Topical Ocular Delivery of TGF-β1 to the Back of the Eye: Implications in Age-Related Neurodegenerative Diseases




 , and
, and 
Abstract
:
1. Introduction
2. Results
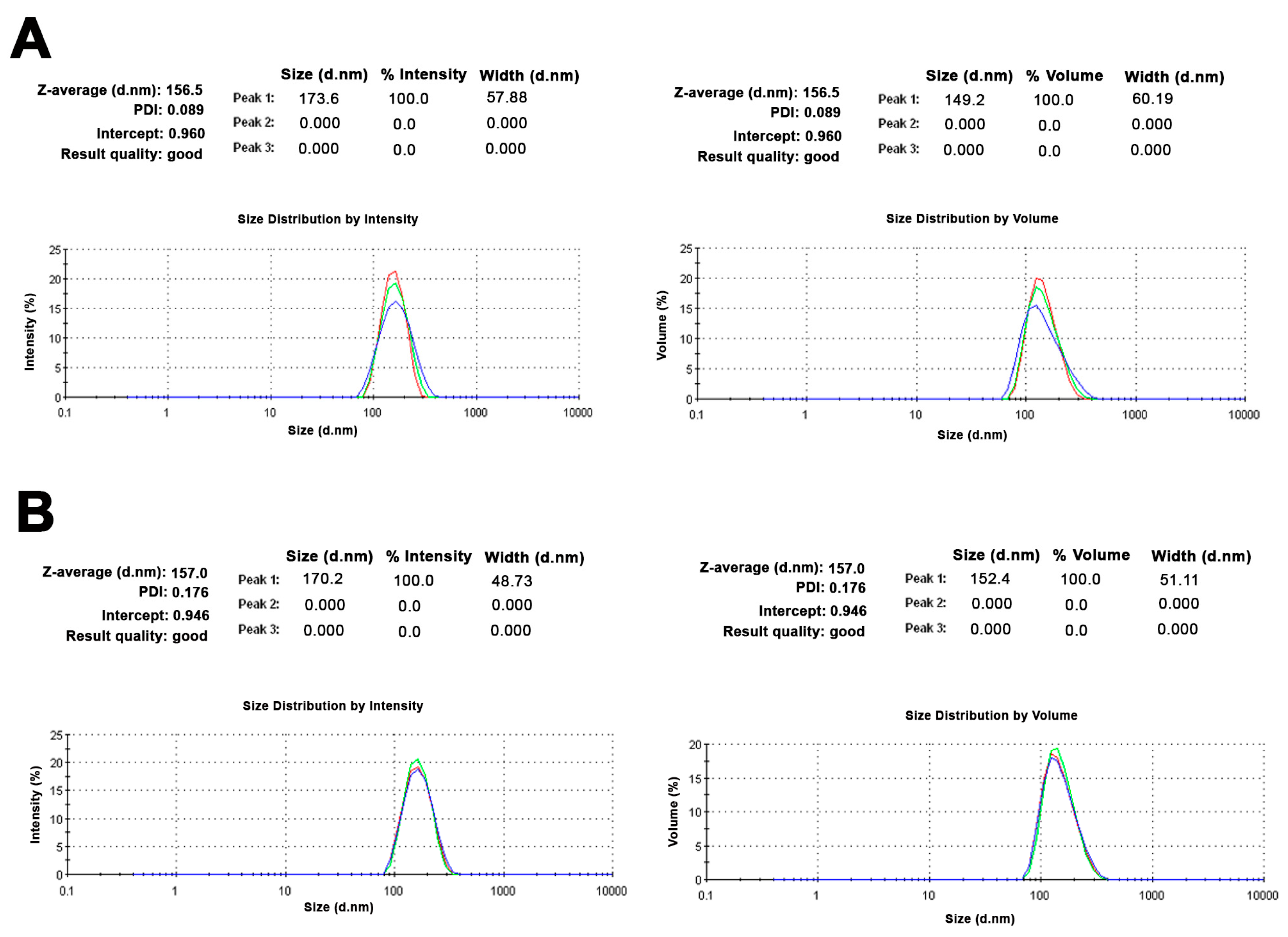
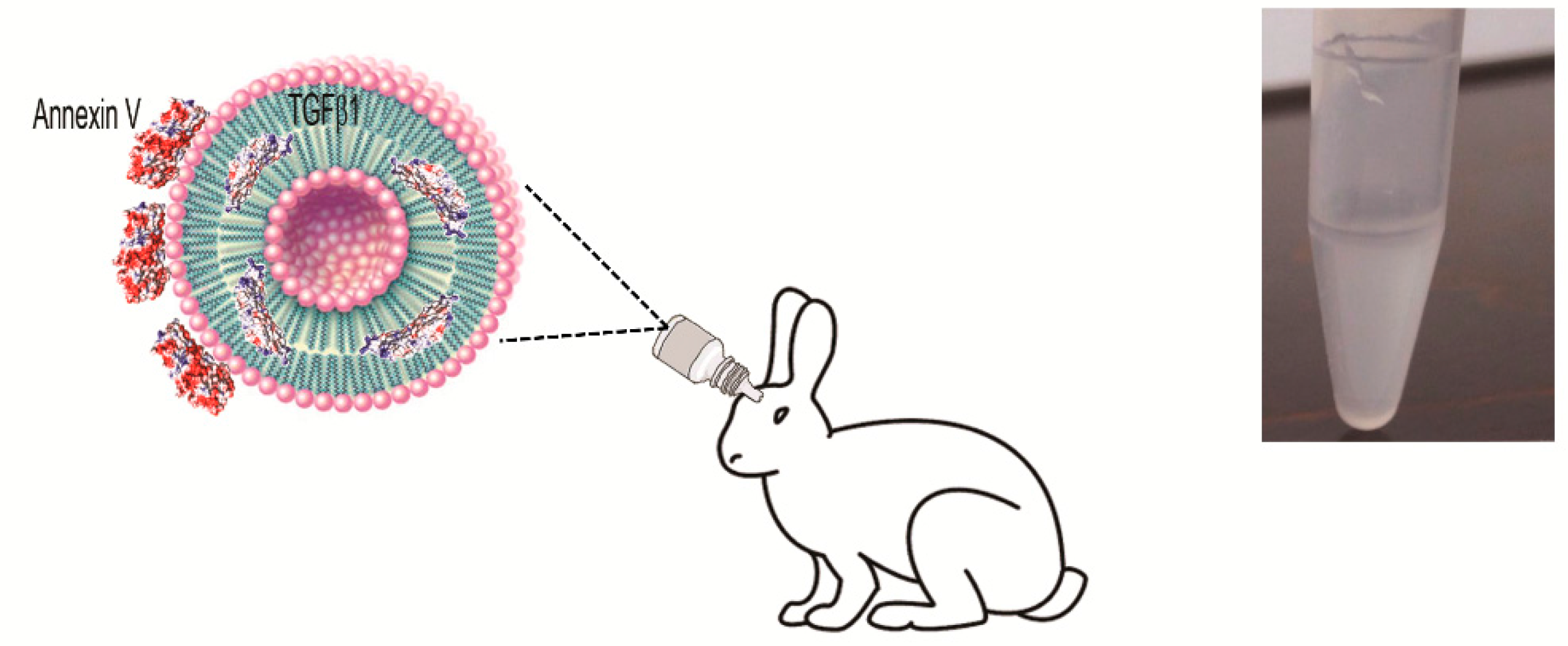
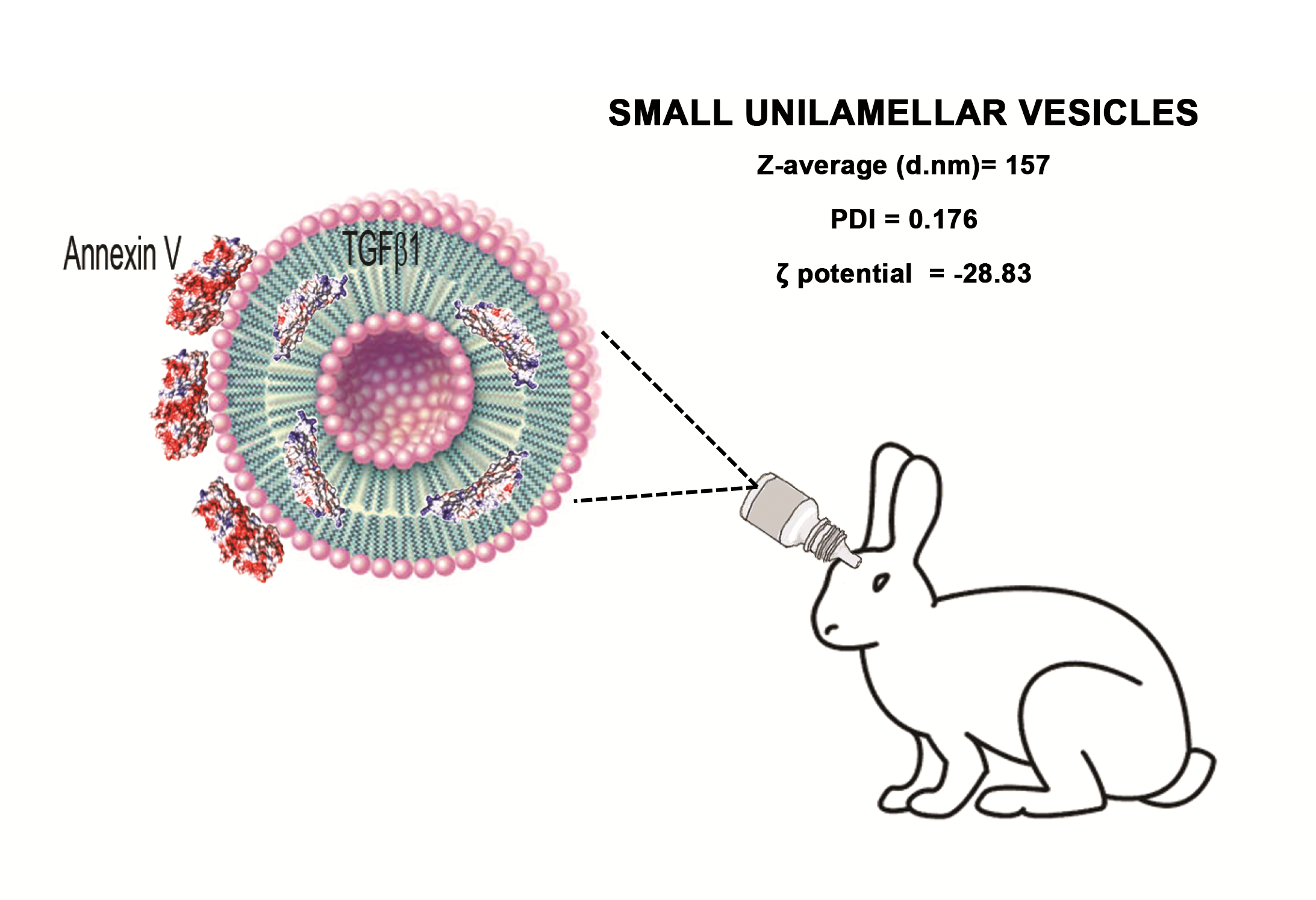
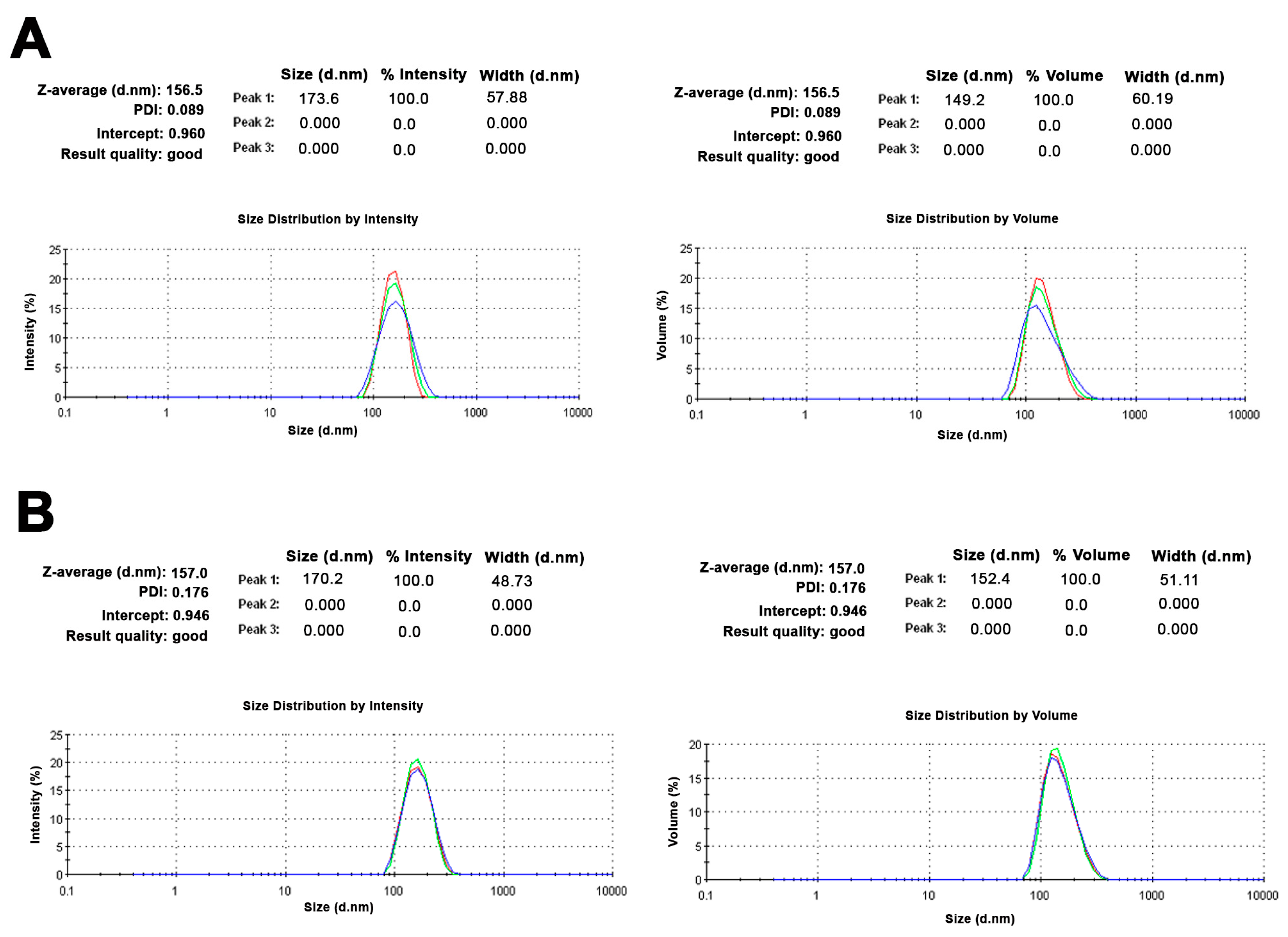
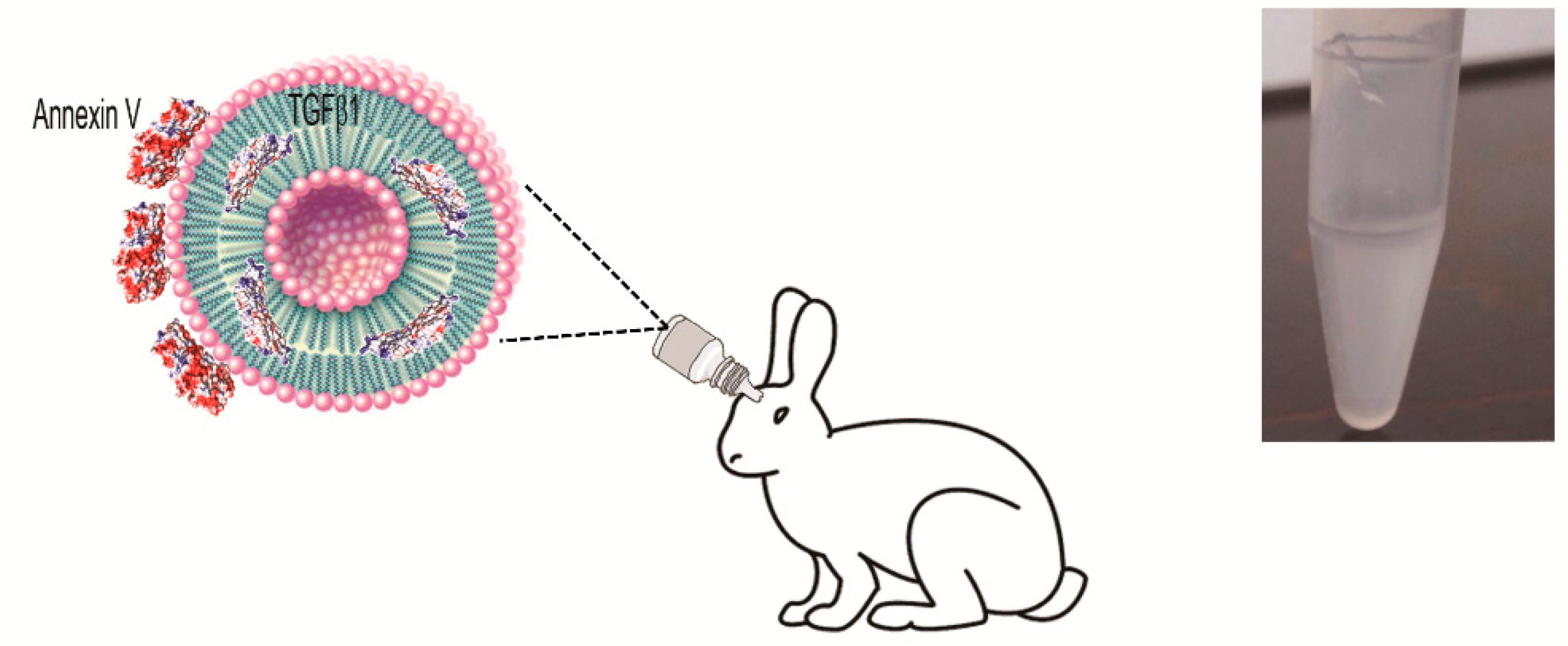
2.1. Small Unilamellar Vesicles Preparation and Characterization
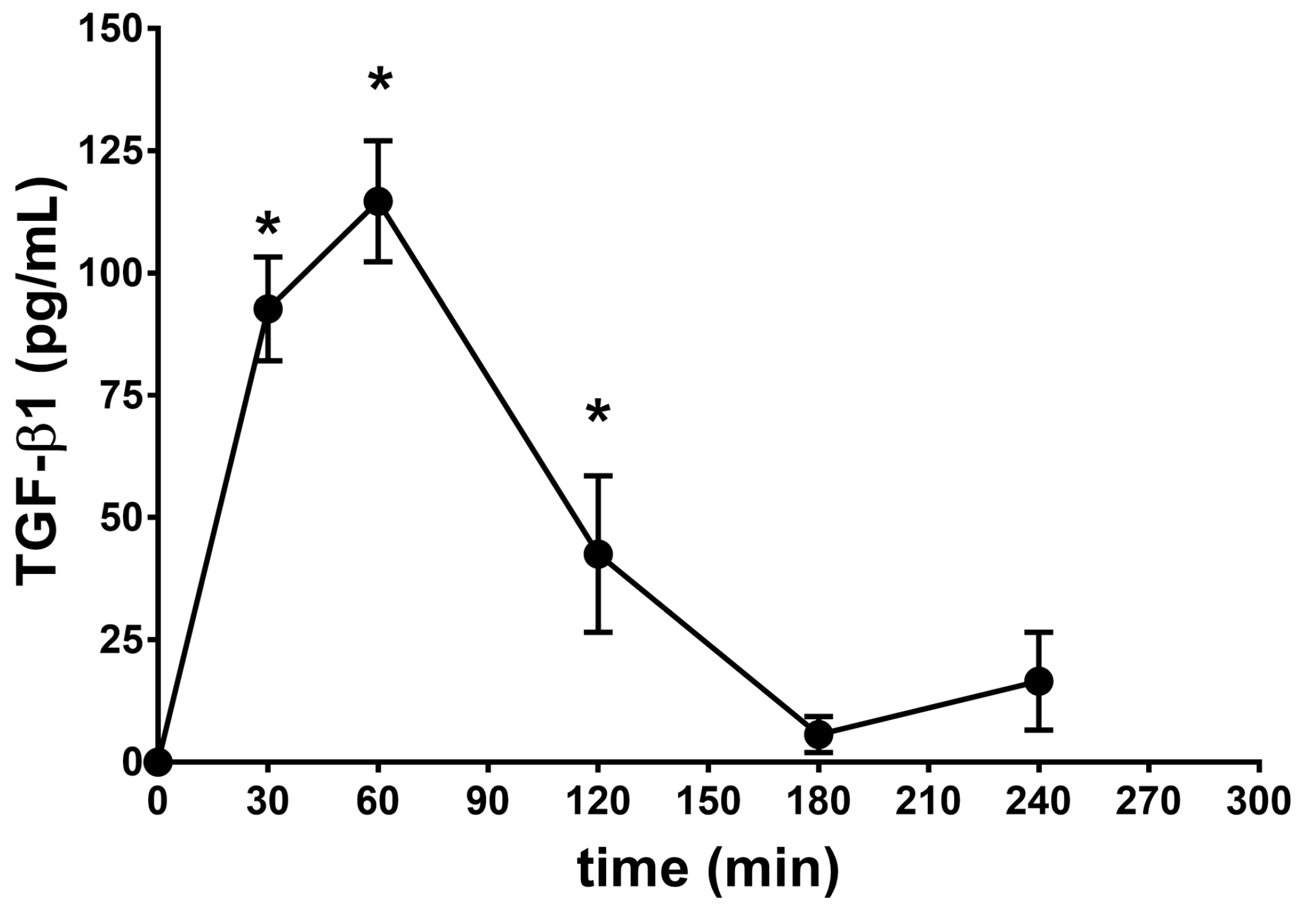
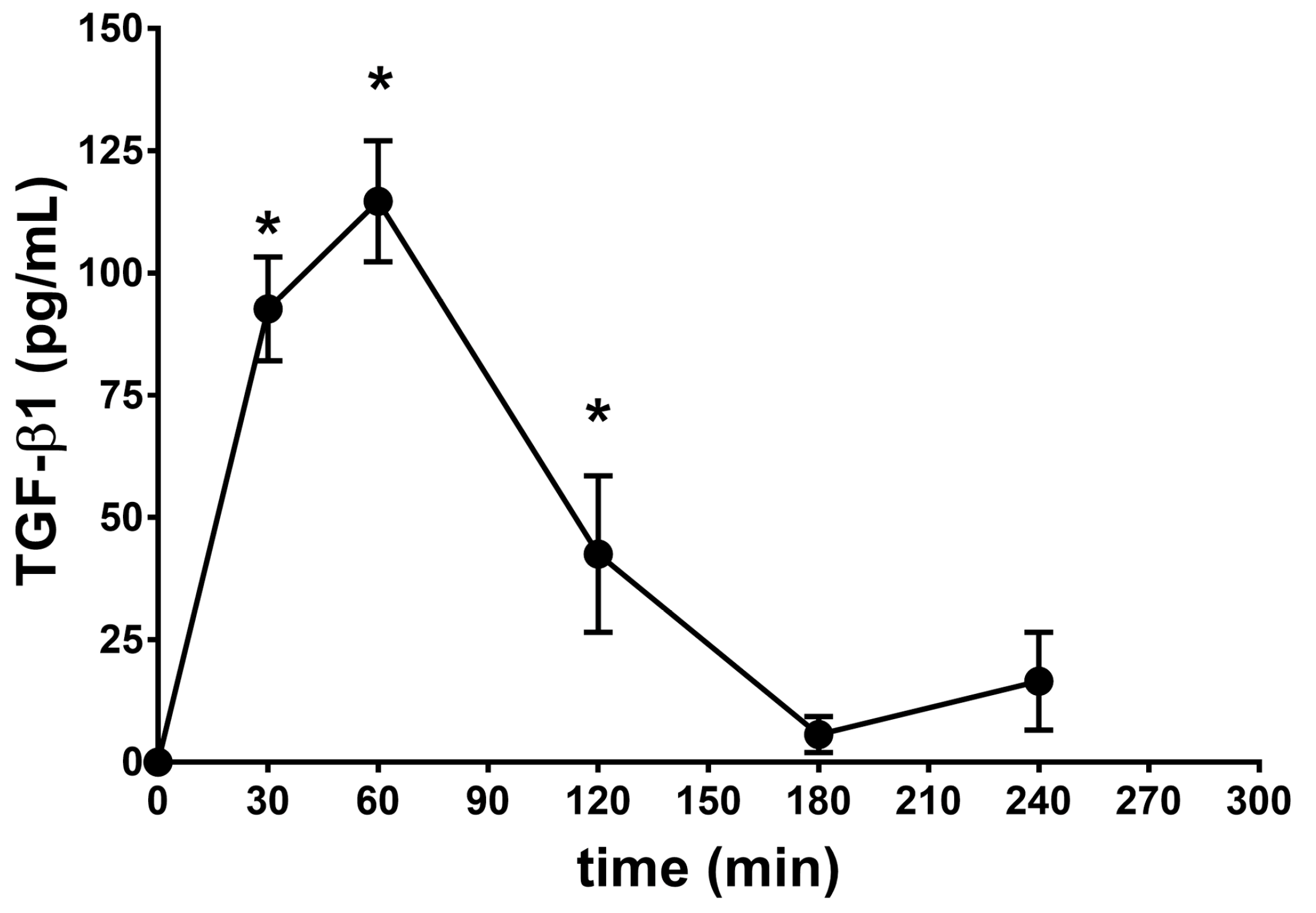
2.2. Vitreous Availability and Ocular Tolerability of TGF-β1 Eye Drops
3. Discussion
4. Material and methods
4.1. Unilamellar Vesicle Preparation
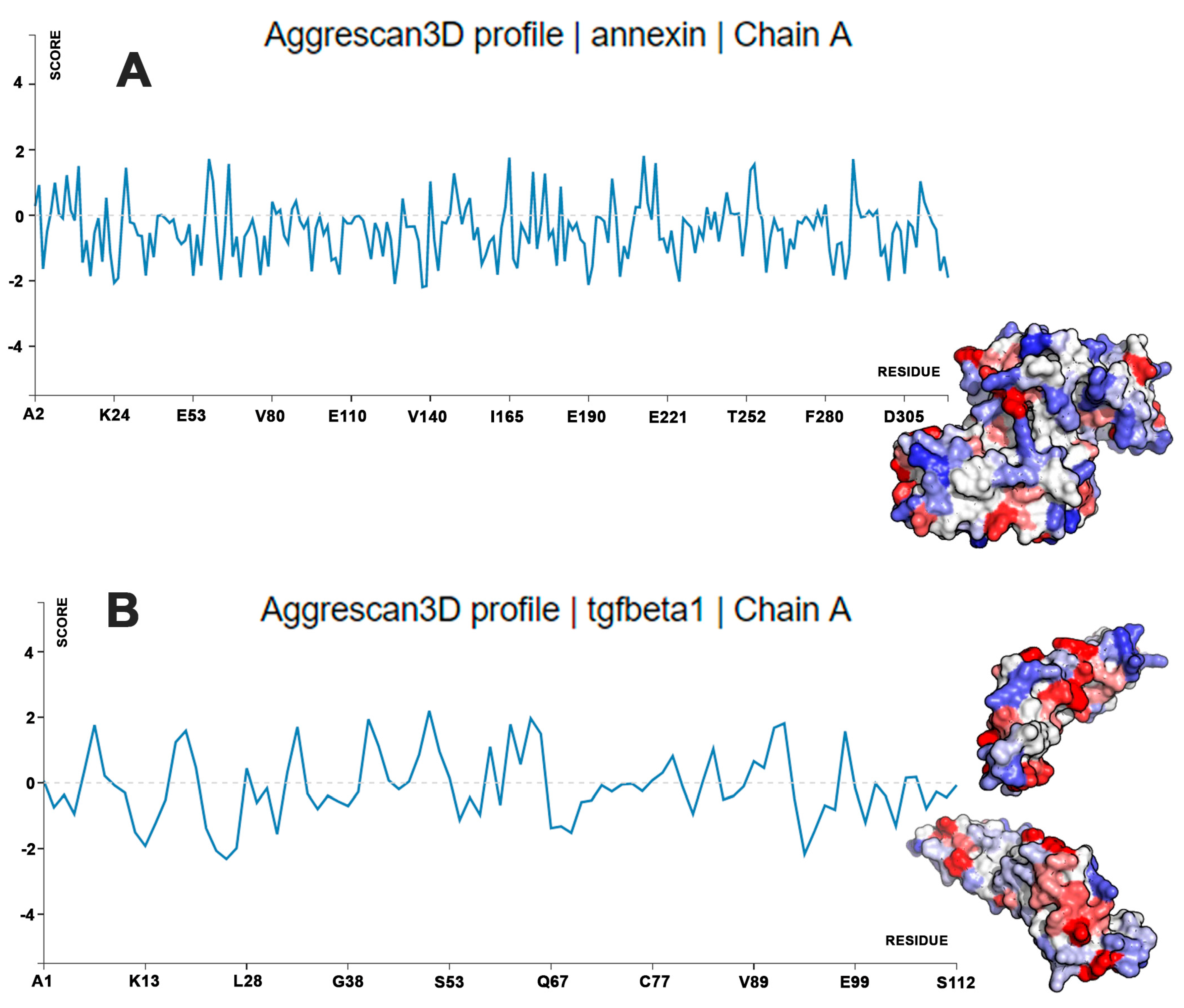
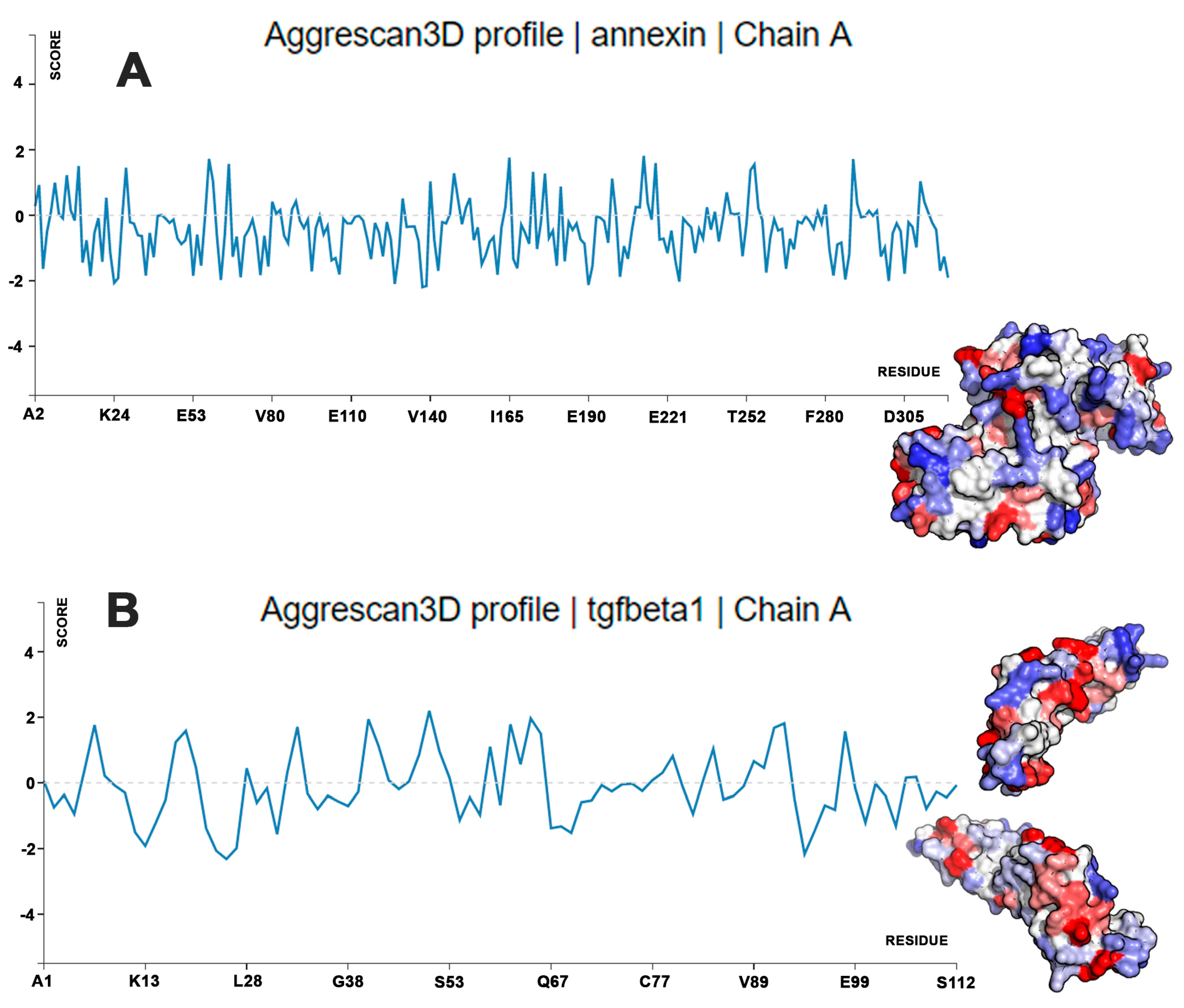
4.2. Calculations of Protein Aggregation Propensity and Water Stability
4.3. In Vivo Study
4.4. TGF-β1 Measurements in Rabbit Vitreous
4.5. Ocular Tolerability Assessment
4.6. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ten Dijke, P.; Hill, C.S. New insights into TGF-β-SMAD signalling. Trends Biochem. Sci. 2004, 29, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Taipale, J.; Saharinen, J.; Keski-Oja, J. Extracellular matrix-associated transforming growth factor-β: Role in cancer cell growth and invasion. Adv. Cancer Res. 1998, 75, 87–134. [Google Scholar] [PubMed]
- Dubois, C.M.; Laprise, M.H.; Blanchette, F.; Gentry, L.E.; Leduc, R. Processing of transforming growth-factor-β-1 precursor by human furin convertase. J. Biol. Chem. 1995, 270, 10618–10624. [Google Scholar] [CrossRef] [PubMed]
- Taipale, J.; Miyazono, K.; Heldin, C.H.; Keskioja, J. Latent transforming growth-factor-β-1 associates to fibroblast extracellular-matrix via latent TGF-β binding-protein. J. Cell Biol. 1994, 124, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Vivien, D.; Ali, C. Transforming growth factor-β signalling in brain disorders. Cytokine Growth F. R. 2006, 17, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Luibl, V.; Isas, J.M.; Kayed, R.; Glabe, C.G.; Langen, R.; Chen, J. Drusen deposits associated with aging and age-related macular degeneration contain nonfibrillar amyloid oligomers. J. Clin. Investig. 2006, 116, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Isas, J.M.; Luibl, V.; Johnson, L.V.; Kayed, R.; Wetzel, R.; Glabe, C.G.; Langen, R.; Chen, J. Soluble and mature amyloid fibrils in drusen deposits. Investig. Ophthalmol. Vis. Sci. 2010, 51, 1304–1310. [Google Scholar] [CrossRef] [PubMed]
- Fisichella, V.; Giurdanella, G.; Platania, C.B.; Romano, G.L.; Leggio, G.M.; Salomone, S.; Drago, F.; Caraci, F.; Bucolo, C. TGF-β1 prevents rat retinal insult induced by amyloid-β1–42 oligomers. Eur. J. Pharmacol. 2016, 787, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Caraci, F.; Spampinato, S.; Sortino, M.A.; Bosco, P.; Battaglia, G.; Bruno, V.; Drago, F.; Nicoletti, F.; Copani, A. Dysfunction of TGF-β1 signaling in alzheimer's disease: Perspectives for neuroprotection. Cell Tissue Res. 2012, 347, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Mocali, A.; Cedrola, S.; Della Malva, N.; Bontempelli, M.; Mitidieri, V.A.M.; Bavazzano, A.; Comolli, R.; Paoletti, F.; La Porta, C.A.M. Increased plasma levels of soluble CD40, together with the decrease of TGF-β1, as possible differential markers of alzheimer disease. Exp. Gerontol. 2004, 39, 1555–1561. [Google Scholar] [CrossRef] [PubMed]
- Juraskova, B.; Andrys, C.; Holmerova, I.; Solichova, D.; Hrnciarikova, D.; Vankova, H.; Vasatko, T.; Krejsek, J. Transforming growth factor β and soluble endoglin in the healthy senior and in alzheimer's disease patients. J. Nutr. Health Aging 2010, 14, 758–761. [Google Scholar] [CrossRef] [PubMed]
- Lauzon, M.A.; Daviau, A.; Marcos, B.; Faucheux, N. Growth factor treatment to overcome alzheimer's dysfunctional signaling. Cell Signal. 2015, 27, 1025–1038. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.H.; Ke, K.F.; Lu, J.H.; Qiu, Y.H.; Peng, Y.P. Protection of TGF-β1 against neuroinflammation and neurodegeneration in a β1–42-induced alzheimer's disease model rats. PLoS ONE 2015, 10, e0116549. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, T.; Suzuki, K.; Matsushima, T.; Yamakawa, N.; Suzuki, T.; Mizushima, T. Suppression of alzheimer's disease-related phenotypes by geranylgeranylacetone in mice. PLoS ONE 2013, 8, e76306. [Google Scholar] [CrossRef] [PubMed]
- Romano, G.L.; Platania, C.B.M.; Drago, F.; Salomone, S.; Ragusa, M.; Barbagallo, C.; Di Pietro, C.; Purrello, M.; Reibaldi, M.; Avitabile, T.; et al. Retinal and circulating mirnas in age-related macular degeneration: An in vivo animal and human study. Front. Pharmacol. 2017, 8, 168. [Google Scholar] [CrossRef] [PubMed]
- Rowe-Rendleman, C.L.; Durazo, S.A.; Kompella, U.B.; Rittenhouse, K.D.; Di Polo, A.; Weiner, A.L.; Grossniklaus, H.E.; Naash, M.I.; Lewin, A.S.; Horsager, A.; et al. Drug and gene delivery to the back of the eye: From bench to bedside. Investig. Ophth Vis. Sci 2014, 55, 2714–2730. [Google Scholar] [CrossRef] [PubMed]
- Caraci, F.; Gili, E.; Calafiore, M.; Failla, M.; La Rosa, C.; Crimi, N.; Sortino, M.A.; Nicoletti, F.; Copani, A.; Vancheri, C. TFG-β1 targets the GSK-3β/β-catenin pathway via ERK activation in the transition of human lung fibroblasts into myofibroblasts. Pharmacol. Res. 2008, 57, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Joseph, M.; Trinh, H.M.; Cholkar, K.; Pal, D.; Mitra, A.K. Recent perspectives on the delivery of biologics to back of the eye. Expert Opin. Drug Del. 2017, 14, 631–645. [Google Scholar] [CrossRef] [PubMed]
- Cholkar, K.; Gilger, B.C.; Mitra, A.K. Topical delivery of aqueous micellar resolvin E1 analog (RX-10045). Int. J. Pharm. 2016, 498, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Cholkar, K.; Gilger, B.C.; Mitra, A.K. Topical, aqueous, clear cyclosporine formulation design for anterior and posterior ocular delivery. Transl. Vis. Sci. Techn. 2015, 4, 1. [Google Scholar] [CrossRef] [PubMed]
- Vadlapudi, A.D.; Cholkar, K.; Vadlapatla, R.K.; Mitra, A.K. Aqueous nanomicellar formulation for topical delivery of biotinylated lipid prodrug of acyclovir: Formulation development and ocular biocompatibility. J. Ocul. Pharmacol. Ther. 2014, 30, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Earla, R.; Cholkar, K.; Gunda, S.; Earla, R.L.; Mitra, A.K. Bioanalytical method validation of rapamycin in ocular matrix by QTRAP LC–MS/MS: Application to rabbit anterior tissue distribution by topical administration of rapamycin nanomicellar formulation. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2012, 908, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Cholkar, K.; Gunda, S.; Earla, R.; Pal, D.; Mitra, A.K. Nanomicellar topical aqueous drop formulation of rapamycin for back-of-the-eye delivery. AAPS Pharm. Sci. Tech. 2015, 16, 610–622. [Google Scholar] [CrossRef] [PubMed]
- Davis, B.M.; Normando, E.M.; Guo, L.; Turner, L.A.; Nizari, S.; O’Shea, P.; Moss, S.E.; Somavarapu, S.; Cordeiro, M.F. Topical delivery of avastin to the posterior segment of the eye in vivo using annexin A5-associated liposomes. Small 2014, 10, 1575–1584. [Google Scholar] [CrossRef] [PubMed]
- Giambanco, I.; Pula, G.; Ceccarelli, P.; Bianchi, R.; Donato, R. Immunohistochemical localization of annexin V (CaBP33) in rat organs. J. Histochem. Cytochem. 1991, 39, 1189–1198. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, M.F.; Migdal, C.; Bloom, P.; Fitzke, F.W.; Moss, S.E. Imaging apoptosis in the eye. Eye 2011, 25, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Concha, N.O.; Head, J.F.; Kaetzel, M.A.; Dedman, J.R.; Seaton, B.A. Annexin-V forms calcium-dependent trimeric units on phospholipid-vesicles. FEBS Lett. 1992, 314, 159–162. [Google Scholar] [CrossRef]
- Langen, R.; Isas, J.M.; Hubbell, W.L.; Haigler, H.T. A transmembrane form of annexin XII detected by site-directed spin labeling. Pro. Natl. Acad. Sci. USA 1998, 95, 14060–14065. [Google Scholar] [CrossRef]
- Kenis, H.; van Genderen, H.; Bennaghmouch, A.; Rinia, H.A.; Frederik, P.; Narula, J.; Hofstra, L.; Reutelingsperger, C.P.M. Cell surface-expressed phosphatidylserine and annexin A5 open a novel portal of cell entry. J. Biol. Chem. 2004, 279, 52623–52629. [Google Scholar] [CrossRef] [PubMed]
- Zambrano, R.; Jamroz, M.; Szczasiuk, A.; Pujols, J.; Kmiecik, S.; Ventura, S. AGGRESCAN3D (A3D): Server for prediction of aggregation properties of protein structures. Nucleic Acids Res. 2015, 43, W306–313. [Google Scholar] [CrossRef] [PubMed]
- Derynck, R.; Zhang, Y.E. Smad-dependent and smad-independent pathways in TGF-β family signalling. Nature 2003, 425, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Caraci, F.; Battaglia, G.; Busceti, C.; Biagioni, F.; Mastrolacovo, F.; Bosco, P.; Drago, F.; Nicoletti, F.; Sortino, M.A.; Copani, A. TGF-β 1 protects against a β-neurotoxicity via the phosphatidylinositol-3-kinase pathway. Neurobiol. Dis. 2008, 30, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Walshe, T.E.; Leach, L.L.; D’Amore, P.A. TGF-β signaling is required for maintenance of retinal ganglion cell differentiation and survival. Neuroscience 2011, 189, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Garcia, C.M.; Darland, D.C.; Massingham, L.J.; D’Amore, P.A. Endothelial cell-astrocyte interactions and TGF β are required for induction of blood-neural barrier properties. Dev. Brain Res. 2004, 152, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Langen, R.; Isas, J.M.; Luecke, H.; Haigler, H.T.; Hubbell, W.L. Membrane-mediated assembly of annexins studied by site-directed spin labeling. J. Biol. Chem. 1998, 273, 22453–22457. [Google Scholar] [CrossRef] [PubMed]
- Pignatello, R. (Ed.) Drug-Biomembrane Interaction Studies, the Application of Calorimetric Techniques, 1st ed.; Woodhead Publishing: Sawaton, UK, 2013. [Google Scholar]
- Bevalot, F.; Cartiser, N.; Bottinelli, C.; Fanton, L.; Guitton, J. Vitreous humor analysis for the detection of xenobiotics in forensic toxicology: A review. Forensic Toxicol. 2016, 34, 12–40. [Google Scholar] [PubMed]
- Saleh, M.; Jehl, F.; Dory, A.; Lefevre, S.; Prevost, G.; Gaucher, D.; Sauer, A.; Speeg-Schatz, C.; Bourcier, T. Ocular penetration of topically applied linezolid in a rabbit model. J. Cataract. Refract. Surg. 2010, 36, 488–492. [Google Scholar] [CrossRef] [PubMed]
- Tuuminen, R.; Loukovaara, S. High intravitreal TGF-β 1 and MMP-9 levels in eyes with retinal vein occlusion. Eye 2014, 28, 1095–1099. [Google Scholar] [CrossRef] [PubMed]
- Gunda, S.; Earla, R.; Cholkar, K.; Mitra, A.K. Pharmacokinetic studies and LC–MS/MS method development of ganciclovir and dipeptide monoester prodrugs in sprague dawley rats. Eur. J. Drug Metab. Pharmacokinet. 2015, 40, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Giannavola, C.; Bucolo, C.; Maltese, A.; Paolino, D.; Vandelli, M.A.; Puglisi, G.; Lee, V.H.; Fresta, M. Influence of preparation conditions on acyclovir-loaded poly-d,l-lactic acid nanospheres and effect of PEG coating on ocular drug bioavailability. Pharm. Res. 2003, 20, 584–590. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Formulation | λz | * t½ (h) | Cmax (pg/mL) | Tmax (min) | AUC0–240min (pg × min/mL) |
|---|---|---|---|---|---|
| SUV-TGF-β1 | 0.90 | 0.77 | 114.7 ± 12.4 | 60 | 11,331 ± 894 |
| Formulation | Conjunctiva | Iris Hyperemia | Corneal Opacity | ||
|---|---|---|---|---|---|
| Congestion | Swelling | Discharge | |||
| SUV unloaded (10 min) | 0 | 0 | 0 | 0 | 0 |
| SUV unloaded (6 h) | 0 | 0 | 0 | 0 | 0 |
| SUV-TGF-β1 (10 min) | 0 | 0 | 0 | 0 | 0 |
| SUV-TGF-β1 (6 h) | 0 | 0 | 0 | 0 | 0 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Platania, C.B.M.; Fisichella, V.; Fidilio, A.; Geraci, F.; Lazzara, F.; Leggio, G.M.; Salomone, S.; Drago, F.; Pignatello, R.; Caraci, F.; et al. Topical Ocular Delivery of TGF-β1 to the Back of the Eye: Implications in Age-Related Neurodegenerative Diseases. Int. J. Mol. Sci. 2017, 18, 2076. https://doi.org/10.3390/ijms18102076
Platania CBM, Fisichella V, Fidilio A, Geraci F, Lazzara F, Leggio GM, Salomone S, Drago F, Pignatello R, Caraci F, et al. Topical Ocular Delivery of TGF-β1 to the Back of the Eye: Implications in Age-Related Neurodegenerative Diseases. International Journal of Molecular Sciences. 2017; 18(10):2076. https://doi.org/10.3390/ijms18102076
Chicago/Turabian StylePlatania, Chiara Bianca Maria, Vincenzo Fisichella, Annamaria Fidilio, Federica Geraci, Francesca Lazzara, Gian Marco Leggio, Salvatore Salomone, Filippo Drago, Rosario Pignatello, Filippo Caraci, and et al. 2017. "Topical Ocular Delivery of TGF-β1 to the Back of the Eye: Implications in Age-Related Neurodegenerative Diseases" International Journal of Molecular Sciences 18, no. 10: 2076. https://doi.org/10.3390/ijms18102076
APA StylePlatania, C. B. M., Fisichella, V., Fidilio, A., Geraci, F., Lazzara, F., Leggio, G. M., Salomone, S., Drago, F., Pignatello, R., Caraci, F., & Bucolo, C. (2017). Topical Ocular Delivery of TGF-β1 to the Back of the Eye: Implications in Age-Related Neurodegenerative Diseases. International Journal of Molecular Sciences, 18(10), 2076. https://doi.org/10.3390/ijms18102076