
Influences of Electromagnetic Energy on Bio-Energy Transport through Protein Molecules in Living Systems and Its Experimental Evidence

Abstract
:1. Introduction
2. Theories of Energy Transport in Protein Molecules and the Mechanism of Influence of EFs on Transport
2.1. Davydov’s Theory of Energy Transport and Its Features
2.2. Pang’s Theory of Energy Transport and Its Properties
2.3. Mechanism of Influence of Electric Field on Energy Transport in Protein Molecules
3. Variation of Properties of Energy Transport Arising from the EF in Protein Molecules
3.1. Analytic Results for Changes of Properties of the Soliton Transporting the Energy under the Influence of EFs
- (1)
- The amplitude () effective mass (Msol), energy (E), rest energy (E0), and binding energy EBP of the soliton are decreased and depressed after EF is applied because the physical parameters in Equations (11)–(14) are inversely proportional to the dipole–dipole interactional energy J. This means that the capability and value of the energy transported by the soliton is decreased because of the increases of dipole–dipole interaction between neighboring amino acid residues. If the strength of an EF in an EMF is very high, then the capability and value of the energy transport will depress considerably. In such a case, we could infer and suppose that some new biological effects will occur and that these new biological effects have just arisen from the EF
- (2)
- The EF varies the form and outline of the soliton in Equation (10) because the form and outline of the solitary wave, , in Equation (10) and its amplitude of envelope, , as well as its phase of the carrier wave, , are all changed with the variation of dipole–dipole interaction J under the influenceof EF. These changes arising from EFs will also affect the proliferation of cells and life bodies because of variations in the bio-energy they obtain.
- (3)
- The biological effects of EFs closely depend on both strength and direction with respect to the dipole moment of amino acid residues because the externally applied electric-field, and dipole moments of the amino acid residue, , all possess a certain strength and direction. In this case, we should consider the direction and strength of the EF, , and the dipole moments of the amino acid residue as well as their relationships. Thus, the electromagnetic energy of interaction between them should be represented by , where θ is the angle between the two vectors, and . This implies that the variations of the dipole–dipole interaction between neighboring amino acid residues caused by the EF should be expressed by , which decreases when the angle (θ) increases. If θ = 0, then . If , then . Therefore, the EF has a stronger biological effect on the former and no biological effect on the latter. This result indicates clearly that the biological effect depends on the direction of externally applied EF with respect to that of electric dipole moments of the amino acid residues. If the directions of EF are different, although their strengths are the same, then their biological effects are also different. This finding implies that the biological effects of EFs depend on both the strength and the direction with respect of the dipole moment of amino acid residues [81,86,97].
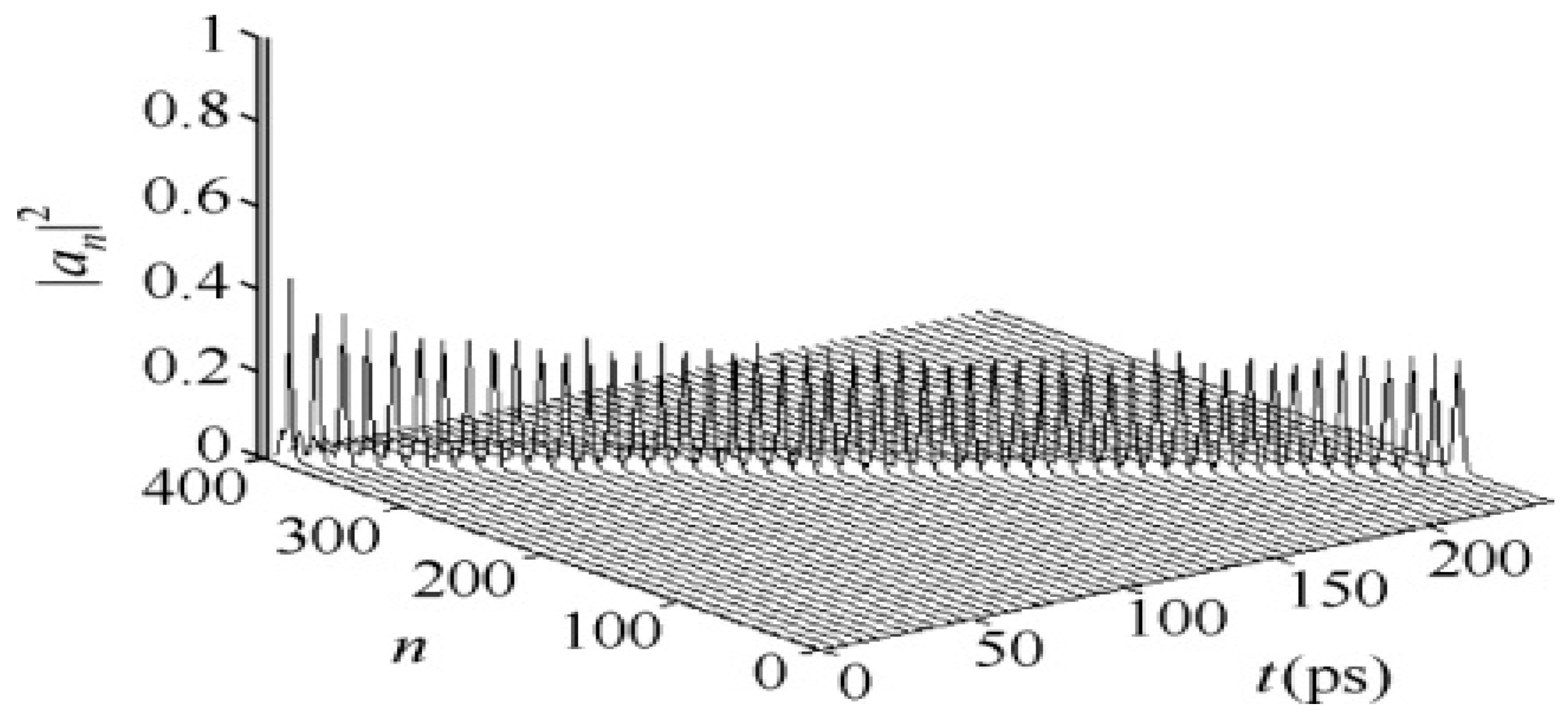
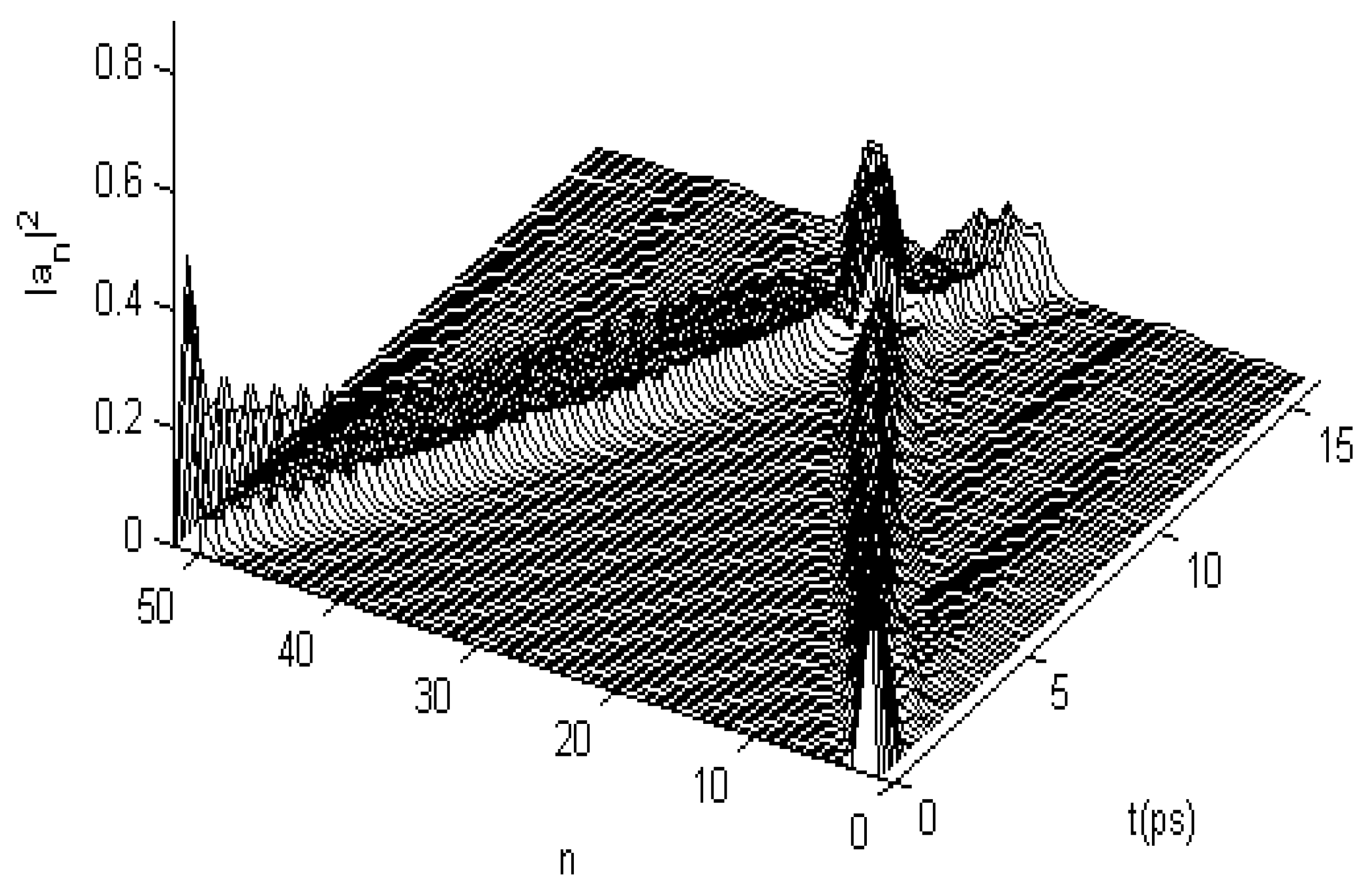
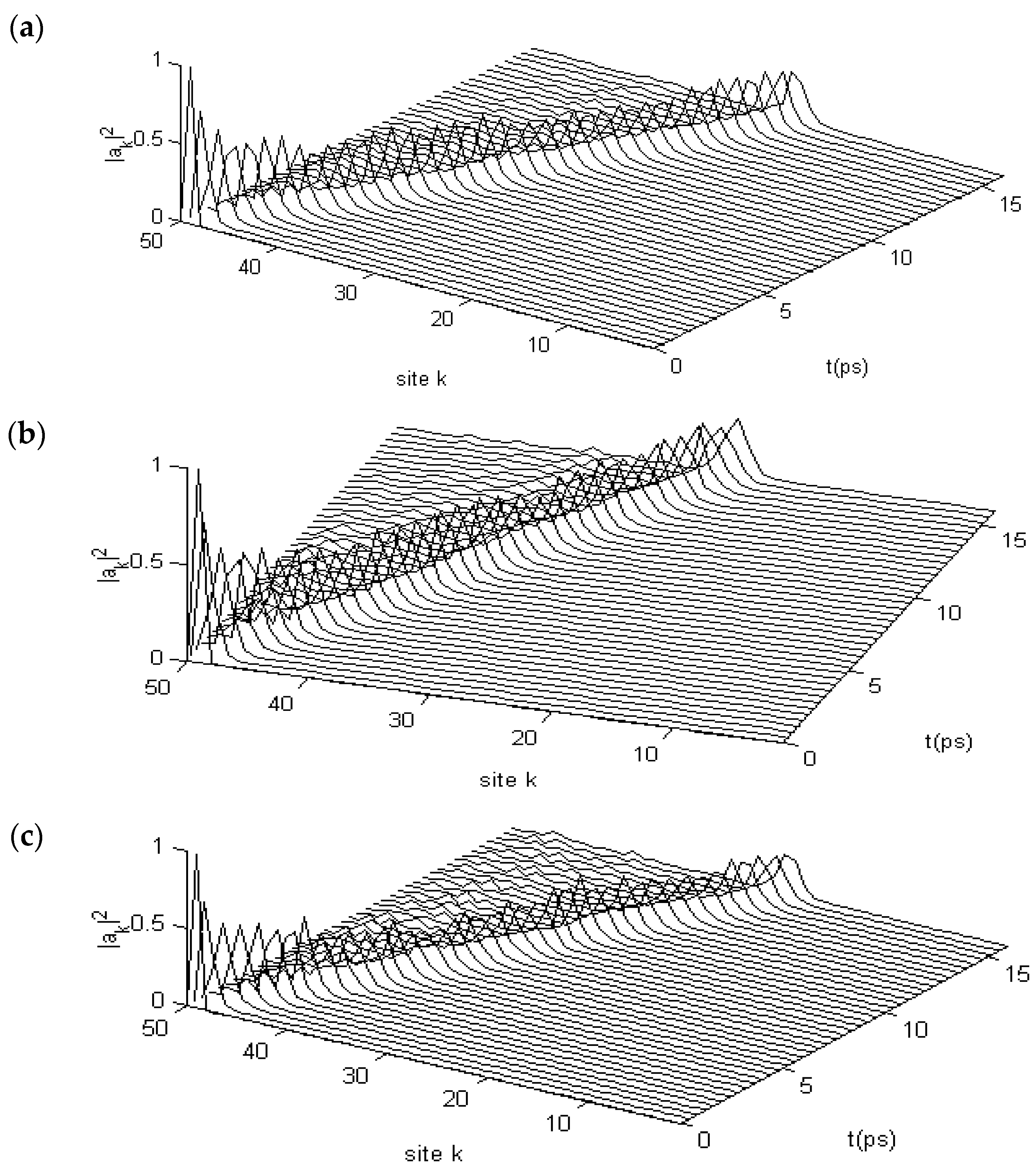
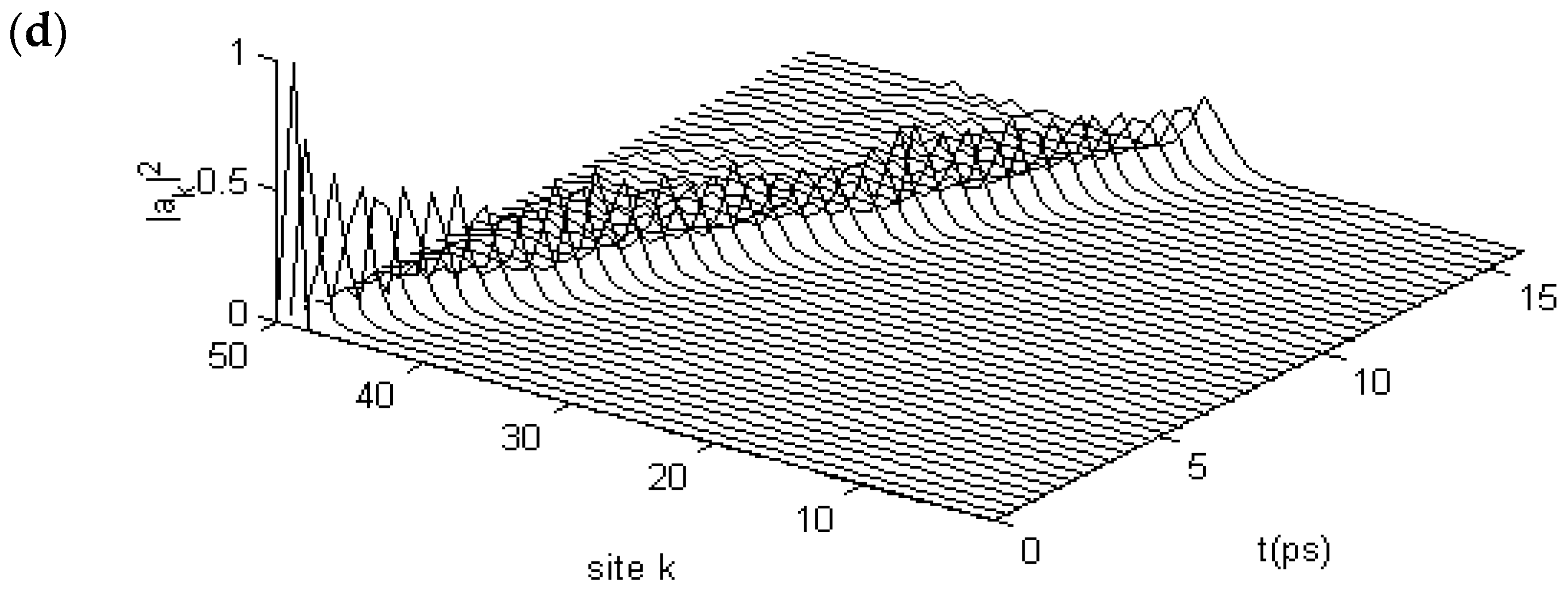
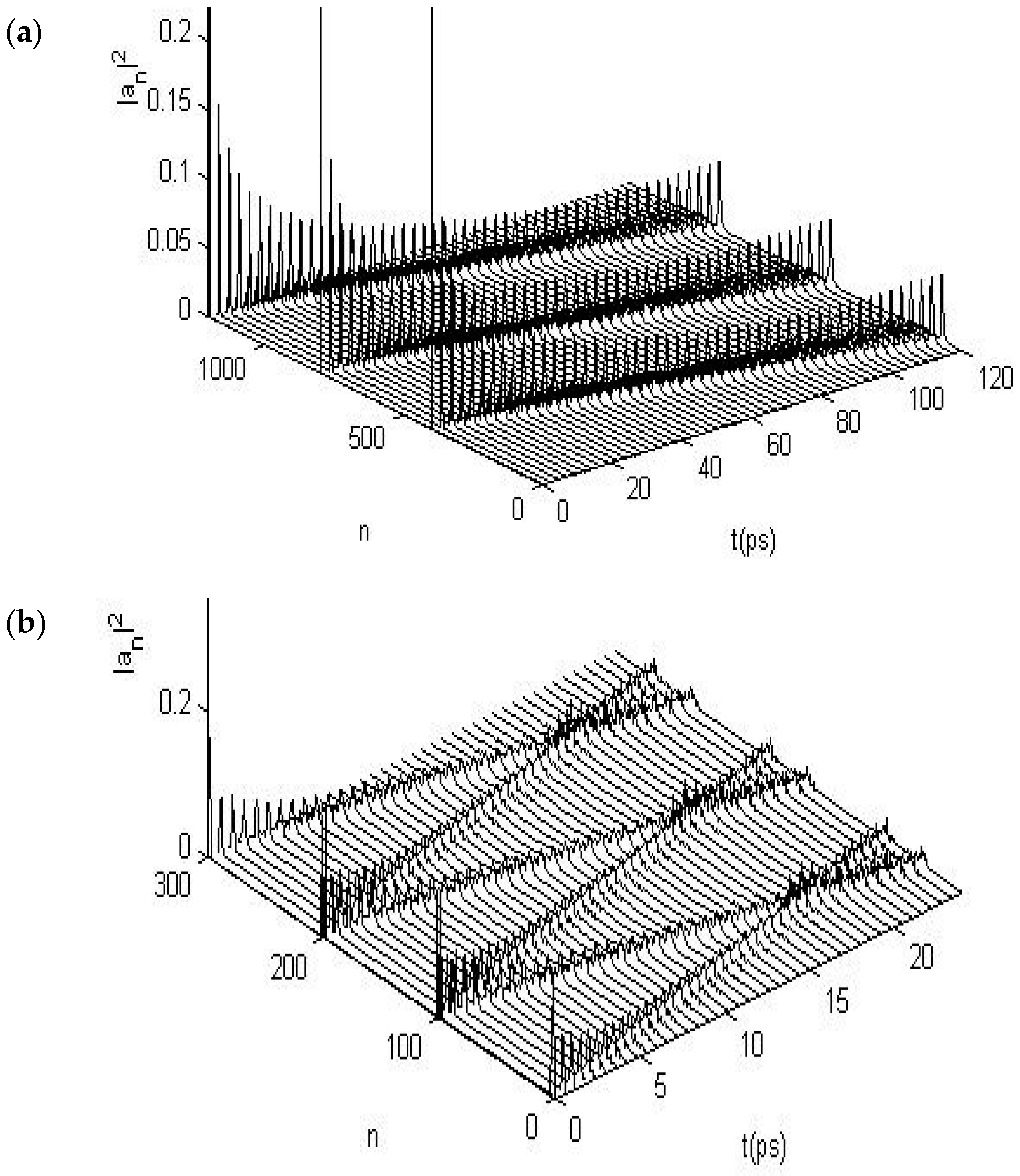
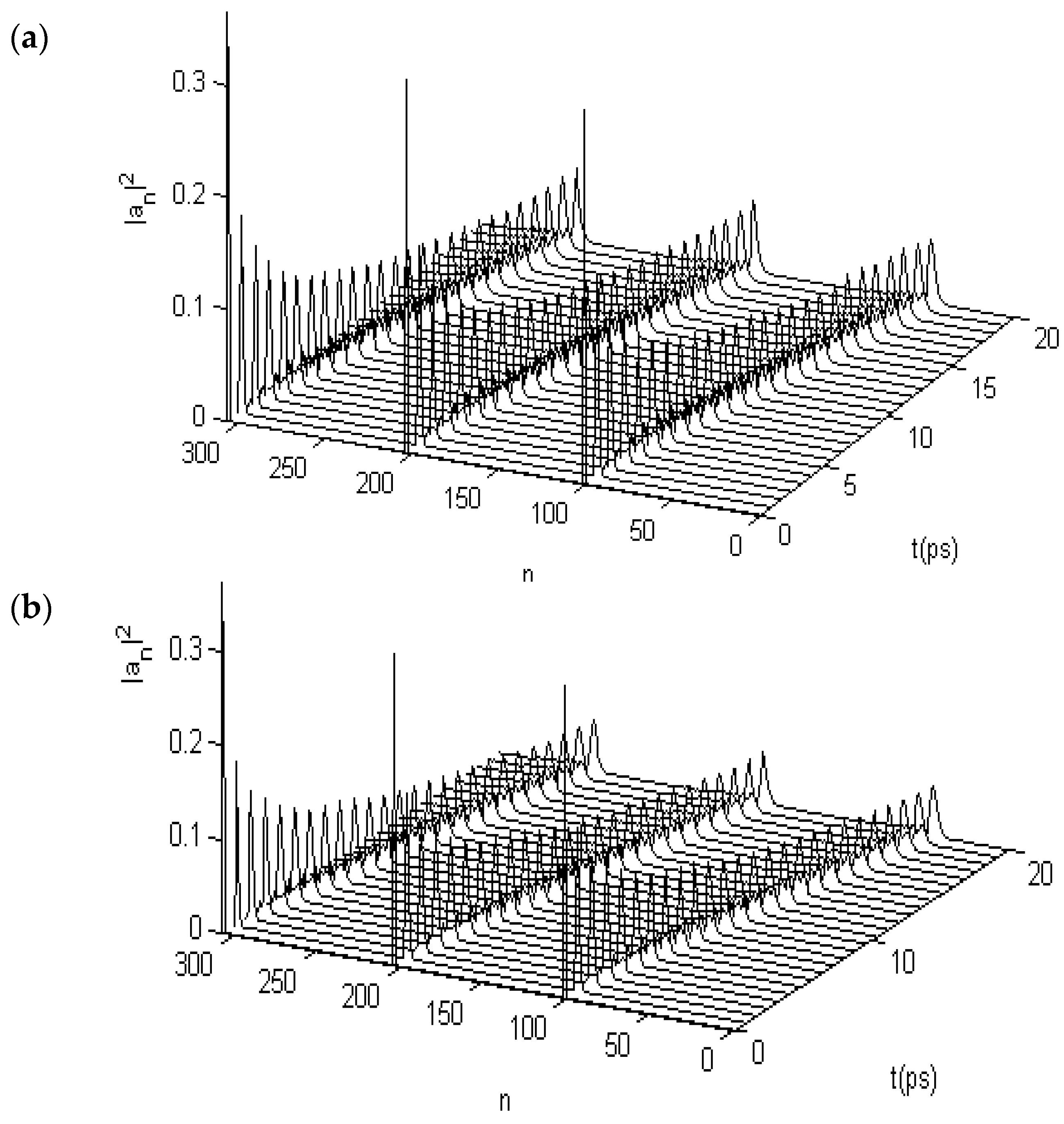
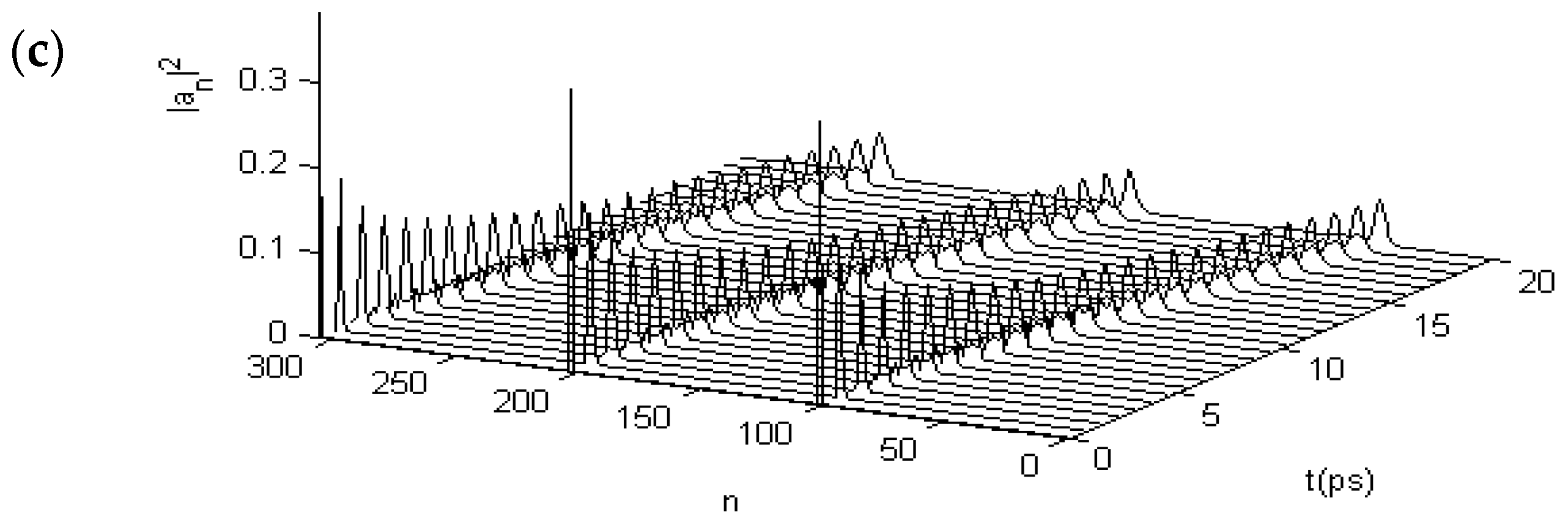
3.2. Results of Numerical Simulation for Changes of Property of the Energy Transport Resulting from EF
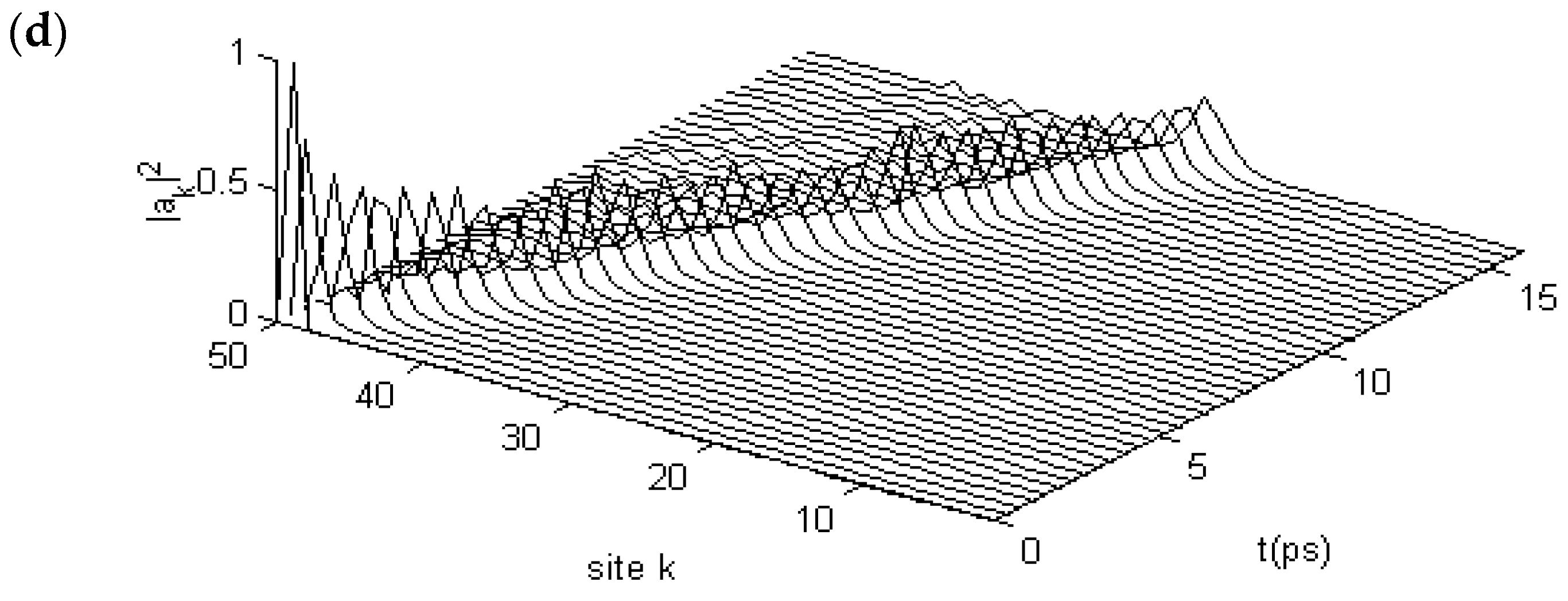
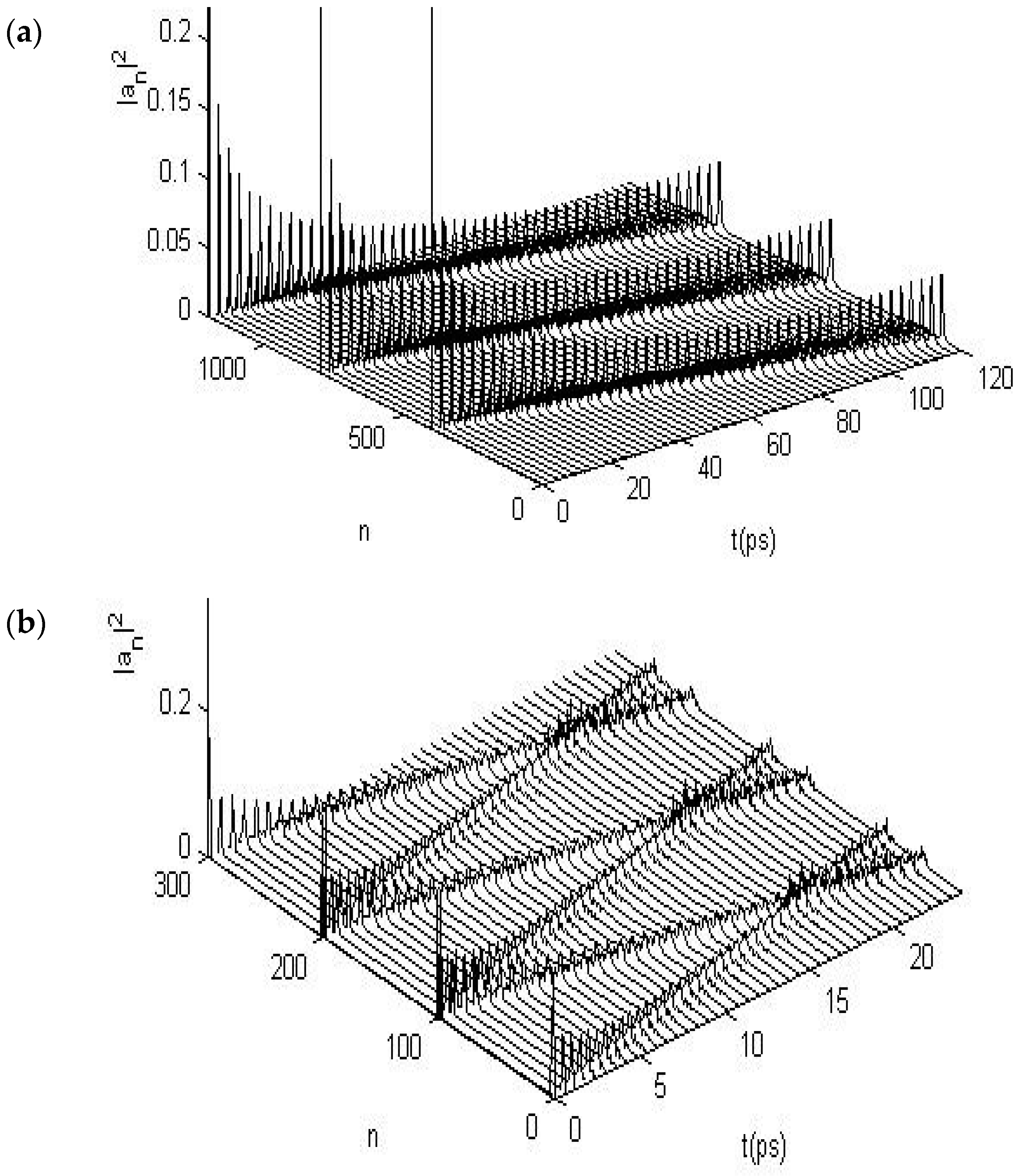
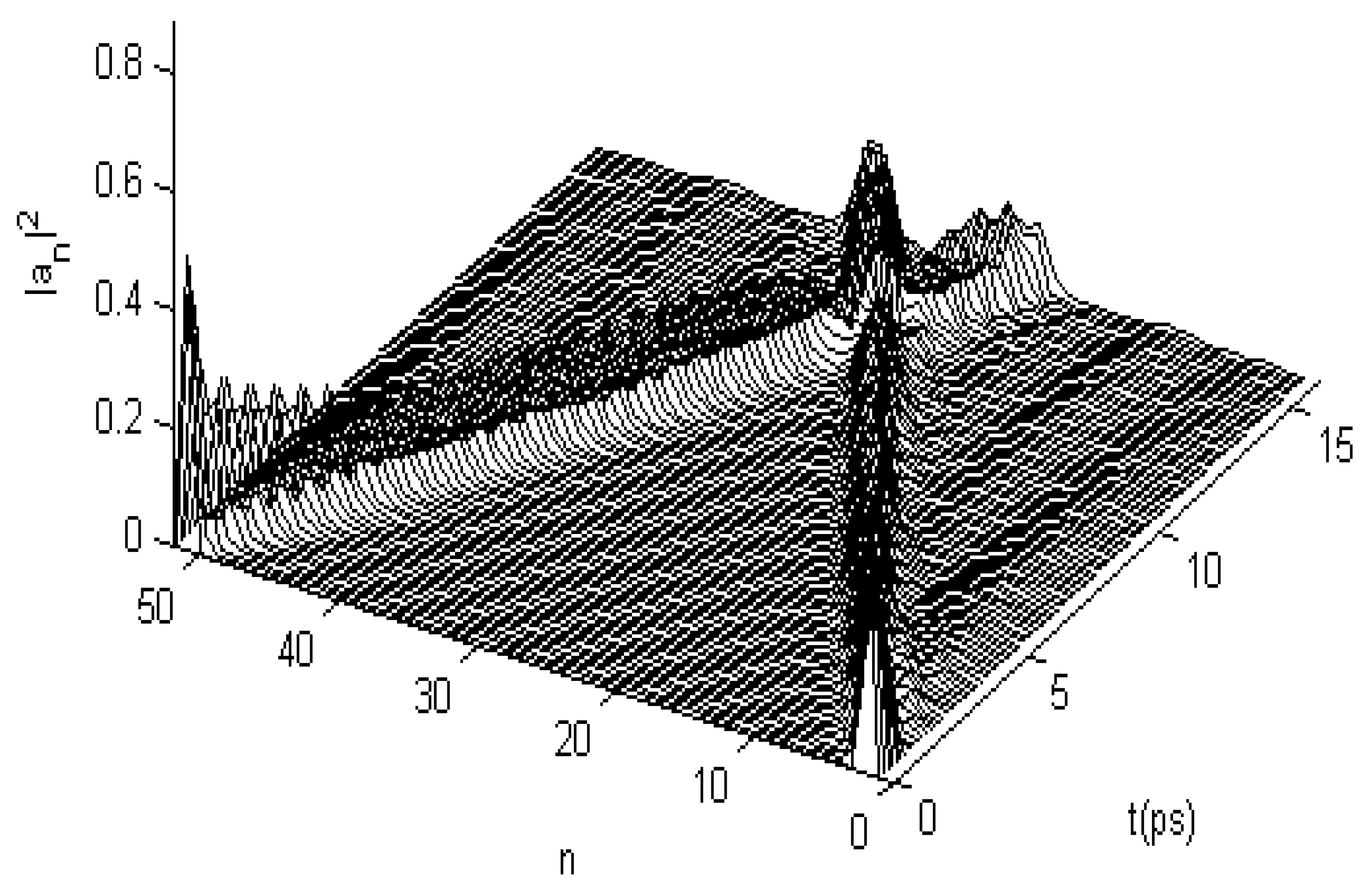
3.2.1. Results in Single-Protein Chains
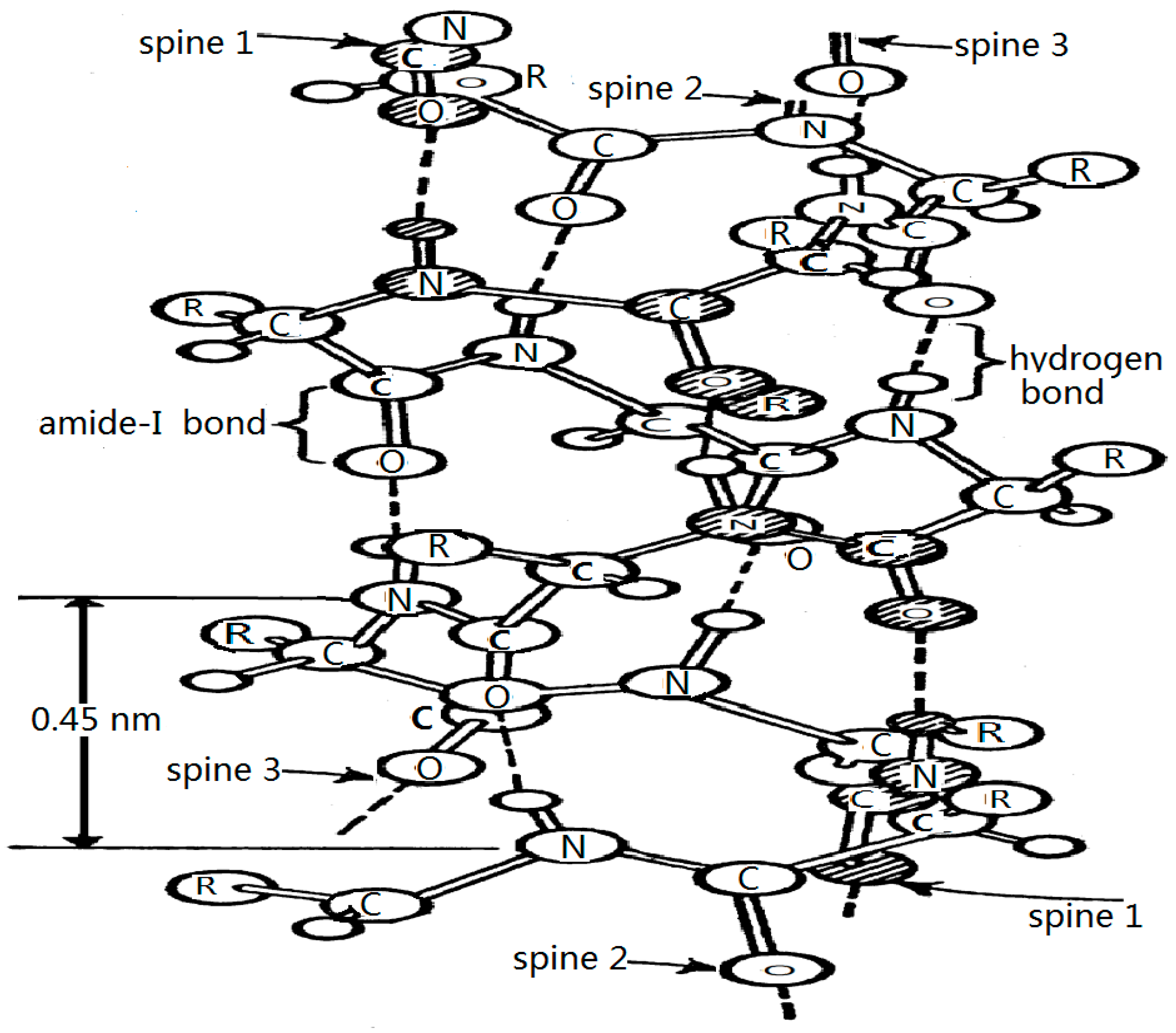
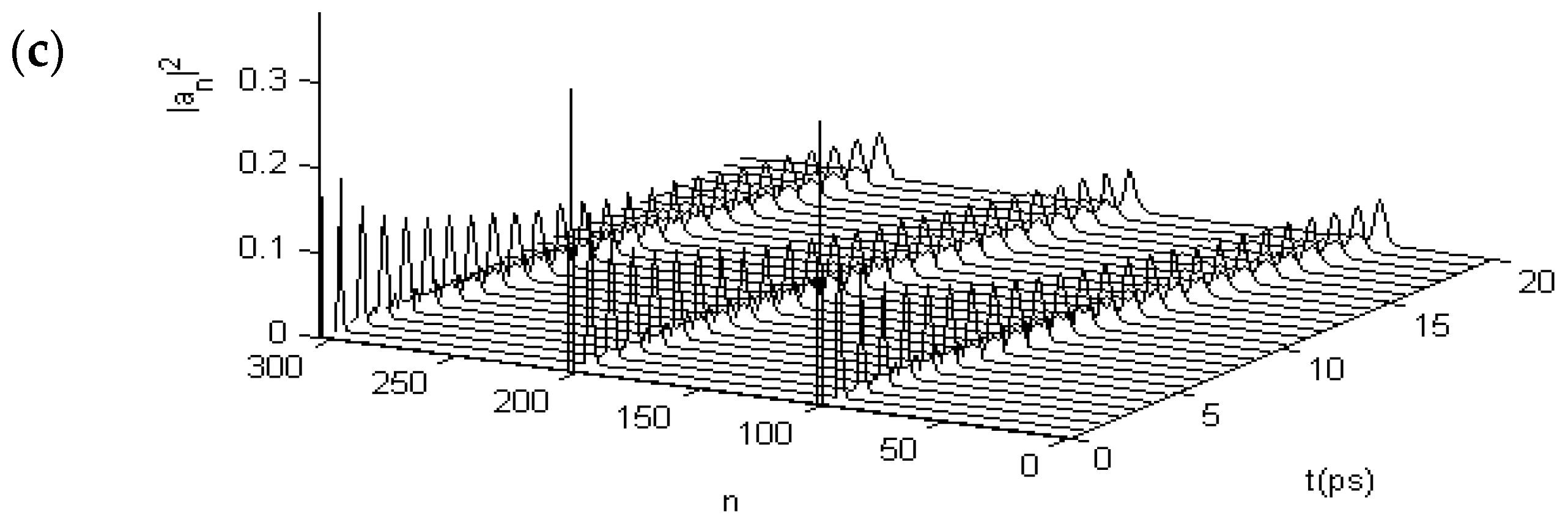
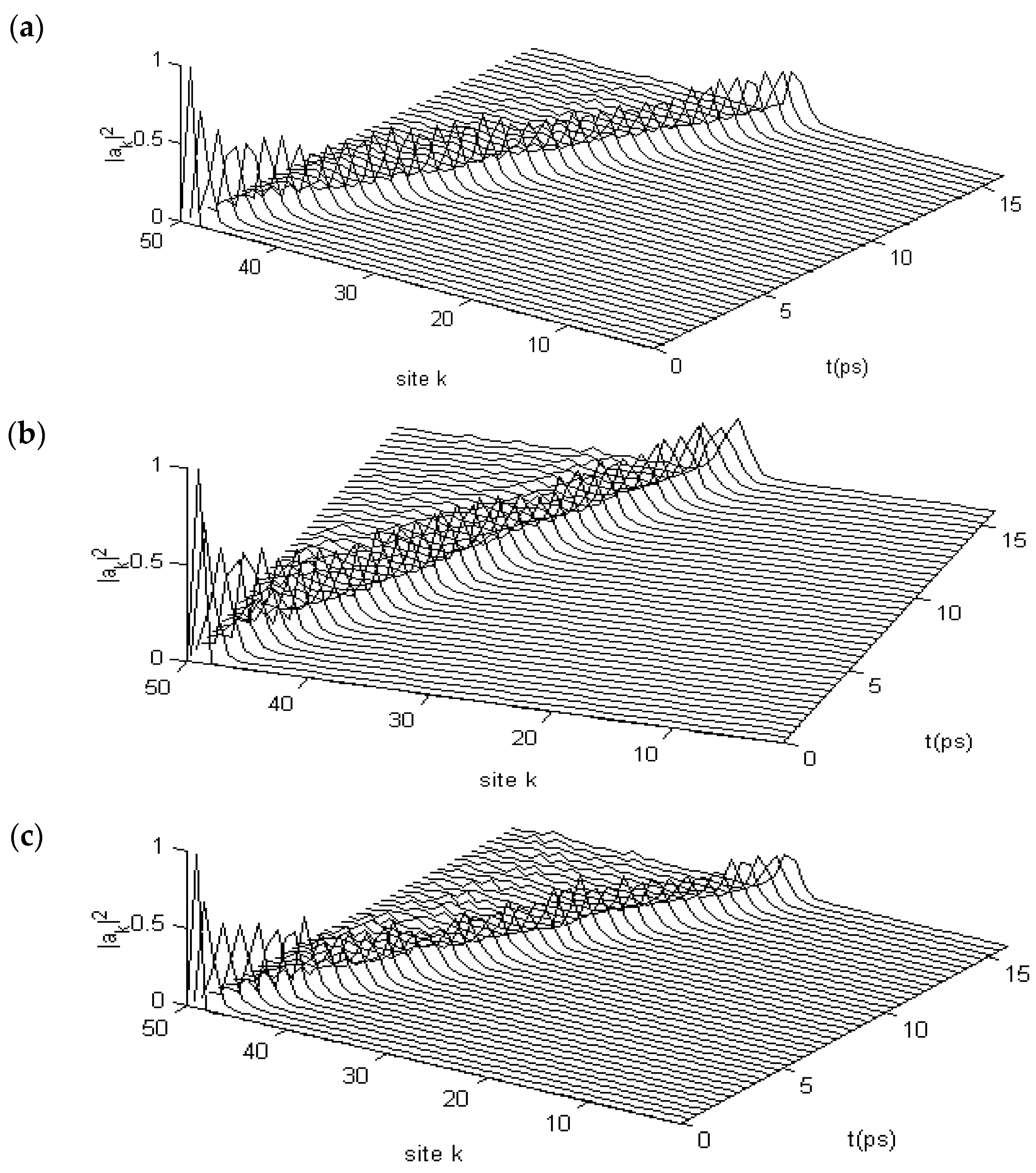
3.2.2. Results in α-Helix Protein Molecules with Three Channels
4. Experimental Evidences for this Theory
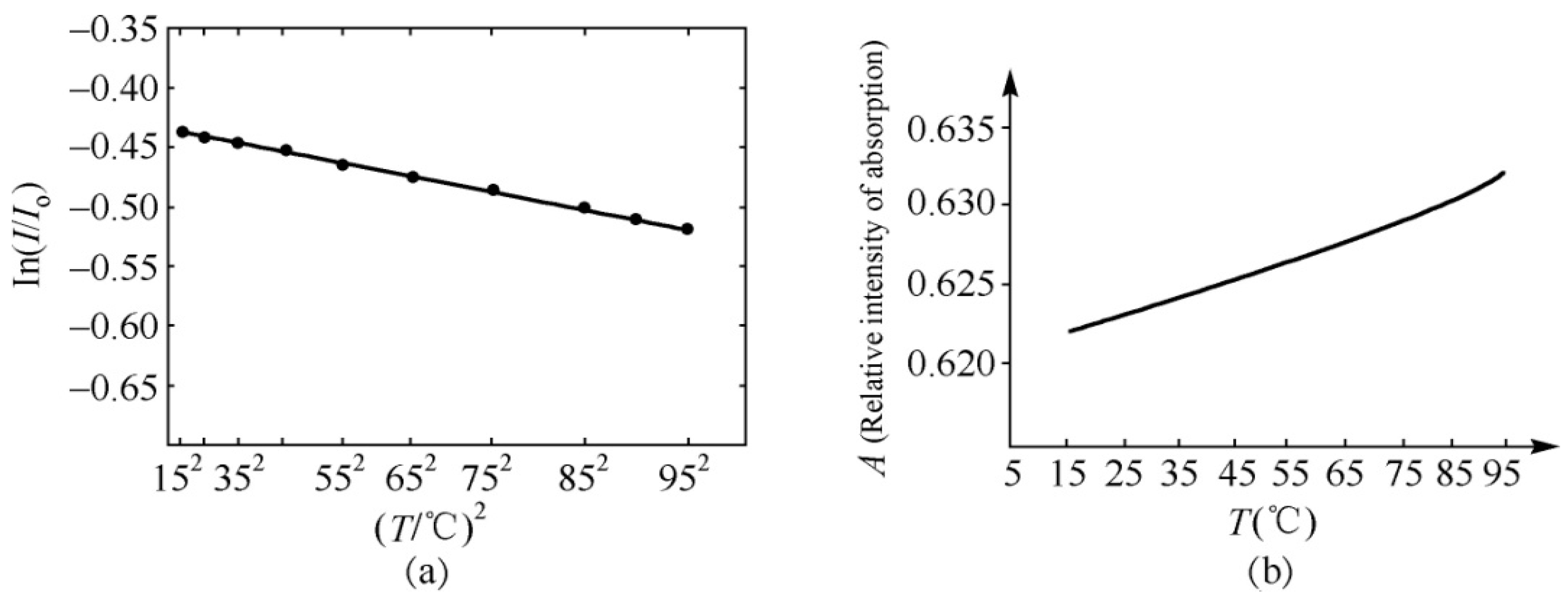
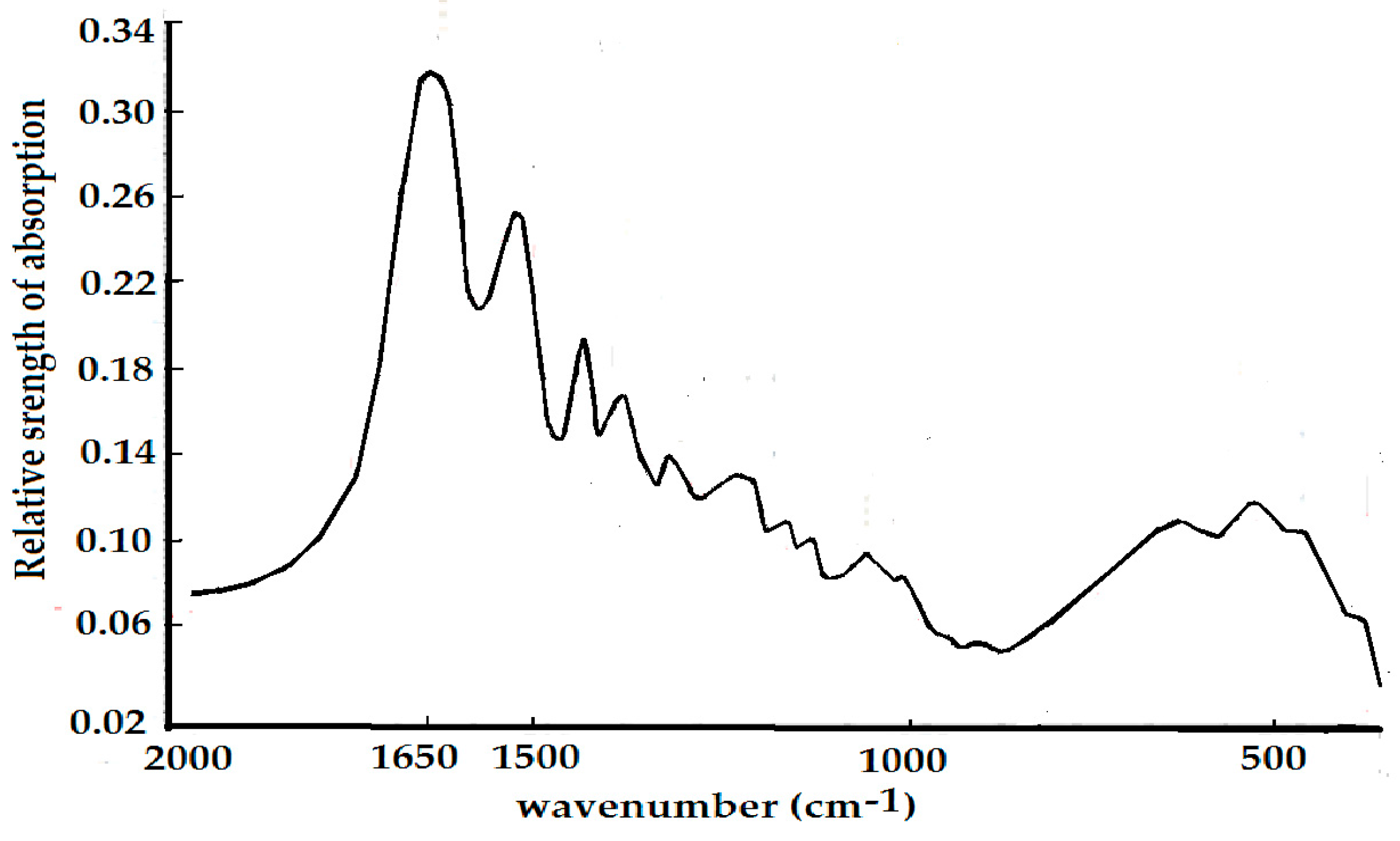
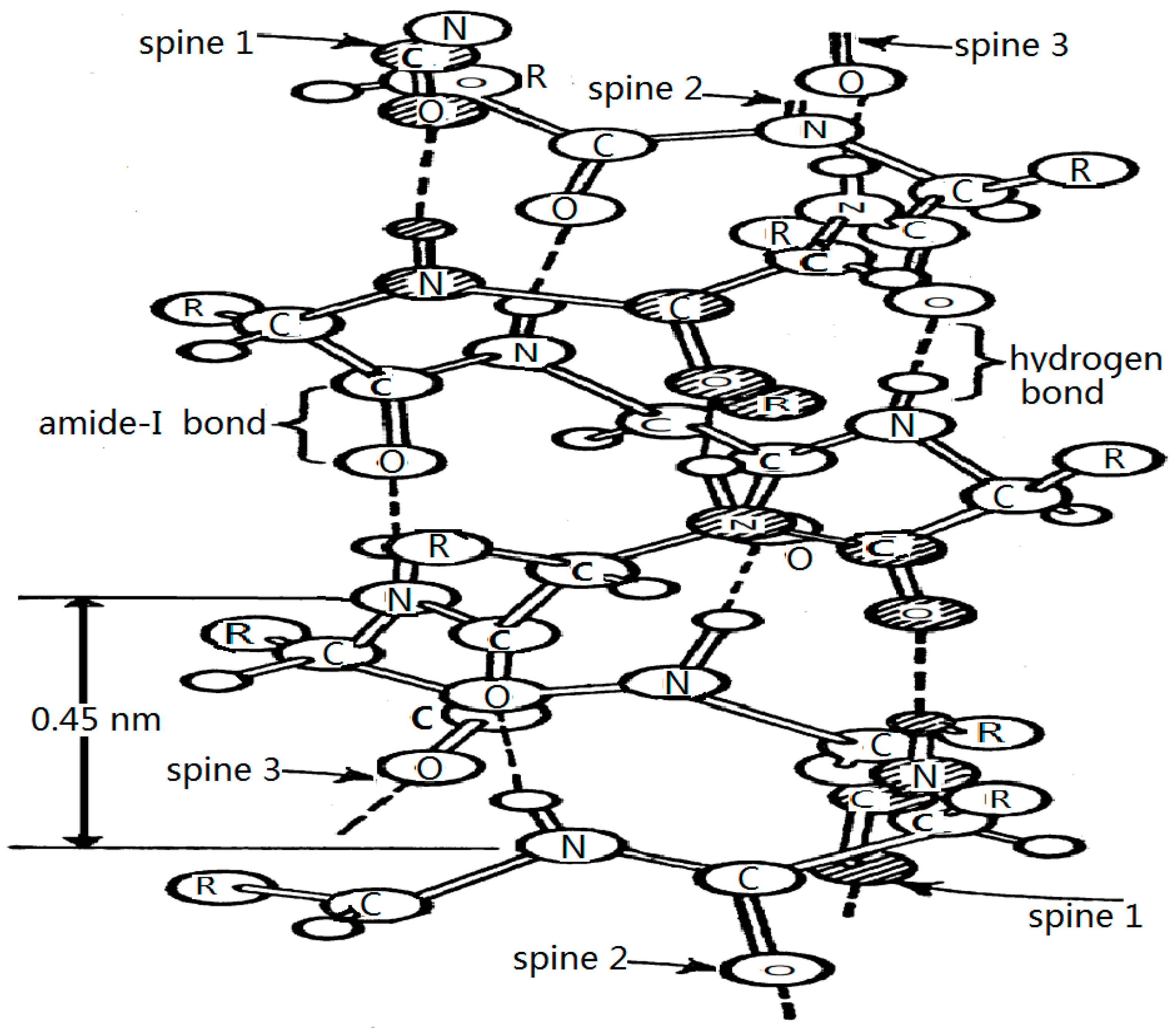
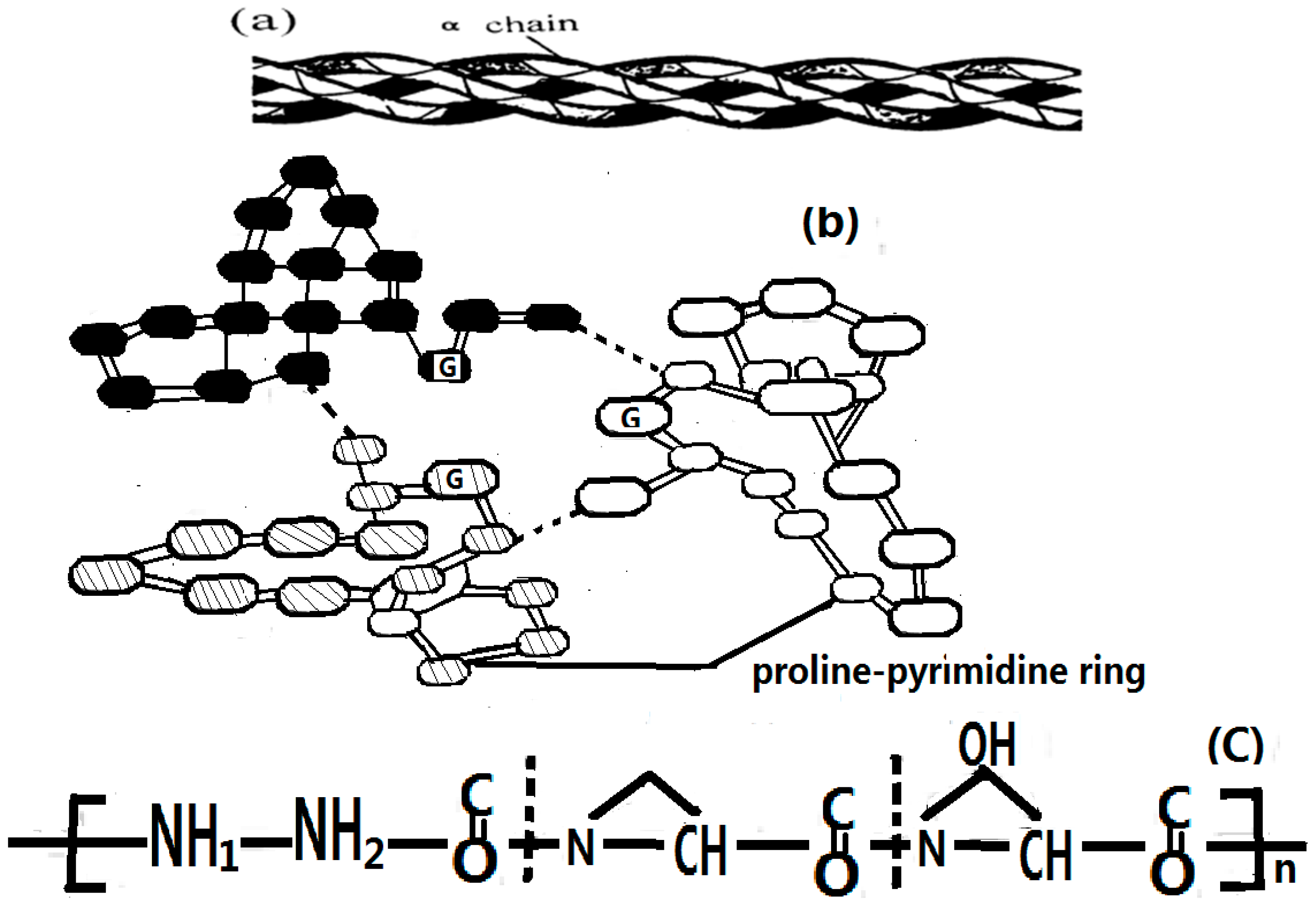
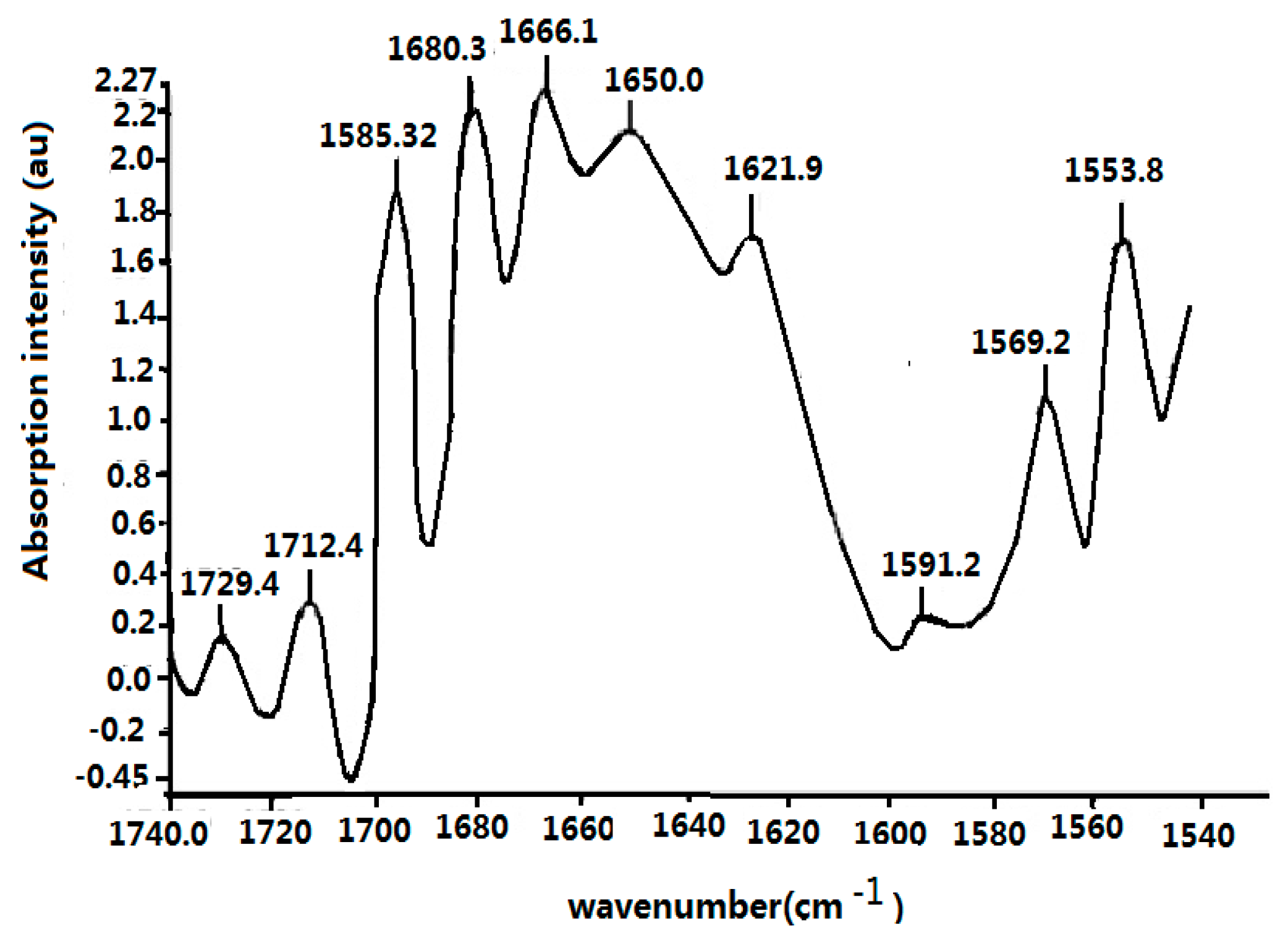
4.1. Experimental Evidence of the Existence of Solitons in Protein Molecules
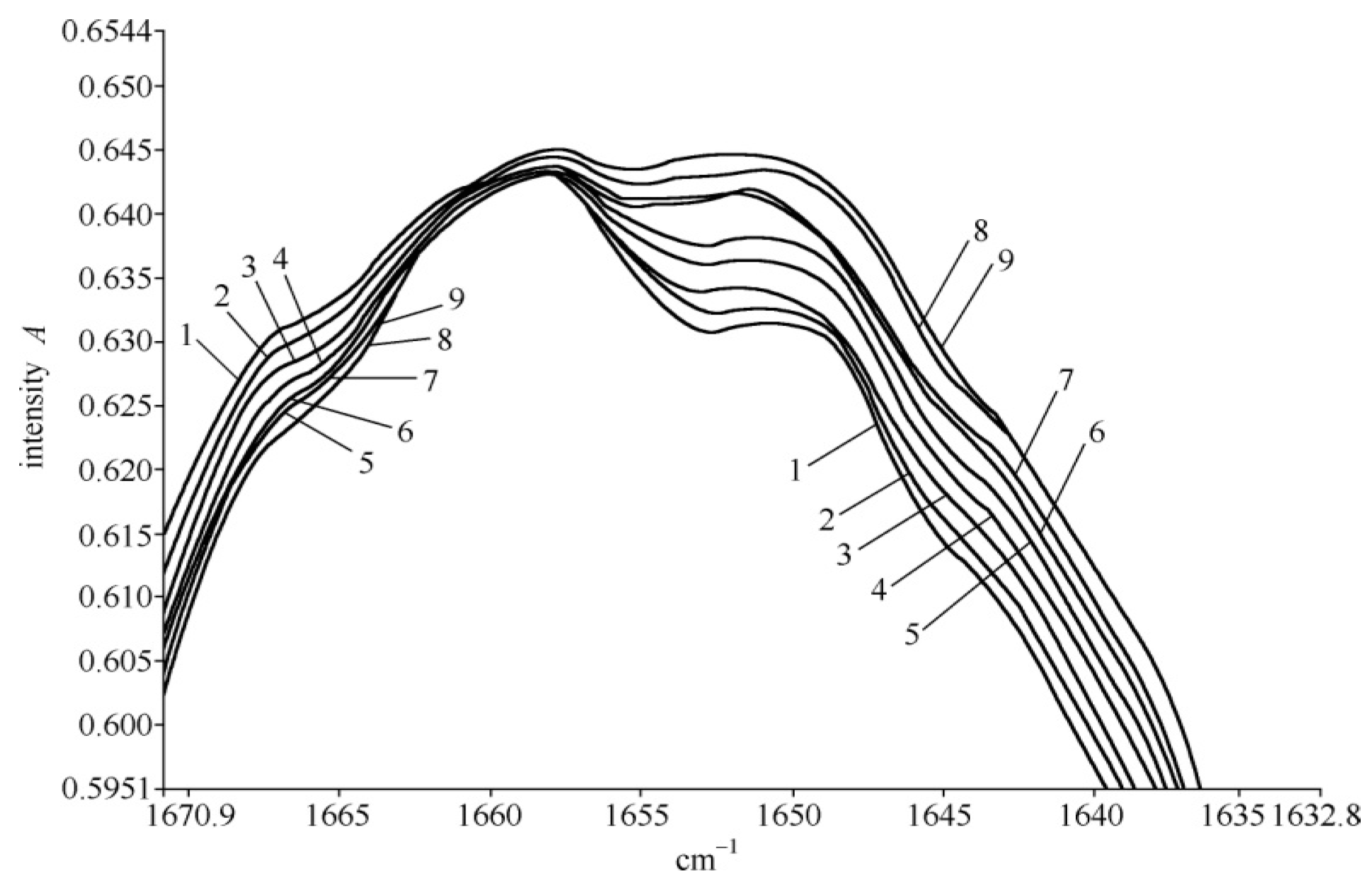
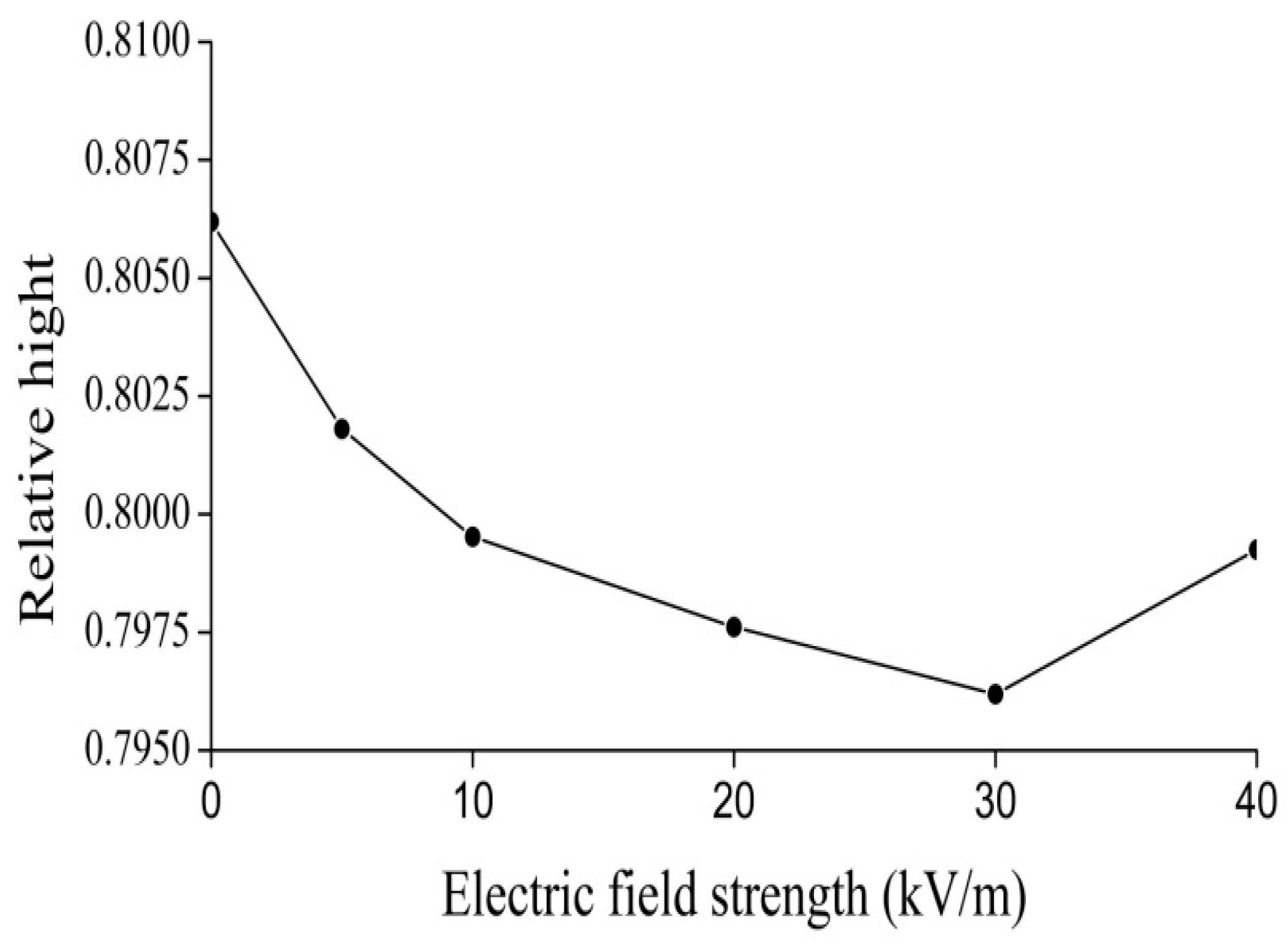
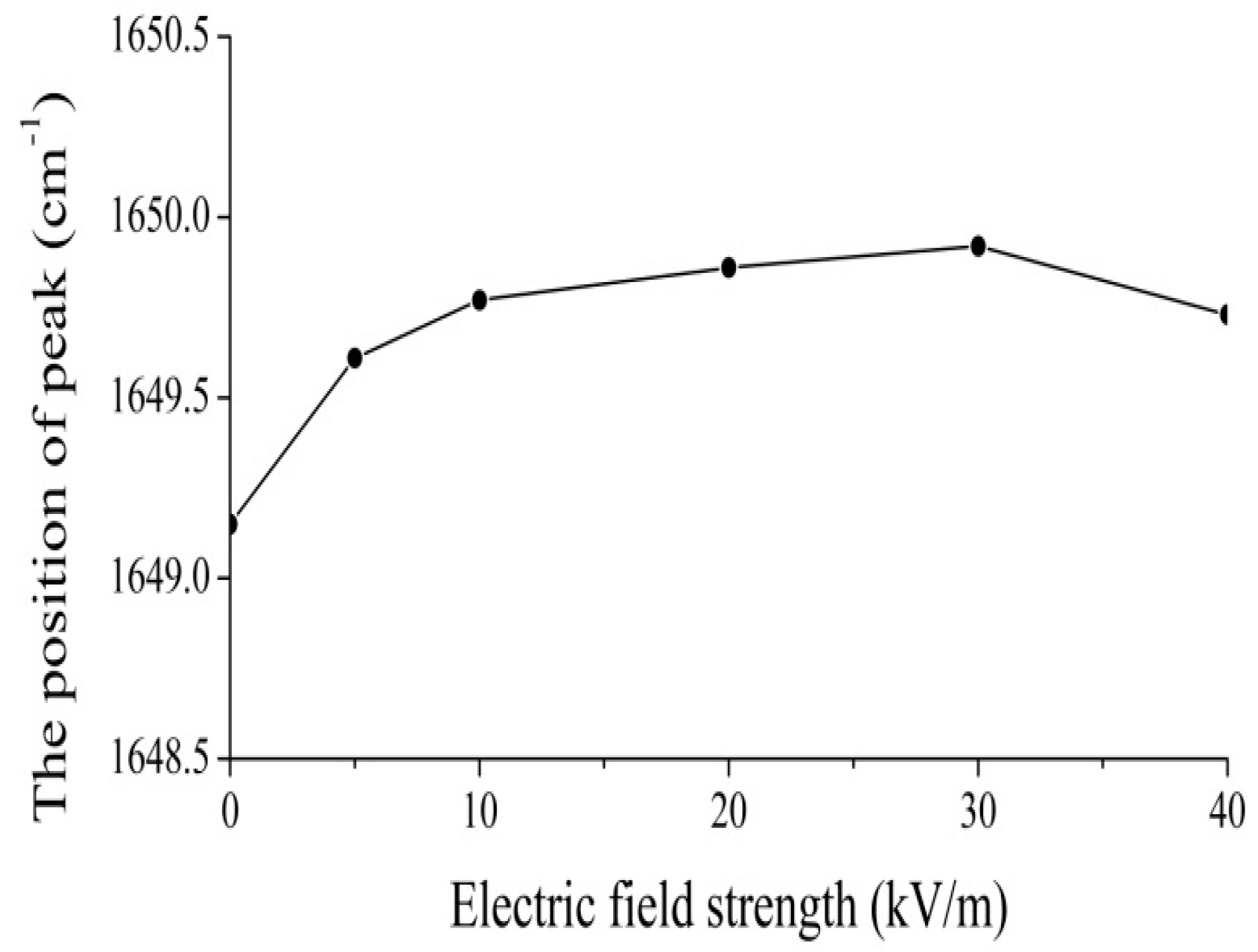
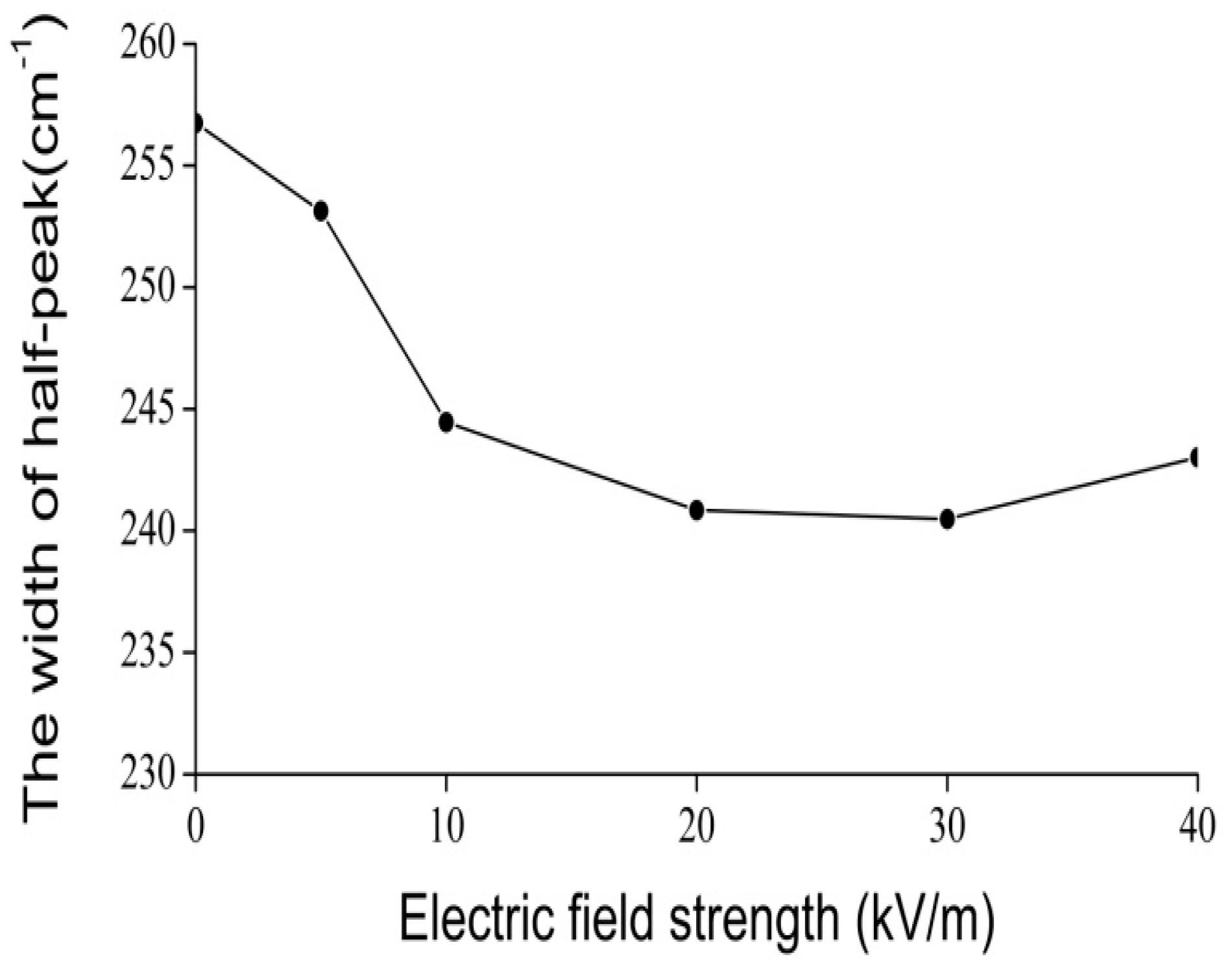
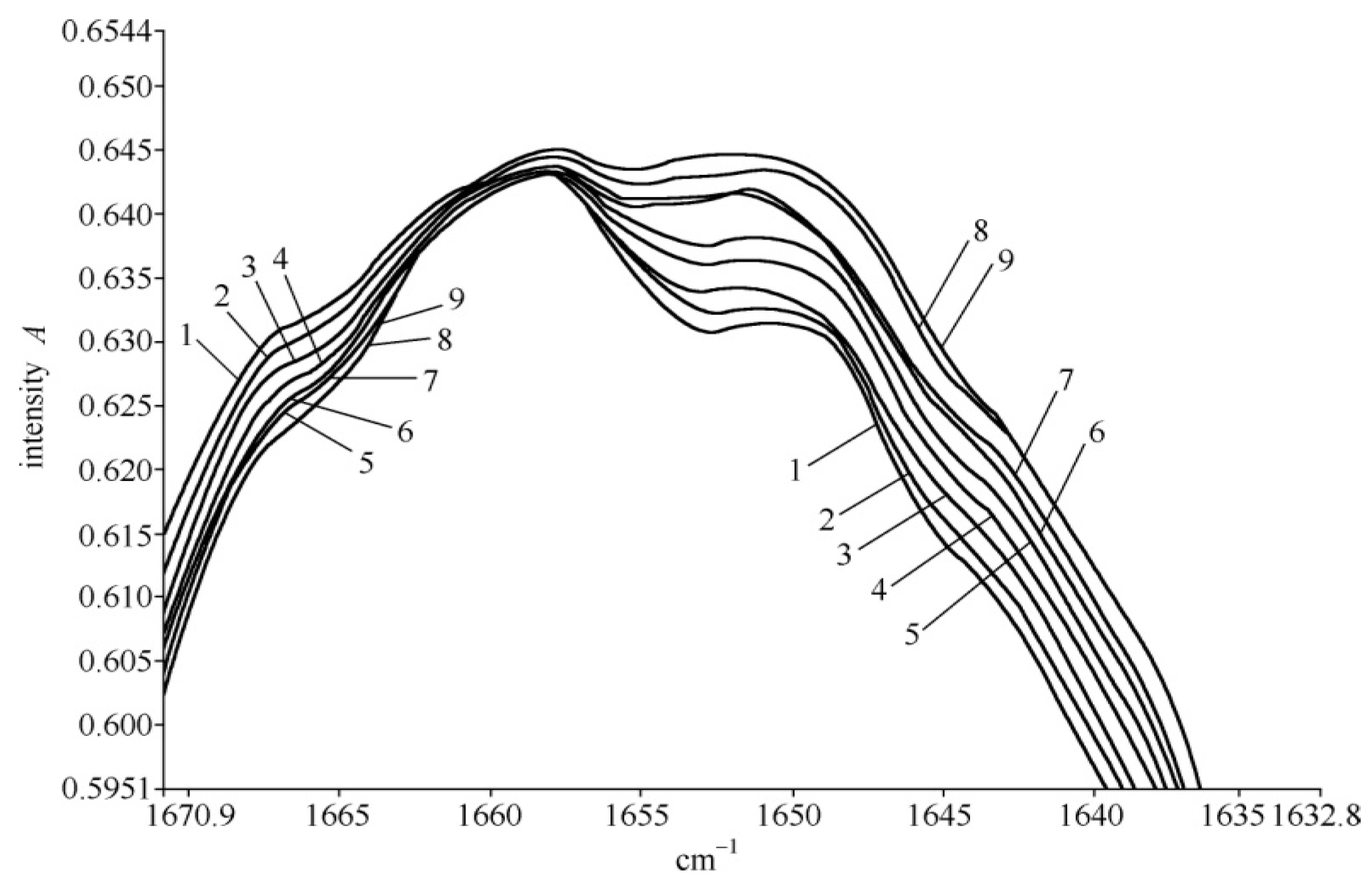
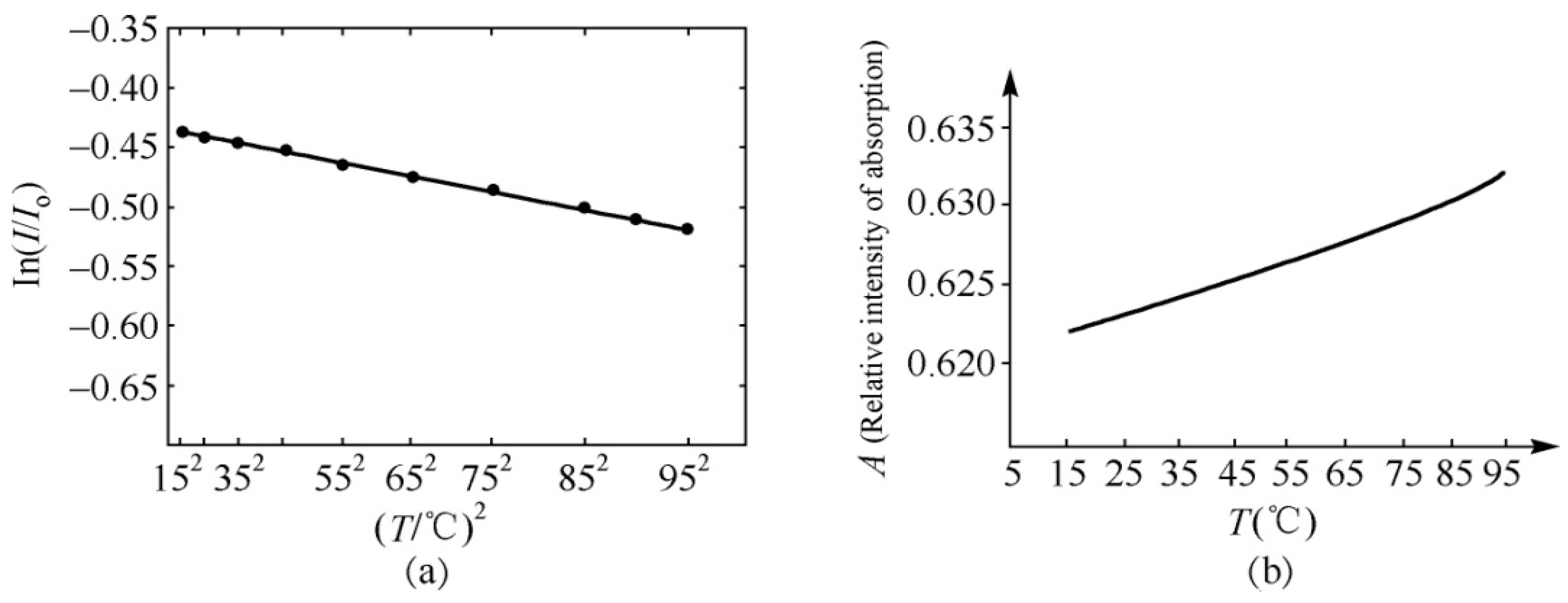
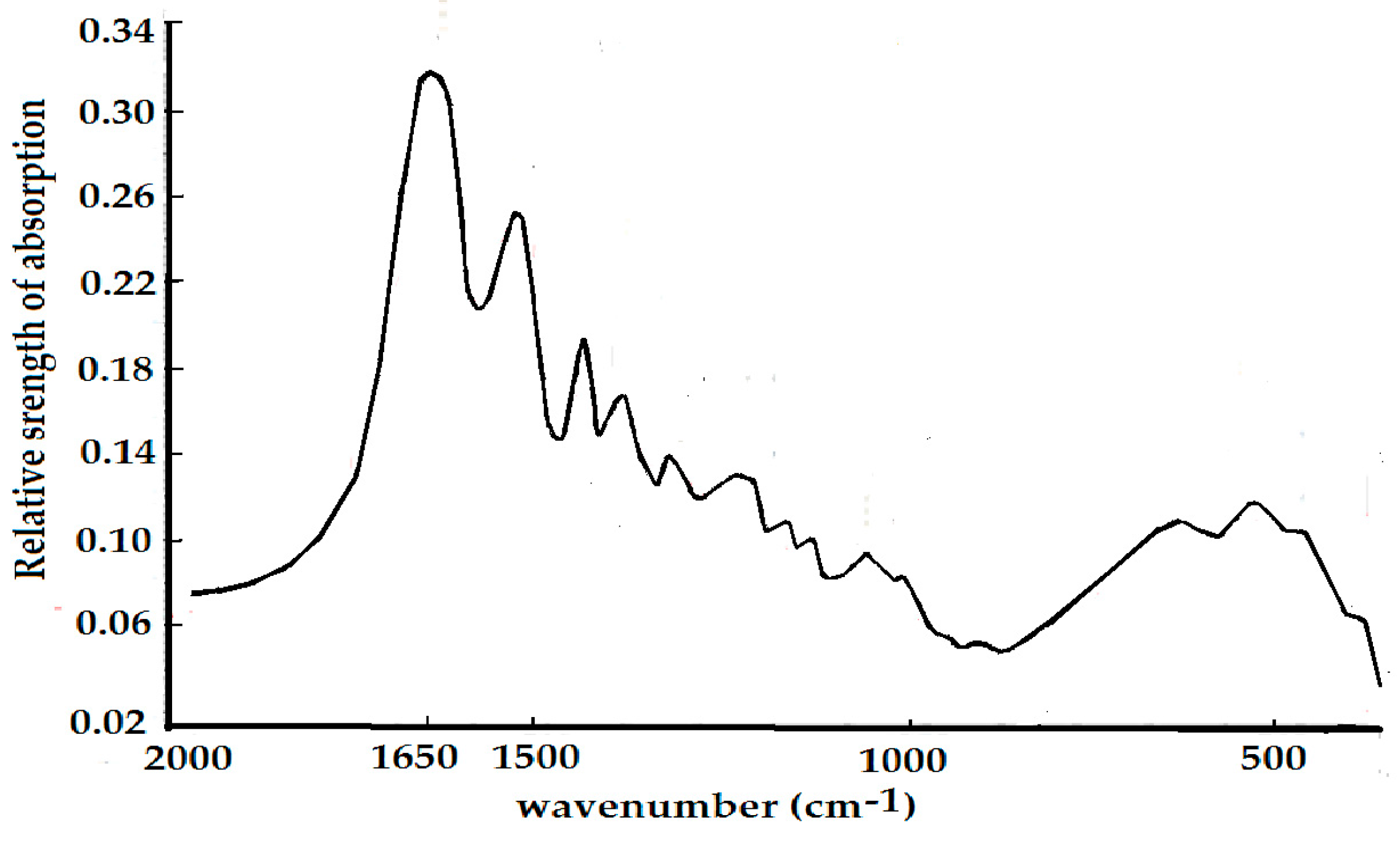
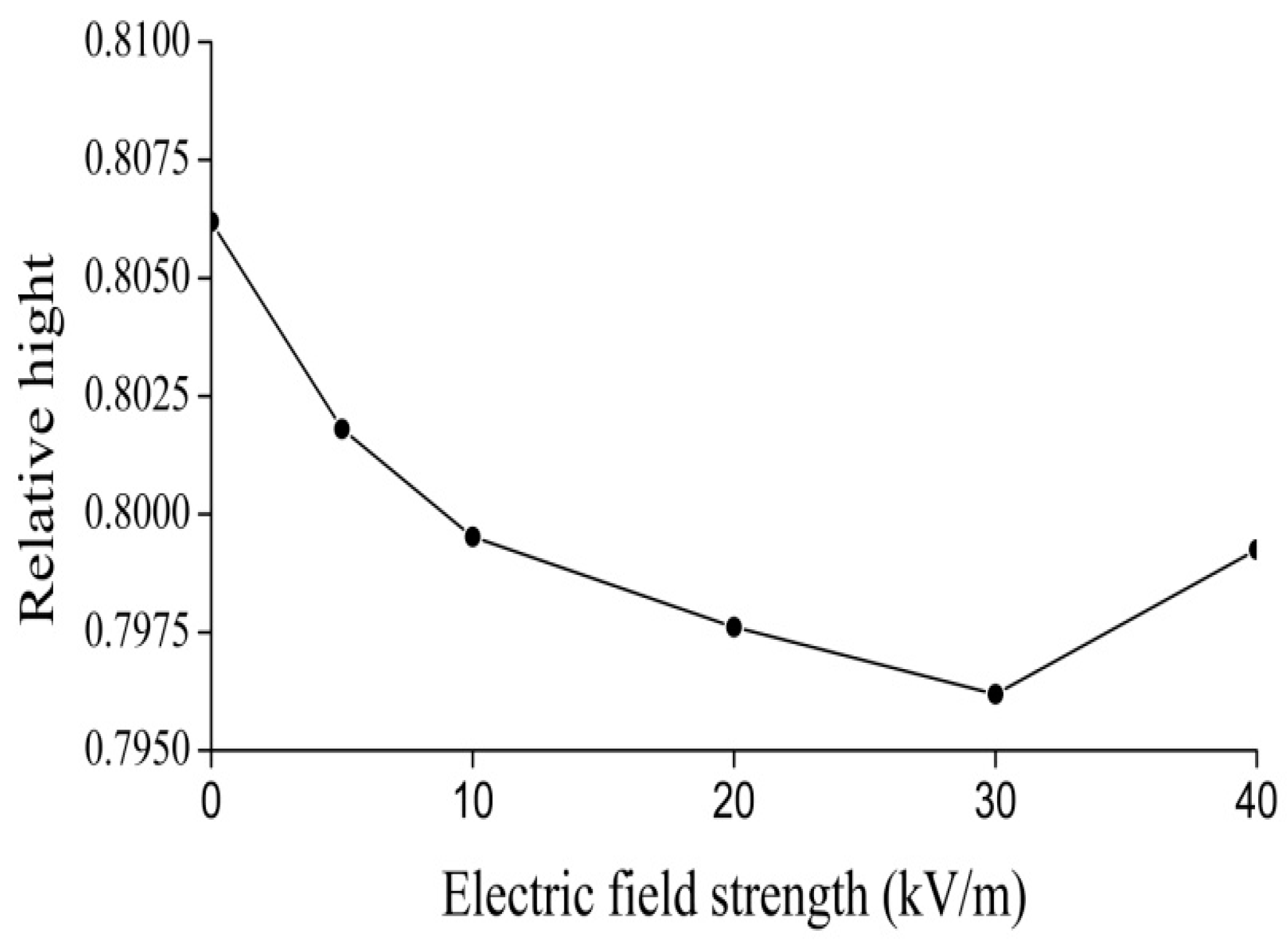
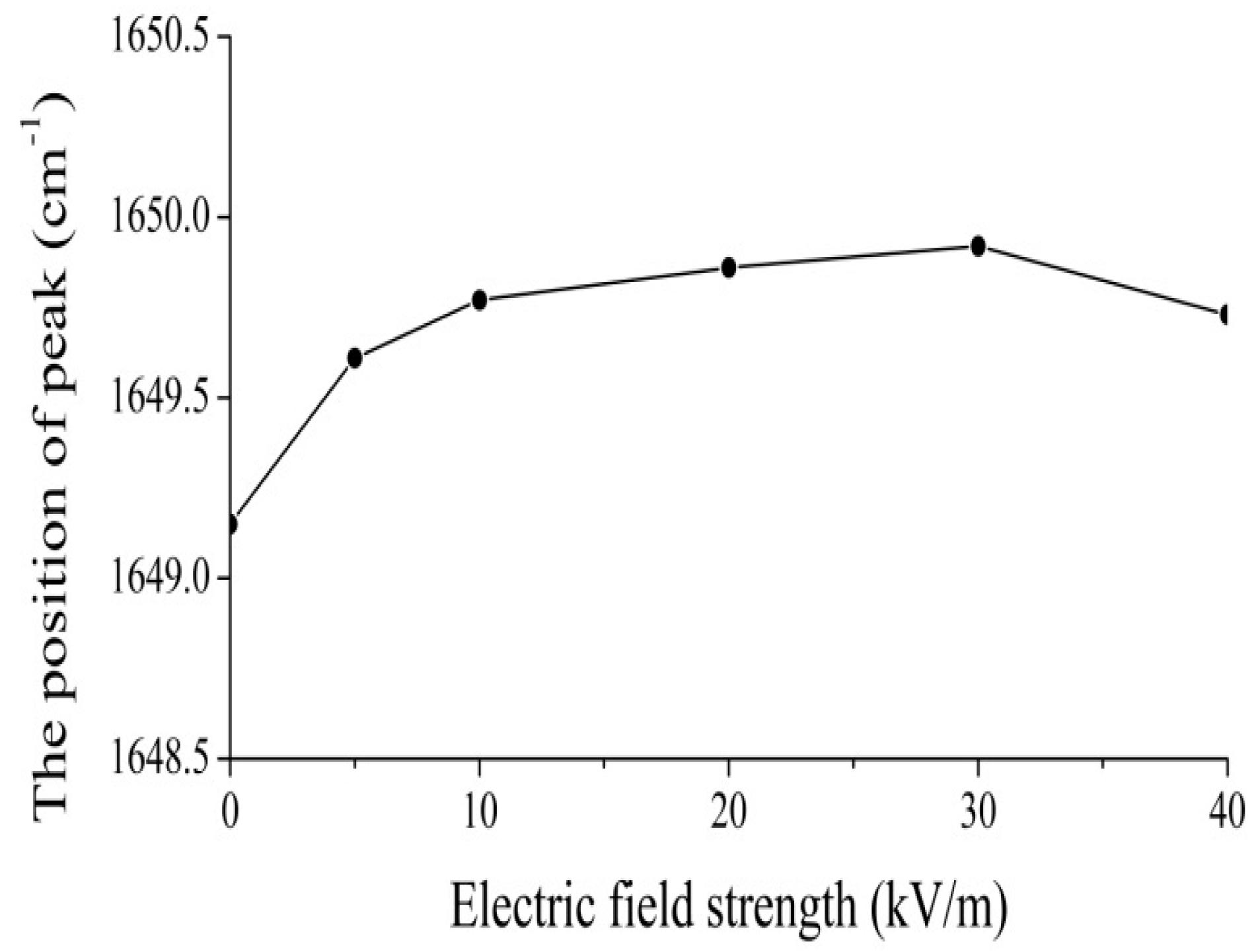
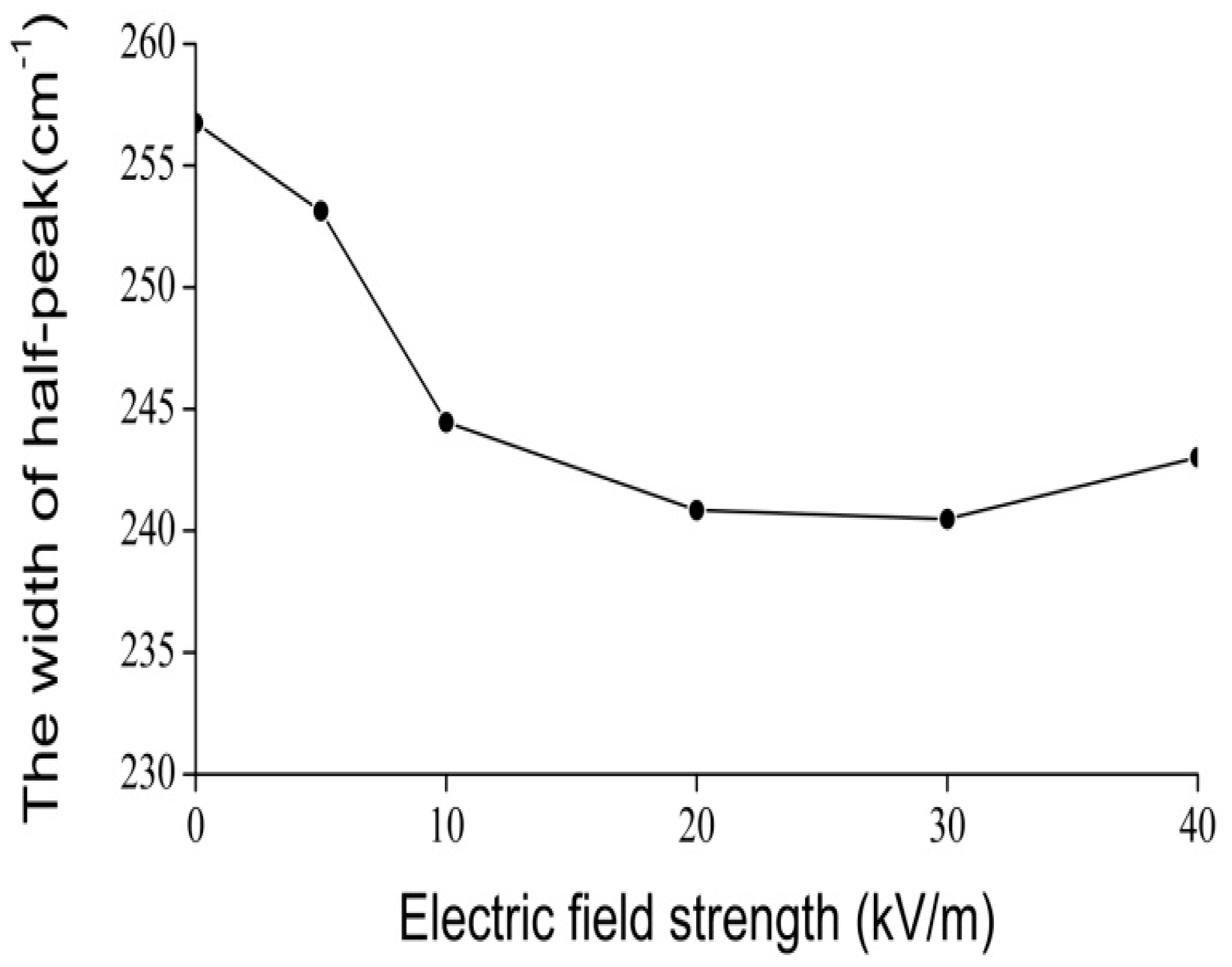
4.2. Experimental Evidence of the Influence of EFs on Solitons in Protein Molecules
5. Conclusions
- (1)
- We have not studied concretely the macroscopic biological effects arising from changes to energy transport under the influences of EMFs and EFs. Thus, further research into the molecular and cellular biology must be pursued. Thus, we cannot confirm that the concrete biological effects arising from variations in the bio-energy transport in protein molecules in the presence of EMFs or EFs are advantageous or harmful to the health of humans and animals.
- (2)
- The mechanism of the biological effects of EFs and EMFs that we propose require additional experimental confirmation, i.e., through instrumentation and novel methods, taking direct measurements of any changed features in the electric dipole moments of α-helix proteins or of the amino acid residues in them arising from EMFs and EFs, which entails many challenges.
- (3)
- Because the strength of EMF frequency is altered in a real EMF, we studied the biological effects of static EFs or DC fields only. Therefore, further investigations would focus on the biological effects of AC fields or altered EFs.
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A. Solutions to Equations (17)–(20)
Appendix B. Solutions to Equations (17)–(20)
References
- Pang, X.F. Bio-Electromagnetism, 1st ed.; The Defense Industry Press of China: Beijing, China, 2009; pp. 14–52. [Google Scholar]
- Fröhlich, H. Biological Coherence and Response to External Stimuli, 1st ed.; Springer-Verlag: New York, NY, USA, 1988; pp. 5–64. [Google Scholar]
- Ahlbom, A.; Day, N.; Feychting, M.; Skinner, R.E.; Dockerty, J.; Linet, J.; McBride, M.; Michaelis, M.; Olsen, J.; Tynes, T.; et al. A pooled analysis of magnetic fields and childhood leukaemia. Br. J. Cancer 2000, 83, 692–698. [Google Scholar] [CrossRef] [PubMed]
- Dowson, D.I.; Lewith, G.; Campbell, M.; Mullee, M.A.; Brewster, L.A. Overhead high-voltage cables and recurrent headache and depressions. Practitioner 1988, 232, 435–436. [Google Scholar] [PubMed]
- Greenland, S.; Sheppard, A.R.; Kaune, W.T.; Poole, C.; Kelsh, M.A. A Pooled analysis of magnetic fields, wire codes, and childhood leukemia. Epidemiology 2000, 11, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Perry, S.; Pearl, L.; Binns, R. Power frequency magnetic field: Depressive Illness and myocardial infarction. Public Health 1989, 103, 177–180. [Google Scholar] [CrossRef]
- Perry, F.S.; Reichmanis, M.; Marino, A.A.; Becker, R.O. Environmental power-frequency magnetic fields and suicide. Health Phys. 1981, 41, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Bromberg, J.; Darnell, J.E. The role of STATs in transcriptional control and their impact on cellular function. Oncogene 2000, 19, 2468–2473. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.F. Biophysics; Press of University of Electronic Science and Technology of China: Chengdu, China, 2008; pp. 67–173. [Google Scholar]
- Li, G.; Pang, X.F. Biological effects of environmental electromagnetic fields. Adv. Mater. Res. 2011, 183–185, 532–536. [Google Scholar] [CrossRef]
- Li, G.; Pang, X.F. The influence of electromagnetic field on the electromagnetic properties of biological tissue. Prog. Biochem. Biophys. (Chin.) 2011, 38, 604–610. [Google Scholar] [CrossRef]
- Li, G.; Li, B.; Yang, S.F.; Pang, X.F. The influences of static magnetic field on the electric features of testicular tissue of rats. Aerosp. Med. Med. Eng. 2012, 25, 172–175. [Google Scholar]
- Li, G.; Yan, Y.Y.; Han, S.G.; Pang, X.F. The influences of static magnetic field on the brain tissue of the rats using infrared spectrum metho. Laser Infrared 2012, 42, 894–896. [Google Scholar]
- Li, G.; Yan, Y.J.; Huan, Y.; Zhou, Y.; Pang, X.F. The influences of electromagnetic field with extremely low frequency on the characteristics of infrared spectrum of the sensitive tissue in rat. Spectrosc. Spectr. Anal. 2012, 32, 1194–1197. [Google Scholar]
- Li, G.; Li, B.; Yan, Y.J.; Yang, S.F.; Pang, X.F. Calculation of equivalent electrical parameters in human tissue and its applications. Mater. Rev. 2011, 25, 142–145. [Google Scholar]
- Zhang, Y.M.; Zhou, Y.; Pang, X.F. The Investigation of effects of pulse electromagnetic field on the fluorescence spectrum of serum in rat. Spectrosc. Spectr. Anal. 2012, 32, 2162–2165. [Google Scholar]
- Zhang, Y.M.; Li, G.; Yan, Y.J.; Pang, X.F. Altered expression of matrix metalloproteinases and tight junction proteins with PEMF-induced BBB permeability change in rats. Biomed. Environ. Sci. 2011, 24, 408–414. [Google Scholar]
- Beebe, S.J.; Fox, P.M.; Rec, L.J.; Somers, K.; Stark, R.H.; Schoenbach, K.H. Nanosecond pulsed electric field (nsPEF) effects on cells and tissues: Apoptosis induction and tumor growth inhibition. IEEE Trans. Plasma Sci. 2002, 30, 286–292. [Google Scholar] [CrossRef]
- Lazzaro, V.D.; Capone, F.; Apollonio, F.; Borea, P.A.; Cadossi, R.; Fassina, L.; Grassi, C.; Liberti, M.; Paffi, A.; Parazzini, M.; et al. A consensus panel review of central nervous system effects of the exposure to low-intensity extremely low-frequency magnetic fields. Brain Stimul. 2013, 6, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Apollonio, F.; Liberti, M.; Paffi, A.; Merla, C.; Marracino, P.; Denzi, A.; Marino, C.; d’Inzeo, G. Feasibility for microwaves energy to affect biological systems via nonthermal mechanisms: A systematic approach. IEEE Trans. Microw. Theory Tech. 2013, 61, 2031–2045. [Google Scholar] [CrossRef]
- Hofmann, G.A.; Dev, S.B.; Dimer, S.; Nanda, G.S. Electroporation therapy: A new approach for treatment of head and neck cancer. IEEE Trans. Biomed. Eng. 1999, 46, 752–759. [Google Scholar] [CrossRef] [PubMed]
- Kekez, M.M.; Savic, P.; Johnson, B.F. Contribution to the biophysics of the lethal effects of electric field on microorganisms. Biochim. Biophys. Acta 1996, 1278, 79–88. [Google Scholar] [CrossRef]
- Pang, X.F.; Zhang, A.Y. Investigations of mechanism and properties of biological thermal effects of the microwave. China J. Atom. Mol. Phys. 2001, 18, 278–281. [Google Scholar]
- Davydov, A.S. Biology and Quantum Mechanics; Pergaman: New York, NY, USA, 1982; pp. 161–211. [Google Scholar]
- Davydov, A.S. Solitons in Molecular Systems, 2nd ed.; Reidel Publishing Company: Dordrecht, The Netherlands, 1991; pp. 23–79. [Google Scholar]
- Davydov, A.S.; Ermakov, V.N. Soliton generation at the boundary of molecular chain. Phys. D 1988, 32, 318–329. [Google Scholar] [CrossRef]
- Davydov, A.S. Solitons in molecular systems. Phys. Scr. 1979, 20, 387–394. [Google Scholar] [CrossRef]
- Davydov, A.S. The lifetime of molecular solitons. J. Biol. Phys. 1991, 18, 111–125. [Google Scholar] [CrossRef]
- Davydov, A.S. Solitons and energy transfer along protein molecules. J. Theor. Biol. 1977, 66, 379–387. [Google Scholar] [CrossRef]
- Scott, A.C. Davydov’s soliton. Phys. Rep. 1992, 217, 1–67. [Google Scholar] [CrossRef]
- Scott, A.C. Dynamics of Davydov’s soliton. Phys. Rev. A 1982, 26, 578–595. [Google Scholar] [CrossRef]
- Brown, D.W.; Ivic, Z. Unification of polaron and soliton theories of exciton transport. Phys. Rev. E 1989, 40, 9876–9887. [Google Scholar] [CrossRef]
- Brown, D.W.; Lindenberg, K.; West, B.J. Nonlinear density-matrix equation for the study of finite-temperature soliton dynamics. Phys. Rev. B 1987, 35, 6169–6180. [Google Scholar] [CrossRef]
- Guo, B.L.; Pang, X.F. Solitons; Chinese Science Press: Beijing, China, 1987; pp. 4–140. [Google Scholar]
- Pang, X.F. The transport of bio-energy in protein molecules. Chin. J. Biochem. Biophys. 1986, 18, 1–8. [Google Scholar]
- Christiansen, P.L.; Scott, A.C. Davydov’s Soliton Revisited; Plenum Press: New York, NY, USA, 1990; pp. 34–103. [Google Scholar]
- Cruzeiro, L.; Halding, J.; Christiansen, P.L.; Skovgard, O.; Scott, A.C. Temperature effects on Davydov soliton. Phys. Rev. A 1985, 37, 880–887. [Google Scholar] [CrossRef]
- Cruzeio-Hansson, L. Mechanism of thermal destabilization of the Davydov soliton. Phys. Rev. A 1992, 45, 4111–4115. [Google Scholar] [CrossRef]
- Forner, W. Quantum and disorder effects in Davydov soliton theory. Phys. Rev. A 1991, 44, 2694–2708. [Google Scholar] [CrossRef] [PubMed]
- Forner, W. Quantum and temperature effects on Davydov soliton dynamics: Averaged Hamiltonian method. J. Phys. Condens. Matter 1992, 4, 1915–1923. [Google Scholar] [CrossRef]
- Forner, W. Davydov soliton dynamics: Two-quantum states and diagonal disorder. J. Phys. Condens. Matter 1991, 3, 3235–3254. [Google Scholar] [CrossRef]
- Lomdahl, P.S.; Kerr, W.C. Do Davydov solitons exist at 300 K? Phys. Rev. Lett. 1985, 55, 1235–1239. [Google Scholar] [CrossRef] [PubMed]
- Kerr, W.C.; Lomdahl, P.S. Quantum-mechanical derivation of the equations of motion for Davydov solitons. Phys. Rev. B 1987, 35, 3629–3632. [Google Scholar] [CrossRef]
- Wang, X.; Brown, D.W.; Lindenberg, K.; West, B. Alternative formulation of Davydov theory of energy transport in biomolecules systems. Phys. Rev. A 1988, 37, 3557–3566. [Google Scholar] [CrossRef]
- Wang, X.; Brown, D.W.; Lindenberg, K. Quantum Monte Carlo simulation of Davydov model. Phys. Rev. Lett. 1989, 62, 1796–1799. [Google Scholar] [CrossRef] [PubMed]
- Cottingham, J.P.; Schweitzer, J.W. Calculation of the lifetime of a Davydov soliton at finite temperature. Phys. Rev. Lett. 1989, 62, 1792–1795. [Google Scholar] [CrossRef] [PubMed]
- Schweitzer, J.W. Lifetime of the Davydov soliton. Phys. Rev. A 1992, 45, 8914–8922. [Google Scholar] [CrossRef] [PubMed]
- Mettle, B.; Shaw, P.B. Evolution of a molecular exciton on a Davydov lattice at T = 0. Phys. Rev. B 1988, 38, 3075–3086. [Google Scholar]
- Ivic, Z.; Przulj, Z.; Kosti, D. Decay and slowing down of the multiquanta Davydov-like solitons in molecular chains. Phys. Rev. E 2000, 61, 6963–6967. [Google Scholar] [CrossRef]
- Teki, J.; Ivic, Z.; Zekovi, S.; Przulj, Z. Kinetic properties of multiquanta Davydov-like solitons inmolecular chains. Phys. Rev. E 1999, 60, 821–825. [Google Scholar] [CrossRef]
- Zekovi, S.; Ivic, Z. Damping and modification of the multiquanta Davydov-like solitons in molecular chains. Bioelectrochem. Bioenerg. 1999, 48, 297–300. [Google Scholar] [CrossRef]
- Pouthier, V. Two-vibron bound states in alpha-helix proteins: The interplay between the intramolecular anharmonicity and the strong vibron-phonon coupling. Phys. Rev. E 2003, 68. [Google Scholar] [CrossRef] [PubMed]
- Pouthier, V.; Falvo, C. Relaxation channels of two-vibron bound states in α-helix proteins. Phys. Rev. E 2004, 69. [Google Scholar] [CrossRef] [PubMed]
- Falvo, C.; Pouthier, V. Vibron–polaron in α-helices. I. Single-vibron states. J. Chem. Phys. 2005, 123, 184709–184716. [Google Scholar] [CrossRef] [PubMed]
- Falvo, C.; Pouthier, V. Vibron–polaron in α-helices. II. Two-vibron states. J. Chem. Phys. 2005, 123. [Google Scholar] [CrossRef] [PubMed]
- Cruzeiro, L. The Davydov/Scott model for energy storage and transport in proteins. J. Biol. Phys. 2009, 35, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Silva, P.A.S.; Cruzeiro, L. Dynamics of a nonconserving Davydov monomer. Phys. Rev. E 2006, 74. [Google Scholar] [CrossRef] [PubMed]
- Silva, P.A.S.; Cruzeiro-Hansson, L. A reduced set of exact equations of motion for a non-number-conserving Hamiltonian. Phys. Lett. A 2003, 315, 447–451. [Google Scholar] [CrossRef]
- Pouthier, V. Energy relaxation of the amide-I mode in hydrogen-bonded peptide units: A route to conformational change. J. Chem. Phys. 2008, 128. [Google Scholar] [CrossRef] [PubMed]
- Pouthier, V.; Tsybin, Y.O. Amide-I relaxation-induced hydrogen bond distortion: Anintermediate in electron capture dissociation mass pectrometry of α-helical peptides? J. Chem. Phys. 2008, 129. [Google Scholar] [CrossRef] [PubMed]
- Moritsugu, K.; Miyashita, O.; Kidera, K. Vibrational energy transfer in a protein molecule. Phys. Rev. Lett. 2000, 85, 3970–3973. [Google Scholar] [CrossRef] [PubMed]
- Fujisaki, H.; Zhang, Y.; Straub, J.E. Time-dependent perturbation theory for vibrational energy relaxation and dephasing in peptides and proteins. J. Chem. Phys. 2006, 124. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.F. The properties of collective excitation in organic protein molecular system. J. Phys. Condens. Matter 1990, 2, 9541–9553. [Google Scholar]
- Pang, X.F. Comment the thermodynamic properties of α-helix protein: A soliton approach. Phys. Rev. 1994, 49, 4747–4751. [Google Scholar]
- Pang, X.F. Nonlinear Quantum Mechanical Theory; Chongqing Press: Chongqing, China, 1994; pp. 356–465. [Google Scholar]
- Pang, X.F. Properties of soliton in protein molecules with nonlinear nearest neighbour interaction. Chin. Sci. Bull. 1993, 38, 1572–1581. [Google Scholar]
- Pang, X.F. The thermodynamic properties of the solitons excited in the protein molecules. Chin. Sci. Bull. 1993, 38, 1665–1677. [Google Scholar]
- Pang, X.F. Quantum-mechamical method for the soliton transported bio-energy in protein. Chin. Phys. Lett. 1993, 10, 437–440. [Google Scholar] [CrossRef]
- Pang, X.F. Stability of the soliton excited in protein in the biological temperature range. Chin. Phys. Lett. 1993, 10, 573–576. [Google Scholar] [CrossRef]
- Pang, X.F. Influence of the soliton in anharmonic molecular crystals with temperature on Mossbauer effect. Eur. Phys. J. B 1999, 10, 415–423. [Google Scholar] [CrossRef]
- Pang, X.F.; Chen, X.R. Distribution of vibrational energy- levels of protein molecular chains. Commun. Theor. Phys. 2001, 35, 323–326. [Google Scholar]
- Pang, X.F. The effect of Raman scattering accompanied by the soliton excitation occurred in the molecular crystals. Phys. D 2001, 154, 138–158. [Google Scholar] [CrossRef]
- Pang, X.F. An improvement of the Davydov theory of bio-energy etransport in the protein molecular systems. Phys. Rev. E 2000, 62, 6989–6998. [Google Scholar]
- Pang, X.F. The lifetime of the soliton in the improved Davydov model at the biological temperature 300 K for protein molecules. Eur. Phys. J. B 2001, 19, 297–316. [Google Scholar]
- Pang, X.F.; Zhang, H.W.; Yu, J.F.; Feng, Y.P. States and properties of the soliton transported bio-energy in nonuniform protein molecules at physiological temperature. Phys. Lett. A 2005, 335, 408–416. [Google Scholar] [CrossRef]
- Pang, X.F.; Luo, Y.H. Stabilization of the soliton transported bio-energy in protein molecules in the improved model. Commun. Theor. Phys. 2004, 41, 470–476. [Google Scholar]
- Pang, X.F.; Feng, Y.P. Quantum Mechanics in Nonlinear Systems; World Science Publisher Co.: Singapore, 2005; pp. 487–556. [Google Scholar]
- Pang, X.F.; Liu, M.J. Features of Motion of Soliton Transported Bio-energy in aperiodic alpha-helix protein molecules with three channels. Commun. Theor. Phys. 2009, 51, 170–180. [Google Scholar]
- Pang, X.F. The effects of damping and temperature of medium on the soliton excited in α-helix protein molecules with three channels. Mod. Phys. Lett. B 2009, 23, 71–88. [Google Scholar] [CrossRef]
- Pang, X.F.; Lui, M.J. The influences of temperature and chain-chain interaction on features of solitons excited in α-Helix protein molecules with three channels. Int. J. Mod. Phys. B 2009, 23, 2303–2322. [Google Scholar] [CrossRef]
- Pang, X.F. Influence of structure disorders and temperatures of systems on the bio-energy transport in protein molecules. Front. Phys. China 2008, 3, 457–488. [Google Scholar] [CrossRef]
- Pang, X.F.; Yu, J.F.; Lao, Y.H. Combination effects of structure nonuniformity of proteins on the soliton transported bio-energy. Int. J. Mod. Phys. B 2007, 21, 13–42. [Google Scholar] [CrossRef]
- Pang, X.F.; Liu, M.J. Properties of soliton-transported bio-energy in α-helix protein molecules with three channels. Commun. Theor. Phys. 2007, 48, 369–376. [Google Scholar]
- Pang, X.F. Theory of bio-energy transport in protein molecules and its experimental evidences as well as applications (I). Front. Phys. China 2007, 2, 469–493. [Google Scholar] [CrossRef]
- Pang, X.F.; Zhang, H.W. Theoretical investigation of properties of infrared absorption of α-helix protein molecules. Int. J. Infrared Millim. Waves 2006, 27, 735–812. [Google Scholar] [CrossRef]
- Pang, X.F.; Zhang, H.W.; Liu, M.J. Influences of heat bath and structure disorder in protein molecules on the soliton transported bio-energy in the improved model. J. Phys. Condens. Matter 2006, 18, 613–627. [Google Scholar] [CrossRef]
- Pang, X.F.; Zhang, H.W.; Yu, J.F.; Luo, Y.H. Influences of variations of characteristic parameters arising from the structure nonuniformity of the protein molecules on states of the soliton transported bio-energy in the improved model. Int. J. Mod. Phys. B 2006, 20, 3027–3041. [Google Scholar] [CrossRef]
- Pang, X.F.; Chen, X.R. The properties of nonlinear energy-spectra of acetanilide. Int. J. Mod. Phys. 2006, 20, 2505–2518. [Google Scholar] [CrossRef]
- Pang, X.F.; Yu, J.F.; Luo, Y.H. Influences of quantum and disorder effects on solitons exited in protein molecules in improved model. Commun. Theor. Phys. 2005, 43, 367–376. [Google Scholar]
- Pang, X.F.; Zhang, H.W. Changes of the Mössbauer effect caused by the excitation of the solitons in the organic molecular crystals at finite temperature. J. Phys. Chem. Solids 2005, 66, 963–972. [Google Scholar]
- Pang, X.F.; Zhang, H.W.; Yu, J.F.; Luo, Y.H. Thermal stability of the new soliton transported bio-energy under influence of fluctuations of characteristic Parameters at biological temperature in the protein molecules. Int. J. Mod. Phys. B 2005, 19, 4677–4699. [Google Scholar] [CrossRef]
- Pang, X.F.; Yu, J.F.; Liu, M.J. Changes of properties of the soliton with temperature under influences of structure disorder in the α-helix protein molecules with three channels. Mol. Phys. 2010, 108, 1297–1315. [Google Scholar] [CrossRef]
- Pang, X.F.; Chen, X.R. Calculation of vibrational energy-spectra of α-helical protein molecules and its properties. Commun. Theor. Phys. 2002, 37, 715–722. [Google Scholar]
- Pang, X.F.; Chen, X.R. Vibrational energy-spectra and infrared absorption of α-helical protein molecules. Chin. Phys. Lett. 2002, 19, 1096–1099. [Google Scholar]
- Pang, X.F. Nonlinear Quantum Mechanics; Chinese Electronic Industry Press: Beijing, China, 2009; pp. 356–412. [Google Scholar]
- Pang, X.F.; Xiao, H.L.; Cue, G.P.; Zhang, H.W.; Dong, B. Experiment studies of properties of infrared absorption of biological tissues. Int. J. Infrared Millim. Waves 2010, 31, 521–532. [Google Scholar]
- Pang, X.F. The properties of bio-energy transport and influence of structure nonuniformity and temperature of systems on energy transport along polypeptide chains. Prog. Biophys. Mol. Biol. 2012, 108, 1–46. [Google Scholar] [PubMed]
- Pang, X.F. The mechanism and properties of bio-photon emission and absorption in protein molecules in living systems. J. Appl. Phys. 2012, 111. [Google Scholar] [CrossRef]
- Pang, X.F. The theory of bio-energy transport in the protein molecules and its properties. Phys. Life Rev. 2011, 8, 264–286. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.F. Correctness and completeness of the theory of bio-energy transport. Phys. Life Rev. 2011, 8, 302–306. [Google Scholar] [CrossRef]
- Pang, X.F. The checkout and verification of theory of bio-energy transport in the protein molecules. Biophys. Rev. Lett. 2014, 9, 1–79. [Google Scholar] [CrossRef]
- Pang, X.F. The features of infrared spectrum of bio-polymer and its theoretical investigation. Int. J. Mod. Phys. B 2014, 29. [Google Scholar] [CrossRef]
- Su, X.D.; Jin, F.J. Davydov-Pang model: An improved Davydov protein soliton theory. Phys. Life Rev. 2011, 8, 300–301. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.D. Physical insights to the bio-energy transport in the protein molecules. Phys. Life Rev. 2011, 8, 287–288. [Google Scholar] [CrossRef] [PubMed]
- Tao, S. The function of soliton on bio-energy transport in the protein molecules. Phys. Life Rev. 2011, 8, 291–292. [Google Scholar]
- He, N.Y. An important biological theory—Solving the transport of bio-energy in living systems. Phys. Life Rev. 2011, 8, 296–297. [Google Scholar] [CrossRef] [PubMed]
- Stiefel, J. Einfuhrung in die Numerische Mathematic; Teubner Verlag: Stuttgart, Germany, 1965; pp. 78–156. [Google Scholar]
- Atkinson, K.E. An Introdution to Numerical Analysis; Wiley New York Inc.: New York, NY, USA, 1987; pp. 34–104. [Google Scholar]
- Kumar, A.; Singh, G. Measurement of dielectric constant and loss factor of the dielectric material at microwave frequencies. Prog. Electromagn. Res. 2007, 69, 47–54. [Google Scholar] [CrossRef]
- Clifford, E.F.; Prilusky, J.; Silman, I.; Sussman, J.L. A server and database for dipole moments of proteins. Nucleic Acids Res. Adv. 2007, 35, W512–W521. [Google Scholar]
- Antosiewicz, J. Computation of the dipole moments of proteins. Biophys. J. 1995, 69, 1344–1354. [Google Scholar] [CrossRef]
- Marracino, P.; Amadei, A.; Apollonio, F.; d’Inzeo, G.; Liberti, M.; diCrescenzo, A.; Fontana, A.; Zappacosta, R.; Aschi, M. Modeling of chemical reactions in micelle: Water-mediated, Keto-Enol Interconversion as a case study. J. Phys. Chem. 2011, 115, 8102–8111. [Google Scholar] [CrossRef] [PubMed]
- Careri, G.; Buontempo, U.; Galluzzi, F.; Scott, A.C.; Gratton, E.; Shyamsunder, E. Spectroscopic evidence for Davydov-like solitons in acetanilide. Phys. Rev. B 1984, 30, 4689–4698. [Google Scholar] [CrossRef]
- Careri, G.A.; Gransanti, A.; Rupley, J.A. Critical exponents of protonic percolation in hydrated lysozyme powders. Phys. Rev. A 1988, 37, 2703–2705. [Google Scholar] [CrossRef]
- Careri, G.A.; Gratton, E.A.; Shyamsunder, E. Fine structure of the amide-I bond in acetanilide. Phys. Rev. A 1988, 37, 4048–4051. [Google Scholar] [CrossRef]
- Careri, G.A.; Buonttempo, U.; Caeta, F.; Scott, A.C. Infrared absorption in acetanilide by solitons. Phys. Rev. Lett. 1983, 51, 304–307. [Google Scholar] [CrossRef]
- Careri, G.A.; Eilbeck, J.C. Stability of solutions of the discrete self-trapping equation. Phys. Lett. A 1985, 109, 201–204. [Google Scholar]
- Scott, A.C. Davydov’s soliton revisited. Phys. D 1990, 51, 333–342. [Google Scholar] [CrossRef]
- Scott, A.C.; Gratton, E.; Shyamsunder, E.; Careri, G. IR overtone spectrum of the vibrational soliton crystalline acetanilide. Phys. Rev. B 1985, 32, 5551–5553. [Google Scholar] [CrossRef]
- Scott, A.C.; Bigio, I.J.; Johnston, C.T. Polarons in acetanide. Phys. Rev. B 1989, 39, 12883–12887. [Google Scholar] [CrossRef]
- Alexander, D.M.; Krumbansl, J.A. Localized excitation in hydeogen-bonded molecular crystals. Phys. Rev. B 1986, 33, 7172–7185. [Google Scholar] [CrossRef]
- Alexander, D.M. Analog of small Holstein polaron in hydrogen-bonded amide systems. Phys. Rev. Lett. 1985, 60, 138–141. [Google Scholar] [CrossRef] [PubMed]
- Weaver, R.F. Molecular Biology; McGraw Hill: Boston, MA, USA, 2002; pp. 41–125. [Google Scholar]
- Chen, H.L. Structure and Function of Biomacromolecules; Press of Shanghu Medicine University: Shanghu, China, 1998; pp. 113–122. [Google Scholar]
- Pang, X.F. Molecular Bio-Physics; Printer of University of Electronic Science and Technology of China: Chengdu, China, 2003; pp. 68–71. [Google Scholar]
- Susi, H.; Ard, T.S.; Carroll, J. The infrared spectrum and water binding of collagen as a function of relative humidity. Biopolymers 1971, 10, 1597–1604. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lifetime at 300 K (S) | Critical Temperature (K) | Number of Amino Acids Traveled by Soliton In Lifetime | Nonlinear Interaction G (×10−21 J) | Amplitude of Soliton | Width of the Soliton (×10−10 m) | Binding Energy of Soliton (×10−21 J) | |
|---|---|---|---|---|---|---|---|
| Pang’s model | 10−9–10−10 | 320 | Several hundreds | 1.18 | 0.974 | 14.88 | −0.188 |
| Davydov model | 10−12–10−13 | <200 | <10 | 3.8 | 1.72 | 4.95 | −7.8 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pang, X.; Chen, S.; Wang, X.; Zhong, L. Influences of Electromagnetic Energy on Bio-Energy Transport through Protein Molecules in Living Systems and Its Experimental Evidence. Int. J. Mol. Sci. 2016, 17, 1130. https://doi.org/10.3390/ijms17081130
Pang X, Chen S, Wang X, Zhong L. Influences of Electromagnetic Energy on Bio-Energy Transport through Protein Molecules in Living Systems and Its Experimental Evidence. International Journal of Molecular Sciences. 2016; 17(8):1130. https://doi.org/10.3390/ijms17081130
Chicago/Turabian StylePang, Xiaofeng, Shude Chen, Xianghui Wang, and Lisheng Zhong. 2016. "Influences of Electromagnetic Energy on Bio-Energy Transport through Protein Molecules in Living Systems and Its Experimental Evidence" International Journal of Molecular Sciences 17, no. 8: 1130. https://doi.org/10.3390/ijms17081130
APA StylePang, X., Chen, S., Wang, X., & Zhong, L. (2016). Influences of Electromagnetic Energy on Bio-Energy Transport through Protein Molecules in Living Systems and Its Experimental Evidence. International Journal of Molecular Sciences, 17(8), 1130. https://doi.org/10.3390/ijms17081130