
Affinin (Spilanthol), Isolated from Heliopsis longipes, Induces Vasodilation via Activation of Gasotransmitters and Prostacyclin Signaling Pathways

 , ,
, ,  and
and
Abstract
:
1. Introduction
2. Results
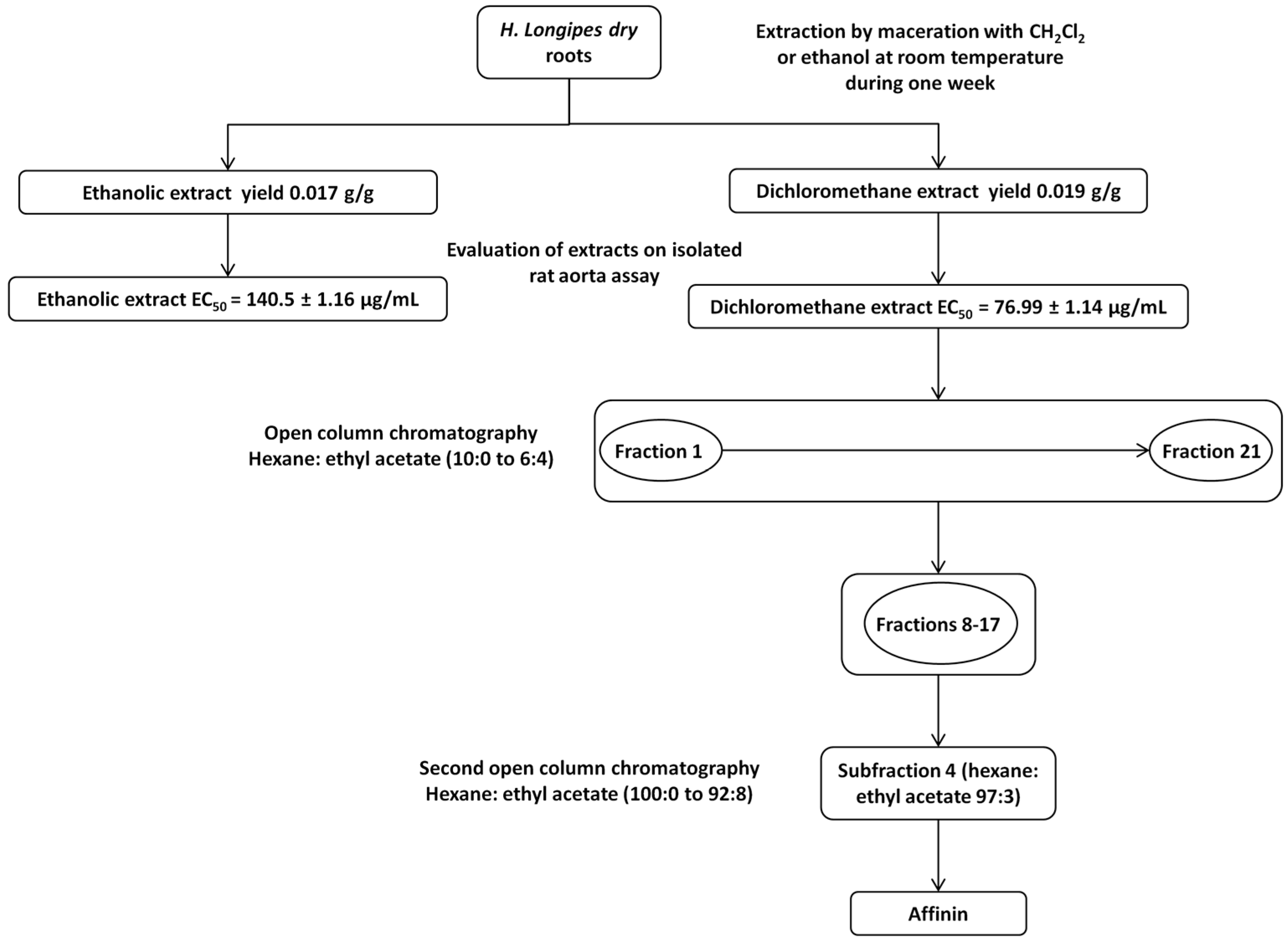
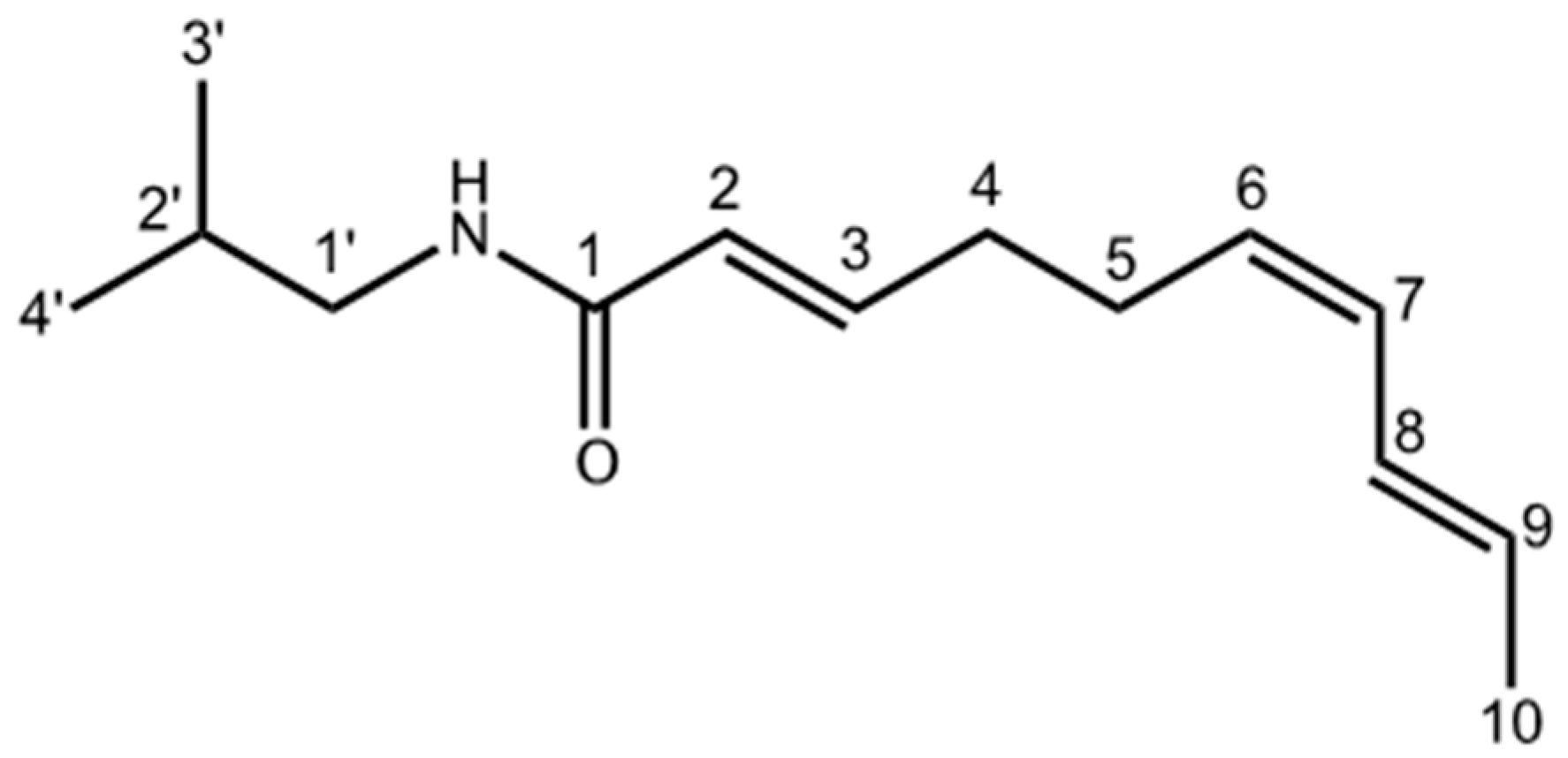
2.1. Phytochemical Study of the Dichloromethane Extract Obtained from H. longipes Roots and Isolation of Affinin
2.2. Determination of the Vasodilator Effect of H. longipes Extracts and Affinin, and Elucidation of the Mechanism of Action of Affinin
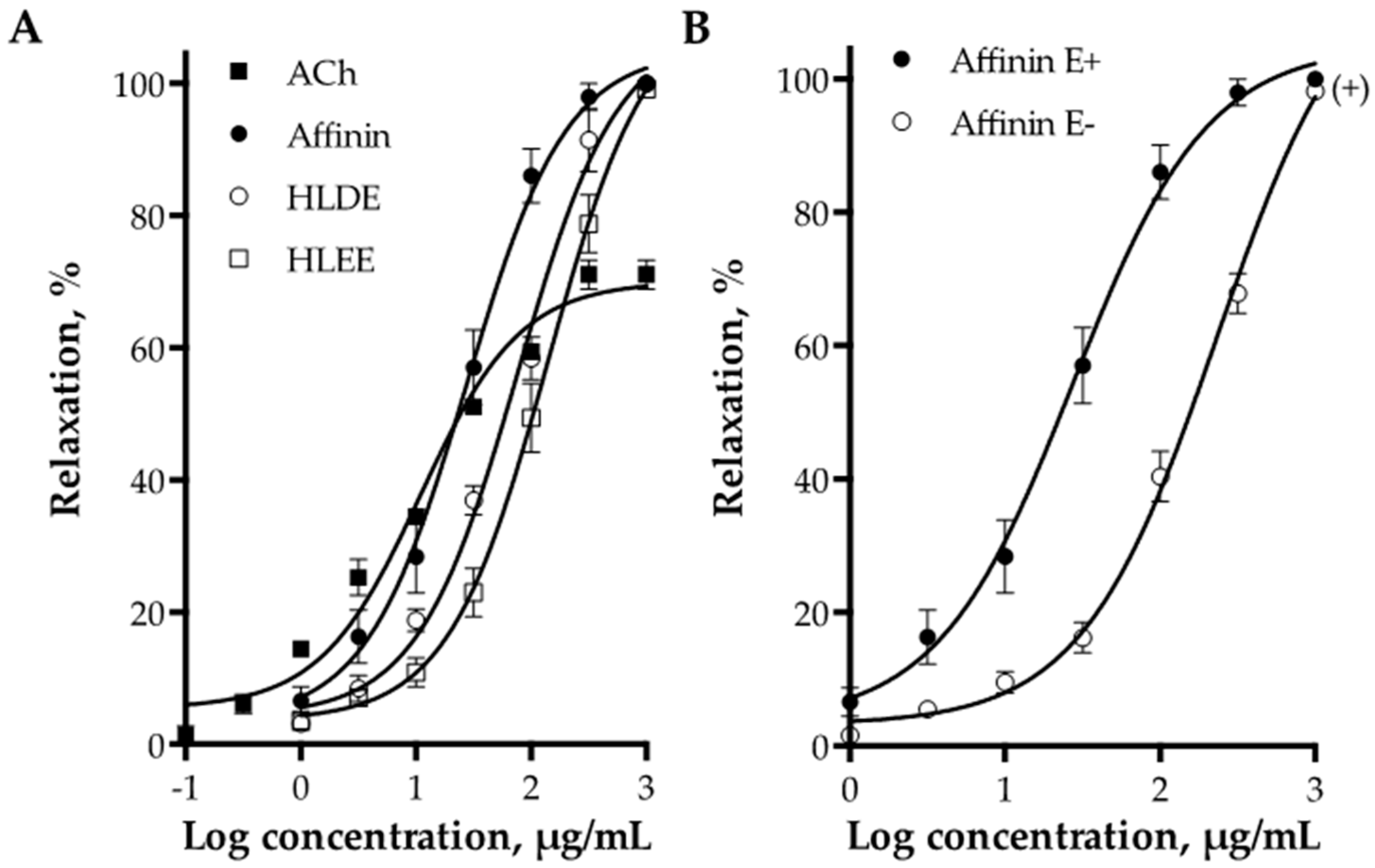
2.2.1. Vasodilator Effect of H. longipes Roots Extracts and Affinin
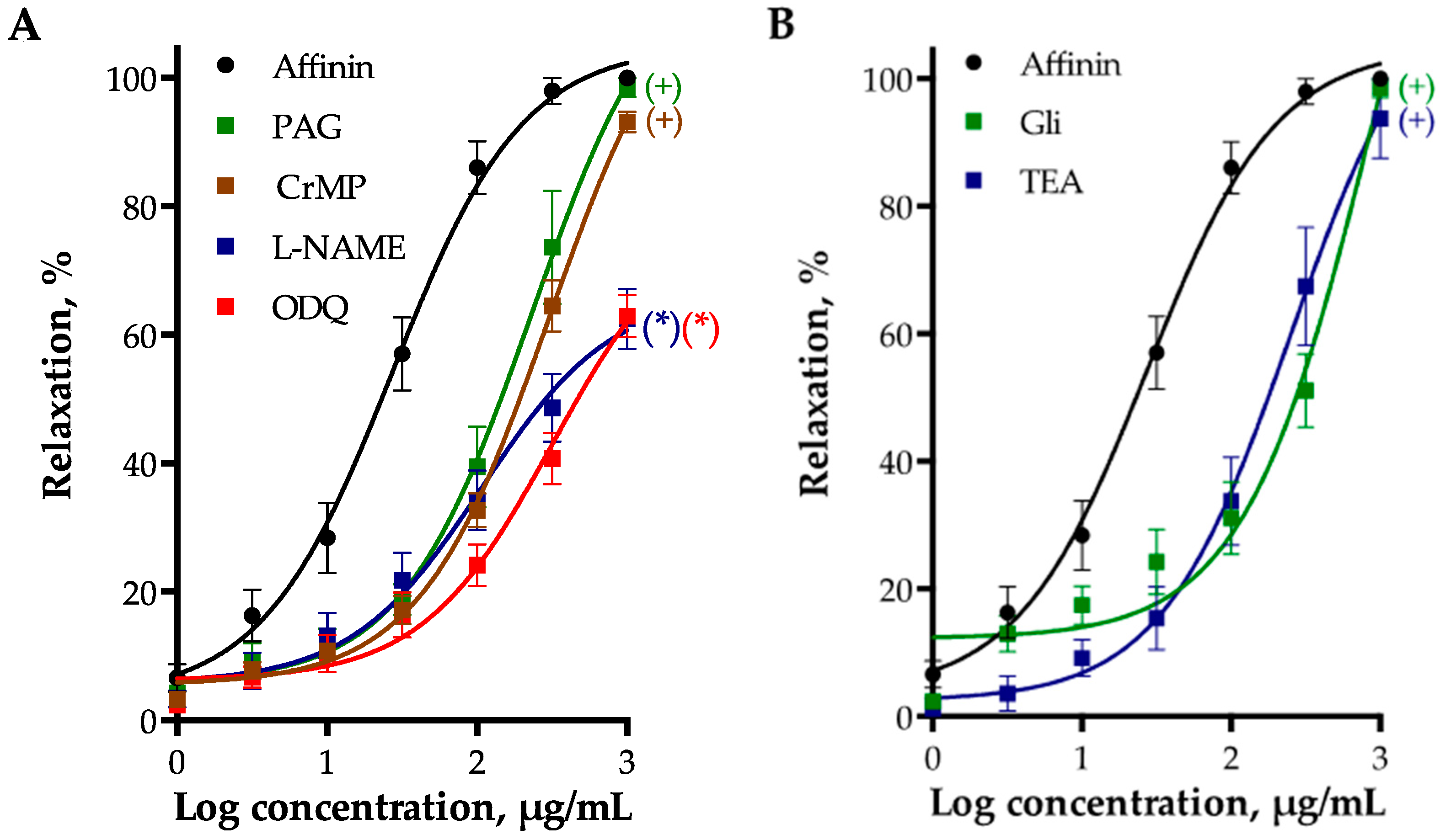
2.2.2. Role of Vascular Endothelium in the Vasodilation Induced by Affinin
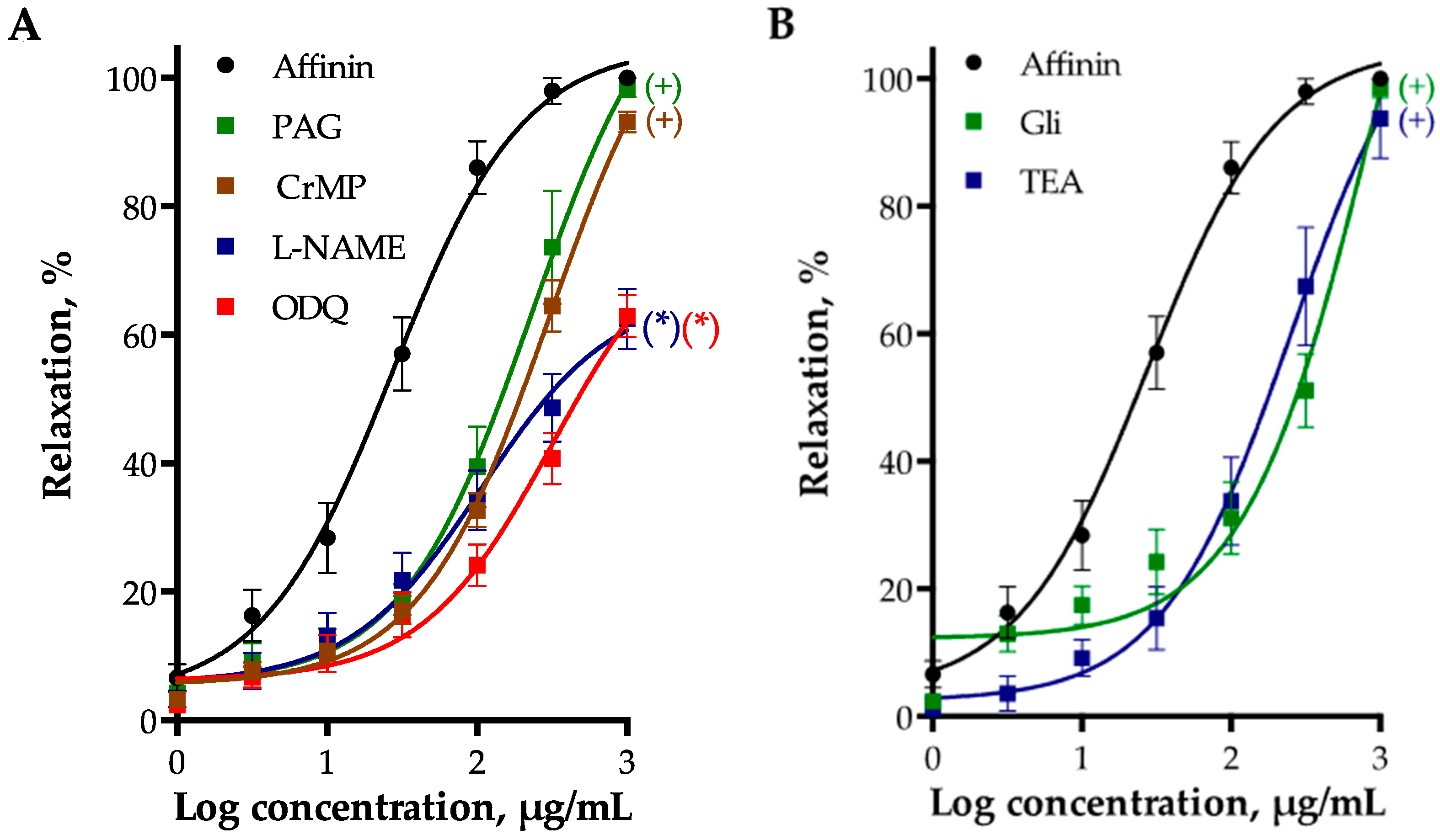
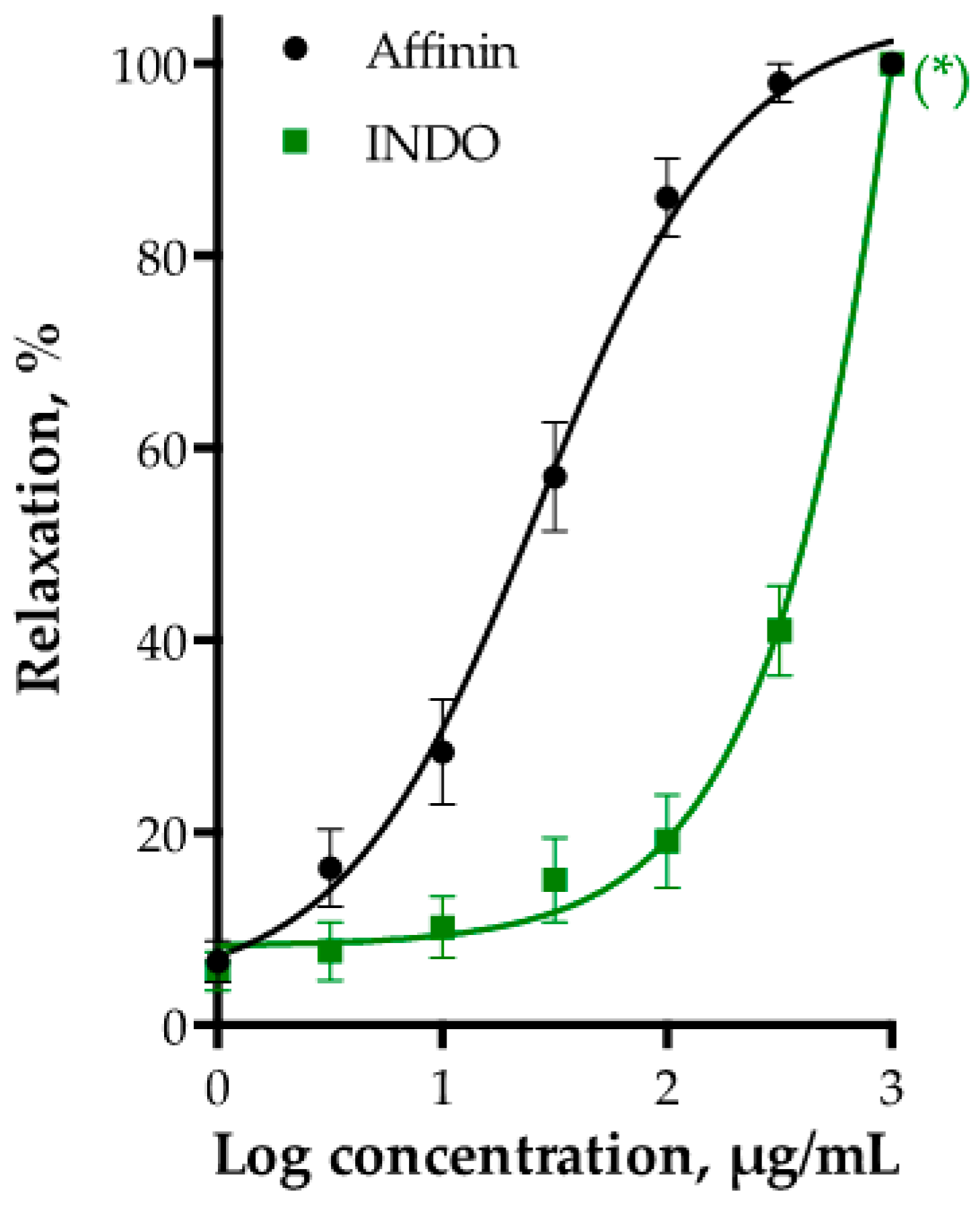
2.2.3. Involvement of Gasotransmitters in the Vasodilation Produced by Affinin
2.2.4. Involvement of K+ Channels in Affinin-Evoked Vasodilation
2.2.5. Effect of PGI2/cAMP Pathway on Affinin-Induced Dilation of Rat Aorta
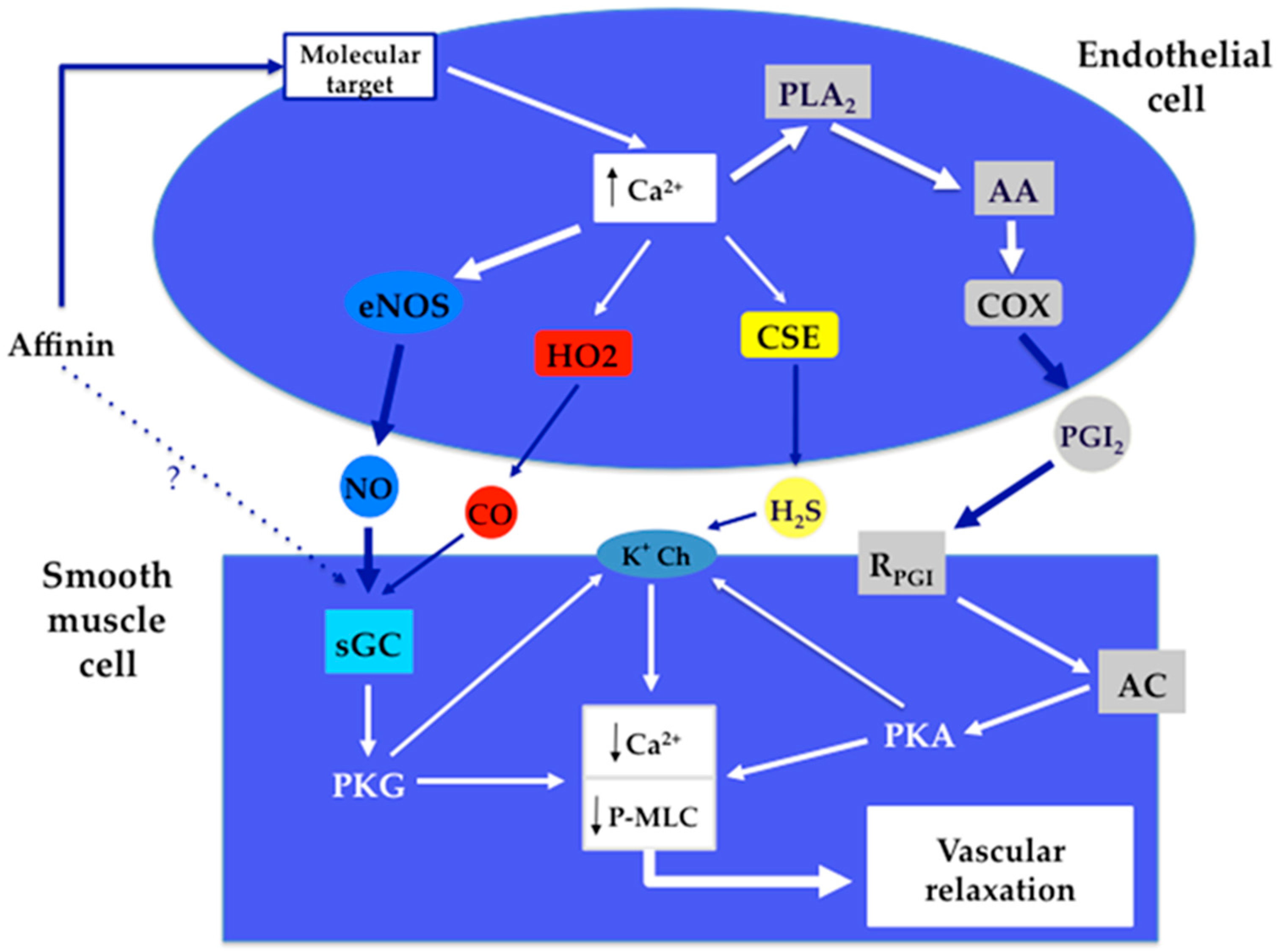
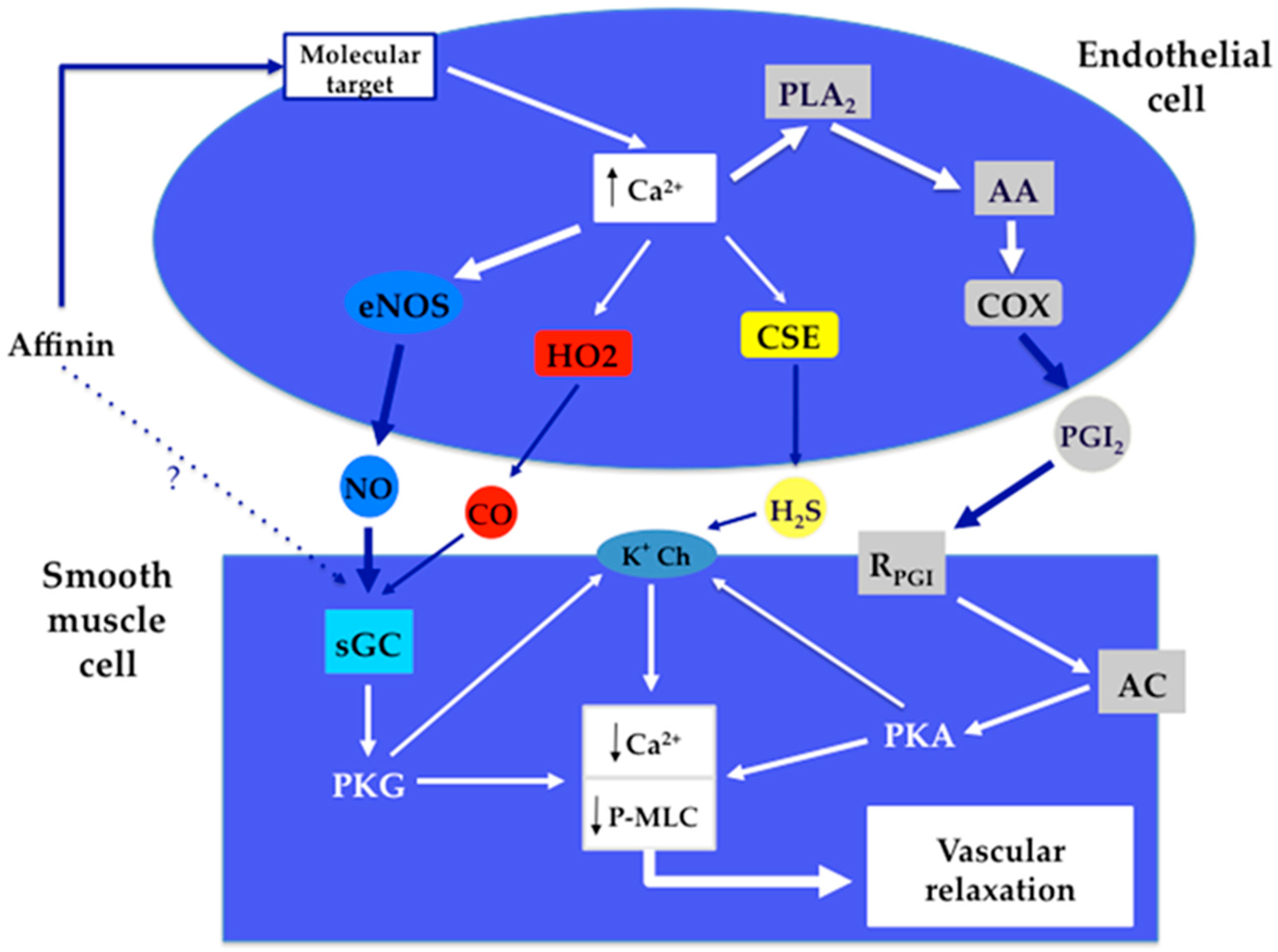
3. Discussion
4. Materials and Methods
4.1. Reagents and Chemicals
4.2. Animals
4.3. Plant Material
4.4. Preparation of the Extracts Employed for the Pharmacological Evaluation
4.5. Chemical Study of the Dichloromethane Extract Obtained from H. longipes Roots
4.5.1. Fractionation of the Dichloromethane Extract Obtained from H. longipes Roots and Purification of Affinin
4.5.2. Determination of the Chemical Structure of Affinin
4.6. Determination of the Vasodilator Effect and Elucidation of the Mechanism of Action of Affinin
4.6.1. Isolated Rat Aorta Assay
4.6.2. Participation of the Endothelium in the Vasodilator Response of Affinin
4.6.3. Evaluation of the Participation of the Gasotransmitters and Prostacyclin Signaling Pathways in the Vasodilator Response of Affinin
4.7. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflict of Interest
References
- Little, E.J. Heliopsis longipes, a Mexican insecticidal plant species. J. Wash. Acad. Sci. 1948, 38, 269–274. [Google Scholar] [PubMed]
- Correa, J.; Roquet, S.; Díaz, E. Multiple NMR analysis of the affinin. Org. Magn. Reson. 1971, 3, 1–5. [Google Scholar] [CrossRef]
- Martínez, M. Chilmecatl. In Las Plantas Medicinales de México; Ediciones Botas: México, D.F., Mexico, 1989. [Google Scholar]
- Molina-Torres, J.; García-Chávez, A.; Ramírez-Chávez, E. Antimicrobial properties of alkamides present in flavouring plants traditionally used in Mesoamerica: Affinin and capsaicin. J. Ethnopharmacol. 1999, 64, 241–248. [Google Scholar] [CrossRef]
- Cilia-López, V.G.; Juárez-Flores, B.I.; Aguirre-Rivera, J.R.; Reyes-Agüero, J.A. Analgesic activity of Heliopsis longipes and its effect on the nervous system. Pharm. Biol. 2010, 48, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Déciga-Campos, M.; Arriaga-Alba, M.; Ventura-Martínez, R.; Aguilar-Guadarrama, B.; Rios, M.Y. Pharmacological and Toxicological Profile of Extract from Heliopsis longipes and Affinin. Drug Dev. Res. 2012, 73, 130–137. [Google Scholar] [CrossRef]
- Acree, F.; Jacobson, M.J.; Haller, H.L. An amide posessing insecticidial properties from the roots of Erigeron affinins DC. J. Org. Chem. 1945, 236–242. [Google Scholar] [CrossRef]
- Molina-Torres, J.; Salazar-Cabrera, C.; Armenta-Salinas, C.; Ramírez-Chávez, E. Fungistatic and bacteriostatic activities of alkamides from Heliopsis longipes roots: Affinin and reduced amides. J. Agric. Food Chem. 2004, 52, 4700–4704. [Google Scholar] [CrossRef] [PubMed]
- Ogura, M.; Cordell, G.A.; Quinn, M.L.; Leon, C.; Benoit, P.S.; Soejarto, D.D.; Farnsworth, N.R. Ethnopharmacologic studies. I. Rapid solution to a problem—Oral use of Heliopsis longipes—By means of a multidisciplinary approach. J. Ethnopharmacol. 1982, 5, 215–219. [Google Scholar] [CrossRef]
- Arriaga-Alba, M.; Rios, M.Y.; Déciga-Campos, M. Antimutagenic properties of affinin isolated from Heliopsis longipes extract. Pharm. Biol. 2013, 51, 1035–1039. [Google Scholar] [CrossRef] [PubMed]
- Molina-Torres, J.; Salgado-Garciglia, R.; Ramírez-Chávez, E.; del Río, R.E. Purely Olefinic Alkamides in Heliopsis longipes and Acmella (Spilanthes) oppositifolia. Biochem. Syst. Ecol. 1996, 24, 43–47. [Google Scholar] [CrossRef]
- López-Martínez, S.; Aguilar-Guadarrama, B.; Ríos, M.Y. Minor alkamides from Heliopsis longipes S.F. Blake (Asteraceae) fresh roots. Phytochem. Lett. 2011, 4, 275–279. [Google Scholar] [CrossRef]
- Ríos, M.Y.; Olivo, H.F. Natural and Synthetic Alkamides: Applications in Pain Therapy. Stud. Nat. Prod. Chem. 2014, 43, 79–121. [Google Scholar]
- Greger, H. Alkamides: Structural relationships, distribution and biological activity. Planta Med. 1984, 50, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Greger, H. Alkamides: A critical reconsideration of a multifunctional class of unsaturated fatty acid amides. Phytochem. Rev. 2016, 15, 729–770. [Google Scholar] [CrossRef]
- Ríos, M.Y.; Aguilar-Guadarrama, A.B.; Gutiérrez, M.D.C. Analgesic activity of affinin, an alkamide from Heliopsis longipes (Compositae). J. Ethnopharmacol. 2007, 110, 364–367. [Google Scholar] [CrossRef] [PubMed]
- Johns, T.; Graham, K.; Towers, G.H.N. Molluscicidal activity of affinin and other isobutylamides from the asteraceae. Phytochemistry 1982, 21, 2737–2738. [Google Scholar] [CrossRef]
- Wu, L.C.; Fan, N.C.; Lin, M.H.; Chu, I.R.; Huang, S.J.; Hu, C.Y.; Han, S.Y. Anti-inflammatory effect of spilanthol from Spilanthes acmella on murine macrophage by down-regulating LPS-induced inflammatory mediators. J. Agric. Food Chem. 2008, 56, 2341–2349. [Google Scholar] [CrossRef] [PubMed]
- Boonen, J.; Baert, B.; Burvenich, C.; Blondeel, P.; de Saeger, S.; de Spiegeleer, B. LC-MS profiling of N-alkylamides in Spilanthes acmella extract and the transmucosal behaviour of its main bio-active spilanthol. J. Pharm. Biomed. Anal. 2010, 53, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, I.; Takeya, K.; Itokawa, H. The geometric structure of spilanthol. Chem. Pharm. Bull. 1980, 28, 2251–2253. [Google Scholar] [CrossRef]
- Nakatani, N.; Nagashima, M. Pungent Alkamides from Spilanthes acmella L. var. oleracea Clarke. Biosci. Biotechnol. Biochem. 1992, 56, 759–762. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Boonen, J.; Chauhan, N.S.; Thakur, M.; de Spiegeleer, B.; Dixit, V.K. Spilanthes acmella ethanolic flower extract: LC–MS alkylamide profiling and its effects on sexual behavior in male rats. Phytomedicine 2011, 18, 1161–1169. [Google Scholar] [CrossRef] [PubMed]
- Veryser, L.; Taevernier, L.; Joshi, T.; Tatke, P.; Wynendaele, E.; Bracke, N.; Stalmans, S.; Peremans, K.; Burvenich, C.; Risseeuw, M.; et al. Mucosal and blood-brain barrier transport kinetics of the plant N-alkylamide spilanthol using in vitro and in vivo models. BMC Complement. Altern. Med. 2016, 16, 177. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.S.; Ehrmann, B.M.; Ettefagh, K.A.; Cech, N.B. A validated liquid chromatography-electrospray ionization-mass spectrometry method for quantification of spilanthol in Spilanthes acmella (L.) Murr. Phytochem. Anal. 2010, 21, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Morales, A.; Arvizu-Gómez, J.L.; Carranza-Álvarez, C.; Gómez-Luna, B.E.; Alvarado-Sánchez, B.; Ramírez-Chávez, E.; Molina-Torres, J. Larvicidal activity of affinin and its derived amides from Heliopsis longipes A. Gray Blake against Anopheles albimanus and Aedes aegypti. J. Asia. Pac. Entomol. 2015, 18, 227–231. [Google Scholar] [CrossRef]
- Déciga-Campos, M.; Rios, M.Y.; Aguilar-Guadarrama, A.B. Antinociceptive effect of Heliopsis longipes extract and affinin in mice. Planta Med. 2010, 76, 665–670. [Google Scholar] [CrossRef] [PubMed]
- Gerbino, A.; Schena, G.; Milano, S.; Milella, L.; Franco Barbosa, A.; Armentano, F.; Procino, G.; Svelto, M.; Carmosino, M. Spilanthol from Acmella oleracea lowers the intracellular levels of cAMP impairing NKCC2 phosphorylation and water channel AQP2 membrane expression in mouse kidney. PLoS ONE 2016, 11, e0156021. [Google Scholar] [CrossRef] [PubMed]
- Cariño-Cortés, R.; Gayosso-De-Lucio, J.A.; Ortiz, M.I.; Sánchez-Gutiérrez, M.; García-Reyna, P.B.; Cilia-López, V.G.; Pérez-Hernández, N.; Moreno, E.; Ponce-Monter, H. Antinociceptive, genotoxic and histopathological study of Heliopsis longipes S.F. Blake in mice. J. Ethnopharmacol. 2010, 130, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Madrid, I.I.; Castañeda-Hernández, G.; Cilia-López, V.G.; Cariño-Cortés, R.; Pérez-Hernández, N.; Fernández-Martínez, E.; Ortiz, M.I. Interaction between Heliopsis longipes extract and diclofenac on the thermal hyperalgesia test. Phytomedicine 2009, 16, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Hernández, I.; Márquez, L.; Martínez, I.; Dieguez, R.; Delporte, C.; Prieto, S.; Molina-Torres, J.; Garrido, G. Anti-inflammatory effects of ethanolic extract and alkamides-derived from Heliopsis longipes roots. J. Ethnopharmacol. 2009, 124, 649–652. [Google Scholar] [CrossRef] [PubMed]
- Hernández, I.; Lemus, Y.; Prieto, S.; Molina-Torres, J.; Garrido, G. Anti-inflammatory effect of an ethanolic root extract of Heliopsis longipes in vitro. Boletín Latinoam. Caribe Plantas Med. Aromáticas 2009, 8, 160–164. [Google Scholar]
- Veryser, L.; Wynendaele, E.; Taevernier, L.; Verbeke, F.; Joshi, T.; Tatke, P.; de Spiegeleer, B. N-alkylamides: From plant to brain. Funct. Foods Heal. Dis. 2014, 4, 264–275. [Google Scholar]
- Boonen, J.; Baert, B.; Roche, N.; Burvenich, C.; de Spiegeleer, B. Transdermal behaviour of the N-alkylamide spilanthol (affinin) from Spilanthes acmella (Compositae) extracts. J. Ethnopharmacol. 2010, 127, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Coletta, C.; Papapetropoulos, A.; Erdelyi, K.; Olah, G.; Modis, K.; Panopoulos, P.; Asimakopoulou, A.; Gero, D.; Sharina, I.; Martin, E.; et al. Hydrogen sulfide and nitric oxide are mutually dependent in the regulation of angiogenesis and endothelium-dependent vasorelaxation. Proc. Natl. Acad. Sci. USA 2012, 109, 9161–9166. [Google Scholar] [CrossRef] [PubMed]
- Bohlen, H.G. Nitric oxide and the cardiovascular system. Compr. Physiol. 2015, 5, 808–823. [Google Scholar] [PubMed]
- Zhao, Y.; Vanhoutte, P.M.; Leung, S.W.S. Vascular nitric oxide: Beyond eNOS. J. Pharmacol. Sci. 2015, 129, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Derbyshire, E.R.; Marletta, M.A. Structure and Regulation of Soluble Guanylate Cyclase. Annu. Rev. Biochem. 2012, 81, 533–559. [Google Scholar] [CrossRef] [PubMed]
- White, R.E.; Kryman, J.P.; El-Mowafy, A.M.; Han, G.; Carrier, G.O. cAMP-dependent vasodilators cross-activate the cGMP-dependent protein kinase to stimulate BK(Ca) channel activity in coronary artery smooth muscle cells. Circ. Res. 2000, 86, 897–905. [Google Scholar] [CrossRef] [PubMed]
- Boerth, N.J.; Dey, N.B.; Cornwell, T.L.; Lincoln, T.M. Cyclic GMP-dependent protein kinase regulates vascular smooth muscle cell phenotype. J. Vasc. Res. 1997, 34, 245–259. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Sattar, M.A.; Rathore, H.A.; Khan, S.A.; Lazhari, M.I.; Afzal, S.; Hashmi, F.; Abdullah, N.A.; Johns, E.J. A critical review of pharmacological significance of Hydrogen Sulfide in hypertension. Indian J. Pharmacol. 2015, 47, 243–247. [Google Scholar] [PubMed]
- Holwerda, K.M.; Karumanchi, S.A.; Lely, A.T. Hydrogen sulfide: Role in vascular physiology and pathology. Curr. Opin. Nephrol. Hypertens. 2015, 24, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Bełtowski, J.; Jamroz-Wis̈niewska, A. Hydrogen sulfide and endothelium-dependent vasorelaxation. Molecules 2014, 19, 21506–21528. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Szabo, C.; Ichinose, F.; Ahmed, A.; Whiteman, M.; Papapetropoulos, A. The role of H2S bioavailability in endothelial dysfunction. Trends Pharmacol. Sci. 2015, 36, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Durante, W.; Johnson, F.K.; Johnson, R.A. Role of carbon monoxide in cardiovascular function. J. Cell. Mol. Med. 2006, 10, 672–686. [Google Scholar] [CrossRef] [PubMed]
- Durante, W. Carbon monoxide and bile pigments: Surprising mediators of vascular function. Vasc. Med. 2002, 7, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Purohit, R.; Fritz, B.G.; The, J.; Issaian, A.; Weichsel, A.; David, C.L.; Campbell, E.; Hausrath, A.C.; Rassouli-Taylor, L.; Garcin, E.D.; et al. YC-1 binding to the β subunit of soluble guanylyl cyclase overcomes allosteric inhibition by the α subunit. Biochemistry 2014, 53, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Huang, H.; Liu, P.; Tang, C.; Wang, J. Hydrogen sulfide contributes to cardioprotection during ischemia-reperfusion injury by opening K ATP channels. Can. J. Physiol. Pharmacol. 2007, 85, 1248–1253. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Zhang, J.; Lu, Y.; Wang, R. The vasorelaxant effect of H2S as a novel endogenous gaseous KATP channel opener. EMBO J. 2001, 20, 6008–6016. [Google Scholar] [CrossRef] [PubMed]
- Yoo, D.; Jupiter, R.C.; Pankey, E.A.; Reddy, V.G.; Edward, J.A.; Swan, K.W.; Peak, T.C.; Mostany, R.; Kadowitz, P.J. Analysis of Cardiovascular Responses to the H2S donors Na2S and NaHS in the Rat. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H605–H614. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, A.K.; Gadalla, M.M.; Snyder, S.H. Signaling by gasotransmitters. Sci. Signal. 2009, 2, re2. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Wu, L.; Wang, Z. The direct effect of carbon monoxide on KCa channels in vascular smooth muscle cells. Pflug. Arch. 1997, 434, 285–291. [Google Scholar] [CrossRef]
- Decaluwé, K.; Pauwels, B.; Verpoest, S.; van de Voorde, J. Divergent mechanisms involved in CO and CORM-2 induced vasorelaxation. Eur. J. Pharmacol. 2012, 674, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Wang, R. Gasotransmitters: Growing pains and joys. Trends Biochem. Sci. 2014, 39, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Ibarra-Alvarado, C.; Galle, J.; Melichar, V.O.; Mameghani, A.; Schmidt, H.H. Phosphorylation of blood vessel vasodilator-stimulated phosphoprotein at serine 239 as a functional biochemical marker of endothelial nitric oxide/cyclic GMP signaling. Mol. Pharmacol. 2002, 61, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Stoner, L.; Erickson, M.L.; Young, J.M.; Fryer, S.; Sabatier, M.J.; Faulkner, J.; Lambrick, D.M.; McCully, K.K. There’s more to flow-mediated dilation than nitric oxide. J. Atheroscler. Thromb. 2012, 19, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Giles, T.D.; Sander, G.E.; Nossaman, B.D.; Kadowitz, P.J. Impaired Vasodilation in the Pathogenesis of Hypertension: Focus on Nitric Oxide, Endothelial-Derived Hyperpolarizing Factors, and Prostaglandins. J. Clin. Hypertens. 2012, 14, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Kawabe, J.; Ushikubi, F.; Hasebe, N. Prostacyclin in Vascular Diseases. Circ. J. 2010, 74, 836–843. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, M.I.; Cariño-Cortés, R.; Pérez-Hernández, N.; Ponce-Monter, H.; Fernández-Martínez, E.; Castañeda-Hernández, G.; Acosta-Madrid, I.I.; Cilia-López, V.G. Antihyperalgesia induced by Heliopsis longipes extract. Proc. West. Pharmacol. Soc. 2009, 52, 75–77. [Google Scholar] [PubMed]
- Wongsawatkul, O.; Prachayasittikul, S.; Isarankura-Na-Ayudhya, C.; Satayavivad, J.; Ruchirawat, S.; Prachayasittikul, V. Vasorelaxant and antioxidant activities of Spilanthes acmella Murr. Int. J. Mol. Sci. 2008, 9, 2724–2744. [Google Scholar] [CrossRef] [PubMed]
- Yetik-Anacak, G.; Sorrentino, R.; Linder, A.E.; Murat, N. Gas what: NO is not the only answer to sexual function. Br. J. Pharmacol. 2015, 172, 1434–1454. [Google Scholar] [CrossRef] [PubMed]
- Nangle, M.R.; Cotter, M.A.; Cameron, N.E. An in vitro study of corpus cavernosum and aorta from mice lacking the inducible nitric oxide synthase gene. Nitric Oxide 2003, 9, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Dalaklioglu, S.; Ozbey, G. The potent relaxant effect of resveratrol in rat corpus cavernosum and its underlying mechanisms. Int. J. Impot. Res. 2013, 25, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Boonen, J.; Bronselaer, A.; Nielandt, J.; Veryser, L.; de Tré, G.; de Spiegeleer, B. Alkamid database: Chemistry, occurrence and functionality of plant N-alkylamides. J. Ethnopharmacol. 2012, 142, 563–590. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Loredo, E.; Izquierdo-Vega, J.A.; Cariño-Cortés, R.; Cilia-López, V.G.; Madrigal-Santillán, E.O.; Zuñiga-Pérez, C.; Valadez-Vega, C.; Moreno, E.; Sánchez-Gutiérrez, M. Effects of Heliopsis longipes ethanolic extract on mouse spermatozoa in vitro. Pharm. Biol. 2016, 54, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Chicca, A.; Raduner, S.; Pellati, F.; Strompen, T.; Altmann, K.-H.; Schoop, R.; Gertsch, J. Synergistic immunomopharmacological effects of N-alkylamides in Echinacea purpurea herbal extracts. Int. Immunopharmacol. 2009, 9, 850–858. [Google Scholar] [CrossRef] [PubMed]
- Sudhahar, V.; Shaw, S.; Imig, J.D. Mechanisms involved in oleamide-induced vasorelaxation in rat mesenteric resistance arteries. Eur. J. Pharmacol. 2009, 607, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Raboune, S.; Stuart, J.M.; Leishman, E.; Takacs, S.M.; Rhodes, B.; Basnet, A.; Jameyfield, E.; McHugh, D.; Widlanski, T.; Bradshaw, H.B. Novel endogenous N-acyl amides activate TRPV1–4 receptors, BV-2 microglia, and are regulated in brain in an acute model of inflammation. Front. Cell. Neurosci. 2014, 8, 195. [Google Scholar] [CrossRef] [PubMed]
- Raduner, S.; Majewska, A.; Chen, J.; Xie, X.; Faller, B.; Altmann, K.; Hamon, J. Alkylamides from Echinacea Are a New Class of Cannabinomimetics. J. Biol. Chem. 2006, 281, 14192–14206. [Google Scholar] [CrossRef] [PubMed]
- Rios, M. Natural Alkamides: Pharmacology, Chemistry and Distribution. In Drug Discovery Research in Pharmacognosy; InTech: Vienna, Austria, 2013; pp. 107–144. [Google Scholar]
- Lu, H.C.; MacKie, K. An introduction to the endogenous cannabinoid system. Biol. Psychiatry 2016, 79, 516–525. [Google Scholar] [CrossRef] [PubMed]
- Zygmunt, P.M.; Petersson, J.; Andersson, D.A.; Chuang, H.; Sørgård, M.; Di Marzo, V.; Julius, D.; Högestätt, E.D. Vanilloid receptors on sensory nerves mediate the vasodilator action of anandamide. Nature 1999, 400, 452–457. [Google Scholar] [PubMed]
- Herradón, E.; Martín, M.I.; López-Miranda, V. Characterization of the vasorelaxant mechanisms of the endocannabinoid anandamide in rat aorta. Br. J. Pharmacol. 2007, 152, 699–708. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, S.E.; Kendall, D.A.; Randall, M.D. Vascular effects of Δ9-tetrahydrocannabinol (THC), anandamide and N-arachidonoyldopamine (NADA) in the rat isolated aorta. Eur. J. Pharmacol. 2005, 507, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Norma Oficial Mexicana, NOM-062-ZOO-1999, Especificaciones Técnicas Para la Producción, Cuidado y uso de los Animales de Laboratorio. Available online: http://www.fmvz.unam.mx/fmvz/principal/archivos/062ZOO.PDF (accessed on 19 January 2017).
- Bauer, R.; Remiger, P. TLC and HPLC Analysis of Alkamides in Echinacea Drugs1,2. Planta Med. 1989, 55, 367–371. [Google Scholar] [CrossRef] [PubMed]
- Ibarra-Alvarado, C.; Rojas, A.; Mendoza, S.; Bah, M.; Gutiérrez, D.M.; Hernández-Sandoval, L.; Martínez, M. Vasoactive and antioxidant activities of plants used in Mexican traditional medicine for the treatment of cardiovascular diseases. Pharm. Biol. 2010, 48, 732–739. [Google Scholar] [CrossRef] [PubMed]
- Andresen, J.J.; Shafi, N.I.; Durante, W.; Bryan, R.M. Effects of carbon monoxide and heme oxygenase inhibitors in cerebral vessels of rats and mice. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, H223–H230. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, C.; Rosas-Hernandez, H.; Jurado-manzano, B.; Ramirez-Lee, M.A.; Salazar-Garcia, S.; Martinez-Cuevas, P.P.; Velarde-salcedo, A.J.; Morales-Loredo, H.; Espinosa-Tanguma, R.; Ali, S.F.; et al. The prolactin family hormones regulate vascular tone through NO and prostacyclin production in isolated rat aortic rings. Acta Pharmacol. Sin. 2015, 36, 572–586. [Google Scholar] [CrossRef] [PubMed]
- Majed, B.H.; Khalil, R.A. Molecular mechanisms regulating the vascular prostacyclin pathways and their adaptation during pregnancy and in the newborn. Pharmacol. Rev. 2012, 64, 540–582. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Zhao, L.; Wu, R.; Yue, S.; Pei, J. The vasorelaxing effect of resveratrol on abdominal aorta from rats and its underlying mechanisms. Vasc. Pharmacol. 2013, 58, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Stott, J.B.; Jepps, T.A.; Greenwood, I.A. KV7 potassium channels: A new therapeutic target in smooth muscle disorders. Drug Discov. Today 2014, 19, 413–424. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| H | δppm | C |
|---|---|---|
| 1 | - | 166.15 |
| 2 | 5.80 (1H, br d, J = 16.0, 8.0 Hz) | 124.30 |
| 3 | 6.80 (1H, dt, J = 16.0, 8.0 Hz) | 143.51 |
| 4 | 2.28 (4H, m) | 32.20 |
| 5 | 2.28 (4H, m) | 26.49 |
| 6 | 5.25 (1H, dt, J = 10.7, 7.1 Hz) | 127.73 |
| 7 | 5.94 (1H, dd, J = 12.0 Hz) | 129.52 |
| 8 | 6.25 (1H, br dd, J = 16.0, 4.0 Hz) | 126.79 |
| 9 | 5.67 (1H, dq, J = 16.0, 6.0 Hz) | 130.00 |
| 10 | 1.76 (3H, d, J = 6.0 Hz) | 18.39 |
| NH | 5.47 (br s) | - |
| 1′ | 3.13 (2H, dd, J = 6.0, 6.0 Hz) | 46.97 |
| 2′ | 1.80 (1H, m) | 28.68 |
| 3′ | 0.93 (6H, d, J = 6.7 Hz) | 20.23 |
| 4′ | 0.93 (6H, d, J = 6.7 Hz) | 18.40 |
| Compound | Emax (%) | EC50 (µg/mL) |
|---|---|---|
| Dichloromethane extract | 100 ± 3.11 | 76.99 ± 1.14 |
| Ethanolic extract | 100 ± 4.54 | 140.5 ± 1.16 |
| Affinin | 100 ± 3.10 | 27.38 ± 1.20 |
| ACh | 70.02 ± 1.43 | 1.094 ± 1.14 |
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castro-Ruiz, J.E.; Rojas-Molina, A.; Luna-Vázquez, F.J.; Rivero-Cruz, F.; García-Gasca, T.; Ibarra-Alvarado, C. Affinin (Spilanthol), Isolated from Heliopsis longipes, Induces Vasodilation via Activation of Gasotransmitters and Prostacyclin Signaling Pathways. Int. J. Mol. Sci. 2017, 18, 218. https://doi.org/10.3390/ijms18010218
Castro-Ruiz JE, Rojas-Molina A, Luna-Vázquez FJ, Rivero-Cruz F, García-Gasca T, Ibarra-Alvarado C. Affinin (Spilanthol), Isolated from Heliopsis longipes, Induces Vasodilation via Activation of Gasotransmitters and Prostacyclin Signaling Pathways. International Journal of Molecular Sciences. 2017; 18(1):218. https://doi.org/10.3390/ijms18010218
Chicago/Turabian StyleCastro-Ruiz, Jesús Eduardo, Alejandra Rojas-Molina, Francisco J. Luna-Vázquez, Fausto Rivero-Cruz, Teresa García-Gasca, and César Ibarra-Alvarado. 2017. "Affinin (Spilanthol), Isolated from Heliopsis longipes, Induces Vasodilation via Activation of Gasotransmitters and Prostacyclin Signaling Pathways" International Journal of Molecular Sciences 18, no. 1: 218. https://doi.org/10.3390/ijms18010218
APA StyleCastro-Ruiz, J. E., Rojas-Molina, A., Luna-Vázquez, F. J., Rivero-Cruz, F., García-Gasca, T., & Ibarra-Alvarado, C. (2017). Affinin (Spilanthol), Isolated from Heliopsis longipes, Induces Vasodilation via Activation of Gasotransmitters and Prostacyclin Signaling Pathways. International Journal of Molecular Sciences, 18(1), 218. https://doi.org/10.3390/ijms18010218