
Downregulation of FOXP1 Inhibits Cell Proliferation in Hepatocellular Carcinoma by Inducing G1/S Phase Cell Cycle Arrest

 and
and
Abstract
:1. Introduction
2. Results
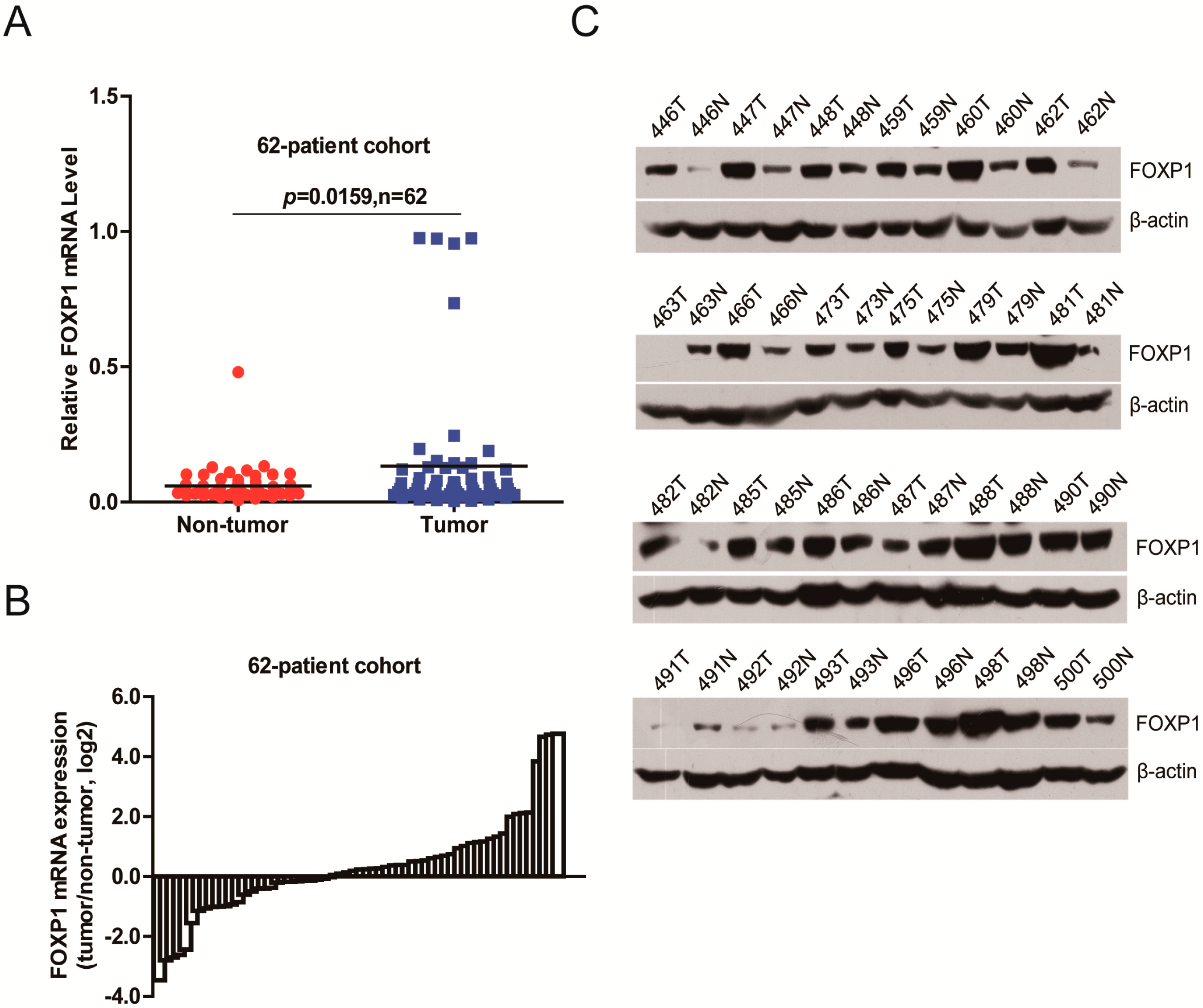
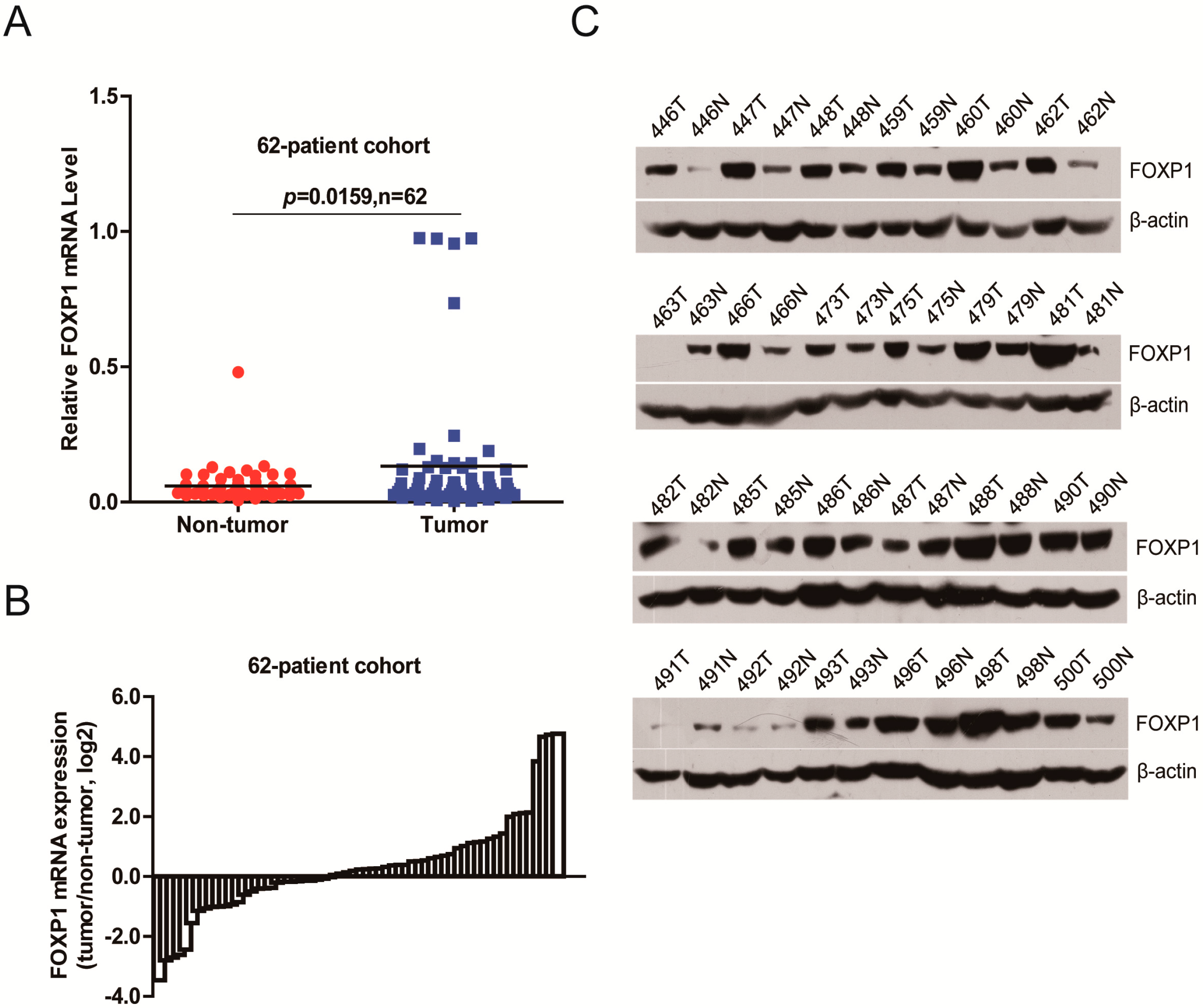
2.1. FOXP1 Is Frequently Upregulated in HCC Tissues
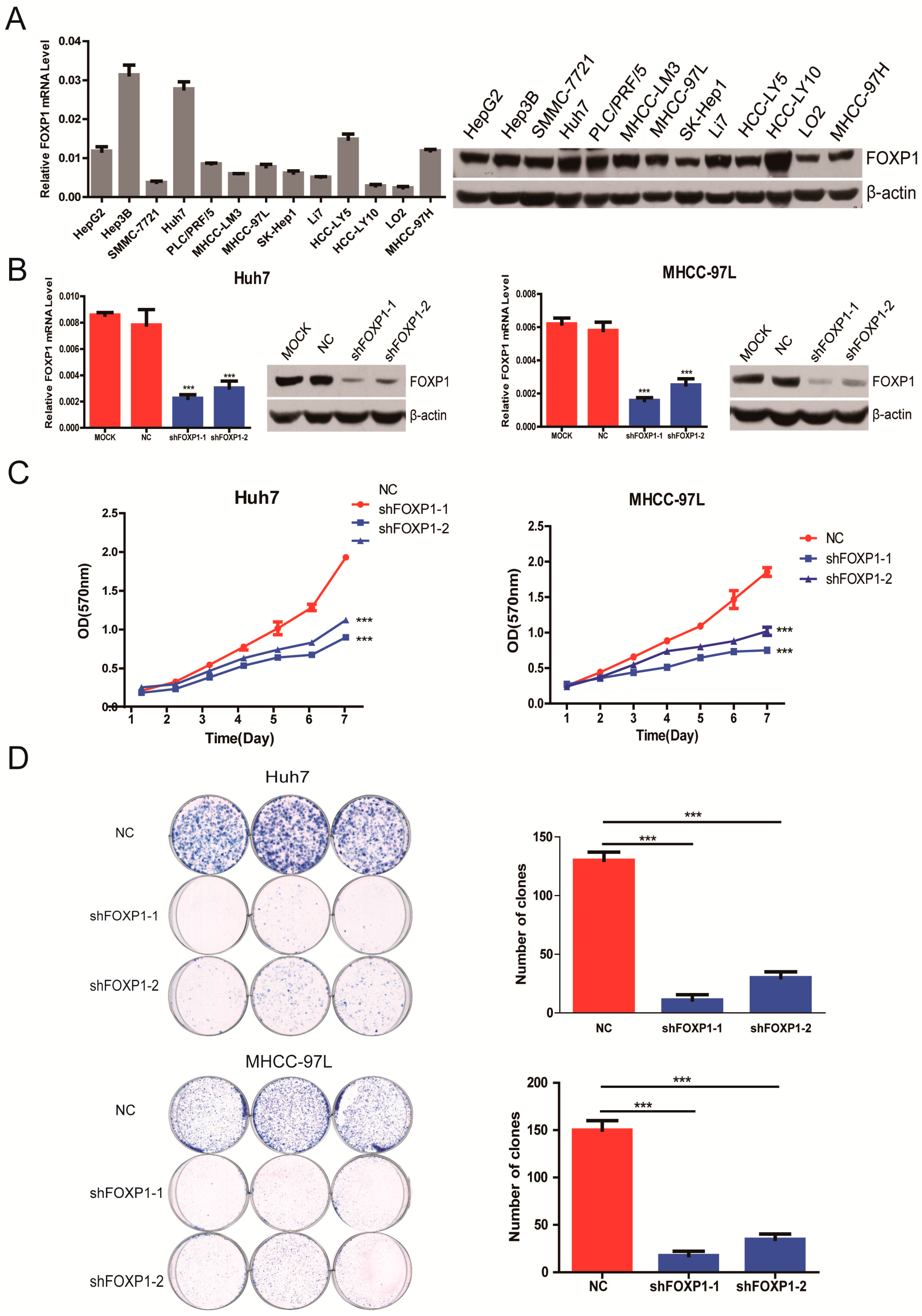
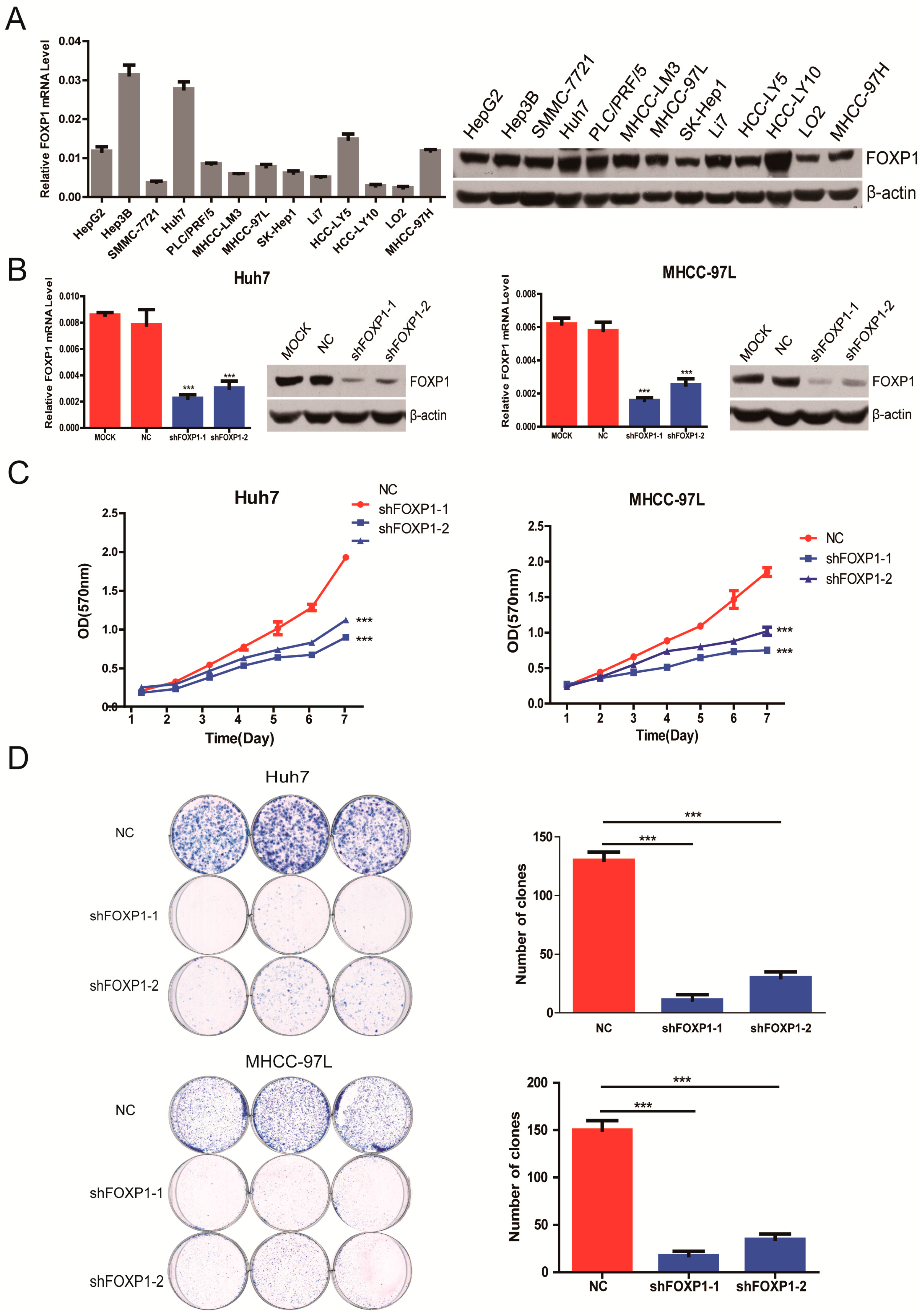
2.2. Silencing FOXP1 Inhibits the Growth Ability of HCC Cells in Vitro
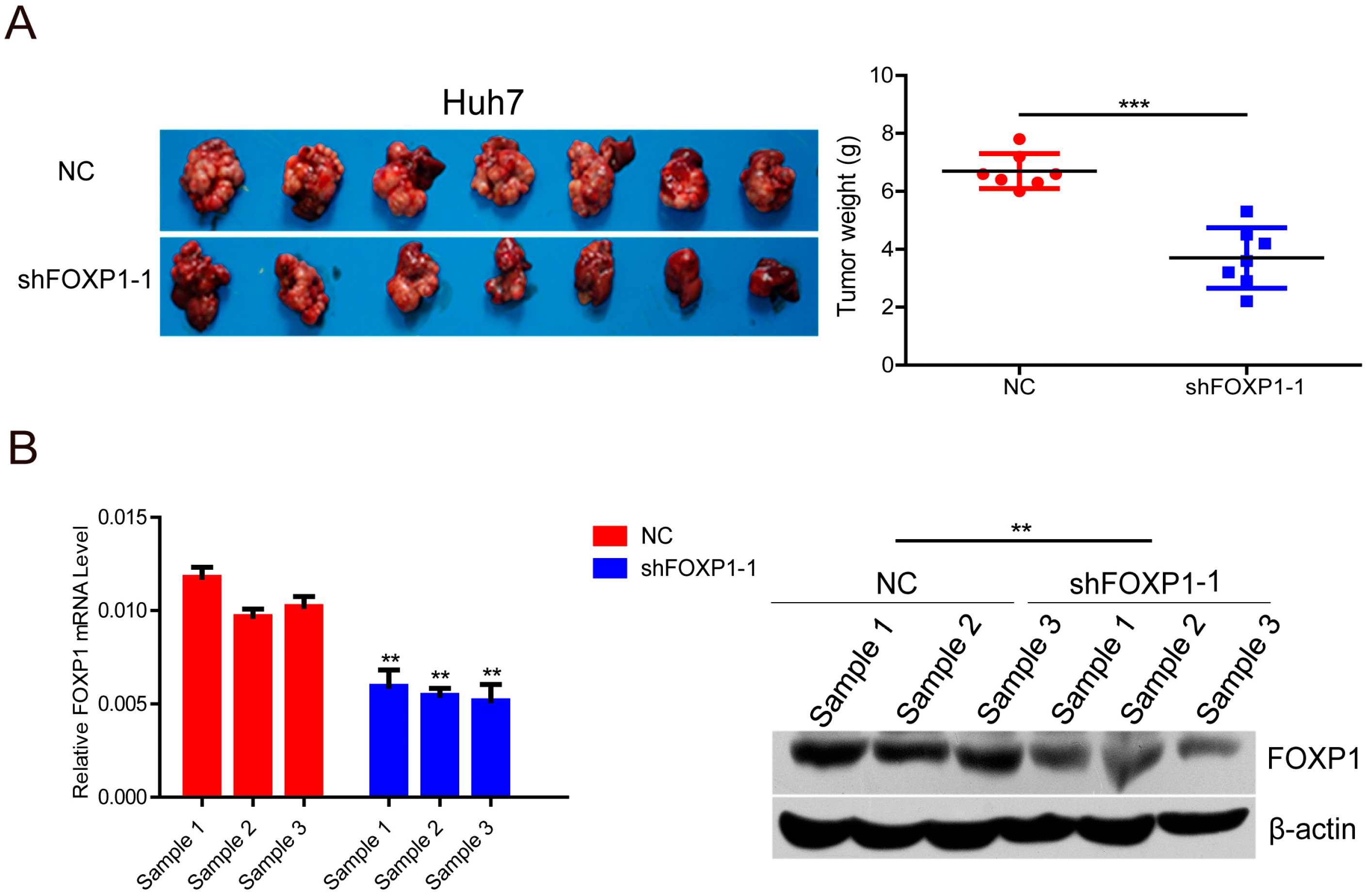
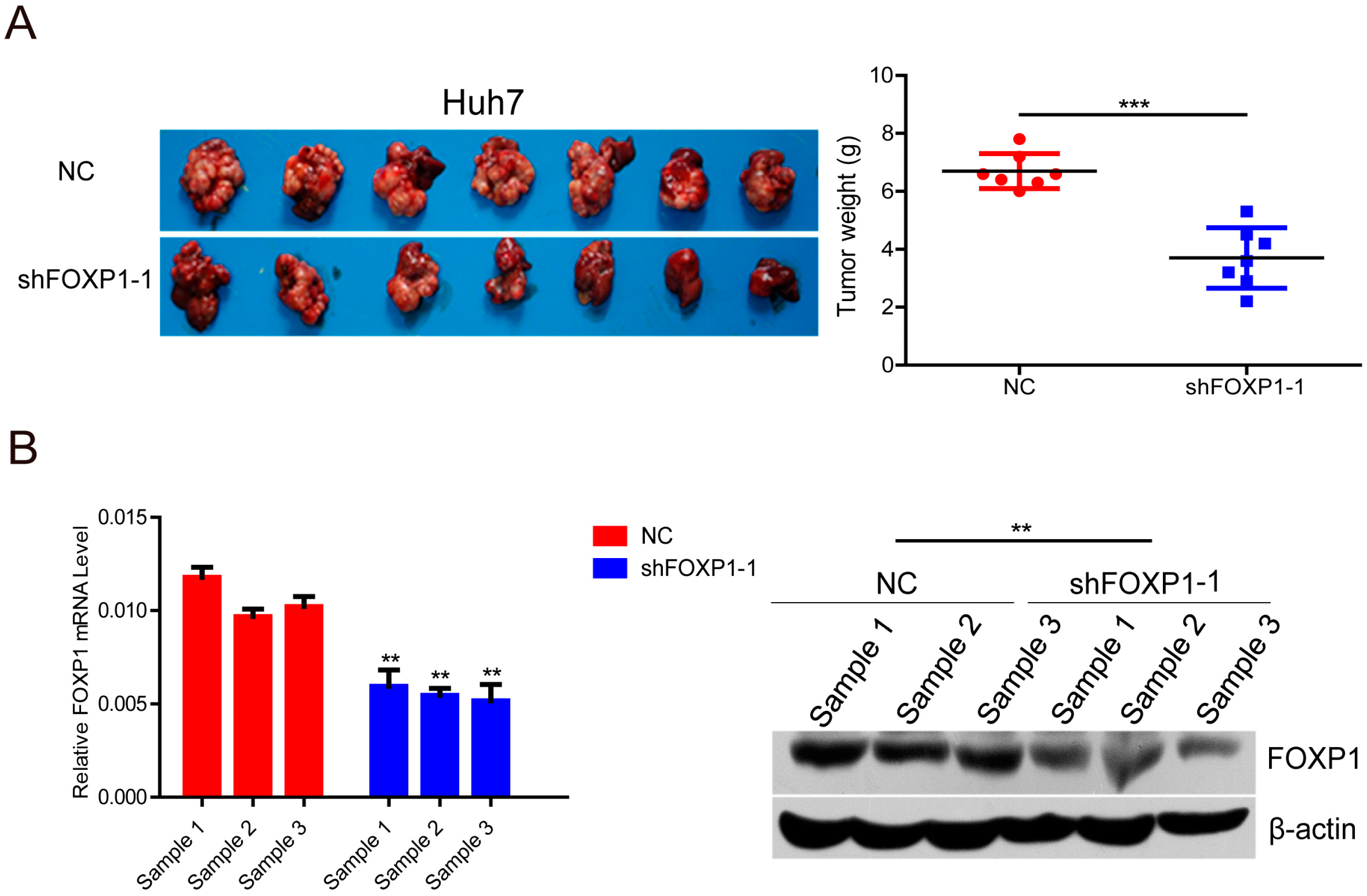
2.3. Knockdown of FOXP1 Decreases Tumorigenicity of HCC Cells in Vivo
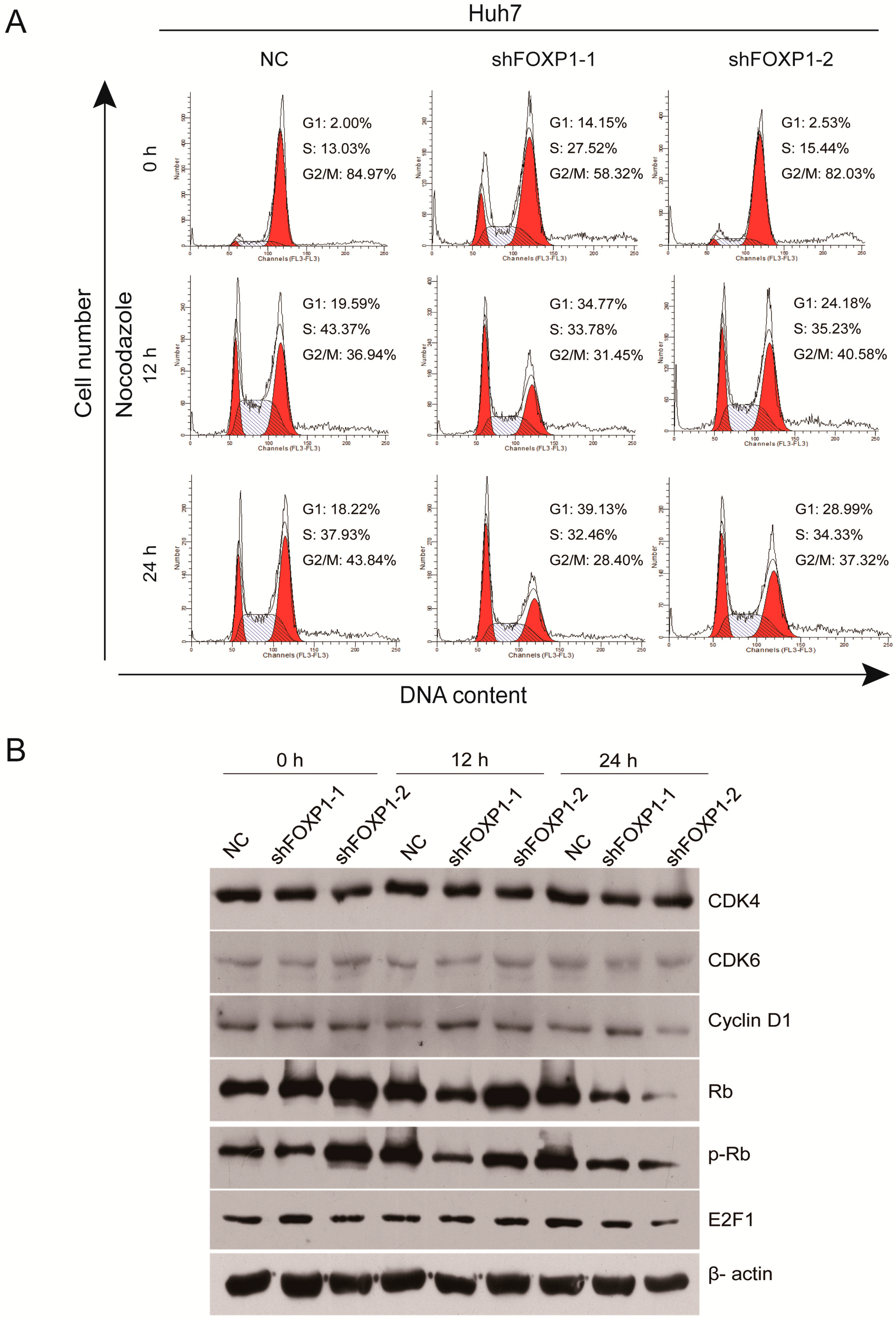
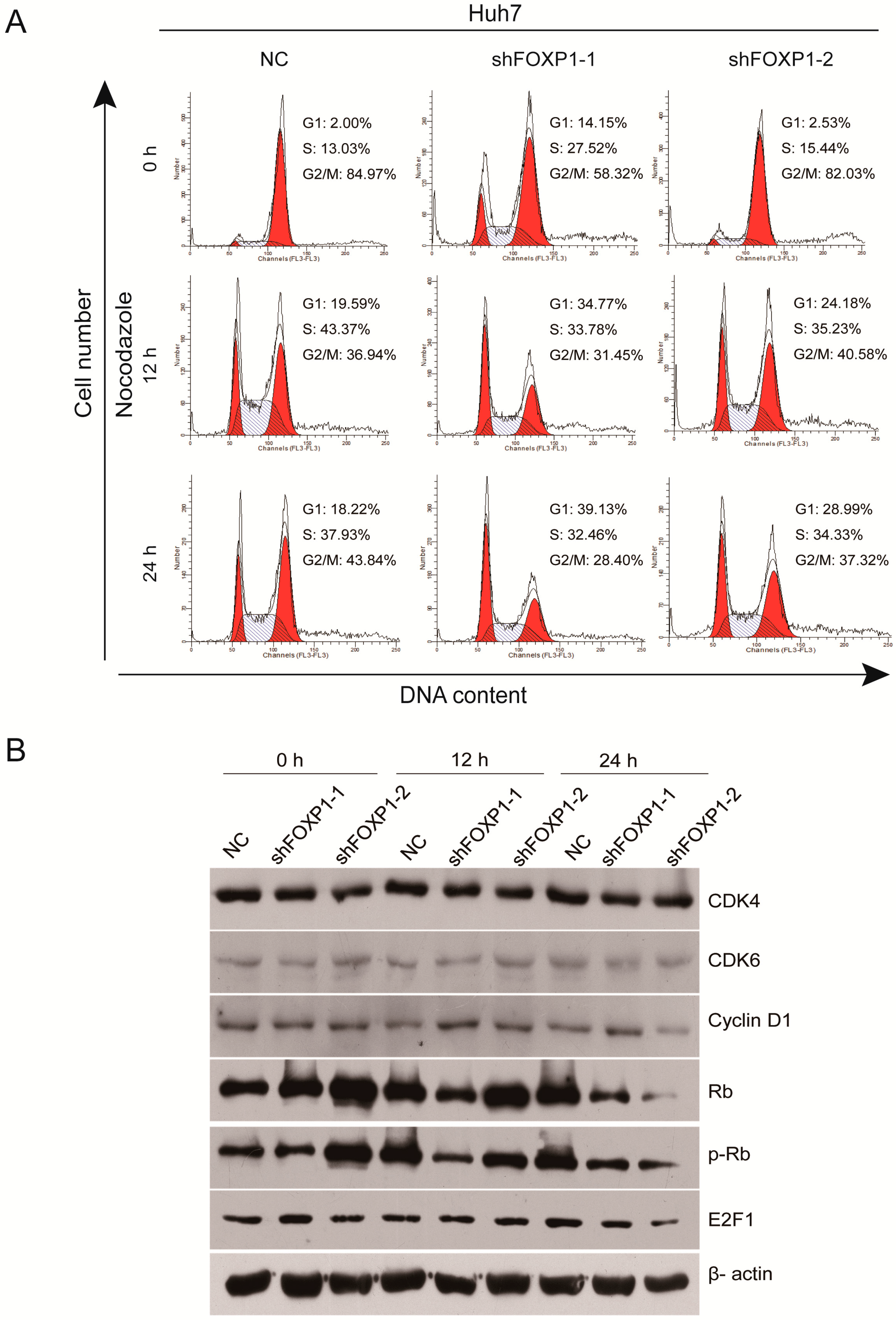
2.4. Downregulation of FOXP1 Induces G1/S Cycle Arrest and Regulates Cell Cycle-Related Proteins in HCC Cells
3. Discussion
4. Materials and Methods
4.1. Clinical Specimens and Expression Data Sets
4.2. Cell Lines and Cell Culture
4.3. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.4. Western Blot Analysis
4.5. RNA Interference-Mediated Gene Silencing
4.6. Colony Formation Assays
4.7. MTT Assays
4.8. In Vitro Wound Healing Assays
4.9. In Vitro Invasion Assays
4.10. Tumor Xenograft Models
4.11. Flow Cytometry Analysis of Cell Cycle
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Forner, A.; Llovet, J.M.; Bruix, J. Hepatocellular carcinoma. Lancet 2012, 379, 1245–1255. [Google Scholar] [CrossRef]
- Faivre, S.; Bouattour, M.; Raymond, E. Novel molecular therapies in hepatocellular carcinoma. Liver Int. 2011, 31, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Katoh, M.; Katoh, M. Human fox gene family (review). Int. J. Oncol. 2004, 25, 1495–1500. [Google Scholar] [CrossRef] [PubMed]
- Myatt, S.S.; Lam, E.W. The emerging roles of forkhead box (FOX) proteins in cancer. Nat. Rev. Cancer 2007, 7, 847–859. [Google Scholar] [CrossRef] [PubMed]
- Lam, E.W.; Brosens, J.J.; Gomes, A.R.; Koo, C.Y. Forkhead box proteins: Tuning forks for transcriptional harmony. Nat. Rev. Cancer 2013, 13, 482–495. [Google Scholar] [CrossRef] [PubMed]
- Shu, W.; Lu, M.M.; Zhang, Y.; Tucker, P.W.; Zhou, D.; Morrisey, E.E. FOXP2 and FOXP1 cooperatively regulate lung and esophagus development. Development 2007, 134, 1991–2000. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Weidenfeld, J.; Lu, M.M.; Maika, S.; Kuziel, W.A.; Morrisey, E.E.; Tucker, P.W. FOXP1 regulates cardiac outflow tract, endocardial cushion morphogenesis and myocyte proliferation and maturation. Development 2004, 131, 4477–4487. [Google Scholar] [CrossRef] [PubMed]
- Tamura, S.; Morikawa, Y.; Iwanishi, H.; Hisaoka, T.; Senba, E. Expression pattern of the winged-helix/forkhead transcription factor FOXP1 in the developing central nervous system. Gene Expr. Patterns 2003, 3, 193–197. [Google Scholar] [CrossRef]
- Hu, H.; Wang, B.; Borde, M.; Nardone, J.; Maika, S.; Allred, L.; Tucker, P.W.; Rao, A. FOXP1 is an essential transcriptional regulator of b cell development. Nat. Immunol. 2006, 7, 819–826. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Geng, J.; Wen, X.; Bi, E.; Kossenkov, A.V.; Wolf, A.I.; Tas, J.; Choi, Y.S.; Takata, H.; Day, T.J.; et al. The transcription factor FOXP1 is a critical negative regulator of the differentiation of follicular helper T cells. Nat. Immunol. 2014, 15, 667–675. [Google Scholar] [CrossRef] [PubMed]
- Streubel, B.; Vinatzer, U.; Lamprecht, A.; Raderer, M.; Chott, A. T(3; 14)(p14.1; q32) involving igh and FOXP1 is a novel recurrent chromosomal aberration in malt lymphoma. Leukemia 2005, 19, 652–658. [Google Scholar] [PubMed]
- Fenton, J.A.; Schuuring, E.; Barrans, S.L.; Banham, A.H.; Rollinson, S.J.; Morgan, G.J.; Jack, A.S.; van Krieken, J.H.; Kluin, P.M. T(3; 14)(p14; q32) results in aberrant expression of FOXP1 in a case of diffuse large B-cell lymphoma. Genes Chromosomes Cancer 2006, 45, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Barrans, S.L.; Fenton, J.A.; Banham, A.; Owen, R.G.; Jack, A.S. Strong expression of foxp1 identifies a distinct subset of diffuse large B-cell lymphoma (DLBCL) patients with poor outcome. Blood 2004, 104, 2933–2935. [Google Scholar] [CrossRef] [PubMed]
- Banham, A.H.; Connors, J.M.; Brown, P.J.; Cordell, J.L.; Ott, G.; Sreenivasan, G.; Farinha, P.; Horsman, D.E.; Gascoyne, R.D. Expression of the FOXP1 transcription factor is strongly associated with inferior survival in patients with diffuse large B-cell lymphoma. Clin. Cancer Res. 2005, 11, 1065–1072. [Google Scholar] [PubMed]
- Han, S.L.; Wu, X.L.; Wan, L.; Zeng, Q.Q.; Li, J.L.; Liu, Z. FOXP1 expression predicts polymorphic histology and poor prognosis in gastric mucosa-associated lymphoid tissue lymphomas. Dig. Surg. 2009, 26, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Yue, S.; Zhang, J. FOXP1 has a low expression in human gliomas and its overexpression inhibits proliferation, invasion and migration of human glioma U251 cells. Mol. Med. Rep. 2014, 10, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Takayama, K.; Suzuki, T.; Tsutsumi, S.; Fujimura, T.; Takahashi, S.; Homma, Y.; Urano, T.; Aburatani, H.; Inoue, S. Integrative analysis of FOXP1 function reveals a tumor-suppressive effect in prostate cancer. Mol. Endocrinol. 2014, 28, 2012–2024. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.; Huang, J.; Chen, J.; Wang, R.; Dong, C.; Lu, S.; Wu, X. Expression and significance of FOXP1, HIF-1A and VEGF in renal clear cell carcinoma. J. BUON 2015, 20, 188–195. [Google Scholar] [PubMed]
- Rayoo, M.; Yan, M.; Takano, E.A.; Bates, G.J.; Brown, P.J.; Banham, A.H.; Fox, S.B. Expression of the forkhead box transcription factor FOXP1 is associated with oestrogen receptor alpha, oestrogen receptor beta and improved survival in familial breast cancers. J. Clin. Pathol. 2009, 62, 896–902. [Google Scholar] [CrossRef] [PubMed]
- Datta, J.; Kutay, H.; Nasser, M.W.; Nuovo, G.J.; Wang, B.; Majumder, S.; Liu, C.G.; Volinia, S.; Croce, C.M.; Schmittgen, T.D.; et al. Methylation mediated silencing of microRNA-1 gene and its role in hepatocellular carcinogenesis. Cancer Res. 2008, 68, 5049–5058. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, S.; Wang, X.; Liu, J.; Yang, L.; He, S.; Chen, L.; Huang, J. Prognostic significance of FOXP1 as an oncogene in hepatocellular carcinoma. J. Clin. Pathol. 2012, 65, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Xu, Y.; Li, X.; Wu, Y.; Zhou, J.; Wang, G.; Chen, L. Effects of lentiviral-mediated FOXP1 and FOXQ1 rnai on the hepatocarcinoma cell. Exp. Mol. Pathol. 2014, 96, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Jordan, M.A.; Thrower, D.; Wilson, L. Effects of vinblastine, podophyllotoxin and nocodazole on mitotic spindles. Implications for the role of microtubule dynamics in mitosis. J. Cell Sci. 1992, 102, 401–416. [Google Scholar] [PubMed]
- De Brabander, M.J.; van de Veire, R.M.; Aerts, F.E.; Borgers, M.; Janssen, P.A. The effects of methyl (5-(2-thienylcarbonyl)-1H-benzimidazol-2-yl) carbamate, (R 17934; NSC 238159), a new synthetic antitumoral drug interfering with microtubules, on mammalian cells cultured in vitro. Cancer Res. 1976, 36, 905–916. [Google Scholar] [PubMed]
- Banham, A.H.; Beasley, N.; Campo, E.; Fernandez, P.L.; Fidler, C.; Gatter, K.; Jones, M.; Mason, D.Y.; Prime, J.E.; Trougouboff, P.; et al. The FOXP1 winged helix transcription factor is a novel candidate tumor suppressor gene on chromosome 3p. Cancer Res. 2001, 61, 8820–8829. [Google Scholar] [PubMed]
- Feng, J.; Zhang, X.; Zhu, H.; Wang, X.; Ni, S.; Huang, J. High expression of FOXP1 is associated with improved survival in patients with non-small cell lung cancer. Am. J. Clin. Pathol. 2012, 138, 230–235. [Google Scholar] [CrossRef] [PubMed]
- De Smedt, L.; Palmans, S.; Govaere, O.; Moisse, M.; Boeckx, B.; de Hertogh, G.; Prenen, H.; van Cutsem, E.; Tejpar, S.; Tousseyn, T.; et al. Expression of FOXP1 and colorectal cancer prognosis. Lab. Med. 2015, 46, 299–311. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Liu, T.; Xiao, C.Y.; Yu, J.X.; Lu, L.G.; Xu, M.Y. FOXP1 and SPINK1 reflect the risk of cirrhosis progression to HCC with HBV infection. Biomed. Pharmacother. 2015, 72, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Leishman, E.; Howard, J.M.; Garcia, G.E.; Miao, Q.; Ku, A.T.; Dekker, J.D.; Tucker, H.; Nguyen, H. FOXP1 maintains hair follicle stem cell quiescence through regulation of FGF18. Development 2013, 140, 3809–3818. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Wang, H.; Takata, H.; Day, T.J.; Willen, J.; Hu, H. Transcription factor FOXP1 exerts essential cell-intrinsic regulation of the quiescence of naive T cells. Nat. Immunol. 2011, 12, 544–550. [Google Scholar] [CrossRef] [PubMed]
- Stewart, Z.A.; Westfall, M.D.; Pietenpol, J.A. Cell-cycle dysregulation and anticancer therapy. Trends Pharmacol. Sci. 2003, 24, 139–145. [Google Scholar] [CrossRef]
- Ackermann, S.; Kocak, H.; Hero, B.; Ehemann, V.; Kahlert, Y.; Oberthuer, A.; Roels, F.; Theissen, J.; Odenthal, M.; Berthold, F.; et al. FOXP1 inhibits cell growth and attenuates tumorigenicity of neuroblastoma. BMC Cancer 2014, 14. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time | Cell Cycle | NC# (%) shFOXP1-1 (%) shFOXP1-2 (%) | ||
|---|---|---|---|---|
| 0 h | G1 | 2.22 ± 0.21 | 14.7 ± 0.57 *** | 2.02 ± 0.44 |
| S | 14.55 ± 1.55 | 27.64 ± 0.40 ** | 15.85 ± 0.86 | |
| G2/M | 83.22 ± 1.68 | 57.65 ± 0.90 ** | 82.13 ± 0.75 | |
| 12 h | G1 | 21.96 ± 2.97 | 34.70 ± 0.91 * | 24.02 ± 1.14 |
| S | 43.52 ± 2.18 | 35.21 ± 1.25 ** | 36.92 ± 1.53 * | |
| G2/M | 34.49 ± 2.12 | 30.08 ± 1.56 * | 39.06 ± 2.07 | |
| 24 h | G1 | 20.37 ± 1.86 | 37.73 ± 1.78 *** | 28.3 ± 0.75 ** |
| S | 37.55 ± 1.88 | 32.28 ± 0.41 * | 35.16 ± 1.07 | |
| G2/M | 42.07 ± 2.42 | 29.99 ± 1.65 ** | 36.76 ± 0.59 | |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Sun, J.; Cui, M.; Zhao, F.; Ge, C.; Chen, T.; Yao, M.; Li, J. Downregulation of FOXP1 Inhibits Cell Proliferation in Hepatocellular Carcinoma by Inducing G1/S Phase Cell Cycle Arrest. Int. J. Mol. Sci. 2016, 17, 1501. https://doi.org/10.3390/ijms17091501
Wang X, Sun J, Cui M, Zhao F, Ge C, Chen T, Yao M, Li J. Downregulation of FOXP1 Inhibits Cell Proliferation in Hepatocellular Carcinoma by Inducing G1/S Phase Cell Cycle Arrest. International Journal of Molecular Sciences. 2016; 17(9):1501. https://doi.org/10.3390/ijms17091501
Chicago/Turabian StyleWang, Xin, Ji Sun, Meiling Cui, Fangyu Zhao, Chao Ge, Taoyang Chen, Ming Yao, and Jinjun Li. 2016. "Downregulation of FOXP1 Inhibits Cell Proliferation in Hepatocellular Carcinoma by Inducing G1/S Phase Cell Cycle Arrest" International Journal of Molecular Sciences 17, no. 9: 1501. https://doi.org/10.3390/ijms17091501
APA StyleWang, X., Sun, J., Cui, M., Zhao, F., Ge, C., Chen, T., Yao, M., & Li, J. (2016). Downregulation of FOXP1 Inhibits Cell Proliferation in Hepatocellular Carcinoma by Inducing G1/S Phase Cell Cycle Arrest. International Journal of Molecular Sciences, 17(9), 1501. https://doi.org/10.3390/ijms17091501