
Zinc as a Signal to Stimulate Red Blood Cell Formation in Fish


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
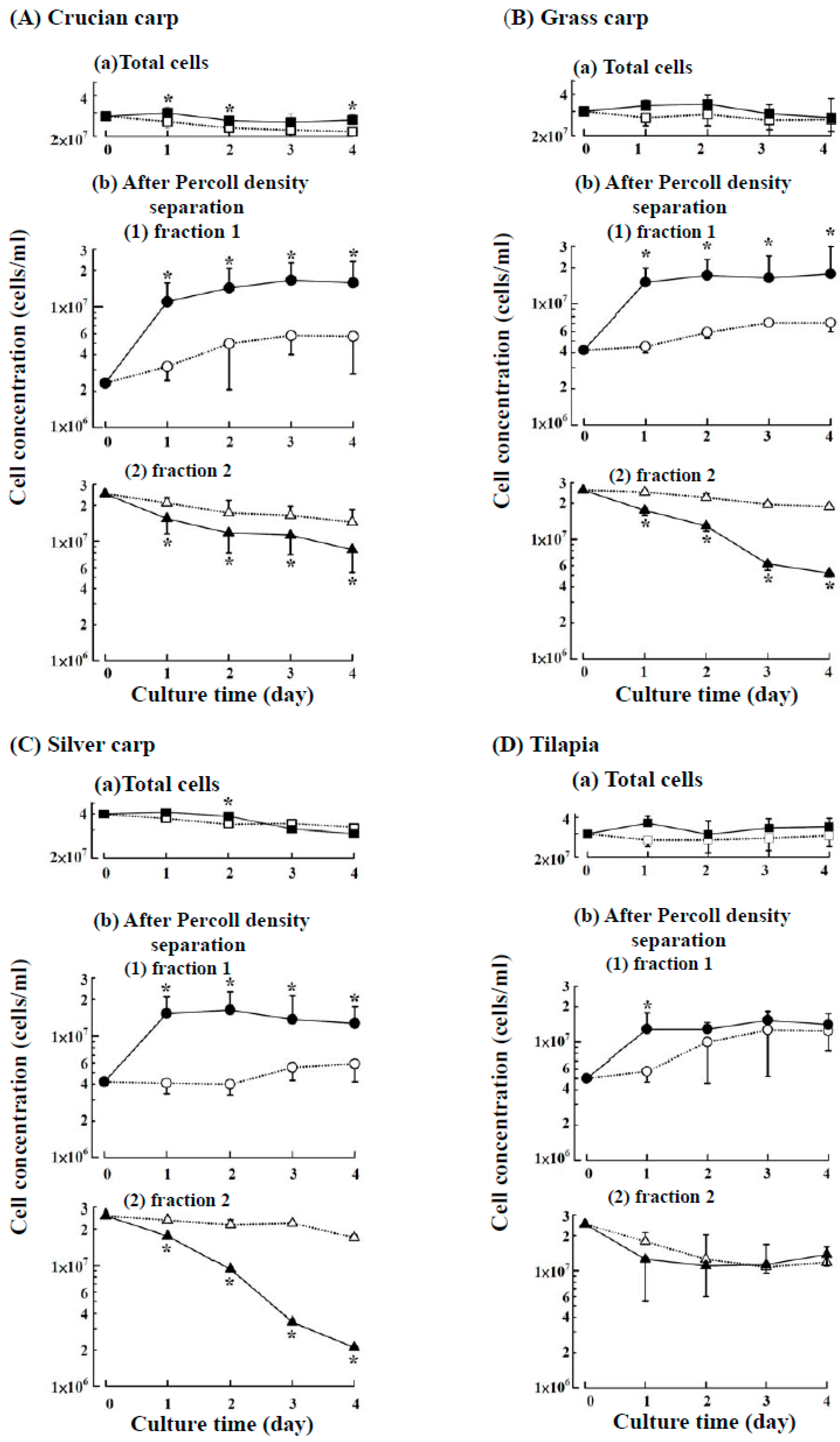
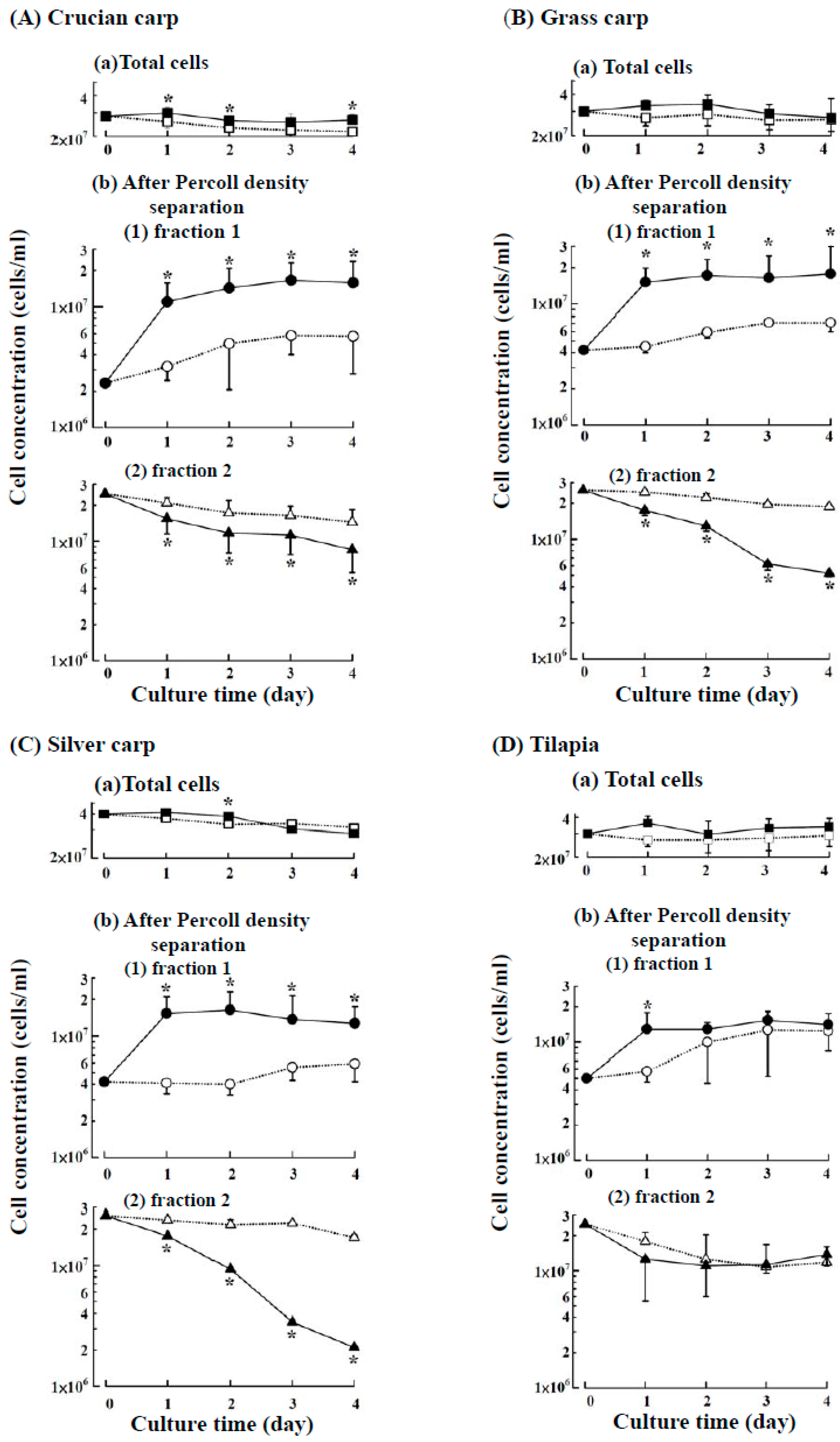
2.1. Suspension Culture of the Head Kidney Cells from Four Fish Species with or without ZnCl2
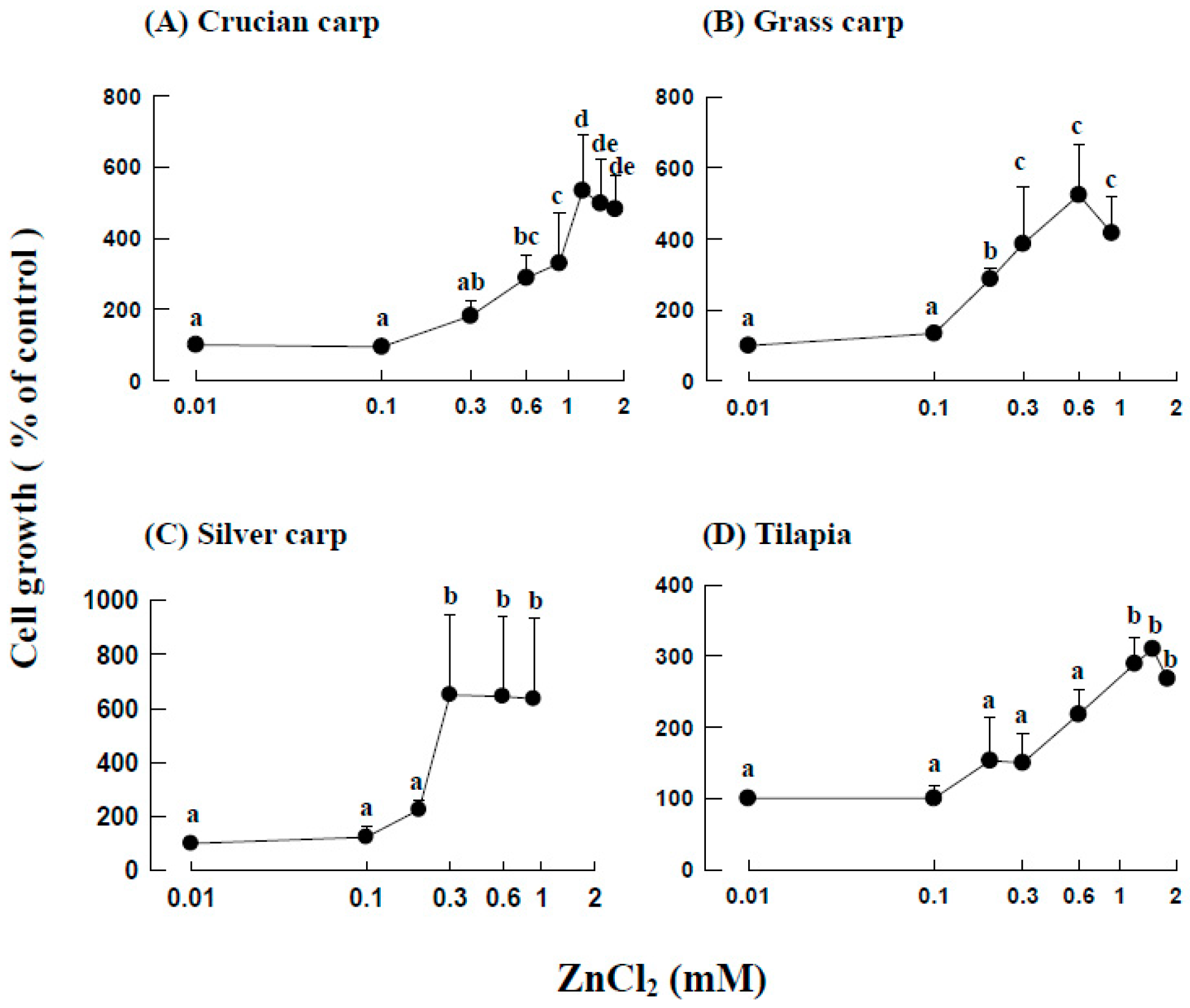
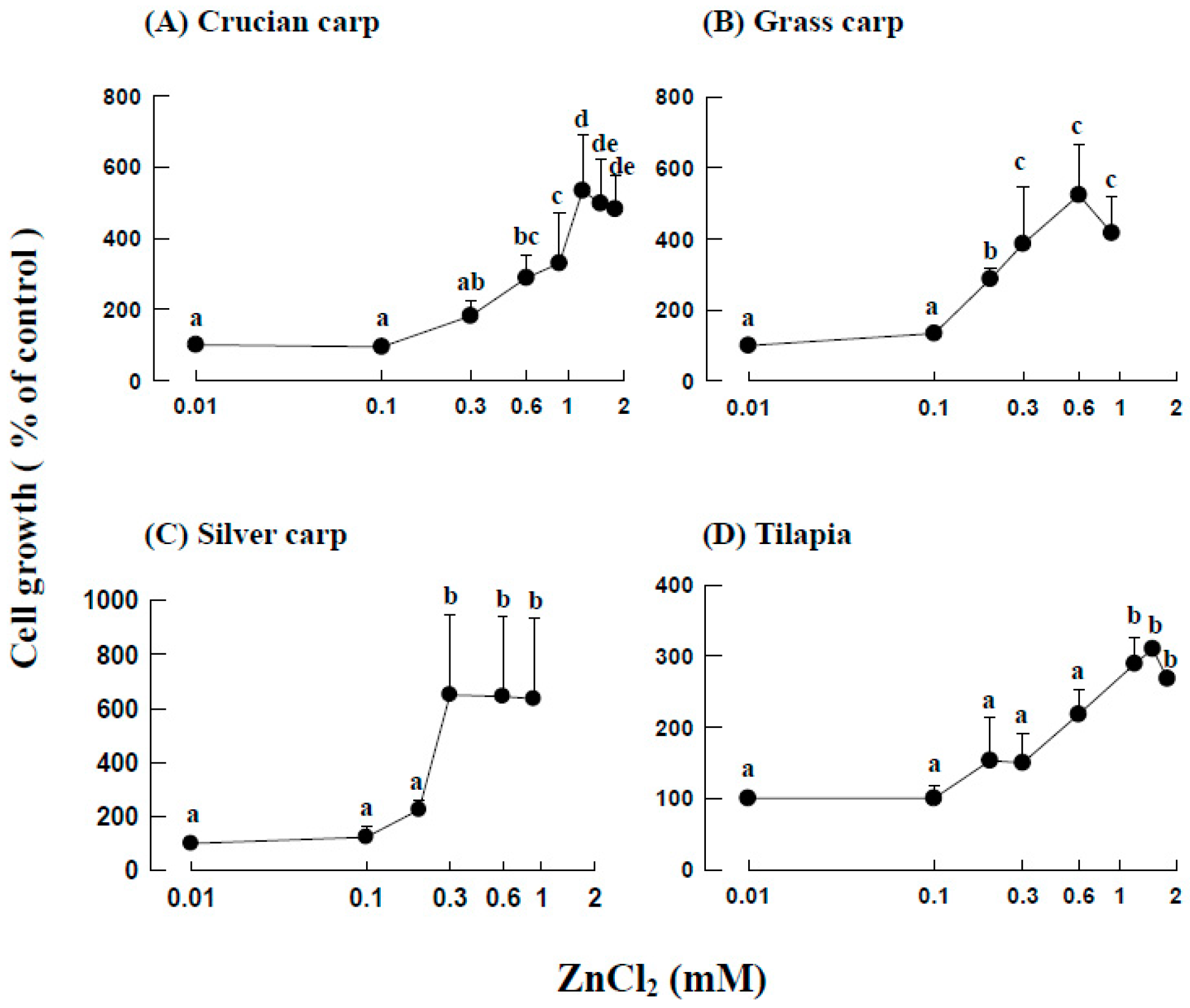
2.2. Effects of ZnCl2 Levels on the Growth of Fraction 1 Cells
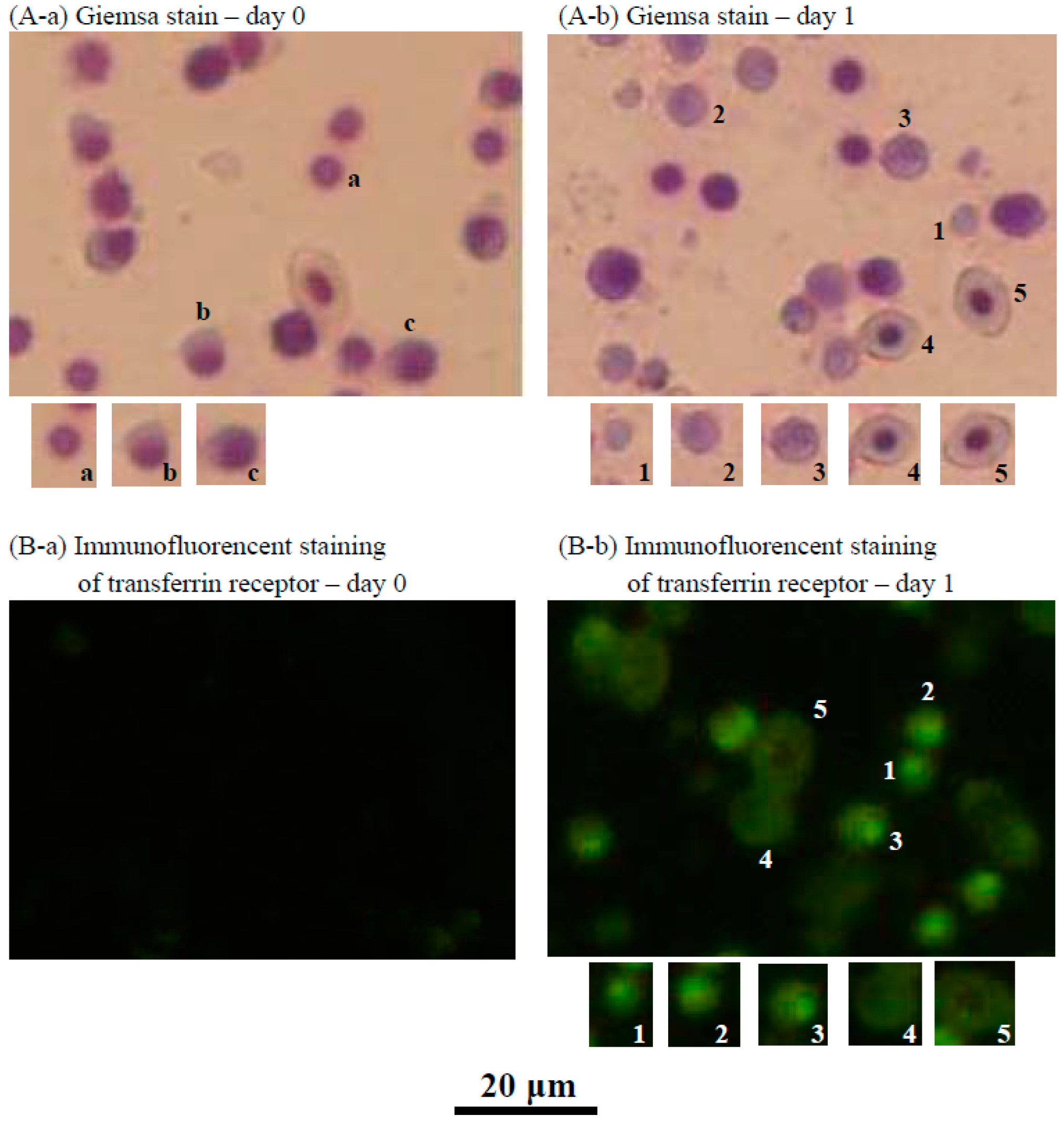
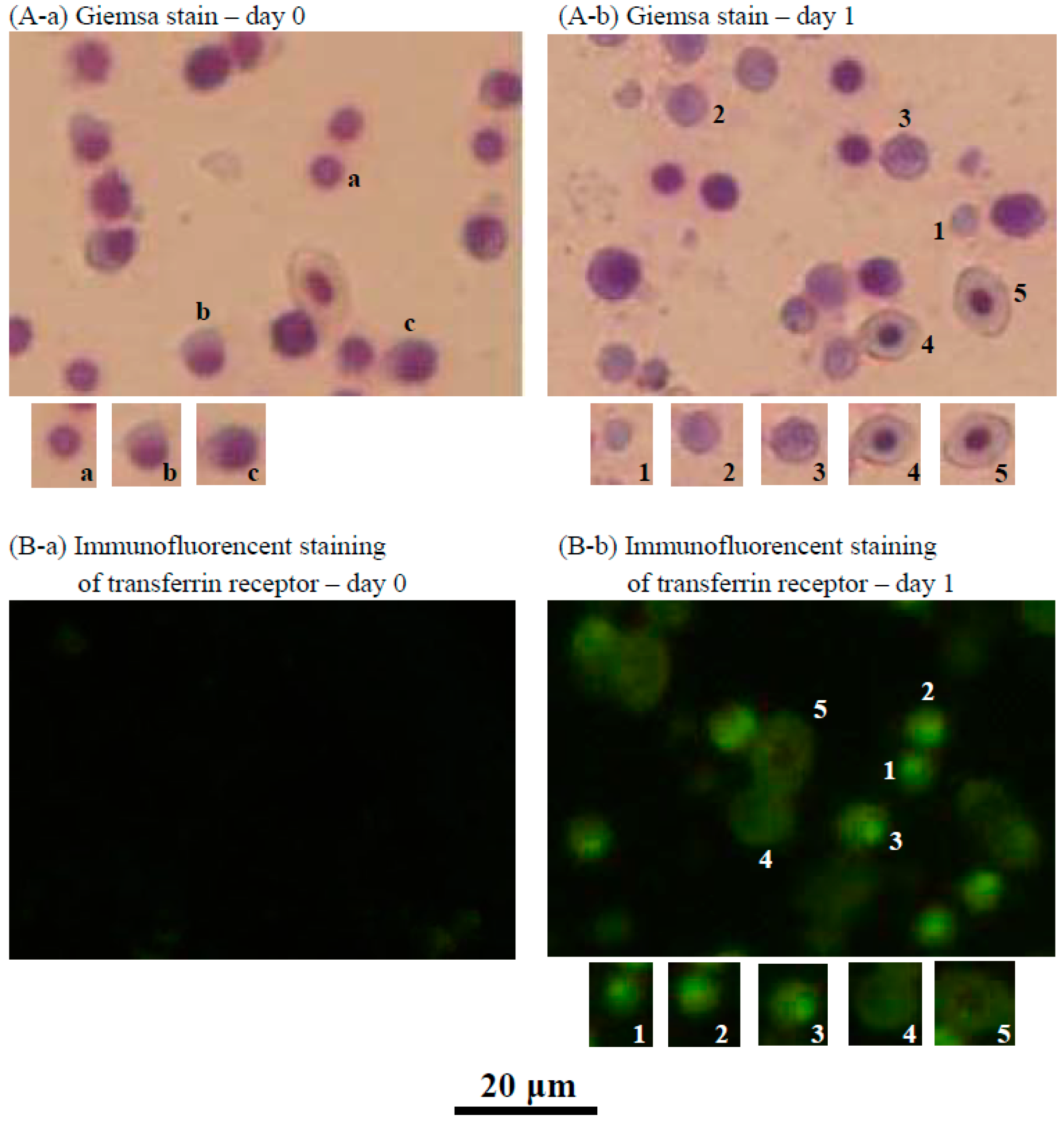
2.3. Characteristics of the Cultured Head Kidney Cells of the Crucian Carp
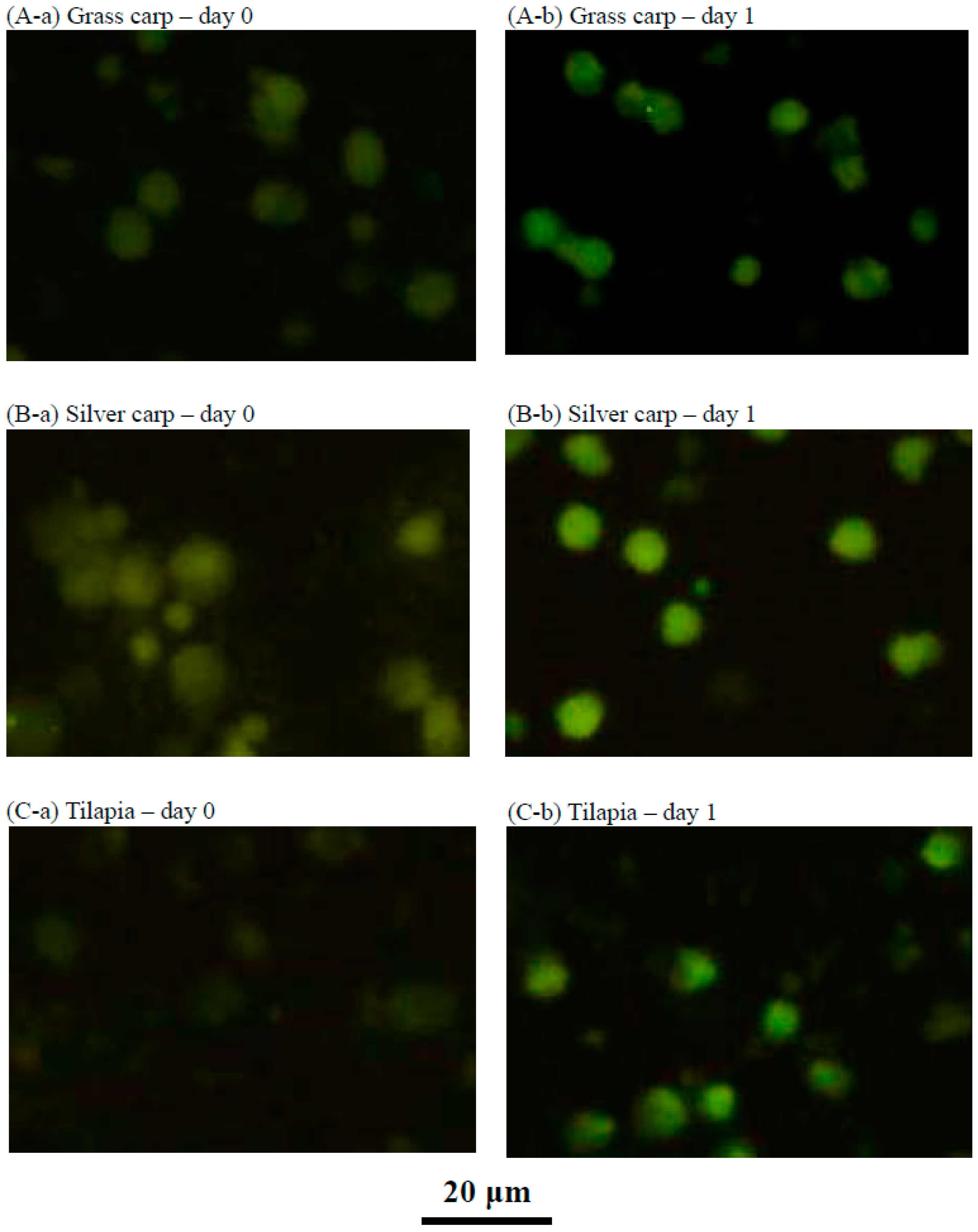
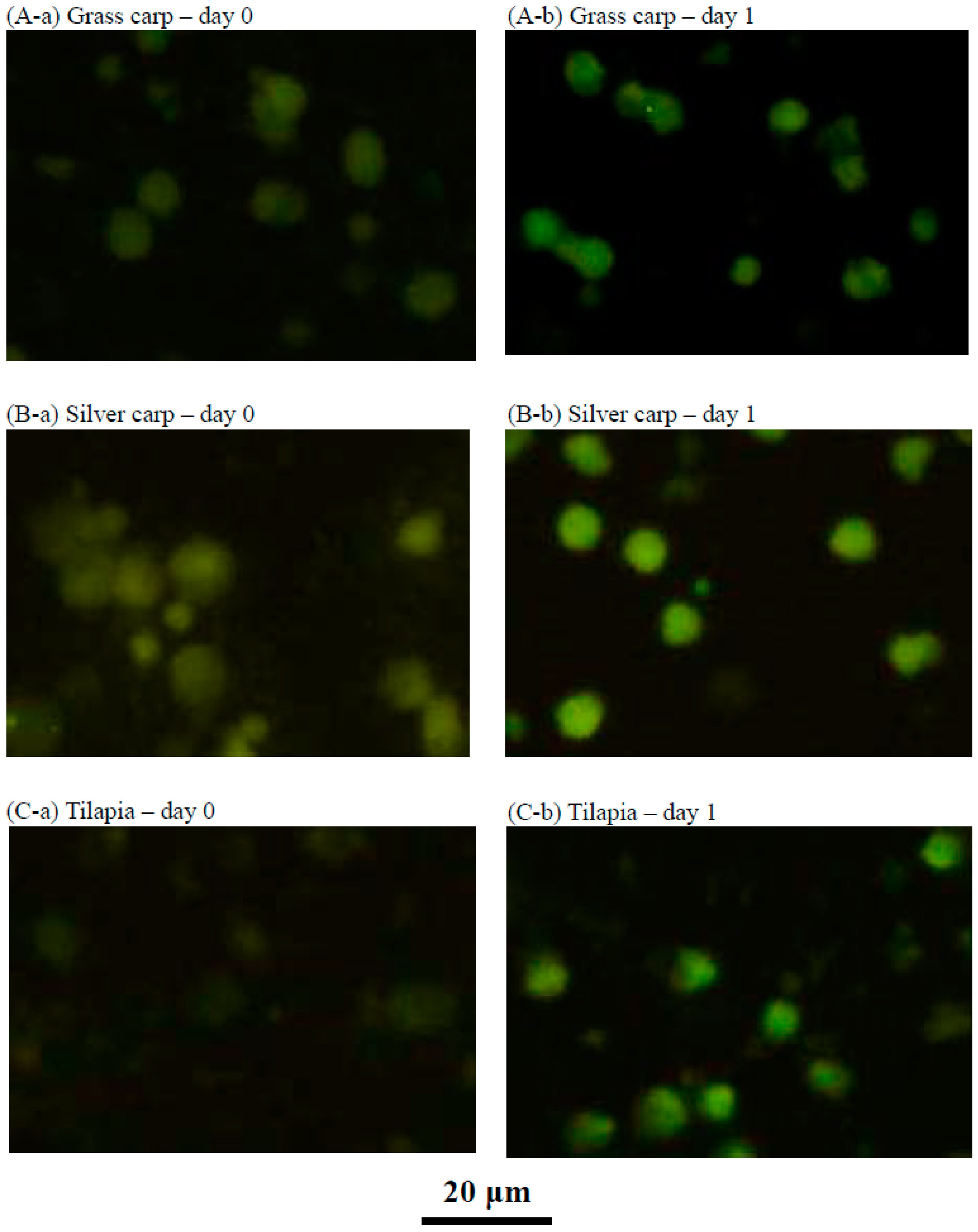
2.4. Characteristics of the Cultured Head Kidney Cells of Grass Carp, Silver Carp and Tilapia
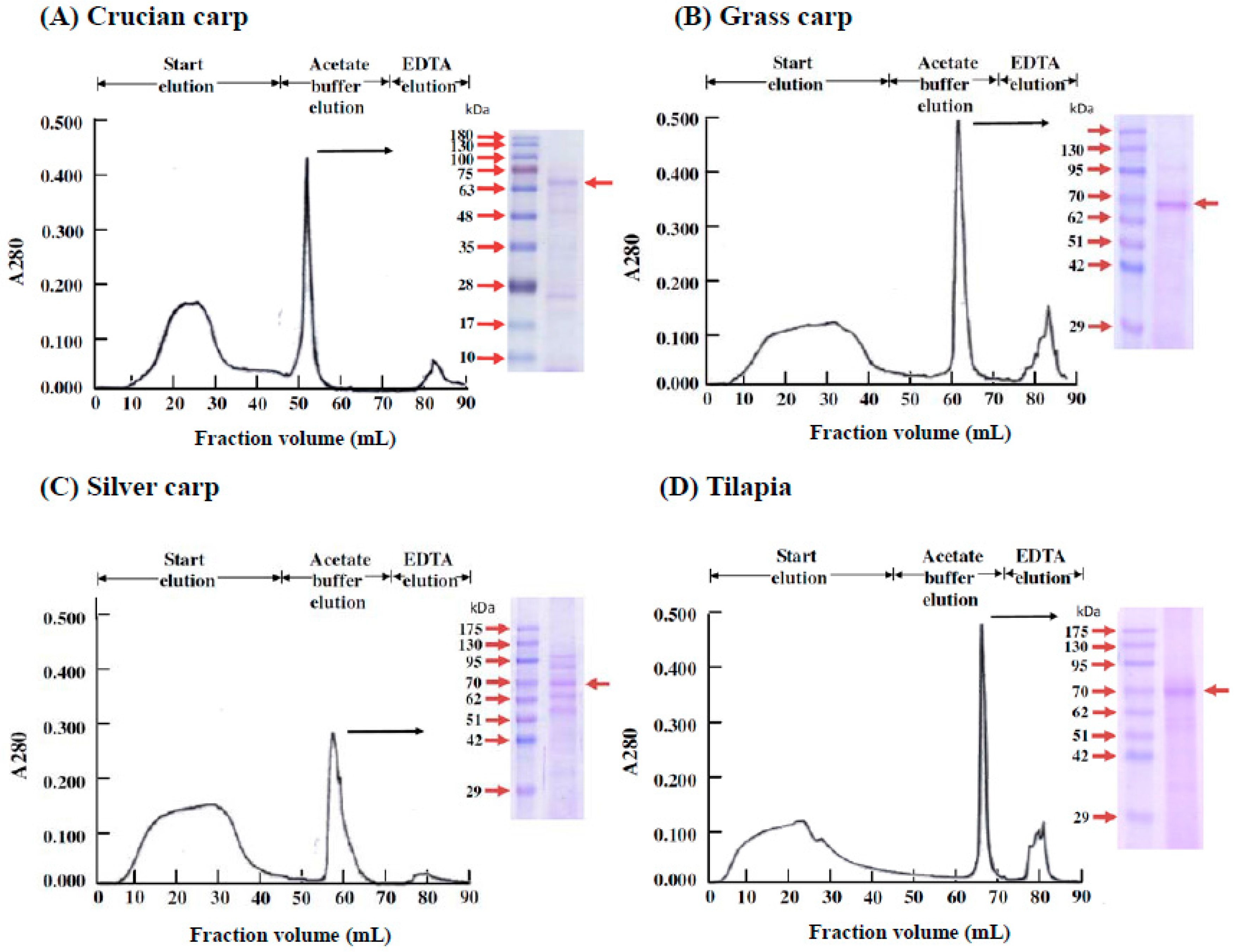
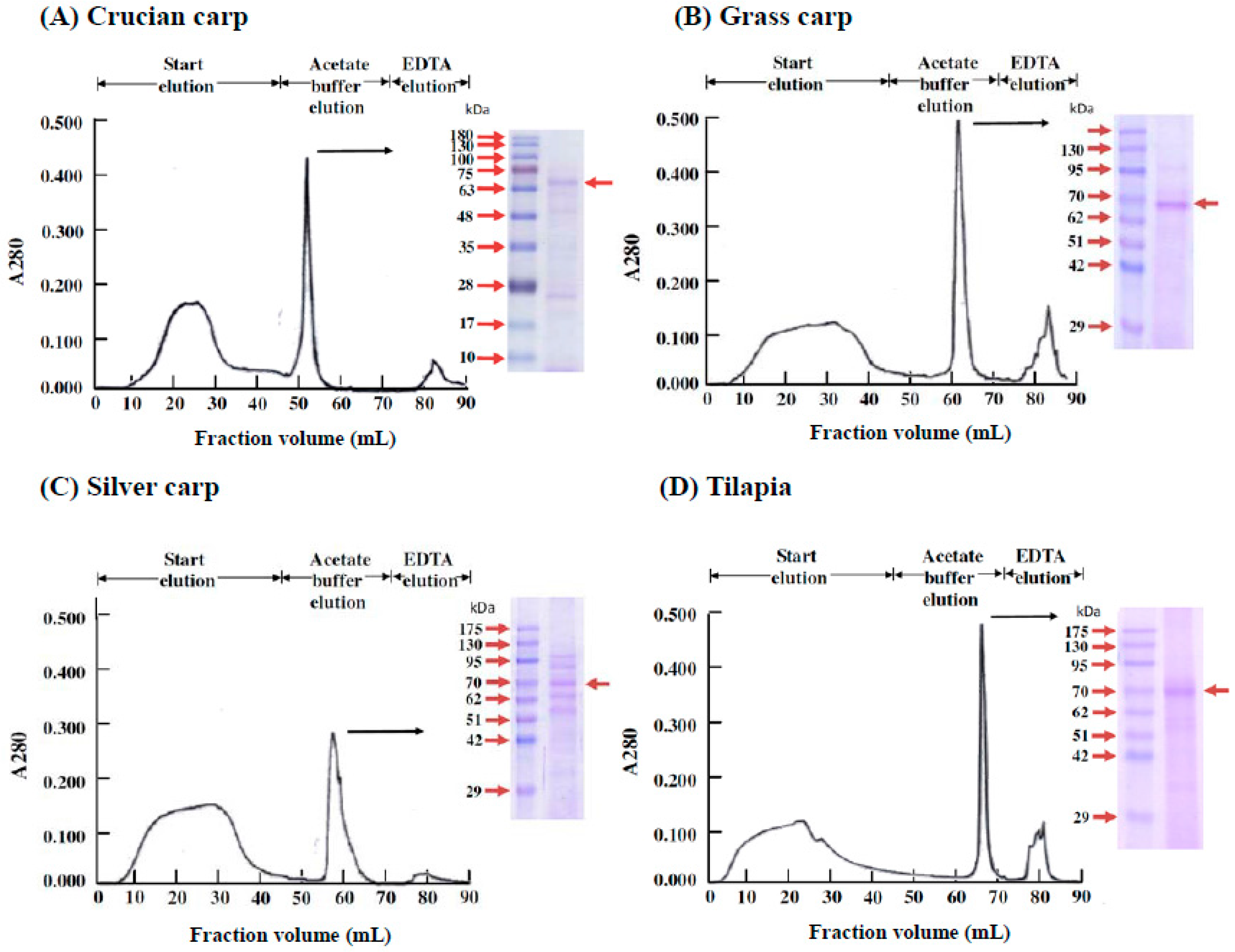
2.5. Isolation of the Active Substance in Fish Serum That Stimulates the Proliferation of Fraction 1 Cells in the Head Kidney Cells of the Four Fish Species
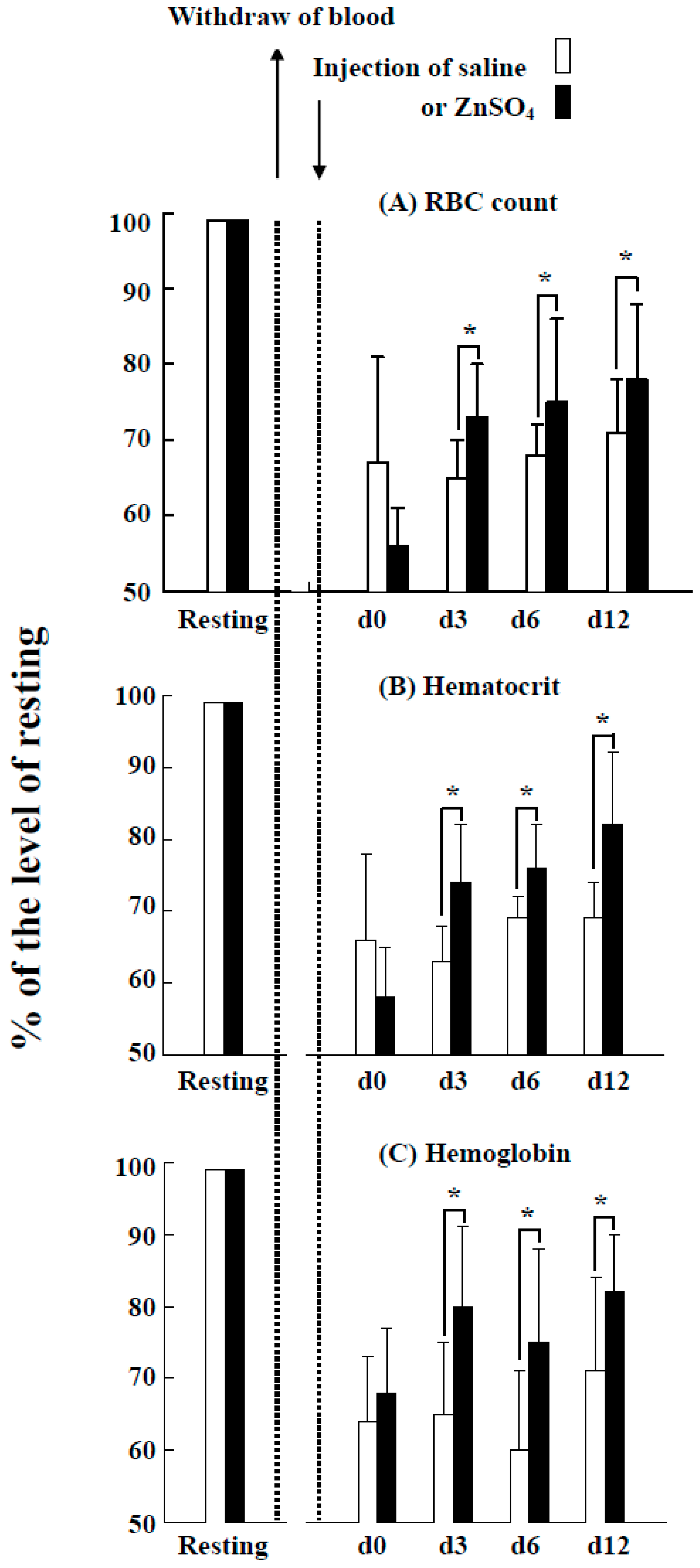
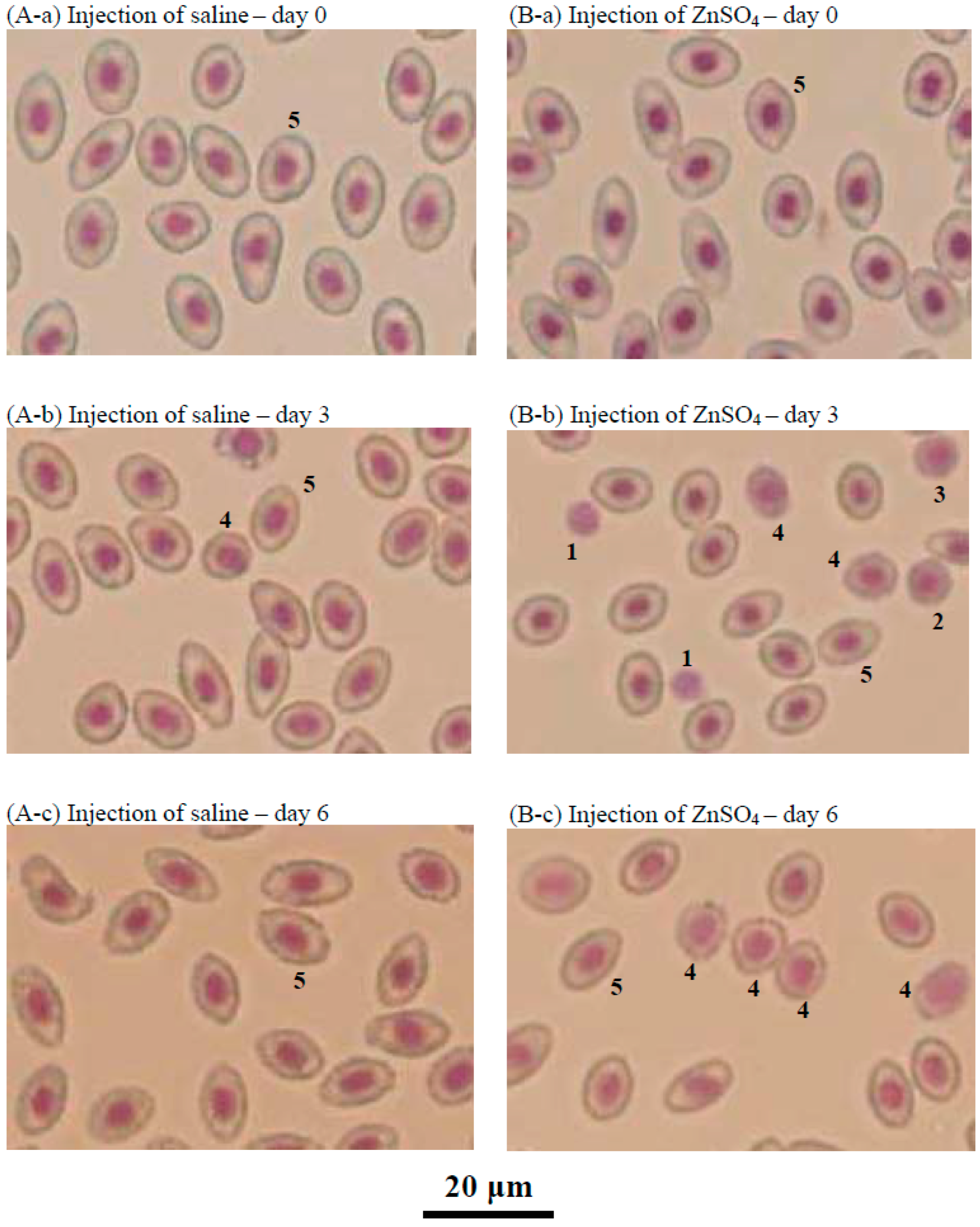
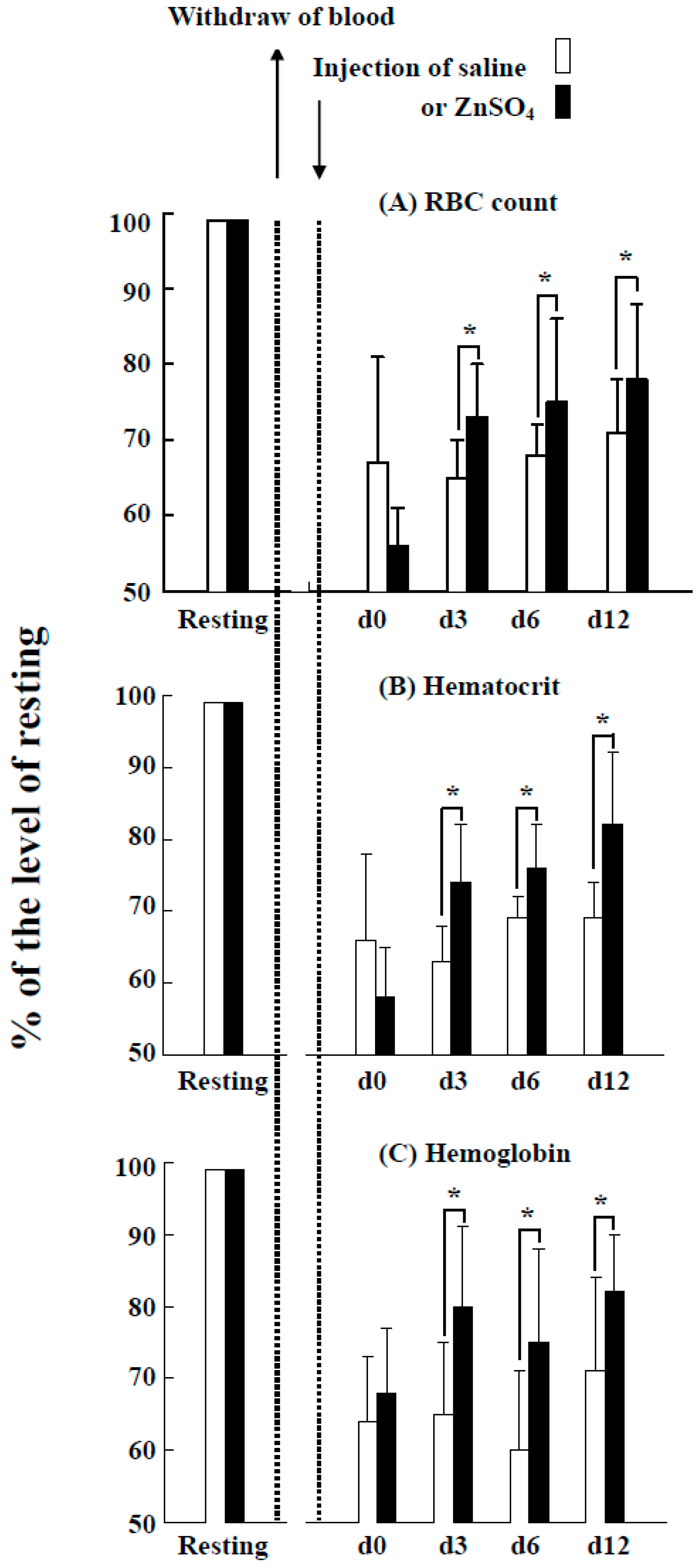
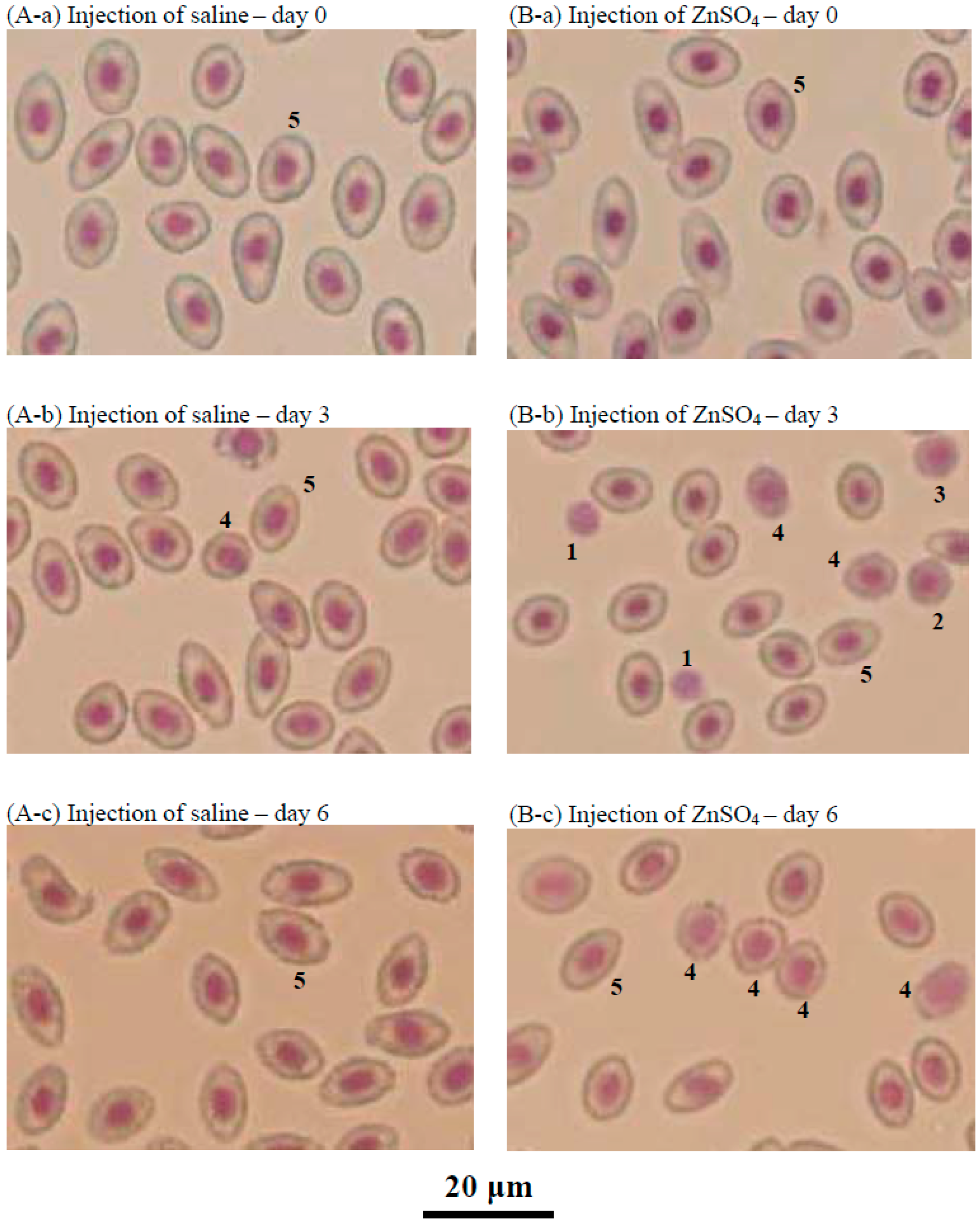
2.6. Effects of the Injection of ZnSO4 on Erythropoiesis in a Blood Loss Tilapia Model
3. Discussion
4. Materials and Methods
4.1. Fish
4.2. Preparation of Head Kidney Cell Suspensions and Fish Serum
4.3. Suspension Culture of Head Kidney Cells with or without ZnCl2 Supplementation
4.4. Separation of Harvested Head Kidney Cells into Two Fractions by Discontinuous Percoll Density Centrifugation
4.5. Effects of ZnCl2 on the Growth of Fraction 1 Cells from the Head Kidneys of Different Fish Species
4.6. Microscopic Observation of Head Kidney Cells or Tilapia Blood by Giemsa Staining
4.7. Immunofluorescence Staining of the Transferrin Receptor in Fish Head Kidney Cells
4.8. Isolation of the Active Substance in Fish Serum That Stimulates the Proliferation of Fraction 1 Cells in Fish Head Kidneys
4.9. Experimental Blood Loss in Tilapia and Injection of ZnSO4 Solution
4.10. Blood Analysis
4.11. Data Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| EPO | Erythropoietin |
| IMAC | Immobilized metal affinity chromatography |
References
- Chen, Y.H.; Fang, S.W.; Jeng, S.S. Zinc transferrin stimulates red blood cell formation in the head kidney of common carp (Cyprinus carpio). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2013, 166, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Fänge, R. Blood cells, haemopoiesis and lymphomyeloid tissues in fish. Fish Shellfish Immunol. 1994, 4, 405–411. [Google Scholar] [CrossRef]
- Fijan, N. Composition of main haematopoietic compartments in normal and bled channel catfish. J. Fish Biol. 2002, 60, 1142–1154. [Google Scholar] [CrossRef]
- Fijan, N. Morphogenesis of blood cell lineages in channel catfish. J. Fish. Biol. 2002, 60, 999–1014. [Google Scholar] [CrossRef]
- Bunn, H.F. Erythropoietin. Cold Spring Harb. Perspect. Med. 2013, 3, a011619. [Google Scholar] [CrossRef] [PubMed]
- Jelkmann, W. Physiology and pharmacology of erythropoietin. Transfus. Med. Hemother. 2013, 40, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.C.; Kakuta, I.; Mok, H.O.; Rummer, J.L.; Randall, D. Effects of moderate and substantial hypoxia on erythropoietin levels in rainbow trout kidney and spleen. J. Exp. Biol. 2006, 209, 2734–2738. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, M.A.; Dunning, S.P.; Bunn, H.F. Regulation of the erythropoietin gene: Evidence that the oxygen sensor is a heme protein. Science 1988, 242, 1412–1415. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Chen, H.H.; Jeng, S.S. Rapid renewal of red blood cells in the common carp following prolonged exposure to air. Fish. Sci. 2015, 81, 255–265. [Google Scholar] [CrossRef]
- Jeng, S.S.; Wang, M.S. Isolation of a Zn-binding protein mediating cell adhesion from common carp. Biochem. Biophys. Res. Commun. 2003, 309, 733–742. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.Y.; Chen, Y.H.; Liu, C.L.; Jeng, S.S. Role of high zinc levels in the stress defense of common carp. Fish. Sci. 2011, 77, 557–574. [Google Scholar] [CrossRef]
- Catton, W.T. Blood cell formation in certain teleost fishes. Blood 1951, 6, 39–60. [Google Scholar] [PubMed]
- Jeng, S.S.; Lo, H.W. High zinc concentration in common carp viscera. Bull. Jpn. Soc. Sci. Fish. 1974, 40, 509. [Google Scholar] [CrossRef]
- Jeng, S.S.; Lin, T.Y.; Wang, M.S.; Chang, Y.Y.; Chen, C.Y.; Chang, C.C. Anoxia survival in common carp and crucian carp is related to high zinc concentration in tissues. Fish. Sci. 2008, 74, 627–634. [Google Scholar] [CrossRef]
- Montero, D.; Tort, L.L.; Izquierdo, M.S.; Socorro, J.; Vergara, J.M.; Robaina, L.; Fernández-Palacios, H. Hematological recovery in Sparus aurata after bleeding. A time course study. Rev. Esp. Fisiol. 1995, 51, 219–226. [Google Scholar] [PubMed]
- Kondera, E.; Dmowska, A.; Rosa, M.; Witeska, M. The effect of bleeding on peripheral blood and head kidney hematopoietic tissue in common carp (Cyprinus carpio). Turk. J. Vet. Anim. Sci. 2012, 36, 169–175. [Google Scholar]
- Fazio, F.; Piccione, G.; Arfuso, F.; Faggio, C. Peripheral blood and head kidney haematopoietic tissue response to experimental blood loss in mullet (Mugil cephalus). Mar. Biol. Res. 2015, 11, 197–202. [Google Scholar] [CrossRef]
- Fukada, T.; Kambe, T. (Eds.) Zinc Signals in Cellular Functions and Disorders; Springer: Tokyo, Japan, 2014.
- Freshney, R.I. Culture of Animal Cells, 4th ed.; Wiley-Liss: New York, NY, USA, 2000. [Google Scholar]
- Kondera, E. Haematopoiesis in the head kidney of common carp (Cyprinus carpio L.): A morphological study. Fish Physiol. Biochem. 2011, 37, 355–362. [Google Scholar] [CrossRef] [PubMed]
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-H.; Shiu, J.-R.; Ho, C.-L.; Jeng, S.-S. Zinc as a Signal to Stimulate Red Blood Cell Formation in Fish. Int. J. Mol. Sci. 2017, 18, 138. https://doi.org/10.3390/ijms18010138
Chen Y-H, Shiu J-R, Ho C-L, Jeng S-S. Zinc as a Signal to Stimulate Red Blood Cell Formation in Fish. International Journal of Molecular Sciences. 2017; 18(1):138. https://doi.org/10.3390/ijms18010138
Chicago/Turabian StyleChen, Yen-Hua, Jhe-Ruei Shiu, Chia-Ling Ho, and Sen-Shyong Jeng. 2017. "Zinc as a Signal to Stimulate Red Blood Cell Formation in Fish" International Journal of Molecular Sciences 18, no. 1: 138. https://doi.org/10.3390/ijms18010138
APA StyleChen, Y.-H., Shiu, J.-R., Ho, C.-L., & Jeng, S.-S. (2017). Zinc as a Signal to Stimulate Red Blood Cell Formation in Fish. International Journal of Molecular Sciences, 18(1), 138. https://doi.org/10.3390/ijms18010138