1. Introduction
Moyamoya disease (MMD) is a chronic cerebrovascular occlusive disorder that results in transient ischemia, cerebral infarcts, and hemorrhagic strokes [
1,
2,
3,
4,
5]. MMD occurs worldwide, but its prevalence is highest in East Asian countries [
5].
MMD has a bimodal age distribution for peak incidence, with peaks in children who are approximately five years of age and adults in their mid-forties [
1,
3,
6,
7,
8]. Most children MMD patients develop transient ischemic attacks or cerebral infarctions [
9], whereas adult MMD patients are more likely to have a hemorrhagic stroke [
8,
10,
11]. This suggests some variant or impairment of the genetic sequence in the same disease. The majority of MMD cases are sporadic, even though familial MMD cases account for approximately 9–15% of all cases [
12,
13]. Genetic associations with loci on chromosome 3, 6, 8, 10, and 17 and a specific human leukocyte antigen (HLA) haplotype have been reported [
14,
15,
16,
17,
18,
19,
20], but questions about genetic penetrations remain.
RNF213, which is located on chromosome 17q25, has been recognized as the major susceptibility gene for MMD in East Asians [
21,
22], as well as Caucasians and East/South Asians [
22,
23,
24,
25]. An association of the p.R4810K polymorphism with intracranial major artery stenosis/occlusion has been reported in Japanese and Korea populations [
26,
27,
28,
29]. However, this genetic variant associated with MMD was also observed in patients with non-MMD intracranial stenosis [
26,
28,
29]. The association between
RNF213 and clinical manifestations of MMD in young children and adults remain uncertain.
The
RNF213 variant p.R4810K (c.14429G>A, rs112735431) was first reported to have a high level of association with MMD on a large scale [
30]. The
RNF213 variants R4859K [
21] and R4810K [
22] correspond to rs112735431, while R4859K is based on the computer predicted open-reading frame in the database [
21].
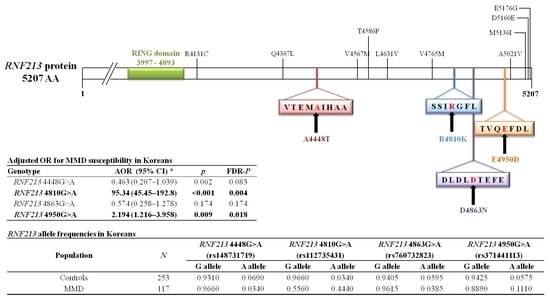
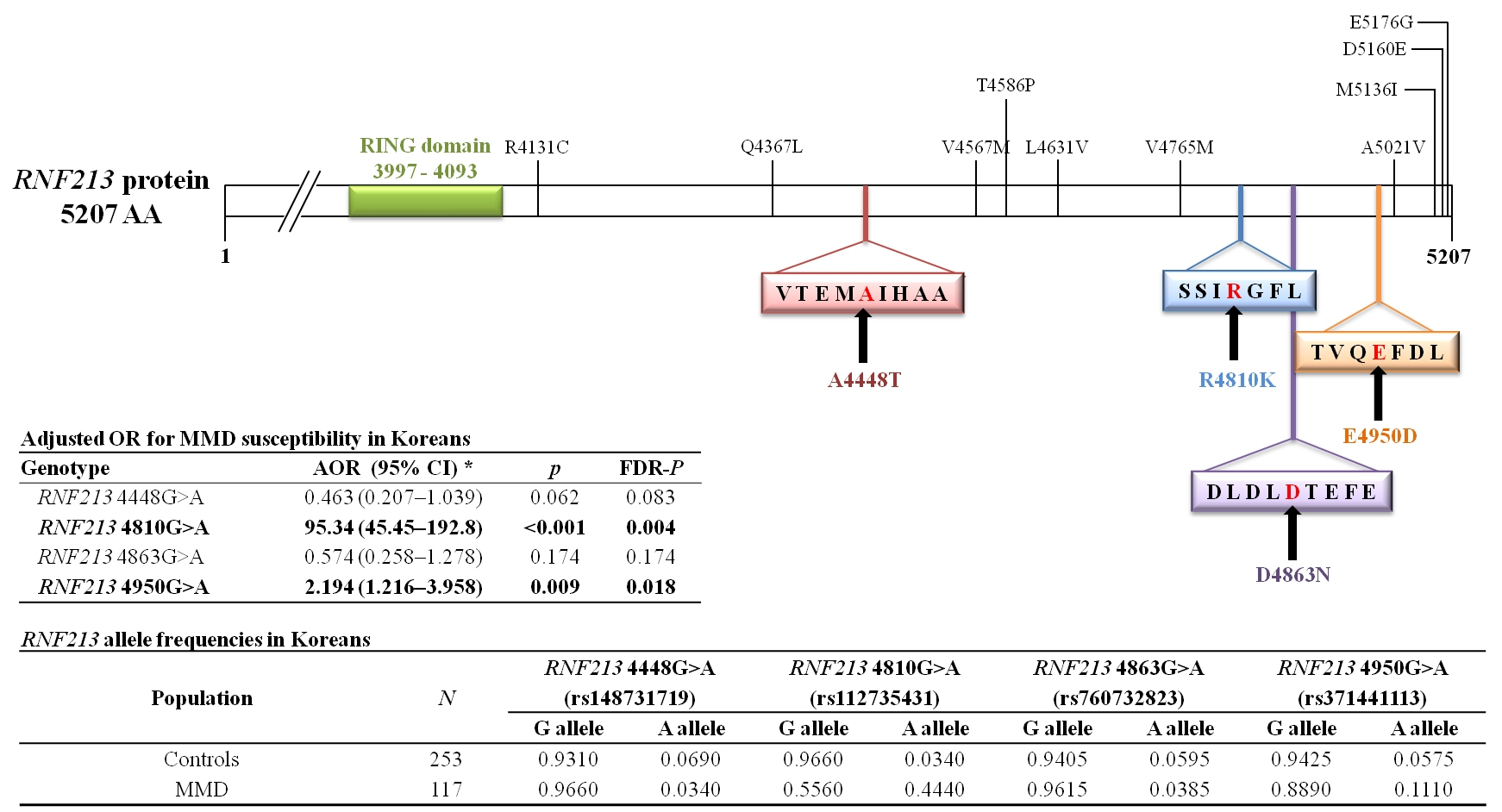
In the present study, four single nucleotide polymorphisms (SNPs) in exon region were identified through previous studies [
21,
22,
29] and a database search:
RNF213 4448G>A,
RNF213 4810G>A,
RNF213 4863G>A, and
RNF213 4950G>A. These four SNPs were selected on the basis of information in the HapMap database (
http://hapmap.ncbi.nlm.nih.gov/). Currently, no study has addressed the frequency of
RNF213 variants (4448, 4810, 4863, 4950) in a Korean population with a high prevalence of MMD. Therefore, we have investigated the frequency of MMD-related
RNF213 variants in a cohort of Korean patients with MMD compared with healthy controls.
3. Discussion
At least 24 genetic changes in the
RNF213 gene have been associated with moyamoya disease [
31,
32]. Three individual studies of MMD patients have revealed high frequencies of the same single base substitution (nonsynonymous mutation) as well as the c.14576G>A (p.R4859K) variant in
RFP213 (a gene located in chromosome 17q) [
21,
22,
23,
24,
25,
26,
27,
28,
29,
30,
31,
32,
33]. The c.14576G>A in
RNF213 is present in ~2% of East Asian populations, which is a relatively higher rate compared with Caucasians [
21,
22,
23,
24,
25,
26,
27,
28,
29,
30,
31,
32,
33]. The
RNF213 gene can be used as a biomarker to predict prognosis, as it has been reported that the
RNF213 gene correlates with the early-onset and severe forms of MMD [
33]. The
RNF213 p.Arg4810Lys variant is reportedly associated with ischemic-type MMD, while
RNF213 non-p.Arg4810Lys variants are associated with hemorrhagic type MMD [
23].
The single-nucleotide polymorphism of c.14576G>A, p.R4859K, in
RNF-213 has been detected in 95% of familial cases and 79% of sporadic cases of MMD [
21]. Nevertheless, some MMD patients do not carry the c.14576G>A variant and this portion is higher in western countries [
34].
Miyawaki et al. found that
RNF213 (14576G>A) was higher in intracranial major artery stenosis/occlusion (ICASO) patients, as in the MMD group [
26], and they suggest ICASO without signs of MMD is a genetic variant associated with MMD. Bang et al. suggest that
RNF213 is a susceptibility gene for both MMD and ICAS in East Asians. Further studies are needed on
RNF213 variants in ICASO patients outside East Asian populations [
27]. However, two variants of
RNF213 (p.Arg2438Cys and p.Ala2826Thr) were found in intracranial aneurysm patients in a French-Canadian population [
35]. Interestingly, our results differed with the results of a previous study [
27] that suggested that 4810G>A is the only variant that is strongly associated with MMD in Korean patients. In this study, our results suggested that
RNF213 4810 and 4950 were associated with MMD risk, furthermore our result found that frequencies of
RNF213 4448G>A, 4863G>A, and 4950G>A hetero genotype. We think that this difference is due to the difference in how the genotypes were identified. We used the classical method of using polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) and confirmed the genotypes of the individual samples, whereas the previous study performed the MALDI-TOF method, which is likely to cause errors because it confirms the genotype of many samples for many variants.
The
RNF213 gene encodes a protein with 5256 amino acids harboring a RING (Really Interesting New Gene) finger motif and an AAA (ATPase associated with a variety of cellular activities) domain, indicating the presence of both E3 ubiquitin ligase activity and an energy-dependent E3 ubiquitin ligase (which has several subtypes), an enzyme that ubiquitinates specific target proteins, resulting in degradation by proteasomes [
26]. The
RNF213 variant associated with MMD prevails, but it is also found in other vascular diseases such as cerebrovascular stenosis [
26], albeit not in the Caucasian MMD population [
36]. In
RNF213-deficient mice, an abnormal vascular network does not develop at the base of the brain [
37]. The
RNF213 variant is an important SNP that is not specific to MMD alone.
The c.14576G>A variant is mainly detected in Japanese, Korean, and Chinese populations [
33,
38,
39]. However, the frequency in the latter population is much lower than those of the former two populations [
34].
RNF213 exhibits strong and obvious ethnic diversity [
32].
The biochemical function and pathological role of
RNF213 have not been completely clarified. Disruption of the RNF213 first B motif disrupts ATP hydrolysis cyclicity, inhibiting angiogenesis, and this reduced anti-angiogenic activity of interferon beta 1 (IFNb) is partially mediated by
RNF213, which acts as a mediator downstream of the IFNb signaling pathway [
32]. The overexpression of
RNF213 R4810K, but not wild type
RNF213, suggests that
RNF213 R4810K over expression mimics IFNb action. Koizumi et al. suggest that
RNF213 R4810K is the major detrimental factor that elicits endothelial cell dysfunction. Pro-inflammatory signals such as IFNs can activate the transcription of
RNF213 [
32]. The model assumes that any of three independent abnormalities, such as endothelial dysfunction, smooth muscle cell dysfunction, and abnormal hemostasis, can exaggerate the proliferation of smooth muscle cells (SMCs) and that each abnormality can result in vascular stenosis [
32].
Genome-wide association study (GWAS) approaches are now being applied to MMD with the hope of uncovering the underlying pathogenic mechanisms [
40]. A GWAS was recently performed in Japanese MMD patients and demonstrated a strong association of MMD risk with chromosome 17q25-ter [
21]. These GWAS studies will need further investigation to solidly replicate the results in modern genetic studies based on familial or non-familial MMD. Subgroup analysis was performed for adult versus pediatric and ischemic versus hemorrhagic groups. The former primarily presents with ischemia, while the latter presents with intracranial hemorrhage [
6,
41]. The progression of MMD has generally been considered to occur exclusively in childhood, with angiographic characteristics completed before adulthood [
42,
43]. Kuroda and colleagues reported in a multicenter observational study on adult onset MMD that the incidence of the disease progression in adult cases was not as rare as originally considered [
44]. Miyatake et al. [
33] reported that patients with childhood-onset MMD and the homozygous polymorphism of c.14576G>A in
RNF213 were more likely to have an earlier onset and more severe mortality because of the rapid progression of vascular stenosis. Han et al. suggested that
RNF213 R4810K is associated with the ischemic type , and A4399T is associated with the hemorrhagic type [
23]. A genetic animal
RNF213 knock-in model has been debated, however, the
RNF213 genetic animal model failed to mimic the MMD model. Liu reported that
RNF213 knock-down zebra fish have abnormal surrounding vessels [
22]. Sonobe et al. [
37] did not observe any modification of angiogenesis after they generated mice that lack
RNF213.
RN 213 knock out (KO) animal models have yielded conflicting results in the cerebrum and hind limbs [
45]. Fujimura et al. speculated that
RNF213 influences vascular remodeling in chronic ischemia [
38]. Allele frequencies of
RNF213 polymorphisms (4448G>A, 4810G>A, 4863G>A, and 4950G>A) in different world populations are presented in
Table 8.
SNP studies have some limitations. First, a question remains about how the p.R4810K variant or nine other variants impair the physiological function of
RNF213, resulting in moyamoya disease [
22]. The second limitation is a lack of a pathologic process for MMD development from
RNF213. Third, large population-based case-control analyses or analyses centered on family-based designs are needed. However, SNP studies have many advantages over other genetic studies, the benefits of which depend on how SNPs will be exploited in relevant study designs and what traits and diseases will be the focus of these studies [
46].
We have considered some of the unique aspects of SNPs and their relative advantages and disadvantages in human population-based analyses [
46]. We believe that wide-scale progress in genetically identifying MMD is needed because MMD appears to be a multifactorial, polygenic spectrum disorder.
4. Materials and Methods
4.1. Subjects
One hundred seventeen consecutive Korean patients with moyamoya disease (mean age, 23.20 ± 17.75 years; 77 females (65.8%), 40 males (34.2%)) were recruited for this study. MMD patients were diagnosed and enrolled based on the presence of clinical ischemic or hemorrhagic symptoms in combination with vascular lesions in magnetic resonance imaging (MRI) or magnetic resonance angiography (MRA) [
47].
The control group was comprised of 253 healthy subjects (mean age 25.60 ± 16.98 years; 145 females (57.3%); 108 males (42.7%)) from the same regional background as the MMD patients. We recruited the age- and sex-matched subjects from outpatient clinics at Severance Hospital, CHA Bundang Medical Center, Chungbuk National University Hospital (Cheongju, Korea).
Table 1 shows demographic characteristics of patients with the disease and control subjects. We divided the MMD patients into pediatric (<18 years) and adult (≥18 years) groups. We further divided the moyamoya patients into ischemic or hemorrhagic groups based on clinical and MRI findings.
All participants gave informed written consent prior to enrollment in the study. The institutional review boards of Severance Hospital (4-2008-0308), CHA Bundang Medical Center (PBC09-103), and Chungbuk National University Hospital (2014-08-010-005) approved this study.
4.2. RNF213 Genotyping
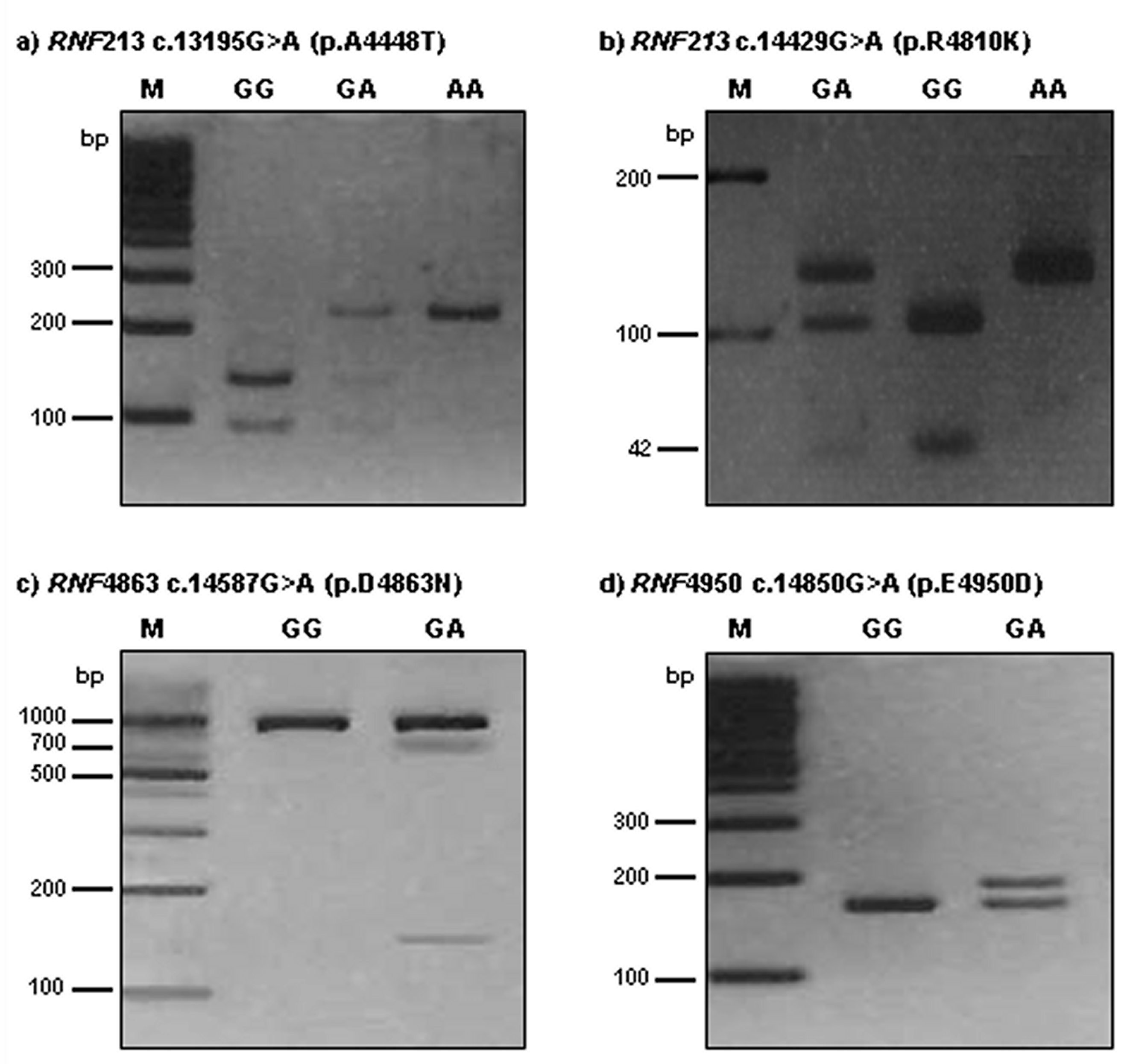
DNA was extracted from leukocytes using a G-DEX II Genomic DNA Extraction kit (Intron Biotechnology, Seongnam, Korea) according to the manufacturer’s instructions. To analyze RNF213 genotypes, we chose polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP).
RNF213 4448G>A was detected using a forward primer (5′-TTG CCA ACT AAG CCC TCG AAA CAA-3′) and a reverse primer (5′-CAA CAA TGG CAC AGA ATT GTC-3′). The 230-bp PCR product was then digested with 5U AluI. A digestion product of 230-bp represented the AA genotype; fragments of 230-bp, 135-bp, and 95-bp represented the AG genotype; 135-bp and 95-bp products represented the GG genotype.
The RNF213 4810G>A polymorphism was detected by PCR-RFLP analysis using forward (5′-AGC AGA GCT GAG GCT GGT AA-3′) and reverse (5′-CTG TCA GAG CAG AGC CAC AC-3′) primers. The 151-bp product was digested with 3U Hpy188I. A restriction fragment of 146-bp and 5-bp represented the AA genotype; fragments of 146-bp, 104-bp, 42-bp, and 5-bp represented the GA genotype; and 104-bp, 42-bp, and 5-bp products represented the GG genotype.
To detect the RNF213 4863G>A genotypes, PCR-RFLP analysis was performed with forward (5′-TGT GTG TGG AGC TGA TGG CT-3′) and reverse (5′-AGG GAG GAG ATA CAG ACC AGA CT-3′) primers. The length of the amplified fragment was 867-bp. PCR products were digested with 5U Hpy188I. For 4863G>A, restriction products of 867-bp identified the GG genotype; products of 867-bp, 719-bp, and 148-bp represented the GA genotype; and 719-bp and 148-bp products represented the AA genotype.
The RNF213 4950G>A polymorphism was detected by PCR-RFLP analysis using forward (5′-GGT GGA GGA GGG CAG AGA GAC CGT GCA CGA-3′) and reverse (5′- CTT CCC TCT CTC GAG AAA CAC ACC AA-3′) primers. The 188-bp product was digested with 3U BssSI. A restriction fragment of 162-bp and 26-bp represented the GG genotype; fragments of 188-bp, 162-bp, and 26-bp represented the GA genotype; and 188-bp products represented the AA genotype. The RNF213 4448G>A, 4810G>A, 4863G>A, and 4950G>A polymorphisms were digested by AluI, Hpy188I, Hpy188I, and BssSI, respectively, for 16 h at 37 °C (New England BioLabs, Beverly, MA, USA). The PCR annealing temperature was 64 °C for all polymorphisms, with 35 amplification cycles for RNF213 polymorphisms. The reaction product (12 μL) was run on a 3.0% ethidium bromide-stained agarose gel and confirmed under ultraviolet illumination.
We randomly repeated approximately 10% of the PCR assays for each of the miRNA polymorphisms and checked the results for concordance by DNA sequencing using an automatic sequencer (ABI3730x1 DNA analyzer; Applied Biosystems, Foster City, CA, USA). The concordance of the quality control samples was 100%. Polymorphism analysis was performed for
RNF213 4448, 4810, 4863, and 4950 in moyamoya disease patients. Polymorphism analysis of
RNF213 gene amplicons was performed by agarose gel electrophoresis after restriction endonuclease digestion (
Figure 1).
In this study, we investigated exome analysis and identified ring finger protein 213 (
RNF213; DDBJ/EMBL/GenBank accession number AB537889) (National Center for Biotechnology Information (NCBI) in
Table S5). Questions remain regarding how the p.R4810K variant or nine other variants impair RNF213 protein, thereby resulting in MMD.
4.3. Statistical Analyses
The genotype frequencies for
RNF213 polymorphism were compared by Hardy-Weinberg equilibrium (HWE) test [
48]. To analyze the demographic characteristics of moyamoya disease, we used the Mann–Whitney and chi-square (χ
2) tests for continuous and categorical data, respectively. The relationships between
RNF213 and MMD patients (pediatric or adult) were calculated according to the odds ratios (ORs) and 95% confidence intervals (CIs) using Fisher’s exact test. The adjusted odds ratios (AORs) for four polymorphisms of the
RNF213 gene were calculated using multiple logistic regression analyses with gender and age. We considered Hardy-Weinberg equilibrium in the genotype distribution. We used GraphPad Prism 4.0 (GraphPad Software, Inc., San Diego, CA, USA) and StatsDirect software (version 2.4.4; StatsDirect Ltd., Altrincham, UK) to perform statistical analyses. We used HAPSTAT (version 3.0; University of North Carolina, Chapel Hill, NC, USA) and Haploview 4.2 (Broad Institute, Cambridge, MA, USA) to perform Haplotype analyses. The adjusted ORs and 95% CIs were calculated by statistical software (version 2.4.4; StatsDirect Ltd., Altrincham, UK).


 ,
, 


{kind=link}
{kind=link}