
Biology, Pest Status, Microbiome and Control of Kudzu Bug (Hemiptera: Heteroptera: Plataspidae): A New Invasive Pest in the U.S.


Abstract
:
1. Introduction
2. Identification of the Kudzu Bug and Its Bacterial Symbionts in the U.S.
3. Kudzu Bug Host Plants Include Soybean
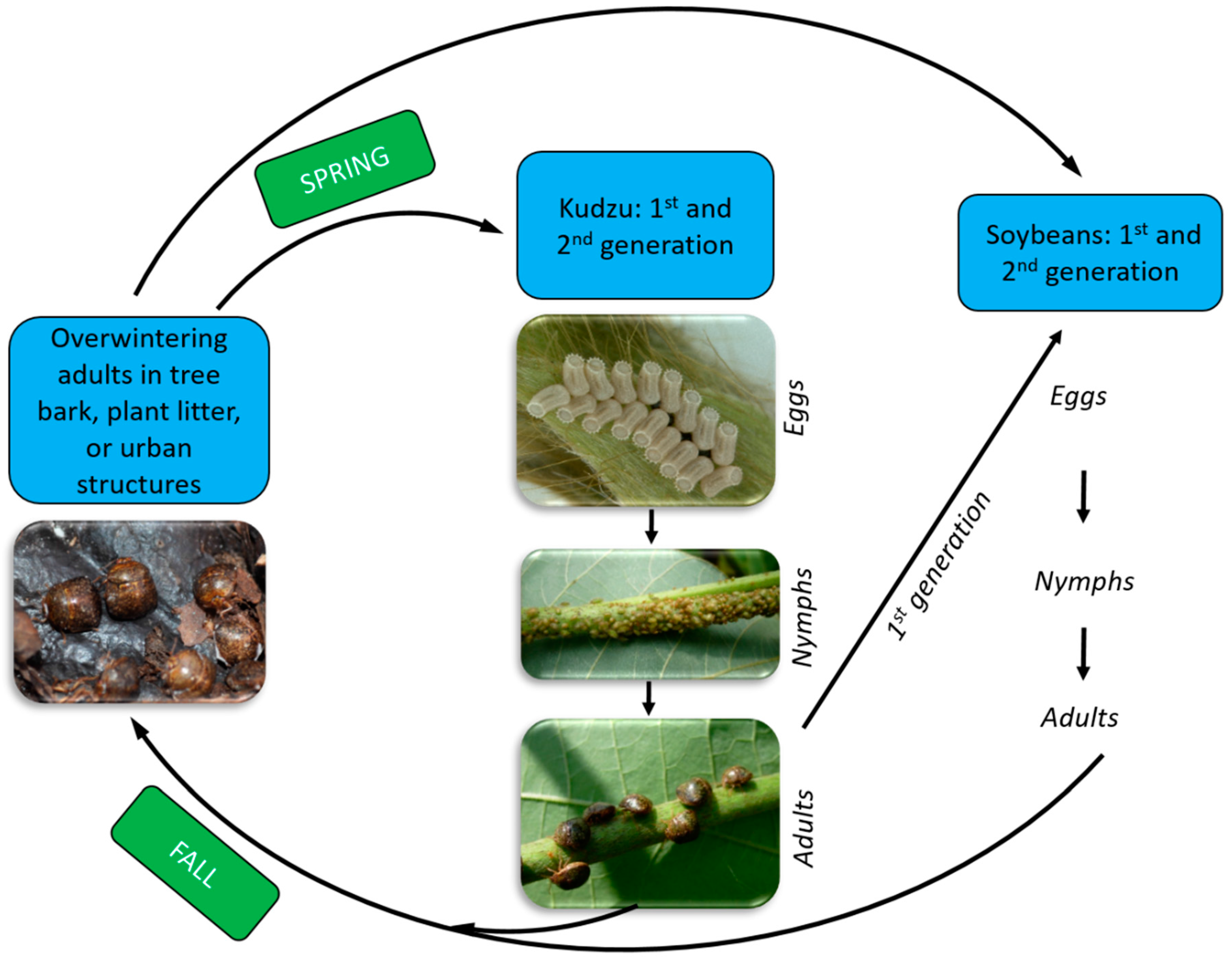
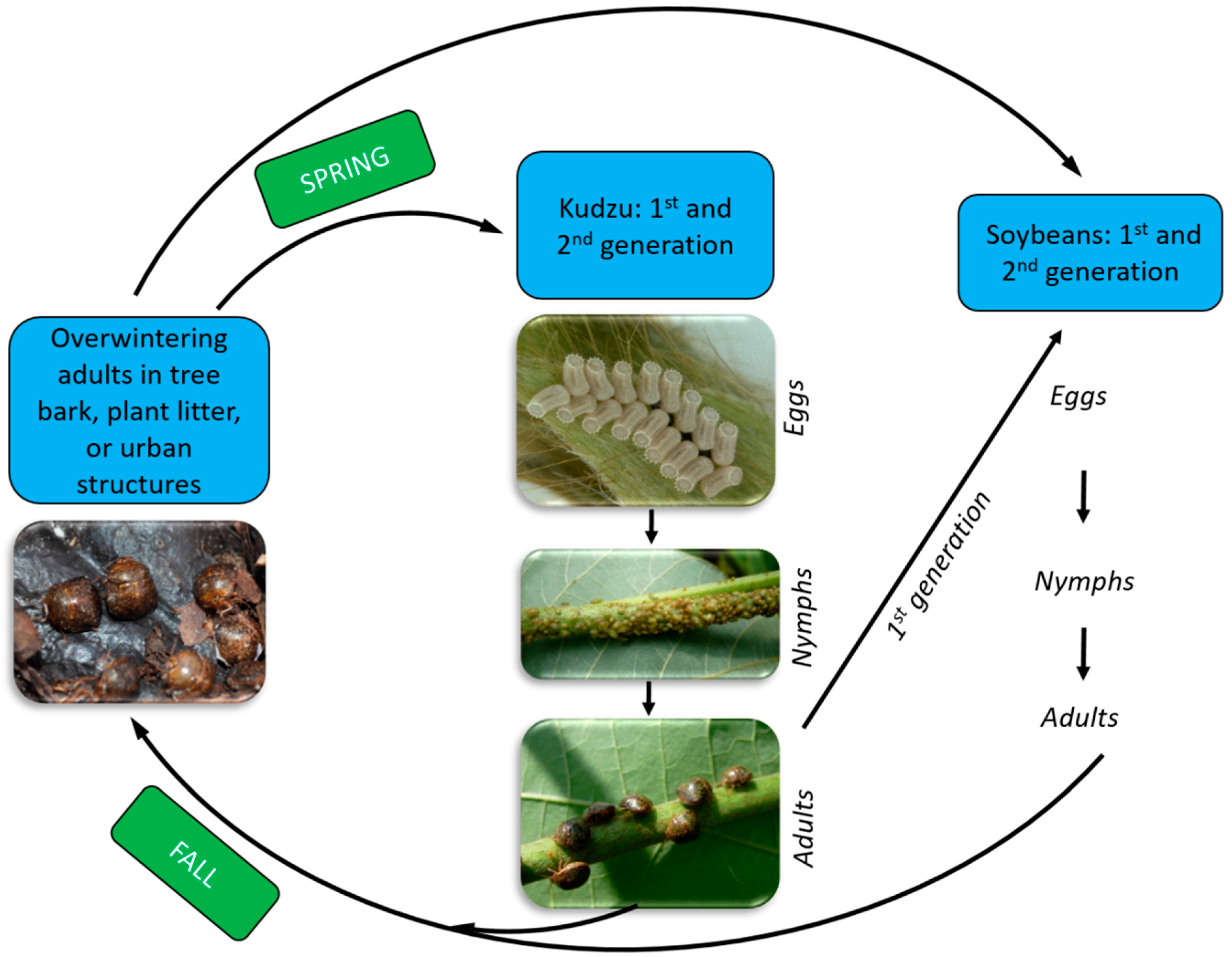
4. Kudzu Bug Biology
5. Soybean Damage from Kudzu Bugs
6. The Identity of Plataspid Endosymbionts
7. The Role of Endosymbionts in Development of Plataspids
8. The Role of Endosymbionts in the Crop-Pest Status of Plataspids
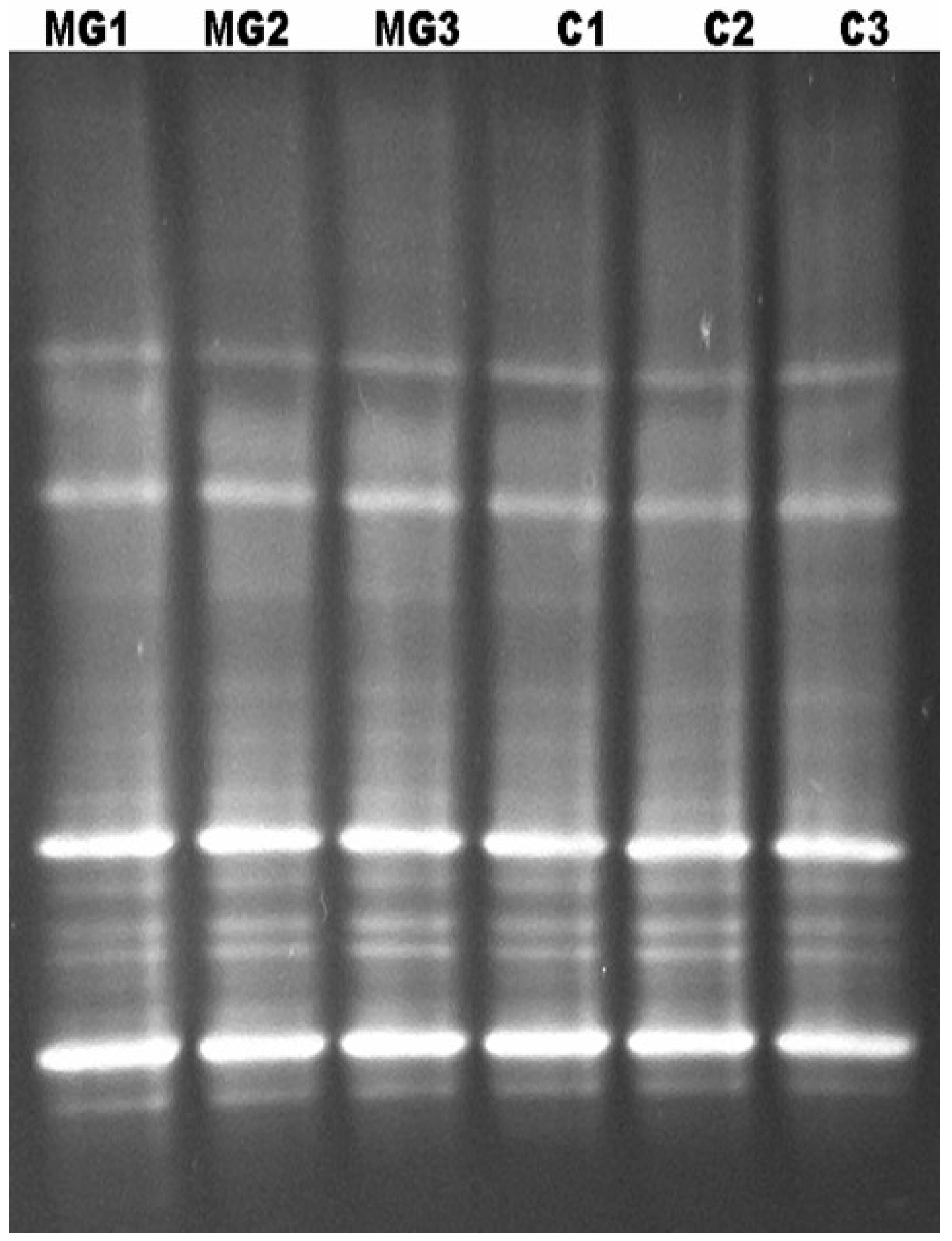
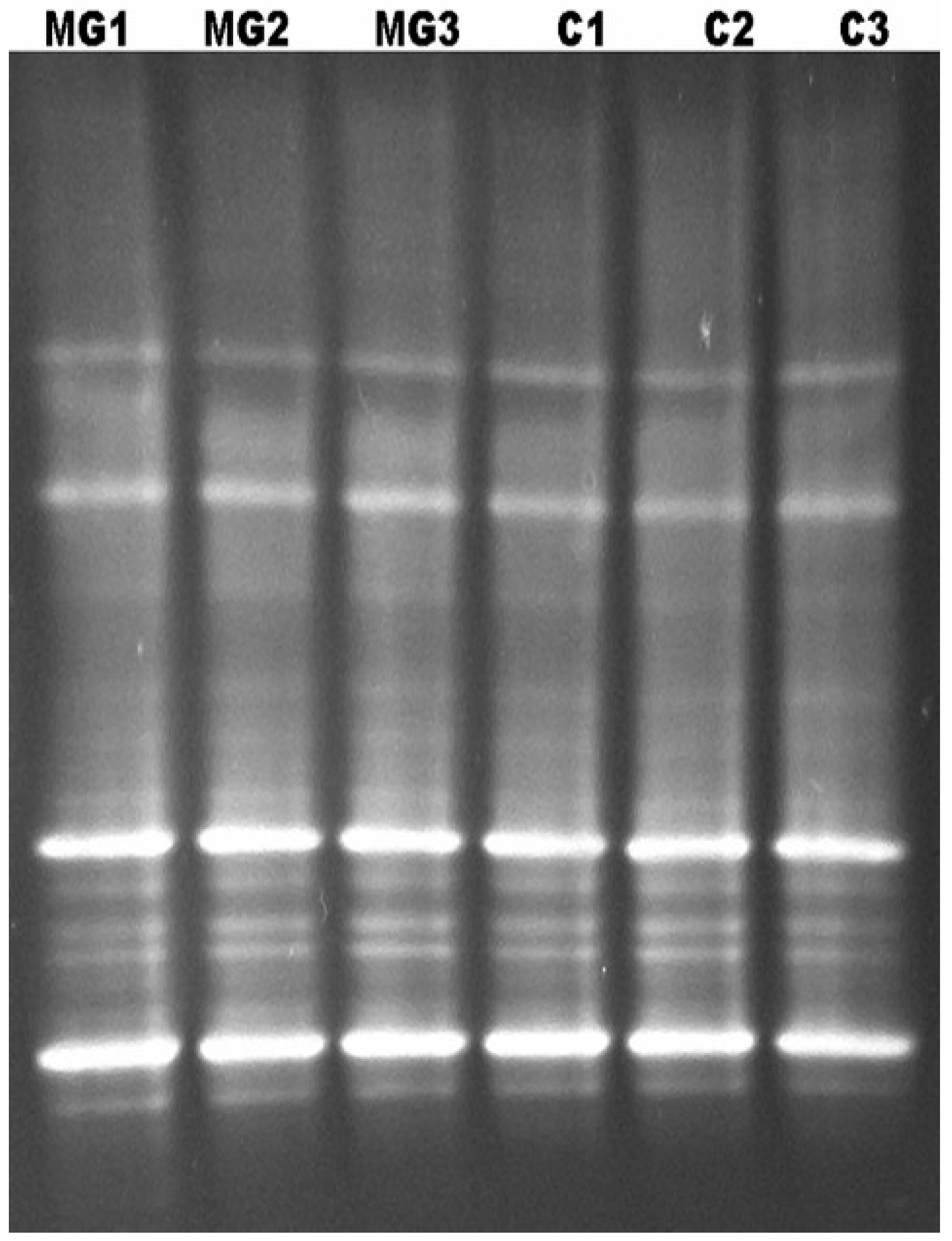
9. Characterization of the Bacterial Diversity of the Kudzu Bug by Denaturing Gradient Gel Electrophoresis (DGGE)
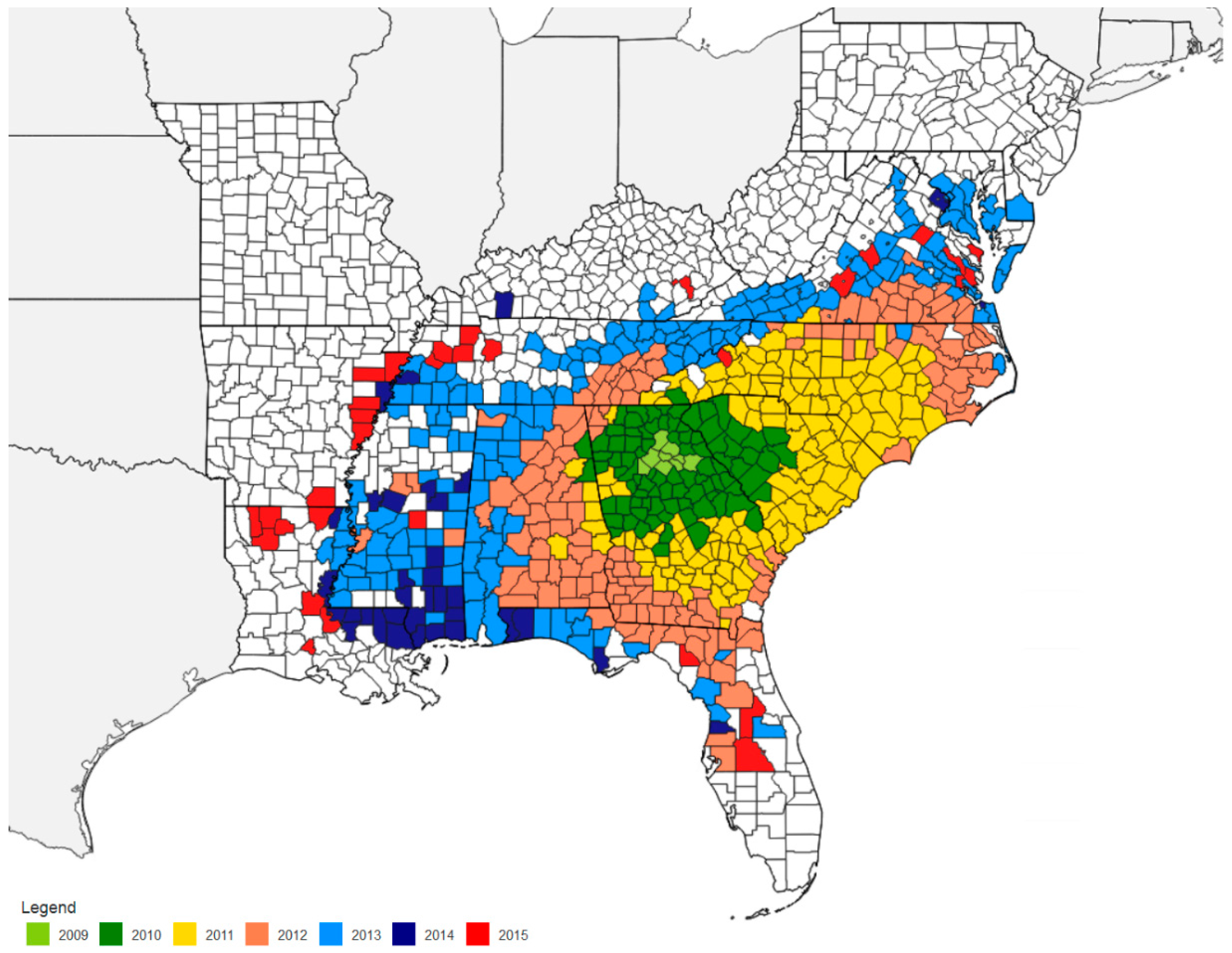
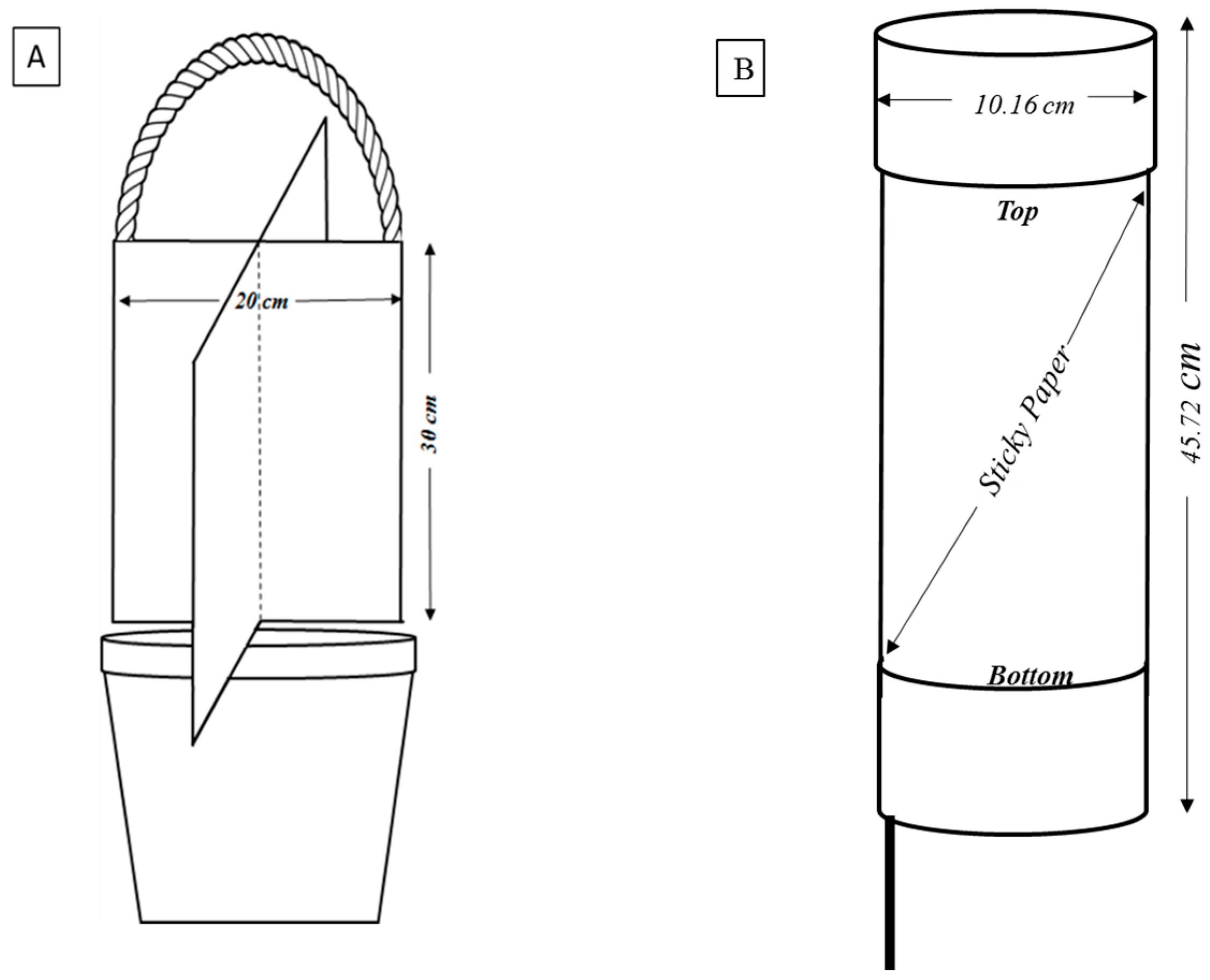
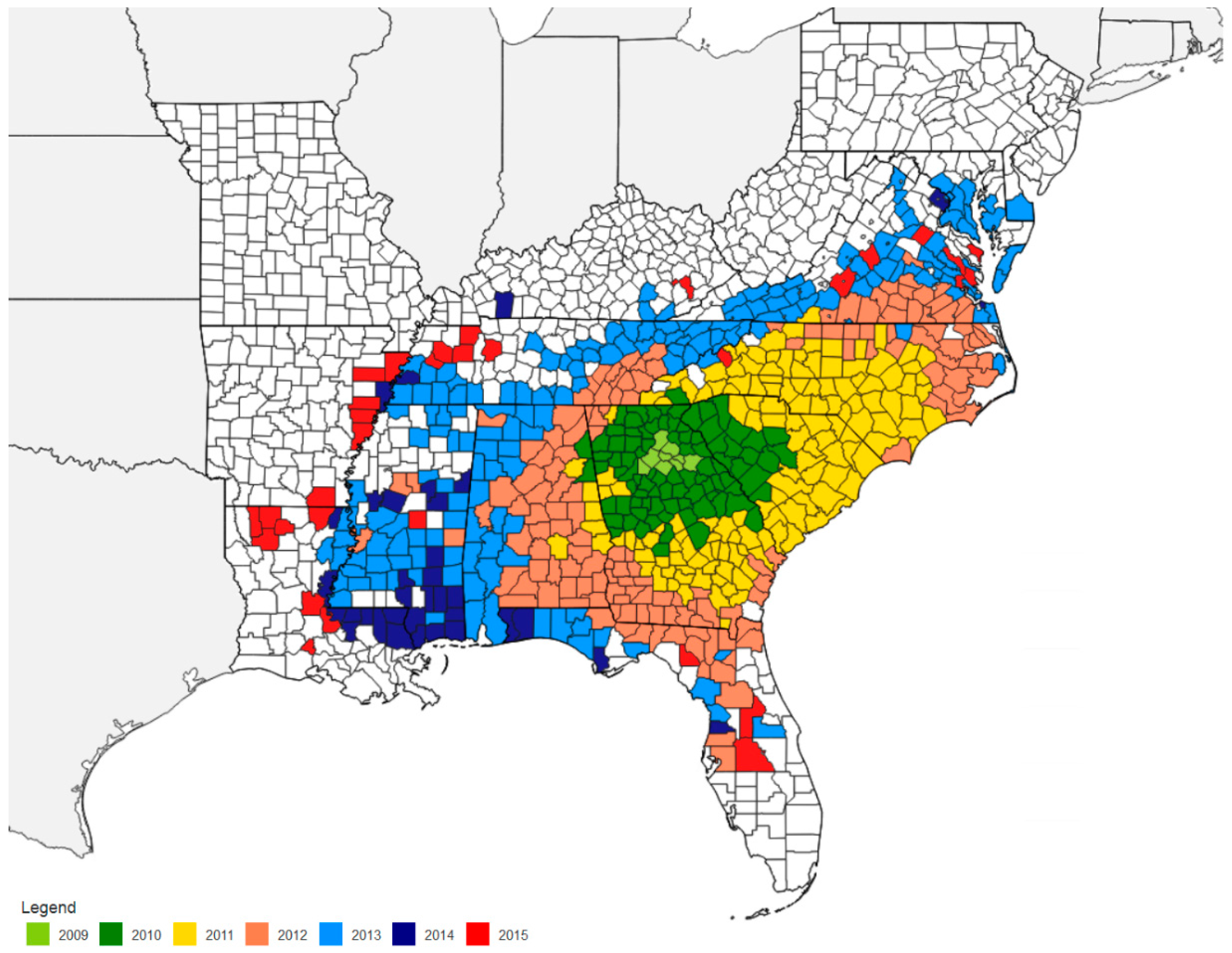
10. Methods for Monitoring Kudzu Bug Population
11. Management of Kudzu Bug in Soybean
12. Impact of Cultural Practices on Kudzu Bug Population Density
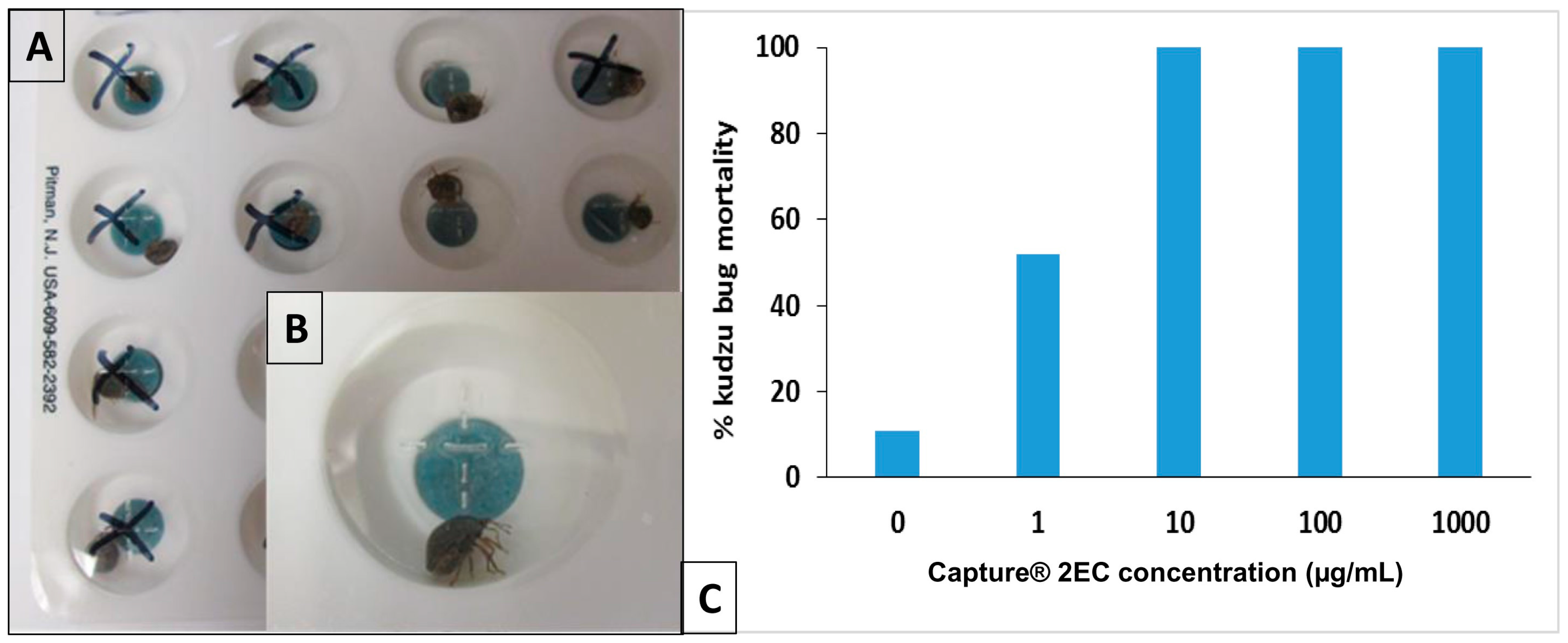
13. Kudzu Bug Feeding Disruption Test (FDT) for Insecticide Susceptibility
14. Kudzu Bug Natural Enemies
15. Future Directions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Eger, J.E., Jr.; Ames, L.M.; Suiter, D.R.; Jenkins, T.M.; Rider, D.A.; Halbert, S.E. Occurrence of the Old World bug Megacopta cribraria (Fabricius) (Heteroptera: Plataspidae) in Georgia: A serious home invader and potential legume pest. Insecta Mundi 2010, 121, 1–11. [Google Scholar]
- Suiter, D.R.; Eger, J.E., Jr.; Gardner, W.A.; Kemerait, R.C.; All, J.N.; Roberts, P.M.; Greene, J.K.; Ames, L.M.; Buntin, G.D.; Jenkins, T.M.; et al. Discovery and distribution of Megacopta cribraria (Hemiptera: Heteroptera: Plataspidae) in northeast Georgia. J. Integr. Pest Manag. 2010, 1, 1–4. [Google Scholar] [CrossRef]
- Zhang, Y.; Hanula, J.L.; Horn, S. The biology and preliminary host range of Megacopta cribraria (Heteroptera: Plataspidae) and its impact on kudzu growth. Environ. Entomol. 2012, 41, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Reisig, D.; Bacheler, J. Kudzu Bug (Megacopta cribraria), a New Potentially Devastating Pest of Soybeans. Available online: https://organicgrains.ces.ncsu.edu/2012/12/kudzu-bug-megacopta-cribraria-a-new-potentially-devastating-pest-of-soybeans/ (accessed on 13 September 2016).
- Gardner, W.A.; Peeler, H.B.; LaForest, J.; Roberts, P.R.; Sparks, A.N., Jr.; Greene, J.K.; Reisig, D.D.; Suiter, D.R.; Bacheler, J.S.; Kidd, K.; et al. Confirmed distribution and occurrence of Megacopta cribraria (F.) (Hemiptera: Plataspidae) in the southeastern United States. J. Entomol. Sci. 2013, 48, 118–127. [Google Scholar] [CrossRef]
- Gardner, W. Megacopta cribraria Distribution Map. 2015. Available online: http://www.kudzubug.org/distribution-map/ (accessed on 20 November 2015).
- Department of Agriculture, Economic Research Service. Soybeans & Oil Crops. Related Data & Statistics, Updated 30 March 2015. Available online: http://www.ers.usda.gov/data-products/oil-crops-yearbook.aspx (accessed on 21 November 2015).
- USDA National Agricultural Statistics Service (NASS). 2012 Census of Agriculture. 2014. Available online: http://quickstats.nass.usda.gov/?source_desc=CENSUS#09FBC30A-9A58-3D06-A044-31FF27D3682C (accessed on 15 November 2015). [Google Scholar]
- Seiter, N.J.; Greene, J.K.; Reay-Jones, F.P. Reduction of soybean yield components by Megacopta cribraria (Hemiptera: Plataspidae). J. Econ. Entomol. 2013, 106, 1676–1683. [Google Scholar] [CrossRef] [PubMed]
- Blount, J.L.; Buntin, G.D.; Roberts, P.M. Effect of planting date and maturity group on soybean yield response to injury by the kudzu bug, Megacopta cribraria F. (Hemiptera: Plataspidae). J. Econ. Entomol. 2016, 109, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Del Pozo-Valdivia, A.I.; Seiter, N.J.; Reisig, D.D.; Greene, J.K.; Reay-Jones, F.P.P.; Bacheler, J.S. Megacopta cribraria (Hemiptera: Plataspidae) Population dynamics in soybeans as influenced by planting date, maturity group, and insecticide use. J. Econ. Entomol. 2016, 109, 1141–1155. [Google Scholar] [CrossRef] [PubMed]
- Ruberson, J.R.; Takatsu, K.; Buntin, G.D.; Eger, J.E., Jr.; Gardner, W.A.; Greene, J.K.; Jenkins, T.M.; Jones, W.A.; Olson, D.M.; Roberts, P.M.; et al. From Asian curiosity to eruptive American pest: Megacopta cribraria (Hemiptera: Plataspidae) and prospects for its biological control. Appl. Entomol. Zool. 2013, 48, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Waldvogel, M.; Alder, P. Kudzu Bug—A Nuisance and Agricultural Pest. North Carolina State University Department of Entomology Residential, Structural and Community Pests Insect Note—ENT/rsc#37. 2011. Available online: https://www.ces.ncsu.edu/depts/ent/notes/Urban/kudzubug.html (accessed on 13 May 2016).
- Del Pozo-Valdivia, A.I.; Reisig, D.D. First generation Megacopta cribraria (Hemiptera: Plataspidae) can develop on soybeans. J. Econ. Entomol. 2013, 106, 533–535. [Google Scholar] [CrossRef] [PubMed]
- Greene, J.K.; Roberts, P.M.; Gardner, W.A.; Reay-Jones, F.P.F.; Seiter, N. Kudzu Bug—Identification and Control in Soybeans. Available online: http://www.kudzubug.org/grower/ (accessed on 13 September 2016).
- Golec, J.R.; Hu, X.P. Preoverwintering copulation and female ratio bias: life history characteristics contributing to the invasiveness and rapid spread of Megacopta cribraria (Heteroptera: Plataspidae). Environ. Entomol. 2015, 44, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.S.; Cui, J.; Zang, L.S. Development, survival, and reproduction of Megacopta cribraria (Heteroptera: Plataspidae) at different constant temperatures. J. Econ. Entomol. 2014, 107, 2061–2066. [Google Scholar] [CrossRef] [PubMed]
- Seiter, N.J.; Greene, J.K.; Reay-Jones, F.P.F. Aggregation and oviposition preferences of Megacopta cribraria (Hemiptera: Plataspidae) in laboratory bioassays. J. Entomol. Sci. 2014, 49, 331–335. [Google Scholar] [CrossRef]
- Del Pozo-Valdivia, A.I. Effects of Agronomic Practices on the Invasive Soybean Pest Megacopta cribraria (Hemiptera: Plataspidae). Ph.D. Thesis, North Carolina State University, Raleigh, NC, USA, 2016. Available online: https://repository.lib.ncsu.edu/handle/1840.16/11321 (accessed on 13 September 2016). [Google Scholar]
- Seiter, N.J.; Reay-Jones, F.P.F.; Greene, J.K. Within-field spatial distribution of Megacopta cribraria (Hemiptera: Plataspidae) in soybean (Fabales: Fabaceae). Environ. Entomol. 2013, 42, 1363–1374. [Google Scholar] [CrossRef] [PubMed]
- Fritz, B.; Reisig, D.D.; Sorenson, C.E.; del Pozo-Valdivia, A.I.; Carter, T.E., Jr. Host plant resistance to Megacopta cribraria (Hemiptera: Plataspidae) in diverse soybean germplasm maturity groups V through VIII. J. Econ. Entomol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Lahiri, S.; Orr, D.; Sorenson, C.; Cardoza, Y. Overwintering refuge sites for Megacopta cribraria (Hemiptera: Plataspidae). J. Entomol. Sci. 2015, 50, 69–73. [Google Scholar] [CrossRef]
- Jenkins, T.M.; Eaton, T.D.; Suiter, D.R.; Eger, J.E., Jr.; Ames, L.M.; Buntin, G.D. Preliminary genetic analysis of a recently-discovered invasive true bug (Hemiptera: Heteroptera: Plataspidae) and its bacterial endosymbiont in Georgia, USA. J. Entomol. Sci. 2010, 45, 1–2. [Google Scholar] [CrossRef]
- Jenkins, T.M.; Eaton, T.D. Population genetic baseline of the first plataspid stink bug symbiosis (Hemiptera: Heteroptera: Plataspidae) reported in North America. Insects 2011, 2, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, T.; Nikoh, N.; Fukatsu, T. Fine-scale geographical origin of an insect pest invading North America. PLoS ONE 2014, 9, e89107. [Google Scholar] [CrossRef] [PubMed]
- Reisig, D.; Bacheler, J. Kudzu Bug (Megacopta cribraria), a New Potentially Devastating Pest of Soybeans. North Carolina State University: Raleigh, NC, USA, 2011. Available online: http://ipm.ncsu.edu/cotton/insectcorner/PDF/Kudzu%20Bug%20Handout_Field%20Crops.Final.pdf (accessed on 10 May 2016).
- Medal, J.; Halbert, S.; Smith, T.; Cruz, A.S. Suitability of selected plants to the bean plataspid, Megacopta cribriaria (Hemiptera: Plataspidae) in No-Choice Tests. Fla. Entomol. 2013, 96, 631–633. [Google Scholar] [CrossRef]
- Blount, J.L.; Buntin, G.D.; Sparks, A.N. Host preference of Megacopta cribraria (Hemiptera: Plataspidae) on selected edible beans and soybean. J. Econ. Entomol. 2015, 109, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Srinivasaperumal, S.; Samuthiravelu, P.; Muthukrishnan, J. Host plant preference and life tale of Megacopta cribraria (Fab.) (Hemiptera: Plataspidae). Proc. Indian Natl. Sci. Acad. 1992, B58, 333–340. [Google Scholar]
- Greene, J.K.; Roberts, P.M.; Garder, W.A.; Reay-Jones, F.P.F.; Seiter, N.J. Kudzu Bug Identification and Control on Soybean. U.S. Soybean Producers Associations, National Soybean Board. Available online: http://www.UnitedSoybean.org (accessed on 13 September 2016).
- Horn, S.; Hanula, J.L. Influence of trap color on collection of the recently-introduced bean plataspid, Megacopta cribraria (Hemiptera: Plataspidae). J. Entomol. Sci. 2011, 46, 85–87. [Google Scholar] [CrossRef]
- Stubbins, F.L.; Seiter, N.J.; Greene, J.K.; Reay-Jones, F.P. Developing sampling plans for the invasive Megacopta cribraria (Hemiptera: Plataspidae) in soybean. J. Econ. Entomol. 2014, 107, 2213–2221. [Google Scholar] [CrossRef] [PubMed]
- Seiter, N.J.; Greene, J.K.; Reay-Jones, F.P.F.; Roberts, P.M.; All, J.N. Insecticidal control of Megacopta cribraria (Hemiptera: Plataspidae) in soybean. J. Entomol. Sci. 2015, 50, 263–283. [Google Scholar] [CrossRef]
- Brown, S.A.; Kerns, D.L.; Williams, T.S.; Emfinger, K.; Jones, N. Evaluation of foliar insecticides for kudzu bug control in soybeans, 2014. Arthropod Manag. Tests 2015, 40, 1. [Google Scholar] [CrossRef]
- Seiter, N.J.; Benson, E.P.; Reay-Jones, F.P.F.; Greene, J.K.; Zungoli, P.A. Residual efficacy of insecticides applied to exterior building material surfaces for control of nuisance infestations of Megacopta cribraria (Hemiptera: Plataspidae). J. Econ. Entomol. 2013, 106, 2448–2456. [Google Scholar] [CrossRef] [PubMed]
- Golec, J.R.; Hu, X.P.; Ray, C.; Woodley, N.E. Strongygaster triangulifera (Diptera: Tachinidae) as a parasitoid of adults of the invasive Megacopta cribraria (Heteroptera: Plataspidae) in Alabama. J. Entomol. Sci. 2013, 48, 352–354. [Google Scholar] [CrossRef]
- Gardner, W.A.; Blount, J.L.; Golec, J.R.; Jones, W.A.; Hu, X.P.; Talamas, E.J.; Evans, R.M.; Dong, X.; Ray, C.H., Jr.; Buntin, G.D.; et al. Discovery of Paratelenomus saccharalis (Dodd) (Hymenoptera: Platygastridae), an egg parasitoid of Megacopta cribraria F. (Hemiptera: Plataspidae) in its expanded North American range. J. Entomol. Sci. 2013, 48, 355–359. [Google Scholar] [CrossRef]
- Medal, J.; Santa Cruz, A.; Williams, K.; Fraser, S.; Wolaver, D.; Smith, T.; Davis, B.J. First record of Paratelenomus saccharalis (Hymenoptera: Platygastridae) on kudzu bug Megacopta cribraria (Heteroptera: Plataspidae) in Florida. Fla. Entomol. 2015, 98, 1250–1251. [Google Scholar] [CrossRef]
- Greenstone, M.H.; Tillman, P.G.; Hu, J.S. Predation of the newly invasive pest Megacopta cribraria (Hemiptera: Plataspidae) in soybean habitats adjacent to cotton by a complex of predators. J. Econ. Entomol. 2014, 107, 947–954. [Google Scholar] [CrossRef] [PubMed]
- Seiter, N.J.; Grabke, A.; Greene, J.K.; Kerrigan, J.L.; Reay-Jones, F.P. Beauveria bassiana is a pathogen of Megacopta cribraria (Hemiptera: Plataspidae) in South Carolina. J. Entomol. Sci. 2014, 49, 326–330. [Google Scholar] [CrossRef]
- Fritz, B.J. Field and Greenhouse Evaluation of Soybean Genotypes for Resistance to the Exotic Invasive Kudzu Bug, Megacopta cribraria Fabricius. Master’s Thesis, North Carolina State University, Raleigh, NC, USA, 2015. Available online: https://repository.lib.ncsu.edu/handle/1840.16/10413 (accessed on 13 September 2016). [Google Scholar]
- Seiter, N.J.; del-Pozo Valdivia, A.I.; Greene, J.K.; Reay-Jones, F.P.F.; Roberts, P.M.; Reisig, D.R. Action thresholds for managing Megacopta cribraria (Hemiptera: Plataspidae) in soybean based on sweep-net sampling. J. Econ. Entomol. 2015, 108, 1818–1829. [Google Scholar] [CrossRef] [PubMed]
- Seiter, N.J.; del-Pozo Valdivia, A.I.; Greene, J.K.; Reay-Jones, F.P.F.; Roberts, P.M.; Reisig, D.D. Management of Megacopta cribraria (Hemiptera: Plataspidae) at different stages of soybean (Fabales: Fabaceae) development. J. Econ. Entomol. 2016, 109, 1167–1176. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, T.; Kikuchi, Y.; Shimada, M.; Fukatsu, T. Obligate symbiont involved in pest status of host insect. Proc. R. Soc. Lond. Ser. B 2007, 274, 1979–1984. [Google Scholar] [CrossRef] [PubMed]
- Fukatsu, T.; Hosokawa, T. Capsule-transmitted gut symbiotic bacterium of the Japanese common plataspid stinkbug, Megacopta punctatissima. Appl. Environ. Microbiol. 2002, 68, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, T.; Kikuchi, Y.; Nikoh, N.; Shimada, M.; Fukatsu, T. Strict host-symbiont cospeciation and reductive genome evolution in insect gut bacteria. PLoS Biol. 2006, 4, e337. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.; Huynh, L.Y.; Bolender, C.M.; Nelson, K.G.; McCutcheon, J.P. Population genomics of a symbiont in the early stages of a pest invasion. Mol. Ecol. 2014, 23, 1516–1530. [Google Scholar] [CrossRef] [PubMed]
- Lovejoy, R.T.; Johnson, D.A. A molecular analysis of herbivory in adults of the invasive bean plataspid, Megacopta cribraria. Southeast. Nat. 2014, 13, 663–672. [Google Scholar] [CrossRef]
- Golec, J.R.; Hu, X.P.; Yang, L.; Eger, J.E. Kudzu-deprived first-generation Megacopta cribraria (F.) (Heteroptera: Plataspidae) are capable of developing on alternative legume species. J. Agric. Urban Entomol. 2015, 31, 52–61. [Google Scholar] [CrossRef]
- Huskisson, S.M.; Fogg, K.L.; Upole, T.L.; Zehnder, C.B. Seasonal dynamics and plant preferences of Megacopta cribraria, an exotic invasive insect species in the Southeast. Southeast. Nat. 2015, 14, 57–65. [Google Scholar] [CrossRef]
- Thippeswamy, C.; Rajagopal, B.K. Life history of lablab bug, Coptosoma cribraria Faricius (Heteroptera: Plataspidae) on field bean, Lablab purpoureus var. lignosus medikus. Karnataka J. Agric. Sci. 2005, 18, 39–43. [Google Scholar]
- Hosokawa, T.; Kikuchi, Y.; Meng, X.Y.; Fukatsu, T. The making of symbiont capsule in the plataspid stinkbug Megacopta punctatissima. FEMS Microbiol. Ecol. 2005, 54, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Ponnusamy, L.; Gonzalez, A.; van Treuren, W.; Weiss, S.; Parobek, C.M.; Juliano, J.J.; Knight, R.; Roe, R.M.; Apperson, C.S.; Meshnick, S.R. Diversity of Rickettsiales in the microbiome of the lone star tick, Amblyomma americanum. Appl. Environ. Microbiol. 2014, 80, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Ponnusamy, L.; Xu, N.; Stav, G.; Wesson, D.M.; Schal, C.; Apperson, C.S. Diversity of bacterial communities in container habitats of mosquitoes. Microb. Ecol. 2008, 56, 593–603. [Google Scholar] [CrossRef] [PubMed]
- Muyzer, G.E.; de Waal, C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified gene coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [PubMed]
- Cocolin, L.; Manzano, M.; Cantoni, C.; Comi, G. Denaturing gradient gel electrophoresis analysis of the 16S rRNA gene V1 region to monitor dynamic changes in the bacterial population during fermentation of Italian sausages. Appl. Environ. Microbiol. 2001, 67, 5113–5121. [Google Scholar] [CrossRef] [PubMed]
- Nübel, U.; Engelen, B.; Felske, A.; Snaidr, J.; Wieshuber, A.; Amann, R.I.; Ludwig, W.; Backhaus, H. Sequence heterogeneities of genes encoding 16S rRNAs in Paenibacillus polymyxa detected by temperature gradient gel electrophoresis. J. Bacteriol. 1996, 178, 5636–5643. [Google Scholar] [PubMed]
- Salles, J.F.; de Souza, F.A.; van Elsas, J.D. Molecular method to assess the diversity of Burkholderia species in environmental samples. Appl. Environ. Microbiol. 2002, 68, 1595–1603. [Google Scholar] [CrossRef] [PubMed]
- Ulyshen, M.D.; Hanula, J.L. A comparison of the beetle (Coleoptera) fauna captured at two heights above the ground in a North American temperate deciduous forest. Am. Midl. Nat. 2007, 158, 260–278. [Google Scholar] [CrossRef]
- Towes, M.; University of Georgia: Athens, GA, USA. Personal communication, 2013.
- Del Pozo-Valdivia, A.I.; Reisig, D.D.; Sorenson, C.E.; Bacheler, J.S. Effect of using early-planted soybean as a trap crop for Megacopta cribraria in commercial soybean. Environ. Entomol. 2016. in preparation. [Google Scholar]
- Bailey, W.D.; Zhao, G.; Carter, L.M.; Gould, F.; Kennedy, G.G.; Roe, R.M. Feeding disruption bioassay for species and Bacillus thuringiensis resistance diagnosis for Heliothis virescens and Helicoverpa zea in cotton (Lepidoptera: Noctuidae). Crop Prot. 1998, 17, 591–598. [Google Scholar] [CrossRef]
- Bailey, W.D.; Young, H.P.; Wyss, C.F.; Bacheler, J.S.; Roe, R.M. Validation of feeding disruption bioassays for species diagnosis and Bt-resistance monitoring of bollworm and tobacco budworm field populations. In Proceedings of the Beltwide Cotton Conferences, San Antonio, TX, USA, 4–8 January 2000; National Cotton Council: Memphis, TN, USA; pp. 918–923.
- Bailey, W.D.; Brownie, C.; Bacheler, J.S.; Gould, F.; Kennedy, G.G.; Sorenson, C.S.; Roe, R.M. Species diagnosis and Bacillus thuringiensis resistance monitoring of Heliothis virescens and Helicoverpa zea (Lepidoptera: Noctuidae) field strains from the Southern United States using feeding disruption bioassays. J. Econ. Entomol. 2001, 94, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Roe, R.M.; Bailey, W.D.; Young, H.P.; Wyss, C.F. Characterization of spinosad (Tracer) resistance in a laboratory strain of the tobacco budworm and development of novel diagnostics for resistance monitoring in the field. In Proceedings of the Beltwide Cotton Conferences, San Antonio, TX, USA, 4–8 January 2000; National Cotton Council: Memphis, TN, USA; pp. 926–929.
- Roe, R.M.; Long, S.; Cawsey, S.; Bacheler, J.S.; Sorenson, C.E. New commercial feeding disruption bioassay kit for species and insecticide resistance diagnosis in the tobacco budworm and cotton bollworm in cotton. In Proceedings of the Beltwide Cotton Conferences, Atlanta, GA, USA, 8–13 January 2002; National Cotton Council: Memphis, TN, USA.
- Roe, R.M.; Bailey, W.D.; Gould, F.; Kennedy, G.G.; Sutula, C.S. Insecticide Resistance Assay. U.S. Patent 6,517,856 B1, 11 February 2003. [Google Scholar]
- Roe, R.M.; Thompson, D.M.; Rhein, J.L.; Donohue, K.V.; Long, S.; Bacheler, J.S.; Sorenson, C.E. Feeding disruption bioassay for field monitoring of insect susceptibility to Bt-transgenic crops and traditional pesticides. In Proceedings of the Beltwide Cotton Conferences, San Antonio, TX, USA, 5–9 January 2004; National Cotton Council: Memphis, TN, USA; p. 326.
- Roe, R.M.; van Kretschmar, J.B.; Thompson, D.M.; Donohue, K.V.; Sorenson, C.E.; Gould, F.; Stumpf, C.F.; van Duyn, J.W.; Thompson, G.D.; Storer, N.P.; et al. Larval feeding disruption test (FDT) for monitoring insect resistance to Cry1Ab, Cry1Ac and Cry1F. In Proceedings of the Beltwide Cotton Conferences, New Orleans, LA, USA, 4–7 January 2005; National Cotton Council: Memphis, TN, USA; pp. 1651–1661.
- Roe, R.M.; Cabrera, A.R.; Ezzeldin, H.; van Kretschmar, J.B.; Bissinger, B.W. Broad-spectrum biosensor for monitoring current and future transgenic plant technologies for insect control. In Proceedings of the Beltwide Cotton Conferences, Atlanta, GA, USA, 4–7 January 2011; National Cotton Council: Memphis, TN, USA; pp. 1153–1159.
- Khalil, S.; Long, S.; Young, H.; Roe, R.M. Development of a laboratory strain of the tobacco budworm resistant to Denim and a field kit for resistance monitoring. In Proceedings of the Beltwide Cotton Conferences, Atlanta, GA, USA, 8–13 January 2002; National Cotton Council: Memphis, TN, USA.
- Van Kretschmar, J.B.; Bradley, J.R.; Stumpf, C.F.; Roe, R.M. Feasibility of adult feeding disruption tests (FDT) for monitoring lepidopteran resistance to chemical insecticides. In Proceedings of the Beltwide Cotton Conferences, New Orleans, LA, USA, 9–12 January 2007; National Cotton Council: Memphis, TN, USA; pp. 136–140.
- Van Kretschmar, J.B.; Bradley, J.R.; Cabrera, A.R.; Roe, R.M. Novel adult assay to detect insecticide resistance of lepidopteran pests in cotton. In Proceedings of the Beltwide Cotton Conferences, Nashville, TN, USA, 8–11 January 2008; National Cotton Council: Memphis, TN, USA.
- Van Kretschmar, J.B.; Magalhaes, L.C.; Zhu, J.; Cohen, A.C.; Roe, R.M. Feasibility of a novel feeding disruption test (FDT) bioassay kit for rapid resistance detection of sucking pests of cotton. In Proceedings of the Beltwide Cotton Conferences, San Antonio, TX, USA, 5–8 January 2009; National Cotton Council: Memphis, TN, USA; pp. 882–892.
- Van Kretschmar, J.B.; Bailey, W.D.; Arellano, C.; Thompson, G.D.; Sutula, C.L.; Roe, R.M. Feeding disruption tests for monitoring the frequency of larval lepidopteran resistance to Cry1Ac, Cry1F, and Cry1Ab. Crop Prot. 2011, 30, 863–870. [Google Scholar] [CrossRef]
- Van Kretschmar, J.B.; Bradley, J.R.; Cabrera, A.R.; Roe, R.M. Lepidopteran adult feeding disruption test (FDT) to detect insecticide resistance. In Proceedings of the Beltwide Cotton Conferences, Atlanta, GA, USA, 4–7 January 2011; National Cotton Council: Memphis, TN, USA; pp. 1010–1015.
- Van Kretschmar, J.B.; Dhammi, A.; Reisig, D.; Roe, R.M. Challenges for rearing of the stink bug: Successes, failures and RNAi screening. In Proceedings of the Beltwide Cotton Conferences, Atlanta, GA, USA, 4–7 January 2011; National Cotton Council: Memphis, TN, USA; pp. 1169–1175.
- Cabrera, A.R.; van Kretschmar, J.; Bacheler, J.S.; Burrack, H.J.; Sorenson, C.E.; Roe, R.M. Development of hydrateable, commercially-relevant artificial cotton leaves and assay architecture for monitoring insect resistance to Bt. In Proceedings of the Beltwide Cotton Conferences, New Orleans, LA, USA, 4–7 January 2010; National Cotton Council: Memphis, TN, USA; pp. 1290–1296.
- Cabrera, A.R.; van Kretschmar, J.; Bacheler, J.S.; Burrack, H.; Sorenson, C.E.; Roe, R.M. Resistance monitoring of Heliothis virescens to pyramided cotton varieties with a hydrateable, “artificial cotton leaf” bioassay. Crop Prot. 2011, 30, 1196–1201. [Google Scholar] [CrossRef]
- Musser, F.R.; Catchot, A.L., Jr.; Davis, J.A.; Herbert, D.A., Jr.; Lorenz, G.M.; Reed, T.; Reisig, D.D.; Stewart, S.D. 2011 soybean insect losses in the southern US. Midsouth Entomol. 2012, 5, 11–22. [Google Scholar]
- Musser, F.R.; Catchot, A.L., Jr.; Davis, J.A.; Herbert, D.A., Jr.; Lorenz, G.M.; Reed, T.; Reisig, D.D.; Stewart, S.D. 2012 Soybean insect losses in the Southern US. Midsouth Entomol. 2013, 6, 12–24. [Google Scholar]
- Musser, F.R.; Catchot, A.L., Jr.; Davis, J.A.; Herbert, D.A., Jr.; Lorenz, G.M.; Reed, T.; Reisig, D.D.; Stewart, S.D. 2013 Soybean insect losses in the Southern US. Midsouth Entomol. 2014, 7, 15–28. [Google Scholar]
- Musser, F.R.; Catchot, A.L., Jr.; Davis, J.A.; Herbert, D.A., Jr.; Lorenz, G.M.; Reed, T.; Reisig, D.D.; Stewart, S.D. 2014 Soybean insect losses in the Southern US. Midsouth Entomol. 2015, 8, 35–48. [Google Scholar]
- Stubbins, F.; Agudelo, P.; Reay-Jones, F.P.F.; Greene, J.K. First report of a mermithid nematode infecting the invasive Megacopta cribraria (Hemiptera: Plataspidae) in the United States. J. Invertebr. Pathol. 2015, 127, 35–37. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
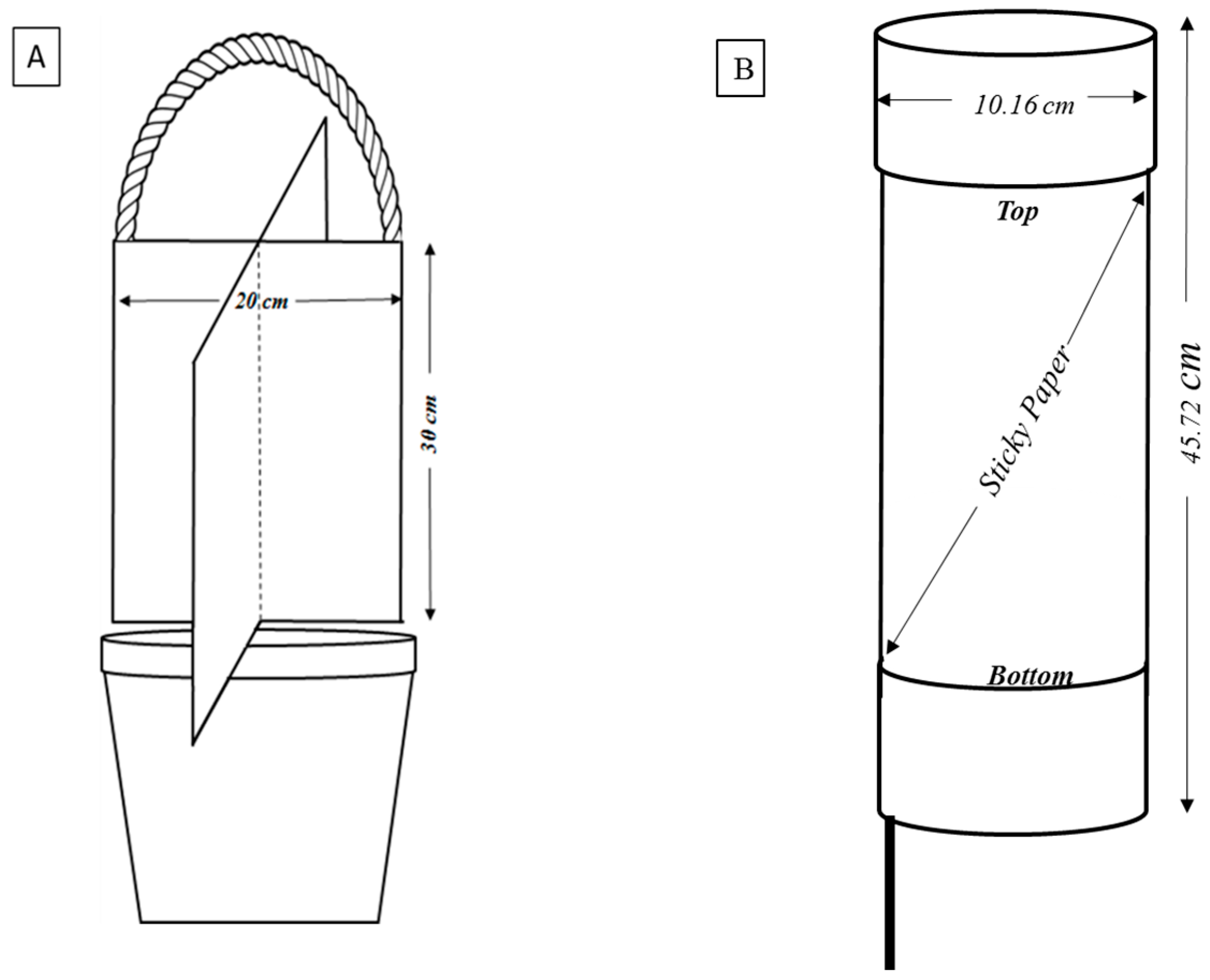{kind=link}
{kind=link}
{kind=link}
| Research Area | Topic | References |
|---|---|---|
| Biology | Morphology | [1,3,13] |
| Life cycle | [1,3,4,13,14,15,16,17] | |
| Spatial distribution | In plants: [18,19] | |
| In the field: [20] | ||
| Population dynamics | [9,20,21] | |
| Overwintering | [12,22] | |
| Identification in US | [1,23,24] | |
| Host and habitat | Native range | [1,25] |
| Discovery and prevalence in the U.S. | [2,3,4,5,13,15] | |
| Host range and preference | [1,3,14,16,26,27,28,29] | |
| Impact as pest | [1,9,12,13,30] | |
| Control and Management | Trap/sampling | [15,26,31,32] |
| Chemical control | [3,30,33,34,35] | |
| Natural enemies | [3,36,37,38,39,40] | |
| Cultural management | [10,11,19,21,41] | |
| Thresholds | [42] | |
| Based on soybean phenology | [43] | |
| Microbial Interaction | Identification and role of symbionts | [44,45,46] |
| Transfer of symbionts | [44] | |
| Host selection | [14,28,44,47,48] | |
| Evolution | [25,44] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dhammi, A.; Van Krestchmar, J.B.; Ponnusamy, L.; Bacheler, J.S.; Reisig, D.D.; Herbert, A.; Del Pozo-Valdivia, A.I.; Roe, R.M. Biology, Pest Status, Microbiome and Control of Kudzu Bug (Hemiptera: Heteroptera: Plataspidae): A New Invasive Pest in the U.S. Int. J. Mol. Sci. 2016, 17, 1570. https://doi.org/10.3390/ijms17091570
Dhammi A, Van Krestchmar JB, Ponnusamy L, Bacheler JS, Reisig DD, Herbert A, Del Pozo-Valdivia AI, Roe RM. Biology, Pest Status, Microbiome and Control of Kudzu Bug (Hemiptera: Heteroptera: Plataspidae): A New Invasive Pest in the U.S. International Journal of Molecular Sciences. 2016; 17(9):1570. https://doi.org/10.3390/ijms17091570
Chicago/Turabian StyleDhammi, Anirudh, Jaap B. Van Krestchmar, Loganathan Ponnusamy, Jack S. Bacheler, Dominic D. Reisig, Ames Herbert, Alejandro I. Del Pozo-Valdivia, and R. Michael Roe. 2016. "Biology, Pest Status, Microbiome and Control of Kudzu Bug (Hemiptera: Heteroptera: Plataspidae): A New Invasive Pest in the U.S." International Journal of Molecular Sciences 17, no. 9: 1570. https://doi.org/10.3390/ijms17091570
APA StyleDhammi, A., Van Krestchmar, J. B., Ponnusamy, L., Bacheler, J. S., Reisig, D. D., Herbert, A., Del Pozo-Valdivia, A. I., & Roe, R. M. (2016). Biology, Pest Status, Microbiome and Control of Kudzu Bug (Hemiptera: Heteroptera: Plataspidae): A New Invasive Pest in the U.S. International Journal of Molecular Sciences, 17(9), 1570. https://doi.org/10.3390/ijms17091570