans. J. Endocrinol. Investig. 2008, 31, 788–794. [Google Scholar] [CrossRef] [PubMed]
- Garin, M.C.; Burns, C.M.; Kaul, S.; Cappola, A.R. Clinical review: The human experience with ghrelin administration. J. Clin. Endocrinol. Metab. 2013, 98, 1826–1837. [Google Scholar] [CrossRef] [PubMed]
- Nesic, D.M.; Stevanovic, D.M.; Ille, T.; Petricevic, S.; Masirevic-Draskovic, G.; Starcevic, V.P. Centrally applied ghrelin affects feeding dynamics in male rats. J. Physiol. Pharmacol. 2008, 59, 489–500. [Google Scholar] [PubMed]
- Kim, M.S.; Namkoong, C.; Kim, H.S.; Jang, P.G.; Kim Pak, Y.M.; Katakami, H.; Park, J.Y.; Lee, K.U. Chronic central administration of ghrelin reverses the effects of leptin. Int. J. Obes. Relat. Metab. Disord. 2004, 28, 1264–1271. [Google Scholar] [CrossRef] [PubMed]
- Heppner, K.M.; Piechowski, C.L.; Muller, A.; Ottaway, N.; Sisley, S.; Smiley, D.L.; Habegger, K.M.; Pfluger, P.T.; Dimarchi, R.; Biebermann, H.; et al. Both acyl and des-acyl ghrelin regulate adiposity and glucose metabolism via central nervous system ghrelin receptors. Diabetes 2014, 63, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Stark, R.; Reichenbach, A.; Lockie, S.H.; Pracht, C.; Wu, Q.; Tups, A.; Andrews, Z.B. Acyl ghrelin acts in the brain to control liver function and peripheral glucose homeostasis in male mice. Endocrinology 2015, 156, 858–868. [Google Scholar] [CrossRef] [PubMed]
- Meyer, C. Final answer: Ghrelin can suppress insulin secretion in humans, but is it clinically relevant? Diabetes 2010, 59, 2726–2728. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.; Ohnuma, H.; Daimon, M.; Susa, S.; Yamaguchi, H.; Kameda, W.; Jimbu, Y.; Oizumi, T.; Kato, T. Ghrelin infused into the portal vein inhibits glucose-stimulated insulin secretion in Wistar rats. Peptides 2008, 29, 1241–1246. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Butte, N.F.; Garcia, J.M.; Smith, R.G. Characterization of adult ghrelin and ghrelin receptor knockout mice under positive and negative energy balance. Endocrinology 2008, 149, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Saha, P.K.; Ma, X.; Henshaw, I.O.; Shao, L.; Chang, B.H.; Buras, E.D.; Tong, Q.; Chan, L.; McGuinness, O.P.; et al. Ablation of ghrelin receptor reduces adiposity and improves insulin sensitivity during aging by regulating fat metabolism in white and brown adipose tissues. Aging Cell 2011, 10, 996–1010. [Google Scholar] [CrossRef] [PubMed]
- Chabot, F.; Caron, A.; Laplante, M.; St-Pierre, D.H. Interrelationships between ghrelin, insulin and glucose homeostasis: Physiological relevance. World J. Diabetes 2014, 5, 328–341. [Google Scholar] [CrossRef] [PubMed]
- Woods, S.C.; Lutz, T.A.; Geary, N.; Langhans, W. Pancreatic signals controlling food intake; insulin, glucagon and amylin. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 1219–1235. [Google Scholar] [CrossRef] [PubMed]
- Lavin, J.H.; Wittert, G.; Sun, W.M.; Horowitz, M.; Morley, J.E.; Read, N.W. Appetite regulation by carbohydrate: Role of blood glucose and gastrointestinal hormones. Am. J. Physiol. 1996, 271, E209–E214. [Google Scholar] [PubMed]
- Flint, A.; Gregersen, N.T.; Gluud, L.L.; Møller, B.K.; Raben, A.; Tetens, I.; Verdich, C.; Astrup, A. Associations between postprandial insulin and blood glucose responses, appetite sensations and energy intake in normal weight and overweight individuals: A meta-analysis of test meal studies. Br. J. Nutr. 2007, 98, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Ferrini, F.; Salio, C.; Lossi, L.; Merighi, A. Ghrelin in central neurons. Curr. Neuropharmacol. 2009, 7, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Guneli, E.; Kazikdas, K.C.; Kolatan, E. Ghrelin may attenuate proinflammatory cytokine-mediated neuropathic pain. Med. Hypotheses 2007, 69, 356–360. [Google Scholar] [CrossRef] [PubMed]
- Vergnano, A.M.; Ferrini, F.; Salio, C.; Lossi, L.; Baratta, M.; Merighi, A. The gastrointestinal hormone ghrelin modulates inhibitory neurotransmission in deep laminae of mouse spinal cord dorsal horn. Endocrinology 2008, 149, 2306–2312. [Google Scholar] [CrossRef] [PubMed]
- Guneli, E.; Onal, A.; Ates, M.; Bagriyanik, H.A.; Resmi, H.; Orhan, C.E.; Kolatan, H.E.; Gumustekin, M. Effects of repeated administered ghrelin on chronic constriction injury of the sciatic nerve in rats. Neurosci. Lett. 2010, 479, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Azizzadeh, F.; Mahmoodi, J.; Sadigh-Eteghad, S.; Farajdokht, F.; Mohaddes, G. Ghrelin exerts analgesic effects through modulation of IL-10 and TGF-β levels in a rat model of inflammatory pain. Iran. Biomed. J. 2017, 21, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Sibilia, V.; Lattuada, N.; Rapetti, D.; Pagani, F.; Vincenza, D.; Bulgarelli, I.; Locatelli, V.; Guidobono, F.; Netti, C. Ghrelin inhibits inflammatory pain in rats: Involvement of the opioid system. Neuropharmacology 2006, 51, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.-M.; Yu, H.; Palyha, O.C.; McKee, K.K.; Feighner, S.D.; Sirinathsinghji, D.J.S.; Smith, R.G.; van der Ploeg, L.H.T.; Howard, A.D. Distribution of mRNA encoding the growth hormone secretagogue receptor in brain and peripheral tissues. Mol. Brain Res. 1997, 48, 23–29. [Google Scholar] [CrossRef]
- Hou, Z.; Miao, Y.; Gao, L.; Pan, H.; Zhu, S. Ghrelin-containing neuron in cerebral cortex and hypothalamus linked with the DVC of brainstem in rat. Regul. Pept. 2006, 134, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.H.; Li, X.; Zhu, Y.Z.; Huang, H.; Li, J.; Liu, L.; Hu, Q.; Ma, T.F.; Shao, Y.; Wu, Y.Q. Ghrelin alleviates neuropathic pain through GHSR-1a-mediated suppression of the p38 MAPK/NF-KappaB pathway in a rat chronic constriction injury model. Reg. Anesth. Pain Med. 2014, 39, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Sibilia, V.; Pagani, F.; Mrak, E.; Dieci, E.; Tulipano, G.; Ferrucci, F. Pharmacological characterization of the ghrelin receptor mediating its inhibitory action on inflammatory pain in rats. Amino Acids 2012, 43, 1751–1759. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Zhi, X.; Wang, X.L.; Zeng, P.; Zou, T.; Yang, B.; Wang, J.L. In vivo characterization of the effects of ghrelin on the modulation of acute pain at the supraspinal level in mice. Peptides 2013, 43, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Luttrell, L. Reviews in molecular biology and biotechnology: Transmembrane signaling by g protein-coupled receptors. Mol. Biotechnol. 2008, 39, 239–264. [Google Scholar] [CrossRef] [PubMed]
- Kohno, D.; Gao, H.Z.; Muroya, S.; Kikuyama, S.; Yada, T. Ghrelin directly interacts with neuropeptide-Y-containing neurons in the rat arcuate nucleus: Ca2+ signaling via protein kinase A and N-type channel-dependent mechanisms and cross-talk with leptin and orexin. Diabetes 2003, 52, 948–956. [Google Scholar] [CrossRef] [PubMed]
- Malagon, M.M.; Luque, R.M.; Ruiz-Guerrero, E.; Rodriguez-Pacheco, F.; Garcia-Navarro, S.; Casanueva, F.F.; Gracia-Navarro, F.; Castano, J.P. Intracellular signaling mechanisms mediating ghrelin-stimulated growth hormone release in somatotropes. Endocrinology 2003, 144, 5372–5380. [Google Scholar] [CrossRef] [PubMed]
- Dezaki, K.; Kakei, M.; Yada, T. Ghrelin uses Galphai2 and activates voltage-dependent K+ channels to attenuate glucose-induced Ca2+ signaling and insulin release in islet β-cells: Novel signal transduction of ghrelin. Diabetes 2007, 56, 2319–2327. [Google Scholar] [CrossRef] [PubMed]
- Schellekens, H.; van Oeffelen, W.E.; Dinan, T.G.; Cryan, J.F. Promiscuous dimerization of the growth hormone secretagogue receptor (GHS-R1a) attenuates ghrelin-mediated signaling. J. Biol. Chem. 2013, 288, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Irani, B.G.; Xiang, Z.; Yarandi, H.N.; Holder, J.R.; Moore, M.C.; Bauzo, R.M.; Proneth, B.; Shaw, A.M.; Millard, W.J.; Chambers, J.B.; et al. Implication of the melanocortin-3 receptor in the regulation of food intake. Eur. J. Pharmacol. 2011, 660, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Rediger, A.; Piechowski, C.L.; Yi, C.X.; Tarnow, P.; Strotmann, R.; Gruters, A.; Krude, H.; Schoneberg, T.; Tschop, M.H.; Kleinau, G.; et al. Mutually opposite signal modulation by hypothalamic heterodimerization of ghrelin and melanocortin-3 receptors. J. Biol. Chem. 2011, 286, 39623–39631. [Google Scholar] [CrossRef] [PubMed]
- Garfield, A.S.; Heisler, L.K. Pharmacological targeting of the serotonergic system for the treatment of obesity. J. Physiol. 2009, 587, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Dutton, A.C.; Barnes, N.M. Anti-obesity pharmacotherapy: Future perspectives utilising 5-HT2C receptor agonists. Drug Discov. Today Ther. Strateg. 2006, 3, 577–583. [Google Scholar] [CrossRef]
- Miller, K.J. Serotonin 5-HT2C receptor agonists: Potential for the treatment of obesity. Mol. Int. 2005, 5, 282–291. [Google Scholar] [CrossRef] [PubMed]
- Kern, A.; Albarran-Zeckler, R.; Walsh, H.E.; Smith, R.G. Apo-ghrelin receptor forms heteromers with DRD2 in hypothalamic neurons and is essential for anorexigenic effects of DRD2 agonism. Neuron 2012, 73, 317–332. [Google Scholar] [CrossRef] [PubMed]
- Chow, K.B.; Sun, J.; Chu, K.M.; Tai Cheung, W.; Cheng, C.H.; Wise, H. The truncated ghrelin receptor polypeptide (GHS-R1b) is localized in the endoplasmic reticulum where it forms heterodimers with ghrelin receptors (GHS-R1a) to attenuate their cell surface expression. Mol. Cell. Endocrinol. 2012, 348, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Mear, Y.; Enjalbert, A.; Thirion, S. GHS-R1a constitutive activity and its physiological relevance. Front. Neurosci. 2013, 7, 87. [Google Scholar] [CrossRef] [PubMed]
- Tsao, P.; von Zastrow, M. Downregulation of G protein-coupled receptors. Curr. Opin. Neurobiol. 2000, 10, 365–369. [Google Scholar] [CrossRef]
- Kaji, H.; Kishimoto, M.; Kirimura, T.; Iguchi, G.; Murata, M.; Yoshioka, S.; Iida, K.; Okimura, Y.; Yoshimoto, Y.; Chihara, K. Hormonal regulation of the human ghrelin receptor gene transcription. Biochem. Biophys. Res. Commun. 2001, 284, 660–666. [Google Scholar] [CrossRef] [PubMed]
- Orkin, R.D.; New, D.I.; Norman, D.; Chew, S.L.; Clark, A.J.; Grossman, A.B.; Korbonits, M. Rapid desensitisation of the GH secretagogue (ghrelin) receptor to hexarelin in vitro. J. Endocrinol. Investig. 2003, 26, 743–747. [Google Scholar] [CrossRef] [PubMed]
- Camina, J.P.; Carreira, M.C.; El Messari, S.; Llorens-Cortes, C.; Smith, R.G.; Casanueva, F.F. Desensitization and endocytosis mechanisms of ghrelin-activated growth hormone secretagogue receptor 1a. Endocrinology 2004, 145, 930–940. [Google Scholar] [CrossRef] [PubMed]
- Guyda, H.J. Human growth hormone: Research and clinical practice. Edited by Roy G Smith and Michael O Thorner. Q. Rev. Biol. 2002, 77, 110–111. [Google Scholar] [CrossRef]
- Gardiner, J.; Bloom, S. Ghrelin gets its GOAT. Cell Metab. 2008, 7, 193–194. [Google Scholar] [CrossRef] [PubMed]
- Lundholm, K.; Gunnebo, L.; Korner, U.; Iresjo, B.M.; Engstrom, C.; Hyltander, A.; Smedh, U.; Bosaeus, I. Effects by daily long term provision of ghrelin to unselected weight-losing cancer patients: A randomized double-blind study. Cancer 2010, 116, 2044–2052. [Google Scholar] [CrossRef] [PubMed]
- Neary, N.M.; Druce, M.R.; Small, C.J.; Bloom, S.R. Acylated ghrelin stimulates food intake in the fed and fasted states but desacylated ghrelin has no effect. Gut 2006, 55, 135. [Google Scholar] [PubMed]
- Wren, A.M.; Seal, L.J.; Cohen, M.A.; Brynes, A.E.; Frost, G.S.; Murphy, K.G.; Dhillo, W.S.; Ghatei, M.A.; Bloom, S.R. Ghrelin enhances appetite and increases food intake in humans. J. Clin. Endocrinol. Metab. 2001, 86, 5992. [Google Scholar] [CrossRef] [PubMed]
- Hewson, A.K.; Tung, L.Y.; Connell, D.W.; Tookman, L.; Dickson, S.L. The rat arcuate nucleus integrates peripheral signals provided by leptin, insulin, and a ghrelin mimetic. Diabetes 2002, 51, 3412–3419. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Guan, J.L.; Wang, Q.P.; Uehara, K.; Yamada, S.; Goto, N.; Date, Y.; Nakazato, M.; Kojima, M.; Kangawa, K.; et al. Immunocytochemical observation of ghrelin-containing neurons in the rat arcuate nucleus. Neurosci. Lett. 2002, 321, 157–160. [Google Scholar] [CrossRef]
- Kageyama, H.; Kitamura, Y.; Hosono, T.; Kintaka, Y.; Seki, M.; Takenoya, F.; Hori, Y.; Nonaka, N.; Arata, S.; Shioda, S. Visualization of ghrelin-producing neurons in the hypothalamic arcuate nucleus using ghrelin-EGFP transgenic mice. Regul. Pept. 2008, 145, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Cabral, A.; de Francesco, P.N.; Perello, M. Brain circuits mediating the orexigenic action of peripheral ghrelin: Narrow gates for a vast kingdom. Front. Endocrinol. 2015, 6, 44. [Google Scholar] [CrossRef] [PubMed]
- Inui, A. Ghrelin: An orexigenic and somatotrophic signal from the stomach. Nat. Rev. Neurosci. 2001, 2, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Banks, W.A. Extent and direction of ghrelin transport across the blood-brain barrier is determined by its unique primary structure. J. Pharmacol. Exp. Ther. 2002, 302, 822–827. [Google Scholar] [CrossRef] [PubMed]
- Takayama, K.; Johno, Y.; Hayashi, K.; Yakabi, K.; Tanaka, T.; Ro, S. Expression of c-Fos protein in the brain after intravenous injection of ghrelin in rats. Neurosci. Lett. 2007, 417, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Saint-Pierre, D.H.; Tache, Y. Peripheral ghrelin selectively increases Fos expression in neuropeptide Y—Synthesizing neurons in mouse hypothalamic arcuate nucleus. Neurosci. Lett. 2002, 325, 47–51. [Google Scholar] [CrossRef]
- Ciofi, P.; Garret, M.; Lapirot, O.; Lafon, P.; Loyens, A.; Prevot, V.; Levine, J.E. Brain-endocrine interactions: A microvascular route in the mediobasal hypothalamus. Endocrinology 2009, 150, 5509–5519. [Google Scholar] [CrossRef] [PubMed]
- Cabral, A.; Valdivia, S.; Fernandez, G.; Reynaldo, M.; Perello, M. Divergent neuronal circuitries underlying acute orexigenic effects of peripheral or central ghrelin: Critical role of brain accessibility. J. Neuroendocrinol. 2014, 26, 542–554. [Google Scholar] [CrossRef] [PubMed]
- Redzic, Z.B.; Preston, J.E.; Duncan, J.A.; Chodobski, A.; Szmydynger—Chodobska, J. The choroid plexus—Cerebrospinal fluid system: From development to aging. Curr. Top. Dev. Biol. 2005, 71, 1–52. [Google Scholar] [PubMed]
- Bolborea, M.; Dale, N. Hypothalamic tanycytes: Potential roles in the control of feeding and energy balance. Trends Neurosci. 2013, 36, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Grill, H.J.; Hayes, M.R. Hindbrain neurons as an essential hub in the neuroanatomically distributed control of energy balance. Cell Metab. 2012, 16, 296–309. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.; Jerome, C.; Cushin, B.; Eterno, R.; Simansky, K. Abdominal vagotomy blocks the satiety effect of cholecystokinin in the rat. Science 1981, 213, 1036–1037. [Google Scholar] [CrossRef] [PubMed]
- Koda, S.; Date, Y.; Murakami, N.; Shimbara, T.; Hanada, T.; Toshinai, K.; Niijima, A.; Furuya, M.; Inomata, N.; Osuye, K. The role of the vagal nerve in peripheral pyy3–36-induced feeding reduction in rats. Endocrinology 2005, 146, 2369–2375. [Google Scholar] [CrossRef] [PubMed]
- Abbott, C.; Small, C.; Sajedi, A.; Smith, K.; Parkinson, J.; Broadhead, L.; Ghatei, M.; Bloom, S. The importance of acclimatisation and habituation to experimental conditions when investigating the anorectic effects of gastrointestinal hormones in the rat. Int. J. Obes. 2006, 30, 288–292. [Google Scholar] [CrossRef] [PubMed]
- Rogers, R.C.; Hermann, G.E. Mechanisms of action of CCK to activate central vagal afferent terminals. Peptides 2008, 29, 1716–1725. [Google Scholar] [CrossRef] [PubMed]
- Bailey, A.R.; Smith, R.G.; Leng, G. The nonpeptide growth hormone secretagogue, MK-0677, activates hypothalamic arcuate nucleus neurons in vivo. J. Neuroendocrinol. 1998, 10, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.G.; Sun, Y.; Jiang, H.; Albarran-Zeckler, R.; Timchenko, N. Ghrelin receptor (GHS-R1a) agonists show potential as interventive agents during aging. Ann. N. Y. Acad. Sci. 2007, 1119, 147–164. [Google Scholar] [CrossRef] [PubMed]
- Date, Y. Ghrelin and the vagus nerve. Methods Enzymol. 2012, 514, 261–269. [Google Scholar] [PubMed]
- Date, Y.; Shimbara, T.; Koda, S.; Toshinai, K.; Ida, T.; Murakami, N.; Miyazato, M.; Kokame, K.; Ishizuka, Y.; Ishida, Y.; et al. Peripheral ghrelin transmits orexigenic signals through the noradrenergic pathway from the hindbrain to the hypothalamus. Cell Metab. 2006, 4, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Le Roux, C.W.; Neary, N.M.; Halsey, T.J.; Small, C.J.; Martinez-Isla, A.M.; Ghatei, M.A.; Theodorou, N.A.; Bloom, S.R. Ghrelin does not stimulate food intake in patients with surgical procedures involving vagotomy. J. Clin. Endocrinol. Metab. 2005, 90, 4521–4524. [Google Scholar] [CrossRef] [PubMed]
- Arnold, M.; Mura, A.; Langhans, W.; Geary, N. Gut vagal afferents are not necessary for the eating-stimulatory effect of intraperitoneally injected ghrelin in the rat. J. Neurosci. 2006, 26, 11052–11060. [Google Scholar] [CrossRef] [PubMed]
- Takeno, R.; Okimura, Y.; Iguchi, G.; Kishimoto, M.; Kudo, T.; Takahashi, K.; Takahashi, Y.; Kaji, H.; Ohno, M.; Ikuta, H.; et al. Intravenous administration of ghrelin stimulates growth hormone secretion in vagotomized patients as well as normal subjects. Eur. J. Endocrinol. 2004, 151, 447–450. [Google Scholar] [CrossRef] [PubMed]
- Akamizu, T.; Takaya, K.; Irako, T.; Hosoda, H.; Teramukai, S.; Matsuyama, A.; Tada, H.; Miura, K.; Shimizu, A.; Fukushima, M.; et al. Pharmacokinetics, safety, and endocrine and appetite effects of ghrelin administration in young healthy subjects. Eur. J. Endocrinol. 2004, 150, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Akamizu, T.; Iwakura, H.; Ariyasu, H.; Hosoda, H.; Murayama, T.; Yokode, M.; Teramukai, S.; Seno, H.; Chiba, T.; Noma, S. Repeated administration of ghrelin to patients with functional dyspepsia: Its effects on food intake and appetite. Eur. J. Endocrinol. 2008, 158, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Ashby, D.R.; Ford, H.E.; Wynne, K.J.; Wren, A.M.; Murphy, K.G.; Busbridge, M.; Brown, E.A.; Taube, D.H.; Ghatei, M.A.; Tam, F.W.K.; et al. Sustained appetite improvement in malnourished dialysis patients by daily ghrelin treatment. Kidney Int. 2009, 76, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Paulo, R.C.; Brundage, R.; Cosma, M.; Mielke, K.L.; Bowers, C.Y.; Veldhuis, J.D. Estrogen elevates the peak overnight production rate of acylated ghrelin. J. Clin. Endocrinol. Metab. 2008, 93, 4440–4447. [Google Scholar] [CrossRef] [PubMed]
- Veldhuis, J.D.; Reynolds, G.A.; Iranmanesh, A.; Bowers, C.Y. Twenty-four hour continuous ghrelin infusion augments physiologically pulsatile, nycthemeral, and entropic (feedback-regulated) modes of growth hormone secretion. J. Clin. Endocrinol. Metab. 2008, 93, 3597–3603. [Google Scholar] [CrossRef] [PubMed]
- Rigamonti, A.; Pincelli, A.; Corra, B.; Viarengo, R.; Bonomo, S.; Galimberti, D.; Scacchi, M.; Scarpini, E.; Cavagnini, F.; Muller, E. Plasma ghrelin concentrations in elderly subjects: Comparison with anorexic and obese patients. J. Endocrinol. 2002, 175, R1–R5. [Google Scholar] [CrossRef] [PubMed]
- Strasser, F.; Lutz, T.A.; Maeder, M.T.; Thuerlimann, B.; Bueche, D.; Tschop, M.; Kaufmann, K.; Holst, B.; Brandle, M.; von Moos, R.; et al. Safety, tolerability and pharmacokinetics of intravenous ghrelin for cancer-related anorexia/cachexia: A randomised, placebo-controlled, double-blind, double-crossover study. Br. J. Cancer 2008, 98, 300–308. [Google Scholar] [CrossRef] [PubMed]
- Lippl, F.; Erdmann, J.; Steiger, A.; Lichter, N.; Czogalla-Peter, C.; Bidlingmaier, M.; Tholl, S.; Schusdziarra, V. Low-dose ghrelin infusion—Evidence against a hormonal role in food intake. Regul. Pept. 2012, 174, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Tong, J.; Dave, N.; Mugundu, G.M.; Davis, H.W.; Gaylinn, B.D.; Thorner, M.O.; Tschop, M.H.; D’Alessio, D.; Desai, P.B. The pharmacokinetics of acyl, des-acyl, and total ghrelin in healthy human subjects. Eur. J. Endocrinol. 2013, 168, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Delhanty, P.J.; Neggers, S.J.; van der Lely, A.J. Should we consider des-acyl ghrelin as a separate hormone and if so, what does it do? Front. Horm. Res. 2014, 42, 163–174. [Google Scholar] [PubMed]
- Vestergaard, E.T.; Hansen, T.K.; Gormsen, L.C.; Jakobsen, P.; Moller, N.; Christiansen, J.S.; Jorgensen, J.O. Constant intravenous ghrelin infusion in healthy young men: Clinical pharmacokinetics and metabolic effects. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E1829–E1836. [Google Scholar] [CrossRef] [PubMed]
- Nagaya, N.; Kojima, M.; Uematsu, M.; Yamagishi, M.; Hosoda, H.; Oya, H.; Hayashi, Y.; Kangawa, K. Hemodynamic and hormonal effects of human ghrelin in healthy volunteers. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 280, R1483–R1487. [Google Scholar] [PubMed]
- Lucidi, P.; Murdolo, G.; Di Loreto, C.; Parlanti, N.; de Cicco, A.; Fatone, C.; Taglioni, C.; Fanelli, C.; Broglio, F.; Ghigo, E.; et al. Metabolic and endocrine effects of physiological increments in plasma ghrelin concentrations. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Druce, M.R.; Neary, N.M.; Small, C.J.; Milton, J.; Monteiro, M.; Patterson, M.; Ghatei, M.A.; Bloom, S.R. Subcutaneous administration of ghrelin stimulates energy intake in healthy lean human volunteers. Int. J. Obes. 2006, 30, 293–296. [Google Scholar] [CrossRef] [PubMed]
- Enomoto, M.; Nagaya, N.; Uematsu, M.; Okumura, H.; Nakagawa, E.; Ono, F.; Hosoda, H.; Oya, H.; Kojima, M.; Kanmatsuse, K.; et al. Cardiovascular and hormonal effects of subcutaneous administration of ghrelin, a novel growth hormone-releasing peptide, in healthy humans. Clin. Sci. 2003, 105, 431–435. [Google Scholar] [CrossRef] [PubMed]
- Delhanty, P.J.; Neggers, S.J.; van der Lely, A.J. Mechanisms in endocrinology: Ghrelin: The differences between acyl-and des-acyl ghrelin. Eur. J. Endocrinol. 2012, 167, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Stark, R.; Santos, V.V.; Geenen, B.; Cabral, A.; Dinan, T.; Bayliss, J.A.; Lockie, S.H.; Reichenbach, A.; Lemus, M.B.; Perello, M.; et al. Des-Acyl ghrelin and ghrelin o-acyltransferase regulate hypothalamic-pituitary-adrenal axis activation and anxiety in response to acute stress. Endocrinology 2016, 157, 3946–3957. [Google Scholar] [CrossRef] [PubMed]
- Bayliss, J.A.; Andrews, Z.B. Ghrelin is neuroprotective in Parkinson’s disease: Molecular mechanisms of metabolic neuroprotection. Ther. Adv. Endocrinol. Metab. 2013, 4, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Bayliss, J.A.; Lemus, M.; Santos, V.V.; Deo, M.; Elsworth, J.D.; Andrews, Z.B. Acylated but not des-acyl ghrelin is neuroprotective in an MPTP mouse model of Parkinson’s disease. J. Neurochem. 2016, 137, 460–471. [Google Scholar] [CrossRef] [PubMed]
- Andrews, Z.B.; Erion, D.; Beiler, R.; Liu, Z.W.; Abizaid, A.; Zigman, J.; Elsworth, J.D.; Savitt, J.M.; DiMarchi, R.; Tschoep, M.; et al. Ghrelin promotes and protects nigrostriatal dopamine function via a UCP2-dependent mitochondrial mechanism. J. Neurosci. 2009, 29, 14057–14065. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Prudom, C.E.; Nass, R.; Pezzoli, S.S.; Oliveri, M.C.; Johnson, M.L.; Veldhuis, P.; Gordon, D.A.; Howard, A.D.; Witcher, D.R.; et al. Novel ghrelin assays provide evidence for independent regulation of ghrelin acylation and secretion in healthy young men. J. Clin. Endocrinol. Metab. 2008, 93, 1980–1987. [Google Scholar] [CrossRef] [PubMed]
- Bednarek, M.A.; Feighner, S.D.; Pong, S.S.; McKee, K.K.; Hreniuk, D.L.; Silva, M.V.; Warren, V.A.; Howard, A.D.; van der Ploeg, L.H.; Heck, J.V. Structure-function studies on the new growth hormone-releasing peptide, ghrelin: Minimal sequence of ghrelin necessary for activation of growth hormone secretagogue receptor 1a. J. Med. Chem. 2000, 43, 4370–4376. [Google Scholar] [CrossRef] [PubMed]
- Veldhuis, J.D.; Bowers, C.Y. Integrating GHS into the ghrelin system. Int. J. Pept. 2010, 2010, 879503. [Google Scholar] [CrossRef] [PubMed]
- Gauna, C.; van de Zande, B.; van Kerkwijk, A.; Themmen, A.P.; van der Lely, A.-J.; Delhanty, P.J. Unacylated ghrelin is not a functional antagonist but a full agonist of the type 1a growth hormone secretagogue receptor (GHS-R). Mol. Cell. Endocrinol. 2007, 274, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Broglio, F.; Gottero, C.; Prodam, F.; Gauna, C.; Muccioli, G.; Papotti, M.; Abribat, T.; van der Lely, A.J.; Ghigo, E. Non-acylated ghrelin counteracts the metabolic but not the neuroendocrine response to acylated ghrelin in humans. J. Clin. Endocrinol. Metab. 2004, 89, 3062–3065. [Google Scholar] [CrossRef] [PubMed]
- Benso, A.; St-Pierre, D.H.; Prodam, F.; Gramaglia, E.; Granata, R.; van der Lely, A.J.; Ghigo, E.; Broglio, F. Metabolic effects of overnight continuous infusion of unacylated ghrelin in humans. Eur. J. Endocrinol. 2012, 166, 911–916. [Google Scholar] [CrossRef] [PubMed]
- Gauna, C.; Meyler, F.M.; Janssen, J.A.; Delhanty, P.J.; Abribat, T.; van Koetsveld, P.; Hofland, L.J.; Broglio, F.; Ghigo, E.; van der Lely, A.J. Administration of acylated ghrelin reduces insulin sensitivity, whereas the combination of acylated plus unacylated ghrelin strongly improves insulin sensitivity. J. Clin. Endocrinol. Metab. 2004, 89, 5035–5042. [Google Scholar] [CrossRef] [PubMed]
- Inhoff, T.; Monnikes, H.; Noetzel, S.; Stengel, A.; Goebel, M.; Dinh, Q.T.; Riedl, A.; Bannert, N.; Wisser, A.S.; Wiedenmann, B.; et al. Desacyl ghrelin inhibits the orexigenic effect of peripherally injected ghrelin in rats. Peptides 2008, 29, 2159–2168. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Salehi, A.; Rehfeld, J.F.; Hoglund, P.; Lindstrom, E.; Hakanson, R. Proghrelin peptides: Desacyl ghrelin is a powerful inhibitor of acylated ghrelin, likely to impair physiological effects of acyl ghrelin but not of obestatin a study of pancreatic polypeptide secretion from mouse islets. Regul. Pept. 2010, 164, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Toshinai, K.; Yamaguchi, H.; Sun, Y.; Smith, R.G.; Yamanaka, A.; Sakurai, T.; Date, Y.; Mondal, M.S.; Shimbara, T.; Kawagoe, T.; et al. Des-acyl ghrelin induces food intake by a mechanism independent of the growth hormone secretagogue receptor. Endocrinology 2006, 147, 2306–2314. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Inui, A.; Asakawa, A.; Fujino, K.; Kato, I.; Chen, C.C.; Ueno, N.; Fujimiya, M. Des-acyl ghrelin acts by crf type 2 receptors to disrupt fasted stomach motility in conscious rats. Gastroenterology 2005, 129, 8–25. [Google Scholar] [CrossRef] [PubMed]
- Soares, J.-B.; Leite-Moreira, A.F. Ghrelin, des-acyl ghrelin and obestatin: Three pieces of the same puzzle. Peptides 2008, 29, 1255–1270. [Google Scholar] [CrossRef] [PubMed]
- Bang, A.S.; Soule, S.G.; Yandle, T.G.; Richards, A.M.; Pemberton, C.J. Characterisation of proghrelin peptides in mammalian tissue and plasma. J. Endocrinol. 2007, 192, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Hosoda, H.; Kojima, M.; Mizushima, T.; Shimizu, S.; Kangawa, K. Structural divergence of human ghrelin. Identification of multiple ghrelin-derived molecules produced by post-translational processing. J. Biol. Chem. 2003, 278, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Takagi, K.; Legrand, R.; Asakawa, A.; Amitani, H.; François, M.; Tennoune, N.; Coëffier, M.; Claeyssens, S.; Do Rego, J.-C.; Déchelotte, P. Anti-ghrelin immunoglobulins modulate ghrelin stability and its orexigenic effect in obese mice and humans. Nat. Commun. 2013, 4, 2685. [Google Scholar] [CrossRef] [PubMed]
- Kuppens, R.J.; Diene, G.; Bakker, N.E.; Molinas, C.; Faye, S.; Nicolino, M.; Bernoux, D.; Delhanty, P.J.; van der Lely, A.J.; Allas, S.; et al. Elevated ratio of acylated to unacylated ghrelin in children and young adults with prader-willi syndrome. Endocrine 2015, 50, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Satou, M.; Nishi, Y.; Yoh, J.; Hattori, Y.; Sugimoto, H. Identification and characterization of acyl-protein thioesterase 1/lysophospholipase I as a ghrelin deacylation/lysophospholipid hydrolyzing enzyme in fetal bovine serum and conditioned medium. Endocrinology 2010, 151, 4765–4775. [Google Scholar] [CrossRef] [PubMed]
- Kirchner, H.; Gutierrez, J.A.; Solenberg, P.J.; Pfluger, P.T.; Czyzyk, T.A.; Willency, J.A.; Schurmann, A.; Joost, H.G.; Jandacek, R.J.; Hale, J.E.; et al. GOAT links dietary lipids with the endocrine control of energy balance. Nat. Med. 2009, 15, 741–745. [Google Scholar] [CrossRef] [PubMed]
- Moulin, A.; Brunel, L.; Verdie, P.; Gavara, L.; Martinez, J.; Fehrentz, J.A. Ghrelin receptor ligands: Design and synthesis of pseudopeptides and peptidomimetics. Curr. Chem. Biol. 2013, 7, 254–270. [Google Scholar] [CrossRef]
- Vodnik, M.; Štrukelj, B.; Lunder, M. Ghrelin receptor ligands reaching clinical trials: From peptides to peptidomimetics; from agonists to antagonists. Horm. Metab. Res. 2016, 48, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Broglio, F.; Benso, A.; Castiglioni, C.; Gottero, C.; Prodam, F.; Destefanis, S.; Gauna, C.; Lely, A.J.; Deghenghi, R.; Bo, M.; et al. The endocrine response to ghrelin as a function of gender in humans in young and elderly subjects. J. Clin. Endocrinol. Metab. 2003, 88, 1537–1542. [Google Scholar] [CrossRef] [PubMed]
- Arvat, E.; Maccario, M.; Vito, L.D.; Broglio, F.; Benso, A.; Gottero, C.; Papotti, M.; Muccioli, G.; Dieguez, C.; Casanueva, F.F.; et al. Endocrine activities of ghrelin, a natural growth hormone secretagogue (GHS), in humans: Comparison and interactions with hexarelin, a nonnatural peptidyl GHS, and GH-releasing hormone. J. Clin. Endocrinol. Metab. 2001, 86, 1169–1174. [Google Scholar] [CrossRef] [PubMed]
- Deghenghi, R.; Cananzi, M.M.; Torsello, A.; Battisti, C.; Muller, E.E.; Locatelli, V. GH-releasing activity of hexarelin, a new growth hormone releasing peptide, in infant and adult rats. Life Sci. 1994, 54, 1321–1328. [Google Scholar] [CrossRef]
- Ghigo, E.; Arvat, E.; Gianotti, L.; Grottoli, S.; Rizzi, G.; Ceda, G.P.; Boghen, M.F.; Deghenghi, R.; Camanni, F. Short-term administration of intranasal or oral hexarelin, a synthetic hexapeptide, does not desensitize the growth hormone responsiveness in human aging. Eur. J. Endocrinol. 1996, 135, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Hansen, B.S.; Raun, K.; Nielsen, K.K.; Johansen, P.B.; Hansen, T.K.; Peschke, B.; Lau, J.; Andersen, P.H.; Ankersen, M. Pharmacological characterisation of a new oral gh secretagogue, NN703. Eur. J. Endocrinol. 1999, 141, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Patchett, A.A.; Nargund, R.P.; Tata, J.R.; Chen, M.H.; Barakat, K.J.; Johnston, D.B.; Cheng, K.; Chan, W.W.; Butler, B.; Hickey, G. Design and biological activities of L-163,191 (MK-0677): A potent, orally active growth hormone secretagogue. Proc. Natl. Acad. Sci. USA 1995, 92, 7001–7005. [Google Scholar] [CrossRef] [PubMed]
- Phung, L.T.; Sasaki, A.; Lee, H.G.; Vega, R.A.; Matsunaga, N.; Hidaka, S.; Kuwayama, H.; Hidari, H. Effects of the administration of growth hormone-releasing peptide-2 (GHRP-2) orally by gavage and in feed on growth hormone release in swine. Domest. Anim. Endocrinol. 2001, 20, 9–19. [Google Scholar] [CrossRef]
- Pihoker, C.; Badger, T.M.; Reynolds, G.A.; Bowers, C.Y. Treatment effects of intranasal growth hormone releasing peptide-2 in children with short stature. J. Endocrinol. 1997, 155, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Roumi, M.; Marleau, S.; du Souich, P.; Maggi, T.; Deghenghi, R.; Ong, H. Kinetics and disposition of hexarelin, a peptidic growth hormone secretagogue, in rats. Drug Metab. Dispos. 2000, 28, 44–50. [Google Scholar] [PubMed]
- Torsello, A.; Luoni, M.; Schweiger, F.; Grilli, R.; Guidi, M.; Bresciani, E.; Deghenghi, R.; Müller, E.E.; Locatelli, V. Novel hexarelin analogs stimulate feeding in the rat through a mechanism not involving growth hormone release. Eur. J. Pharmacol. 1998, 360, 123–129. [Google Scholar] [CrossRef]
- Laferrere, B.; Abraham, C.; Russell, C.D.; Bowers, C.Y. Growth hormone releasing peptide-2 (GHRP-2), like ghrelin, increases food intake in healthy men. J. Clin. Endocrinol. Metab. 2005, 90, 611–614. [Google Scholar] [CrossRef] [PubMed]
- Okada, K.; Ishii, S.; Minami, S.; Sugihara, H.; Shibasaki, T.; Wakabayashi, I. Intracerebroventricular administration of the growth hormone-releasing peptide KP-102 increases food intake in free-feeding rats. Endocrinology 1996, 137, 5155–5158. [Google Scholar] [PubMed]
- Lee, H.G.; Vega, R.A.; Phung, L.T.; Matsunaga, N.; Kuwayama, H.; Hidari, H. The effect of growth hormone-releasing peptide-2 (KP102) administration on plasma insulin-like growth factor (IGF)-1 and IGF-binding proteins in holstein steers on different planes of nutrition. Domest. Anim. Endocrinol. 2000, 18, 293–308. [Google Scholar] [CrossRef]
- Lawrence, C.B.; Snape, A.C.; Baudoin, F.M.; Luckman, S.M. Acute central ghrelin and GH secretagogues induce feeding and activate brain appetite centers. Endocrinology 2002, 143, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Inui, A.; Asakawa, A.; Bowers, C.Y.; Mantovani, G.; Laviano, A.; Meguid, M.M.; Fujimiya, M. Ghrelin, appetite, and gastric motility: The emerging role of the stomach as an endocrine organ. FASEB J. 2004, 18, 439–456. [Google Scholar] [CrossRef] [PubMed]
- Egecioglu, E.; Jerlhag, E.; Salome, N.; Skibicka, K.P.; Haage, D.; Bohlooly-Y, M.; Andersson, D.; Bjursell, M.; Perrissoud, D.; Engel, J.A.; et al. Ghrelin increases intake of rewarding food in rodents. Addict. Biol. 2010, 15, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Skibicka, K.P.; Hansson, C.; Alvarez-Crespo, M.; Friberg, P.A.; Dickson, S.L. Ghrelin directly targets the ventral tegmental area to increase food motivation. Neuroscience 2011, 180, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, Y.; Kawahara, H.; Kaneko, F.; Yamada, M.; Nishi, Y.; Tanaka, E.; Nishi, A. Peripherally administered ghrelin induces bimodal effects on the mesolimbic dopamine system depending on food-consumptive states. Neuroscience 2009, 161, 855–864. [Google Scholar] [CrossRef] [PubMed]
- Sanger, G.J. Motilin, ghrelin and related neuropeptides as targets for the treatment of GI diseases. Drug Discov. Today 2008, 13, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Charoenthongtrakul, S.; Giuliana, D.; Longo, K.A.; Govek, E.K.; Nolan, A.; Gagne, S.; Morgan, K.; Hixon, J.; Flynn, N.; Murphy, B.J.; et al. Enhanced gastrointestinal motility with orally active ghrelin receptor agonists. J. Pharmacol. Exp. Ther. 2009, 329, 1178–1186. [Google Scholar] [CrossRef] [PubMed]
- Sivertsen, B.; Lang, M.; Frimurer, T.M.; Holliday, N.D.; Bach, A.; Els, S.; Engelstoft, M.S.; Petersen, P.S.; Madsen, A.N.; Schwartz, T.W.; et al. Unique interaction pattern for a functionally biased ghrelin receptor agonist. J. Biol. Chem. 2011, 286, 20845–20860. [Google Scholar] [CrossRef] [PubMed]
- Hoveyda, H.R.; Marsault, E.; Gagnon, R.; Mathieu, A.P.; Vezina, M.; Landry, A.; Wang, Z.; Benakli, K.; Beaubien, S.; Saint-Louis, C.; et al. Optimization of the potency and pharmacokinetic properties of a macrocyclic ghrelin receptor agonist (Part I): Development of ulimorelin (TZP-101) from hit to clinic. J. Med. Chem. 2011, 54, 8305–8320. [Google Scholar] [CrossRef] [PubMed]
- Pietra, C.; Takeda, Y.; Tazawa-Ogata, N.; Minami, M.; Xia, Y.; Duus, E.M.; Northrup, R. Anamorelin HCL (ONO-7643), a novel ghrelin receptor agonist, for the treatment of cancer anorexia-cachexia syndrome: Preclinical profile. J. Cachexia Sarcopenia Muscle 2014, 5, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Halem, H.A.; Taylor, J.E.; Dong, J.Z.; Shen, Y.; Datta, R.; Abizaid, A.; Diano, S.; Horvath, T.L.; Culler, M.D. A novel growth hormone secretagogue-1a receptor antagonist that blocks ghrelin-induced growth hormone secretion but induces increased body weight gain. Neuroendocrinology 2005, 81, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Hassouna, R.; Labarthe, A.; Zizzari, P.; Videau, C.; Culler, M.; Epelbaum, J.; Tolle, V. Actions of agonists and antagonists of the ghrelin/GHS-R pathway on GH secretion, appetite, and cFos activity. Front. Endocrinol. 2013, 4, 25. [Google Scholar] [CrossRef] [PubMed]
- Costantini, V.J.; Vicentini, E.; Sabbatini, F.M.; Valerio, E.; Lepore, S.; Tessari, M.; Sartori, M.; Michielin, F.; Melotto, S.; Bifone, A.; et al. GSK1614343, a novel ghrelin receptor antagonist, produces an unexpected increase of food intake and body weight in rodents and dogs. Neuroendocrinology 2011, 94, 158–168. [Google Scholar] [CrossRef] [PubMed]
- M’Kadmi, C.; Leyris, J.-P.; Onfroy, L.; Galès, C.; Saulière, A.; Gagne, D.; Damian, M.; Mary, S.; Maingot, M.; Denoyelle, S.; et al. Agonism, antagonism and inverse agonism bias at the Ghrelin receptor signaling. J. Biol. Chem. 2015, 290, 27021–27039. [Google Scholar] [CrossRef] [PubMed]
- Depoortere, I. Targeting the ghrelin receptor to regulate food intake. Regul. Pept. 2009, 156, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Moulin, A.; Ryan, J.; Martinez, J.; Fehrentz, J.-A. Recent developments in ghrelin receptor ligands. ChemMedChem 2007, 2, 1242–1259. [Google Scholar] [CrossRef] [PubMed]
- Walker, R.F.; Codd, E.E.; Barone, F.C.; Nelson, A.H.; Goodwin, T.; Campbell, S.A. Oral activity of the growth hormone releasing peptide His-D-Trp-Ala-Trp-D-Phe-Lys-NH2 in rats, dogs and monkeys. Life Sci. 1990, 47, 29–36. [Google Scholar] [CrossRef]
- Bowers, C.Y.; Momany, F.A.; Reynolds, G.A.; Hong, A. On the in vitro and in vivo activity of a new synthetic hexapeptide that acts on the pituitary to specifically release growth hormone. Endocrinology 1984, 114, 1537–1545. [Google Scholar] [CrossRef] [PubMed]
- Ghigo, E.; Arvat, E.; Gianotti, L.; Imbimbo, B.P.; Lenaerts, V.; Deghenghi, R.; Camanni, F. Growth hormone-releasing activity of hexarelin, a new synthetic hexapeptide, after intravenous, subcutaneous, intranasal, and oral administration in man. J. Clin. Endocrinol. Metab. 1994, 78, 693–698. [Google Scholar] [PubMed]
- Laron, Z.; Frenkel, J.; Deghenghi, R.; Anin, S.; Klinger, B.; Silbergeld, A. Intranasal administration of the GHRP hexarelin accelerates growth in short children. Clin. Endocrinol. 1995, 43, 631–635. [Google Scholar] [CrossRef]
- Imbimbo, B.P.; Mant, T.; Edwards, M.; Amin, D.; Dalton, N.; Boutignon, F.; Lenaerts, V.; Wuthrich, P.; Deghenghi, R. Growth hormone-releasing activity of hexarelin in humans. A dose-response study. Eur. J. Clin. Pharmacol. 1994, 46, 421–425. [Google Scholar] [CrossRef] [PubMed]
- Arvat, E.; di Vito, L.; Maccagno, B.; Broglio, F.; Boghen, M.F.; Deghenghi, R.; Camanni, F.; Ghigo, E. Effects of GHRP-2 and hexarelin, two synthetic GH-releasing peptides, on GH, prolactin, ACTH and cortisol levels in man. Comparison with the effects of GHRH, TRH and HCRH. Peptides 1997, 18, 885–891. [Google Scholar] [CrossRef]
- Bowers, C.Y. Gh releasing peptides—Structure and kinetics. J. Pediatr. Endocrinol. 1993, 6, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Pihoker, C.; Kearns, G.L.; French, D.; Bowers, C.Y. Pharmacokinetics and pharmacodynamics of growth hormone-releasing peptide-2: A phase I study in children. J. Clin. Endocrinol. Metab. 1998, 83, 1168–1172. [Google Scholar] [CrossRef] [PubMed]
- Broglio, F.; Benso, A.; Gottero, C.; Muccioli, G.; Deghenghi, R.; Ghigo, E.; Arvat, E. Endocrine activities of alexamorelin (Ala-His-D-2-Methyl-Trp-Ala-Trp-D-Phe-Lys-Nh2), a synthetic gh secretagogue, in humans. Eur. J. Endocrinol. 2000, 143, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Ankersen, M.; Johansen, N.L.; Madsen, K.; Hansen, B.S.; Raun, K.; Nielsen, K.K.; Thogersen, H.; Hansen, T.K.; Peschke, B.; Lau, J.; et al. A new series of highly potent growth hormone-releasing peptides derived from ipamorelin. J. Med. Chem. 1998, 41, 3699–3704. [Google Scholar] [CrossRef] [PubMed]
- Gobburu, J.V.; Agerso, H.; Jusko, W.J.; Ynddal, L. Pharmacokinetic-pharmacodynamic modeling of ipamorelin, a growth hormone releasing peptide, in human volunteers. Pharm. Res. 1999, 16, 1412–1416. [Google Scholar] [CrossRef] [PubMed]
- Johansen, P.B.; Nowak, J.; Skjaerbaek, C.; Flyvbjerg, A.; Andreassen, T.T.; Wilken, M.; Orskov, H. Ipamorelin, a new growth-hormone-releasing peptide, induces longitudinal bone growth in rats. Growth Horm. IGF Res. 1999, 9, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Polvino, W.J.; Nelson, R.; Mann, W.R. Method of Stimulating the Motility of the Gastrointestinal System using Ipamorelin. U.S. Patent U.S. 20090143310 A1, 4 July 2009. [Google Scholar]
- Carpino, P.A.; Lefker, B.A.; Toler, S.M.; Pan, L.C.; Hadcock, J.R.; Cook, E.R.; DiBrino, J.N.; Campeta, A.M.; DeNinno, S.L.; Chidsey-Frink, K.L.; et al. Pyrazolinone-piperidine dipeptide growth hormone secretagogues (GHSS). Discovery of capromorelin. Bioorg. Med. Chem. 2003, 11, 581–590. [Google Scholar] [CrossRef]
- Khojasteh-Bakht, S.C.; O’Donnell, J.P.; Fouda, H.G.; Potchoiba, M.J. Metabolism, pharmacokinetics, tissue distribution, and excretion of [14c]Cp-424391 in rats. Drug Metab. Dispos. Biol. Fate Chem. 2005, 33, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.G.; Pong, S.S.; Hickey, G.; Jacks, T.; Cheng, K.; Leonard, R.; Cohen, C.J.; Arena, J.P.; Chang, C.H.; Drisko, J.; et al. Modulation of pulsatile GH release through a novel receptor in hypothalamus and pituitary gland. Recent Prog. Horm. Res. 1996, 51, 261–285. [Google Scholar] [PubMed]
- Pan, L.C.; Carpino, P.A.; Lefker, B.A.; Ragan, J.A.; Toler, S.M.; Pettersen, J.C.; Nettleton, D.O.; Ng, O.; Pirie, C.M.; Chidsey-Frink, K.; et al. Preclinical pharmacology of CP-424,391, an orally active pyrazolinone-piperidine [correction of pyrazolidinone-piperidine] growth hormone secretagogue. Endocrine 2001, 14, 121–132. [Google Scholar] [CrossRef]
- Lembo, A.; Camilleri, M.; McCallum, R.; Sastre, R.; Breton, C.; Spence, S.; White, J.; Currie, M.; Gottesdiener, K.; Stoner, E. Relamorelin reduces vomiting frequency and severity and accelerates gastric emptying in adults with diabetic gastroparesis. Gastroenterology 2016, 151, 87.e86–96.e86. [Google Scholar] [CrossRef] [PubMed]
- Palus, S.; Schur, R.; Akashi, Y.J.; Bockmeyer, B.; Datta, R.; Halem, H.; Dong, J.; Culler, M.D.; Adams, V.; Anker, S.D.; et al. Ghrelin and its analogues, BIM-28131 and BIM-28125, improve body weight and regulate the expression of MuRF-1 and MAFbx in a rat heart failure model. PLoS ONE 2011, 6, e26865. [Google Scholar] [CrossRef] [PubMed]
- Strassburg, S.; Anker, S.D.; Castaneda, T.R.; Burget, L.; Perez-Tilve, D.; Pfluger, P.T.; Nogueiras, R.; Halem, H.; Dong, J.Z.; Culler, M.D.; et al. Long-term effects of ghrelin and ghrelin receptor agonists on energy balance in rats. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E78–E84. [Google Scholar] [CrossRef] [PubMed]
- Fischer, K.; Finan, B.; Clemmensen, C.; van der Ploeg, L.H.T.; Tschöp, M.H.; Müller, T.D. The pentapeptide rm-131 promotes food intake and adiposity in wildtype mice but not in mice lacking the ghrelin receptor. Front. Nutr. 2014, 1, 31. [Google Scholar] [CrossRef] [PubMed]
- Palus, S.; von Haehling, S.; Doehner, W.; Datta, R.; Zhang, J.; Dong, J.Z.; Culler, M.D.; Anker, S.D.; Springer, J. Effect of application route of the ghrelin analog BIM-28131 (RM-131) on body weight and body composition in a rat heart failure model. Int. J. Cardiol. 2013, 168, 2369–2374. [Google Scholar] [CrossRef] [PubMed]
- Van der Ploeg, L.; Laken, H.; Sharma, S.; Datta, R.; Halem, H.; Dong, J.; Touvay, C.; Teillot, M.; Noonan, P.; Tartaglia, L.; et al. Preclinical gastrointestinal prokinetic efficacy and endocrine effects of the ghrelin mimetic RM-131. Life Sci. 2014, 109, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Shin, A.; Camilleri, M.; Busciglio, I.; Burton, D.; Smith, S.A.; Vella, A.; Ryks, M.; Rhoten, D.; Zinsmeister, A.R. The ghrelin agonist RM-131 accelerates gastric emptying of solids and reduces symptoms in patients with type 1 diabetes mellitus. Clin. Gastroenterol. Hepatol. 2013, 11, 1453–1459. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.M.; Swerdloff, R.; Wang, C.; Kyle, M.; Kipnes, M.; Biller, B.M.; Cook, D.; Yuen, K.C.; Bonert, V.; Dobs, A.; et al. Macimorelin (AEZS-130)-stimulated growth hormone (GH) test: Validation of a novel oral stimulation test for the diagnosis of adult GH deficiency. J. Clin. Endocrinol. Metab. 2013, 98, 2422–2429. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.A.; Garcia, J.M. Randomized clinical trial of the novel oral ghrelin mimetic macimorelin in the treatment of cancer cachexia: Study design and preliminary results. In Proceedings of the Endocrine Society’s 95th Annual Meeting and Expo, San Francisco, CA, USA, 15–18 June 2013.
- Piccoli, F.; Degen, L.; MacLean, C.; Peter, S.; Baselgia, L.; Larsen, F.; Beglinger, C.; Drewe, J. Pharmacokinetics and pharmacodynamic effects of an oral ghrelin agonist in healthy subjects. J. Clin. Endocrinol. Metab. 2007, 92, 1814–1820. [Google Scholar] [CrossRef] [PubMed]
- Broglio, F.; Boutignon, F.; Benso, A.; Gottero, C.; Prodam, F.; Arvat, E.; Ghè, C.; Catapano, F.; Torsello, A.; Locatelli, V.; et al. EP1572: A novel peptido-mimetic GH secretagogue with potent and selective GH-releasing activity in man. J. Endocrinol. Investig. 2002, 25, RC26–RC28. [Google Scholar] [CrossRef] [PubMed]
- Ankersen, M.; Kramer Nielsen, K.; Kruse Hansen, T.; Raun, K.; Sehested Hansen, B. Growth hormone secretagogues derived from NN703 with hydrazidesas c-terminal. Eur. J. Med. Chem. 2000, 35, 487–497. [Google Scholar] [CrossRef]
- Zdravkovic, M.; Søgaard, B.; Ynddal, L.; Christiansen, T.; Agersø, H.; Thomsen, M.S.; Falch, J.E.; Ilondo, M.M. The pharmacokinetics, pharmacodynamics, safety and tolerability of a single dose of NN703, a novel orally active growth hormone secretagogue in healthy male volunteers. Growth Horm. IGF Res. 2000, 10, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Zdravkovic, M.; Christiansen, T.; Eliot, L.; Agersoe, H.; Thomsen, M.S.; Falch, J.F.; Søgaard, B.; Ynddal, L.; Ilondo, M.M. The pharmacokinetics, pharmacodynamics, safety and tolerability following 7 days daily oral treatment with NN703 in healthy male subjects. Growth Horm. IGF Res. 2001, 11, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.M.; Polvino, W.J. Pharmacodynamic hormonal effects of anamorelin, a novel oral ghrelin mimetic and growth hormone secretagogue in healthy volunteers. Growth Horm. IGF Res. 2009, 19, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.M.; Polvino, W.J. Effect on body weight and safety of rc-1291, a novel, orally available ghrelin mimetic and growth hormone secretagogue: Results of a phase I, randomized, placebo-controlled, multiple-dose study in healthy volunteers. Oncologist 2007, 12, 594–600. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.; Boccia, R.V.; Graham, C.; Kumor, K.; Polvino, W. A phase II randomized, placebo-controlled, double-blind study of the efficacy and safety of RC-1291 (RC) for the treatment of cancer cachexia. J. Clin. Oncol. 2007, 25, 9133. [Google Scholar]
- Garcia, J.M.; Friend, J.; Allen, S. Therapeutic potential of anamorelin, a novel, oral ghrelin mimetic, in patients with cancer-related cachexia: A multicenter, randomized, double-blind, crossover, pilot study. Support Care Cancer 2013, 21, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Adunsky, A.; Chandler, J.; Heyden, N.; Lutkiewicz, J.; Scott, B.B.; Berd, Y.; Liu, N.; Papanicolaou, D.A. Mk-0677 (ibutamoren mesylate) for the treatment of patients recovering from hip fracture: A multicenter, randomized, placebo-controlled phase IIb study. Arch. Gerontol. Geriatr. 2011, 53, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Svensson, J.; Lonn, L.; Jansson, J.O.; Murphy, G.; Wyss, D.; Krupa, D.; Cerchio, K.; Polvino, W.; Gertz, B.; Boseaus, I.; et al. Two-month treatment of obese subjects with the oral growth hormone (GH) secretagogue MK-677 increases GH secretion, fat-free mass, and energy expenditure. J. Clin. Endocrinol. Metab. 1998, 83, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Jacks, T.; Smith, R.; Judith, F.; Schleim, K.; Frazier, E.; Chen, H.; Krupa, D.; Hora, D., Jr.; Nargund, R.; Patchett, A.; et al. Mk-0677, a potent, novel, orally active growth hormone (GH) secretagogue: GH, insulin-like growth factor i, and other hormonal responses in beagles. Endocrinology 1996, 137, 5284–5289. [Google Scholar] [PubMed]
- Prahalada, S.; Block, G.; Handt, L.; DeBurlet, G.; Cahill, M.; Hoe, C.M.; van Zwieten, M.J. Insulin-like growth factor-1 and growth hormone (GH) levels in canine cerebrospinal fluid are unaffected by GH or gh secretagogue (MK-0677) administration. Horm. Metab. Res. 1999, 31, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Fraser, G.L.; Hoveyda, H.R.; Tannenbaum, G.S. Pharmacological demarcation of the growth hormone, gut motility and feeding effects of ghrelin using a novel ghrelin receptor agonist. Endocrinology 2008, 149, 6280–6288. [Google Scholar] [CrossRef] [PubMed]
- Venkova, K.; Fraser, G.; Hoveyda, H.R.; Greenwood-Van Meerveld, B. Prokinetic effects of a new ghrelin receptor agonist TZP-101 in a rat model of postoperative ileus. Dig. Dis. Sci. 2007, 52, 2241–2248. [Google Scholar] [CrossRef] [PubMed]
- Fraser, G.L.; Venkova, K.; Hoveyda, H.R.; Thomas, H.; Greenwood-Van Meerveld, B. Effect of the ghrelin receptor agonist TZP-101 on colonic transit in a rat model of postoperative ileus. Eur. J. Pharmacol. 2009, 604, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Lasseter, K.C.; Shaughnessy, L.; Cummings, D.; Pezzullo, J.C.; Wargin, W.; Gagnon, R.; Oliva, J.; Kosutic, G. Ghrelin agonist (TZP-101): Safety, pharmacokinetics and pharmacodynamic evaluation in healthy volunteers: A phase I, first-in-human study. J. Clin. Pharmacol. 2008, 48, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Wo, J.M.; Ejskjaer, N.; Hellström, P.M.; Malik, R.A.; Pezzullo, J.C.; Shaughnessy, L.; Charlton, P.; Kosutic, G.; McCallum, R.W. Randomised clinical trial: Ghrelin agonist TZP-101 relieves gastroparesis associated with severe nausea and vomiting—Randomised clinical study subset data. Aliment. Pharmacol. Ther. 2011, 33, 679–688. [Google Scholar] [CrossRef] [PubMed]
- Ejskjaer, N.; Dimcevski, G.; Wo, J.; Hellstrom, P.M.; Gormsen, L.C.; Sarosiek, I.; Softeland, E.; Nowak, T.; Pezzullo, J.C.; Shaughnessy, L.; et al. Safety and efficacy of ghrelin agonist TZP-101 in relieving symptoms in patients with diabetic gastroparesis: A randomized, placebo-controlled study. Neurogastroenterol. Motil. 2010, 22, e1069–e1281. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Garcia, J.M. Anamorelin hydrochloride for the treatment of cancer-anorexia-cachexia in NSCLC. Expert Opin. Pharmacother. 2015, 16, 1245–1253. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.M.; Boccia, R.V.; Graham, C.D.; Yan, Y.; Duus, E.M.; Allen, S.; Friend, J. Anamorelin for patients with cancer cachexia: An integrated analysis of two phase 2, randomised, placebo-controlled, double-blind trials. Lancet Oncol. 2015, 16, 108–116. [Google Scholar] [CrossRef]
- Shimizu, Y.; Chang, E.C.; Shafton, A.D.; Ferens, D.M.; Sanger, G.J.; Witherington, J.; Furness, J.B. Evidence that stimulation of ghrelin receptors in the spinal cord initiates propulsive activity in the colon of the rat. J. Physiol. 2006, 576, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Shafton, A.D.; Sanger, G.J.; Witherington, J.; Brown, J.D.; Muir, A.; Butler, S.; Abberley, L.; Shimizu, Y.; Furness, J.B. Oral administration of a centrally acting ghrelin receptor agonist to conscious rats triggers defecation. Neurogastroenterol. Motil. 2009, 21, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Naitou, K.; Mamerto, T.; Pustovit, R.V.; Rivera, L.R.; Callaghan, B.; Ringuet, M.T.; Chan, A.; Pietra, C.; Furness, J.B. The ghrelin receptor agonist, HM01, activates the innervation of the colon to initiate coordinated propulsive contractions and bowel emptying. Auton. Neurosci. Basic Clin. 2015, 192, 58–59. [Google Scholar] [CrossRef]
- Karasawa, H.; Pietra, C.; Giuliano, C.; Garcia-Rubio, S.; Xu, X.; Yakabi, S.; Taché, Y.; Wang, L. New ghrelin agonist, HM01 alleviates constipation and l-dopa-delayed gastric emptying in 6-hydroxydopamine rat model of Parkinson’s disease. Neurogastroenterol. Motil. 2014, 26, 1771–1782. [Google Scholar] [CrossRef] [PubMed]
- Naitou, K.; Mamerto, T.P.; Pustovit, R.V.; Callaghan, B.; Rivera, L.R.; Chan, A.J.; Ringuet, M.T.; Pietra, C.; Furness, J.B. Site and mechanism of the colokinetic action of the ghrelin receptor agonist, HM01. Neurogastroenterol. Motil. 2015, 27, 1764–1771. [Google Scholar] [CrossRef] [PubMed]
- Borner, T.; Loi, L.; Pietra, C.; Giuliano, C.; Lutz, T.A.; Riediger, T. The ghrelin receptor agonist HM01 mimics the neuronal effects of ghrelin in the arcuate nucleus and attenuates anorexia-cachexia syndrome in tumor-bearing rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 311, R89–R96. [Google Scholar] [CrossRef] [PubMed]
- Esler, W.P.; Rudolph, J.; Claus, T.H.; Tang, W.; Barucci, N.; Brown, S.-E.; Bullock, W.; Daly, M.; DeCarr, L.; Li, Y.; et al. Small-molecule ghrelin receptor antagonists improve glucose tolerance, suppress appetite, and promote weight loss. Endocrinology 2007, 148, 5175–5185. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, J.; Esler, W.P.; O’Connor, S.; Coish, P.D.; Wickens, P.L.; Brands, M.; Bierer, D.E.; Bloomquist, B.T.; Bondar, G.; Chen, L.; et al. Quinazolinone derivatives as orally available ghrelin receptor antagonists for the treatment of diabetes and obesity. J. Med. Chem. 2007, 50, 5202–5216. [Google Scholar] [CrossRef] [PubMed]
- Moran, T.H.; Dailey, M.J. Gut peptides: Targets for antiobesity drug development? Endocrinology 2009, 150, 2526–2530. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, B.; Funami, H.; Iwaki, T.; Maruoka, H.; Shibata, M.; Koyama, M.; Nagahira, A.; Kamiide, Y.; Kanki, S.; Igawa, Y.; et al. Orally active ghrelin receptor inverse agonists and their actions on a rat obesity model. Bioorg. Med. Chem. 2015, 23, 4792–4803. [Google Scholar] [CrossRef] [PubMed]
- McCoull, W.; Barton, P.; Brown, A.J.H.; Bowker, S.S.; Cameron, J.; Clarke, D.S.; Davies, R.D.M.; Dossetter, A.G.; Ertan, A.; Fenwick, M.; et al. Identification, optimization, and pharmacology of acylurea GHS-R1a inverse agonists. J. Med. Chem. 2014, 57, 6128–6140. [Google Scholar] [CrossRef] [PubMed]
- Engel, J.A.; Nylander, I.; Jerlhag, E. A ghrelin receptor (GHS-R1a) antagonist attenuates the rewarding properties of morphine and increases opioid peptide levels in reward areas in mice. Eur. Neuropsychopharmacol. 2015, 25, 2364–2371. [Google Scholar] [CrossRef] [PubMed]
- Jerlhag, E.; Engel, J.A. Ghrelin receptor antagonism attenuates nicotine-induced locomotor stimulation, accumbal dopamine release and conditioned place preference in mice. Drug Alcohol. Depend. 2011, 117, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Skibicka, K.P.; Hansson, C.; Egecioglu, E.; Dickson, S.L. Role of ghrelin in food reward: Impact of ghrelin on sucrose self-administration and mesolimbic dopamine and acetylcholine receptor gene expression. Addict. Biol. 2012, 17, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Sustkova-Fiserova, M.; Jerabek, P.; Havlickova, T.; Kacer, P.; Krsiak, M. Ghrelin receptor antagonism of morphine-induced accumbens dopamine release and behavioral stimulation in rats. Psychopharmacology 2014, 231, 2899–2908. [Google Scholar] [CrossRef] [PubMed]
- Jerlhag, E.; Egecioglu, E.; Landgren, S.; Salome, N.; Heilig, M.; Moechars, D.; Datta, R.; Perrissoud, D.; Dickson, S.L.; Engel, J.A. Requirement of central ghrelin signaling for alcohol reward. Proc. Natl. Acad. Sci. USA 2009, 106, 11318–11323. [Google Scholar] [CrossRef] [PubMed]
- Lenard, N.R.; Berthoud, H.-R. Central and peripheral regulation of food intake and physical activity: Pathways and genes. Obesity 2008, 16, S11–S22. [Google Scholar] [CrossRef] [PubMed]
- Wise, R.A. Role of brain dopamine in food reward and reinforcement. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 1149–1158. [Google Scholar] [CrossRef] [PubMed]
- Fields, H.L.; Hjelmstad, G.O.; Margolis, E.B.; Nicola, S.M. Ventral tegmental area neurons in learned appetitive behavior and positive reinforcement. Annu. Rev. Neurosci. 2007, 30, 289–316. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, N.S.; Guarnieri, D.J.; DiLeone, R.J. Metabolic hormones, dopamine circuits, and feeding. Front. Neuroendocrinol. 2010, 31, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Naleid, A.M.; Grace, M.K.; Cummings, D.E.; Levine, A.S. Ghrelin induces feeding in the mesolimbic reward pathway between the ventral tegmental area and the nucleus accumbens. Peptides 2005, 26, 2274–2279. [Google Scholar] [CrossRef] [PubMed]
- Skibicka, K.P.; Shirazi, R.H.; Rabasa-Papio, C.; Alvarez-Crespo, M.; Neuber, C.; Vogel, H.; Dickson, S.L. Divergent circuitry underlying food reward and intake effects of ghrelin: Dopaminergic VTA-accumbens projection mediates ghrelin’s effect on food reward but not food intake. Neuropharmacology 2013, 73, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Parent, M.A.; Amarante, L.M.; Swanson, K.; Laubach, M. Cholinergic and ghrelinergic receptors and kcnq channels in the medial PFC regulate the expression of palatability. Front. Behav. Neurosci. 2015, 9, 284. [Google Scholar] [CrossRef] [PubMed]
- Jerlhag, E.; Egecioglu, E.; Dickson, S.L.; Douhan, A.; Svensson, L.; Engel, J.A. Ghrelin administration into tegmental areas stimulates locomotor activity and increases extracellular concentration of dopamine in the nucleus accumbens. Addict. Biol. 2007, 12, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Van der Plasse, G.; van Zessen, R.; Luijendijk, M.C.; Erkan, H.; Stuber, G.D.; Ramakers, G.M.; Adan, R.A. Modulation of cue-induced firing of ventral tegmental area dopamine neurons by leptin and ghrelin. Int. J. Obes. 2015, 39, 1742–1749. [Google Scholar] [CrossRef] [PubMed]
- Jerlhag, E. Systemic administration of ghrelin induces conditioned place preference and stimulates accumbal dopamine. Addict. Biol. 2008, 13, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Schaeffer, M.; Langlet, F.; Lafont, C.; Molino, F.; Hodson, D.J.; Roux, T.; Lamarque, L.; Verdie, P.; Bourrier, E.; Dehouck, B.; et al. Rapid sensing of circulating ghrelin by hypothalamic appetite-modifying neurons. Proc. Natl. Acad. Sci. USA 2013, 110, 1512–1517. [Google Scholar] [CrossRef] [PubMed]
- Banks, W.A.; Burney, B.O.; Robinson, S.M. Effects of triglycerides, obesity, and starvation on ghrelin transport across the blood-brain barrier. Peptides 2008, 29, 2061–2065. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.; Tu, H.; Kastin, A.J. Differential BBB interactions of three ingestive peptides: Obestatin, ghrelin, and adiponectin. Peptides 2006, 27, 911–916. [Google Scholar] [CrossRef] [PubMed]
- Drewes, L.R. Biology of the blood-brain glucose transporter. In Introduction to the Blood-Brain Barrier: Methodology; Biology and Pathology; Cambridge University Press: Cambridge, UK, 1998; pp. 165–174. [Google Scholar]
- Schwartz, M.W.; Sipols, A.; Kahn, S.E.; Lattemann, D.F.; Taborsky, G.J.; Bergman, R.N.; Woods, S.C.; Porte, D. Kinetics and specificity of insulin uptake from plasma into cerebrospinal fluid. Am. J. Physiol. Endocrinol. Metab. 1990, 259, E378. [Google Scholar]
- Currie, P.J.; Khelemsky, R.; Rigsbee, E.M.; Dono, L.M.; Coiro, C.D.; Chapman, C.D.; Hinchcliff, K. Ghrelin is an orexigenic peptide and elicits anxiety-like behaviors following administration into discrete regions of the hypothalamus. Behav. Brain Res. 2012, 226, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Olszewski, P.K.; Grace, M.K.; Billington, C.J.; Levine, A.S. Hypothalamic paraventricular injections of ghrelin: Effect on feeding and c-Fos immunoreactivity. Peptides 2003, 24, 919–923. [Google Scholar] [CrossRef]
- Nieh, E.H.; Matthews, G.A.; Allsop, S.A.; Presbrey, K.N.; Leppla, C.A.; Wichmann, R.; Neve, R.; Wildes, C.P.; Tye, K.M. Decoding neural circuits that control compulsive sucrose seeking. Cell 2015, 160, 528–541. [Google Scholar] [CrossRef] [PubMed]
- Stuber, G.D.; Wise, R.A. Lateral hypothalamic circuits for feeding and reward. Nat. Neurosci. 2016, 19, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Lutter, M.; Nestler, E.J. Homeostatic and hedonic signals interact in the regulation of food intake. J. Nutr. 2009, 139, 629–632. [Google Scholar] [CrossRef] [PubMed]
- Harris, G.C.; Wimmer, M.; Aston-Jones, G. A role for lateral hypothalamic orexin neurons in reward seeking. Nature 2005, 437, 556–559. [Google Scholar] [CrossRef] [PubMed]
- Cason, A.M.; Smith, R.J.; Tahsili-Fahadan, P.; Moorman, D.E.; Sartor, G.C.; Aston-Jones, G. Role of orexin/hypocretin in reward-seeking and addiction: Implications for obesity. Physiol. Behav. 2010, 100, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Aston-Jones, G.; Smith, R.J.; Sartor, G.C.; Moorman, D.E.; Massi, L.; Tahsili-Fahadan, P.; Richardson, K.A. Lateral hypothalamic orexin/hypocretin neurons: A role in reward-seeking and addiction. Brain Res. 2010, 1314, 74–90. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Z.; Santiago, A.M.; Thomas, M.P.; Routh, V.H. Metabolic regulation of lateral hypothalamic glucose-inhibited orexin neurons may influence midbrain reward neurocircuitry. Mol. Cell. Neurosci. 2014, 62, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Saper, C.B.; Loewy, A.D. Efferent connections of the parabrachial nucleus in the rat. Brain Res. 1980, 197, 291–317. [Google Scholar] [CrossRef]
- Krukoff, T.L.; Harris, K.H.; Jhamandas, J.H. Efferent projections from the parabrachial nucleus demonstrated with the anterograde tracer phaseolus vulgaris leucoagglutinin. Brain Res. Bull. 1993, 30, 163–172. [Google Scholar] [CrossRef]
- Grill, H.J.; Friedman, M.I.; Norgren, R.; Scalera, G.; Seeley, R. Parabrachial nucleus lesions impair feeding response elicited by 2,5-anhydro-d-mannitol. Am. J. Physiol. 1995, 268, R676–R682. [Google Scholar] [PubMed]
- Cornwall, J.; Cooper, J.D.; Phillipson, O.T. Afferent and efferent connections of the laterodorsal tegmental nucleus in the rat. Brain Res. Bull. 1990, 25, 271–284. [Google Scholar] [CrossRef]
- Wu, Q.; Boyle, M.P.; Palmiter, R.D. Loss of Gabaergic signaling by AgRP neurons to the parabrachial nucleus leads to starvation. Cell 2009, 137, 1225–1234. [Google Scholar] [CrossRef] [PubMed]
- Scalera, G.; Spector, A.C.; Norgren, R. Excitotoxic lesions of the parabrachial nuclei prevent conditioned taste aversions and sodium appetite in rats. Behav. Neurosci. 1995, 109, 997. [Google Scholar] [CrossRef] [PubMed]
- Spector, A.C.; Scalera, G.; Grill, H.J.; Norgren, R. Gustatory detection thresholds after parabrachial nuclei lesions in rats. Behav. Neurosci. 1995, 109, 939–954. [Google Scholar] [CrossRef] [PubMed]
- Berridge, K.C.; Robinson, T.E. Parsing reward. Trends Neurosci. 2003, 26, 507–513. [Google Scholar] [CrossRef]
- Abizaid, A.; Horvath, T.L. Ghrelin and the central regulation of feeding and energy balance. Indian J. Endocrinol. Metab. 2012, 16, S617–S626. [Google Scholar] [PubMed]
- Oliveira-Maia, A.J.; Roberts, C.D.; Simon, S.A.; Nicolelis, M.A.L. Gustatory and reward brain circuits in the control of food intake. Adv. Tech. Stand. Neurosurg. 2011, 36, 31–59. [Google Scholar] [PubMed]
- Jhamandas, J.H.; Harris, K.H.; Petrov, T.; Krukoff, T.L. Characterization of the parabrachial nucleus input to the hypothalamic paraventricular nucleus in the rat. J. Neuroendocrinol. 1992, 4, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Coizet, V.; Dommett, E.J.; Klop, E.M.; Redgrave, P.; Overton, P.G. The parabrachial nucleus is a critical link in the transmission of short latency nociceptive information to midbrain dopaminergic neurons. Neuroscience 2010, 168, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Igelstrom, K.M.; Herbison, A.E.; Hyland, B.I. Enhanced c-Fos expression in superior colliculus, paraventricular thalamus and septum during learning of cue-reward association. Neuroscience 2010, 168, 706–714. [Google Scholar] [CrossRef] [PubMed]
- Sárvári, M.; Kocsis, P.; Deli, L.; Gajári, D.; Dávid, S.; Pozsgay, Z.; Hegedűs, N.; Tihanyi, K.; Liposits, Z. Ghrelin modulates the fmri bold response of homeostatic and hedonic brain centers regulating energy balance in the rat. PLoS ONE 2014, 9, e97651. [Google Scholar] [CrossRef] [PubMed]
- Skibicka, K.P.; Dickson, S.L. Ghrelin and food reward: The story of potential underlying substrates. Peptides 2011, 32, 2265–2273. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Clark, M.S.; Palmiter, R.D. Deciphering a neuronal circuit that mediates appetite. Nature 2012, 483, 594–597. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Nakajima, K.; Oomura, Y.; Wayner, M.J.; Sasaki, K. Orexin-A and ghrelin depolarize the same pedunculopontine tegmental neurons in rats: An in vitro study. Peptides 2009, 30, 1328–1335. [Google Scholar] [CrossRef] [PubMed]
- Lanca, A.J.; Adamson, K.L.; Coen, K.M.; Chow, B.L.; Corrigall, W.A. The pedunculopontine tegmental nucleus and the role of cholinergic neurons in nicotine self-administration in the rat: A correlative neuroanatomical and behavioral study. Neuroscience 2000, 96, 735–742. [Google Scholar] [CrossRef]
- Kim, J.; Nakajima, K.; Oomura, Y.; Wayner, M.J.; Sasaki, K. Electrophysiological effects of ghrelin on pedunculopontine tegmental neurons in rats: An in vitro study. Peptides 2009, 30, 745–757. [Google Scholar] [CrossRef] [PubMed]
- Ferrario, C.R.; Labouèbe, G.; Liu, S.; Nieh, E.H.; Routh, V.H.; Xu, S.; Connor, E.C. Homeostasis meets motivation in the battle to control food intake. J. Neurosci. 2016, 36, 11469. [Google Scholar] [CrossRef] [PubMed]
- Bouret, S.G.; Gorski, J.N.; Patterson, C.M.; Chen, S.; Levin, B.E.; Simerly, R.B. Hypothalamic neural projections are permanently disrupted in diet-induced obese rats. Cell Metab. 2008, 7, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Figlewicz, D.; Evans, S.; Murphy, J.; Hoen, M.; Baskin, D. Expression of receptors for insulin and leptin in the ventral tegmental area/substantia nigra (VTA/SN) of the rat. Brain Res. 2003, 964, 107–115. [Google Scholar] [CrossRef]
- Su, J.; Geng, J.; Bao, J.; Tang, Y.; Liu, M.; Yu, H.; Han, Y.; Huang, W.; Zhou, S. Two ghrelin receptor agonists for adults with malnutrition: A systematic review and meta-analysis. Nutr. J. 2016, 15, 97. [Google Scholar] [CrossRef] [PubMed]
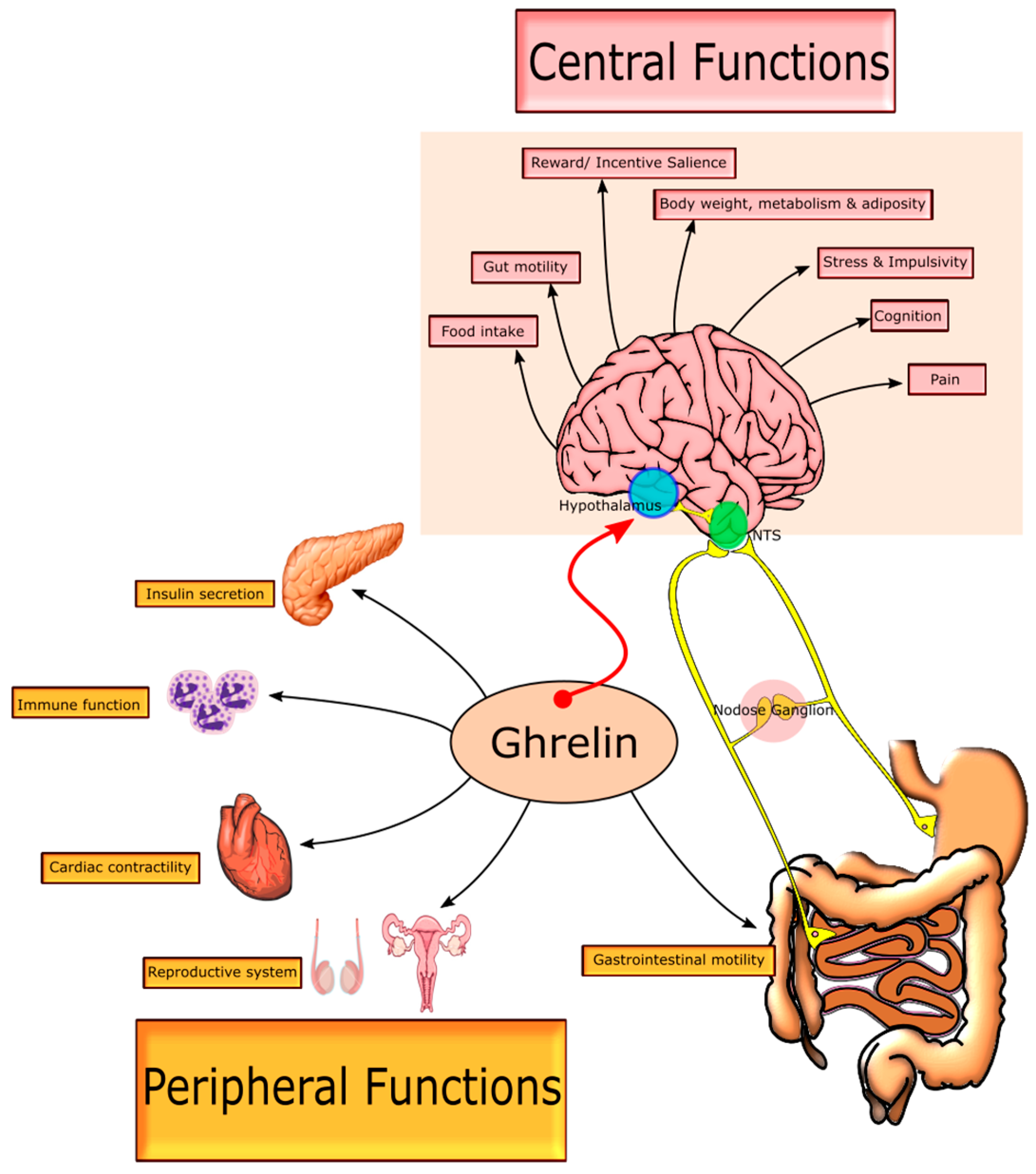
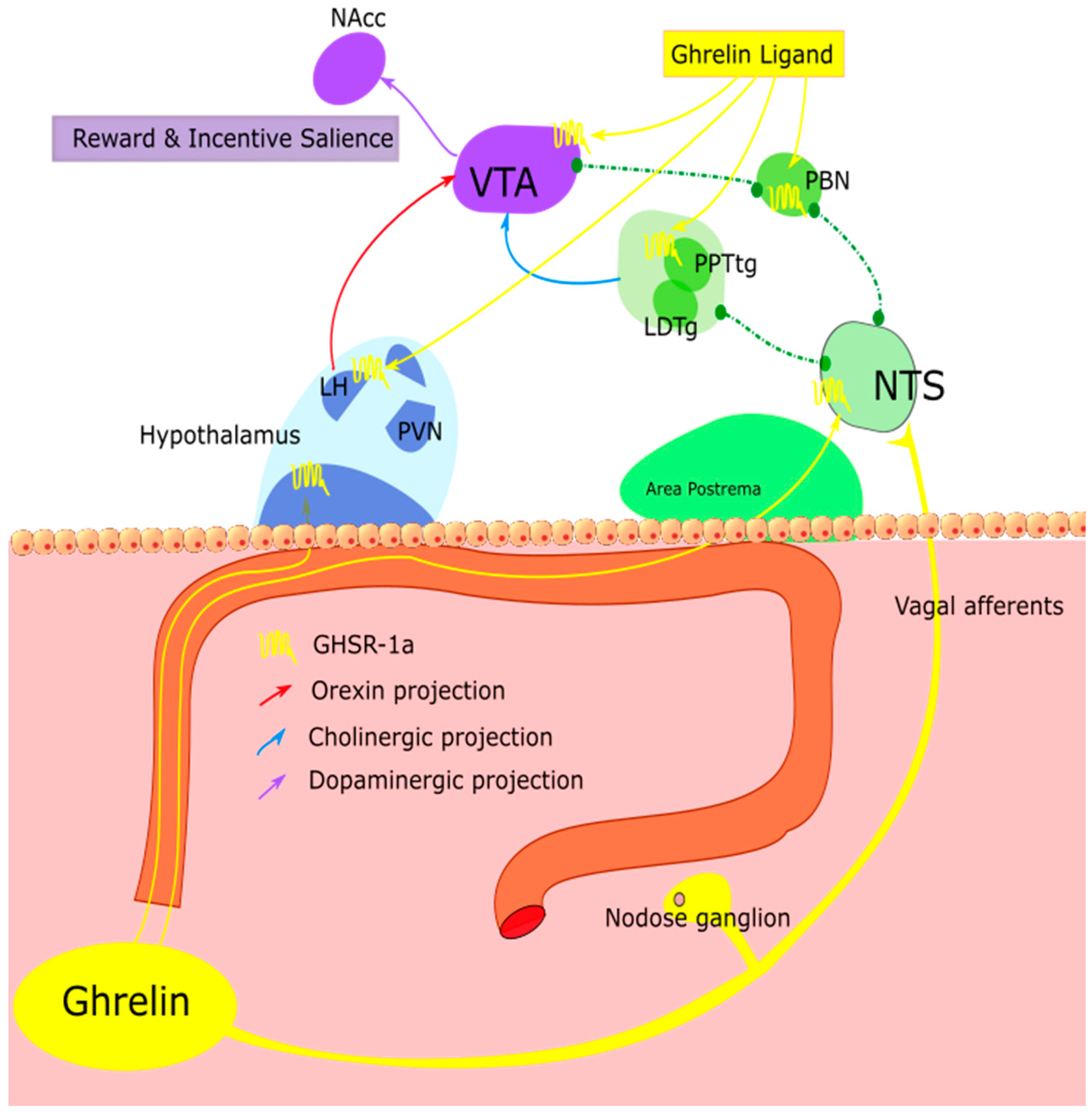

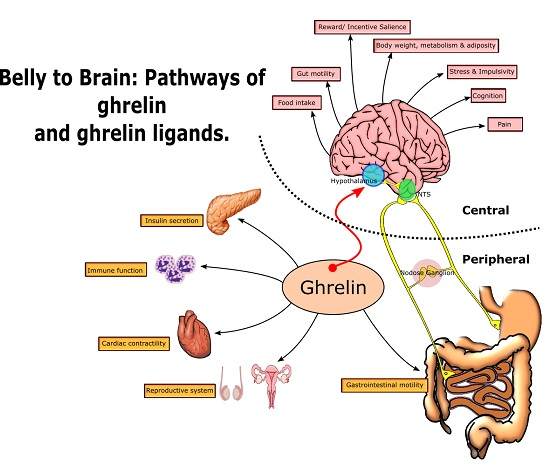{kind=link}
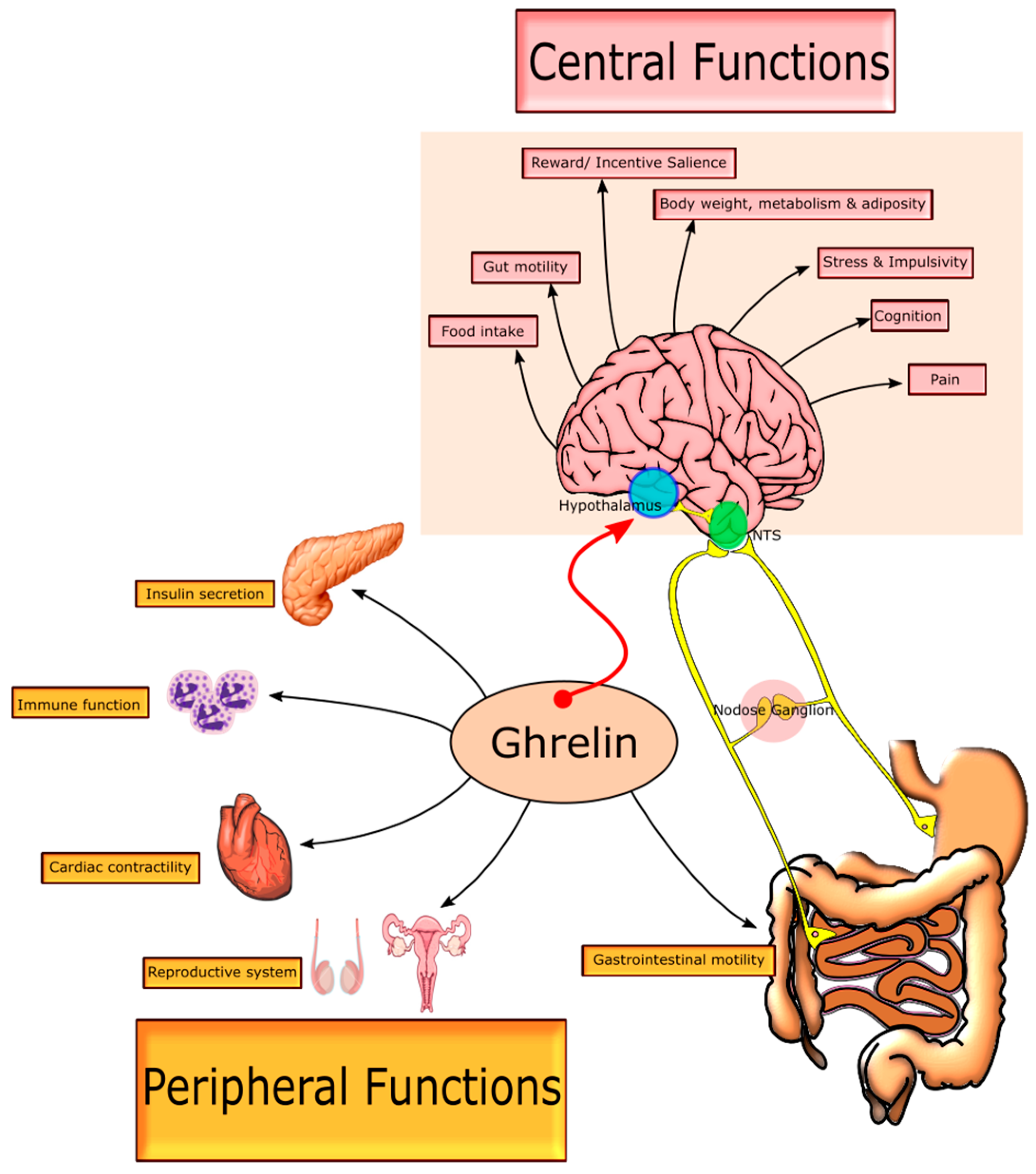{kind=link}
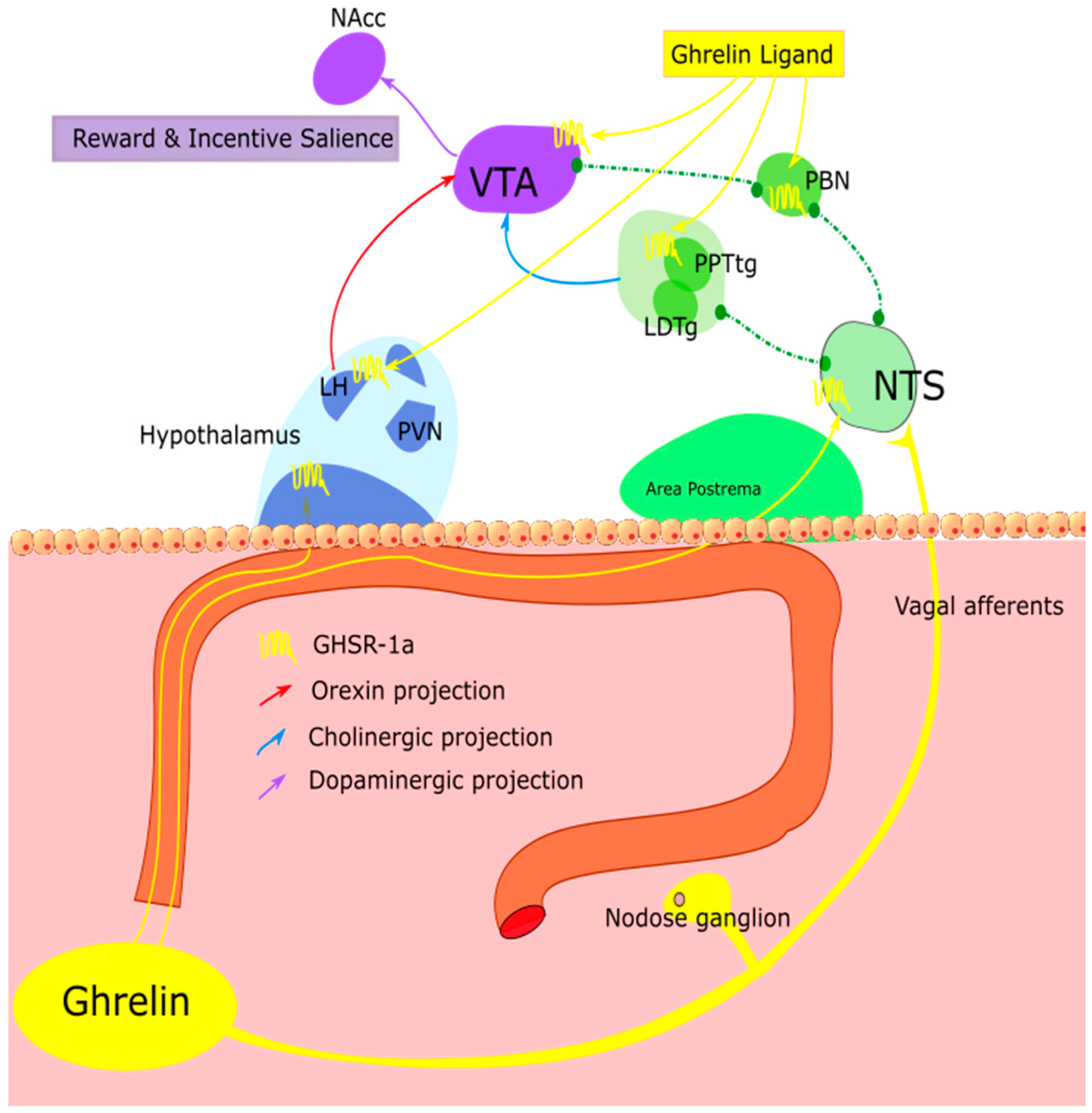{kind=link}
| Status | Dose of Infusion (Duration) | Fed Status | Form Assayed | Mean Serum Ghrelin (pmol/mL) | Average Fold Increase | Time Post-Dose (min) | Reference |
|---|---|---|---|---|---|---|---|
| Acylated | 5 pmol/kg/min (180 min) | Overnight fasted | Total | 1.32 | Not reported | 180 (Tmax) | [219] |
| Acylated | 300 pmol/kg (Bolus) 1500 pmol/kg (Bolus) | Overnight fasted | Total and active | Total: 1.06 Acylated: 0.447 Total: 6.598 Acylated: 3.454 | 4.58 18.7 28.6 145.1 | 15 (Tmax) 15 (Tmax) | [209] |
| Acylated | 3000 pmol/kg (Bolus) | Overnight fasted | Total | 44.5 | 61 | 1 | [220] |
| Acylated | 5 pmol/kg/min (65 min) 15 pmol/kg/min (65 min) 25 pmol/kg/min (65 min) | Overnight fasted | Total and active | Total: 1.647 Acylated: 1.170 Total: 5.139 Acylated: 3.510 Total: 8.619 Acylated: 5.880 | Not reported 118 Not reported 355 Not reported 594 | 45 (Tmax) | [217] |
| Acylated | 84 pmol/kg (Bolus) + 5 pmol/kg/min (65 min) | Overnight fasted | Active and inactive | Acylated: 0.579 Desacylated: 0.350 | 44 17 | 30 (Tmax) | [217] |
| Desacylated | 343 pmol/kg + 20.8 pmol/kg/min (65 min) | Overnight fasted | Active and inactive | Acylated: 0.006 Desacylated: 4.955 | No change 233 | Not specified | [217] |
| Acylated and Desacylated | Acylated: 84 pmol/kg (Bolus) + 5 pmol/kg/min (65 min) Desacylated: 343 pmol/kg + 20.8 pmol/kg/min (65 min) | Overnight fasted | Active and inactive | Acylated: 0.495 Desacylated: 4.644 | 54 272 | Not specified | [217] |
| Acylated | 1 pmol/kg/min (75 min) 5 pmol/kg/min (75 min) | Overnight fasted | Total | 0.725 1.598 | 1.6 3.6 | 45 (Tmax) 45 (Tmax) | [58] |
| Acylated | 1 pmol/kg/min (120 min) 5 pmol/kg/min (120 min) | Not specified | Total | 0.958 4.087 | 3.54 15.13 | 90 90 | [206] |
| Acylated | 0.3 pmol/kg/min (300 min) | Fed | Active | 0.057 | 2.4 | 210 (Tmax) | [216] |
| Acylated | 7.5 pmol/kg/min (120 min) 15 pmol/kg/min (120 min) | Overnight fasted | Total | 0.300 0.494 | 2 3 | 120 (Tmax) 120 (Tmax) | [221] |
| Acylated | 3600 pmol/kg (Subcutaneous) | Overnight fasted | Total and active | Total: 0.988 Acylated: 0.355 | 5.15 10.23 | 15 (Tmax) 30 (Tmax) | [222] |
| Acylated | 300 pmol/kg (Subcutaneous) 1500 pmol/kg 3000 pmol/kg | Overnight fasted | Total | ~0.350 ~0.900 ~1.400 | 2 8 12 | 30 (Tmax) | [223] |
| Agonist | Class of Compound | Oral Bioavailability (Species) | Half Life | Centrally Regulated Parameters Reported |
|---|---|---|---|---|
| Growth Hormone Releasing Peptide 6 (GHRP-6) | Synthetic peptide | 0.3% (Human) [247,278] | 0.3 h [247] | Food intake [262], Body weight [262,279], Gastric emptying [119], Growth hormone [251,279] |
| Hexarelin | Synthetic peptide | <0.3% (Human) [280] | 1.15 h [257,280] | Food intake [258], Growth velocity [281,282,283] |
| Pralmorelin (GHRP-2) | Synthetic peptide | Not reported, but has been dosed orally [284] | 0.52 h [285] | Food intake [86,259], Growth hormone [284,285] |
| Alexamorelin | Synthetic peptide | Not reported | Not reported | Growth hormone [286] |
| Ipamorelin | Synthetic peptide | 1%–6% (Rat, Dog) [287] | 2 h [288] | Growth hormone [287,289], Body weight [287], Gastointestinal motility [290] |
| Capromorelin | Small molecule | 65% [291] (Rat) [292] | 2.4 h [291] | Growth hormone [291,293], Body weight [294], Gastric emptying [119] |
| Relamorelin | Synthetic peptide | Not reported | 19.4 h [295] | Growth hormone [296], Food intake, Body weight [297,298,299], Gastric emptying [300,301] |
| Macimorelin | Small molecule | Not reported, but has been dosed orally [302,303] | 3.8 h [304] | Growth hormone [302,305] |
| Tabimorelin | Synthetic peptide | 30%–35% (Rat) [253,306] | 20.8 h [307,308] | Growth hormone [306,307,308] Body weight [253] |
| Anamorelin | Small molecule | Not reported, but has been dosed orally [309,310,311] | 7 h [309] | Growth hormone [309,312], Food intake [271,310,311,312] |
| Ibutamoren (MK-0677) | Small molecule | >60% (Dog) [254,313,314] | 6 h [180] | Growth hormone [254,313,315], Body weight [316], Fat free mass [314] |
| Ulimorelin | Synthetic peptide | 24% (Rat) [270] | 1.6 h [317,318,319] | Growth hormone (no effect), Food intake, Gastrointestinal motility [270,317,319,320,321,322] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Howick, K.; Griffin, B.T.; Cryan, J.F.; Schellekens, H. From Belly to Brain: Targeting the Ghrelin Receptor in Appetite and Food Intake Regulation. Int. J. Mol. Sci. 2017, 18, 273. https://doi.org/10.3390/ijms18020273
Howick K, Griffin BT, Cryan JF, Schellekens H. From Belly to Brain: Targeting the Ghrelin Receptor in Appetite and Food Intake Regulation. International Journal of Molecular Sciences. 2017; 18(2):273. https://doi.org/10.3390/ijms18020273
Chicago/Turabian StyleHowick, Ken, Brendan T. Griffin, John F. Cryan, and Harriët Schellekens. 2017. "From Belly to Brain: Targeting the Ghrelin Receptor in Appetite and Food Intake Regulation" International Journal of Molecular Sciences 18, no. 2: 273. https://doi.org/10.3390/ijms18020273
APA StyleHowick, K., Griffin, B. T., Cryan, J. F., & Schellekens, H. (2017). From Belly to Brain: Targeting the Ghrelin Receptor in Appetite and Food Intake Regulation. International Journal of Molecular Sciences, 18(2), 273. https://doi.org/10.3390/ijms18020273