
Impact of Prematurity and Perinatal Antibiotics on the Developing Intestinal Microbiota: A Functional Inference Study

 , , ,
, , ,
Abstract
:
1. Introduction
2. Results and Discussion
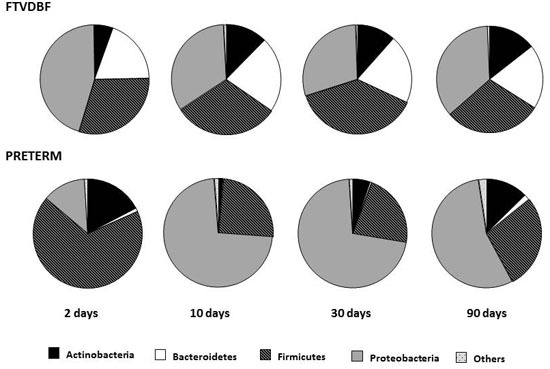
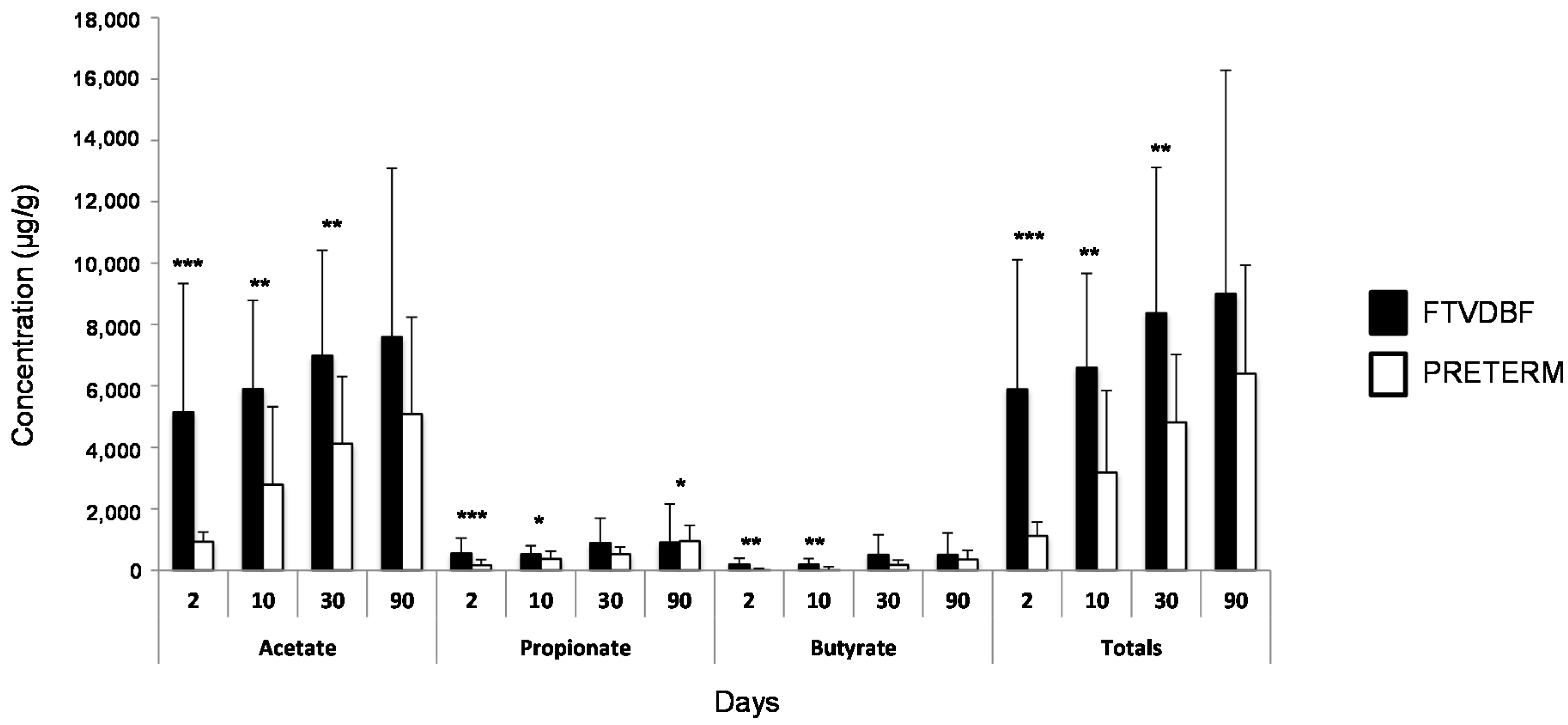
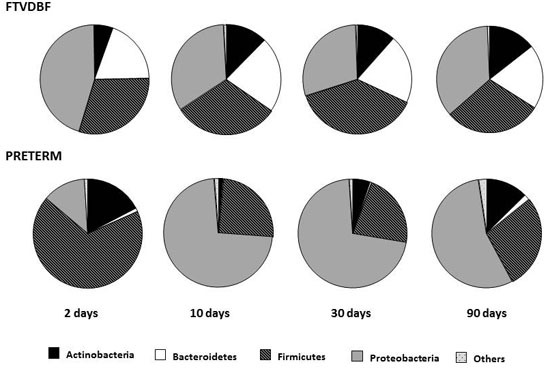
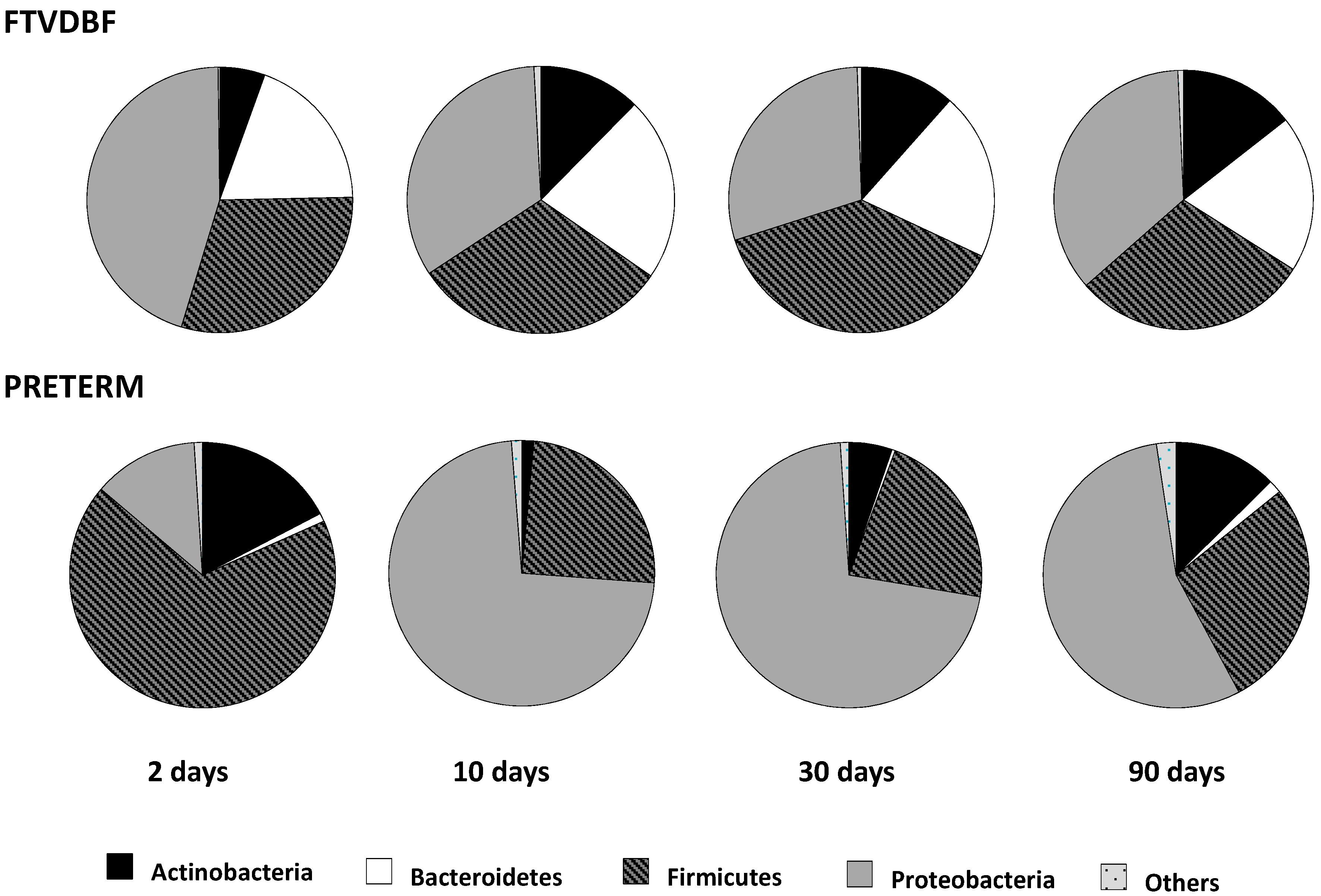
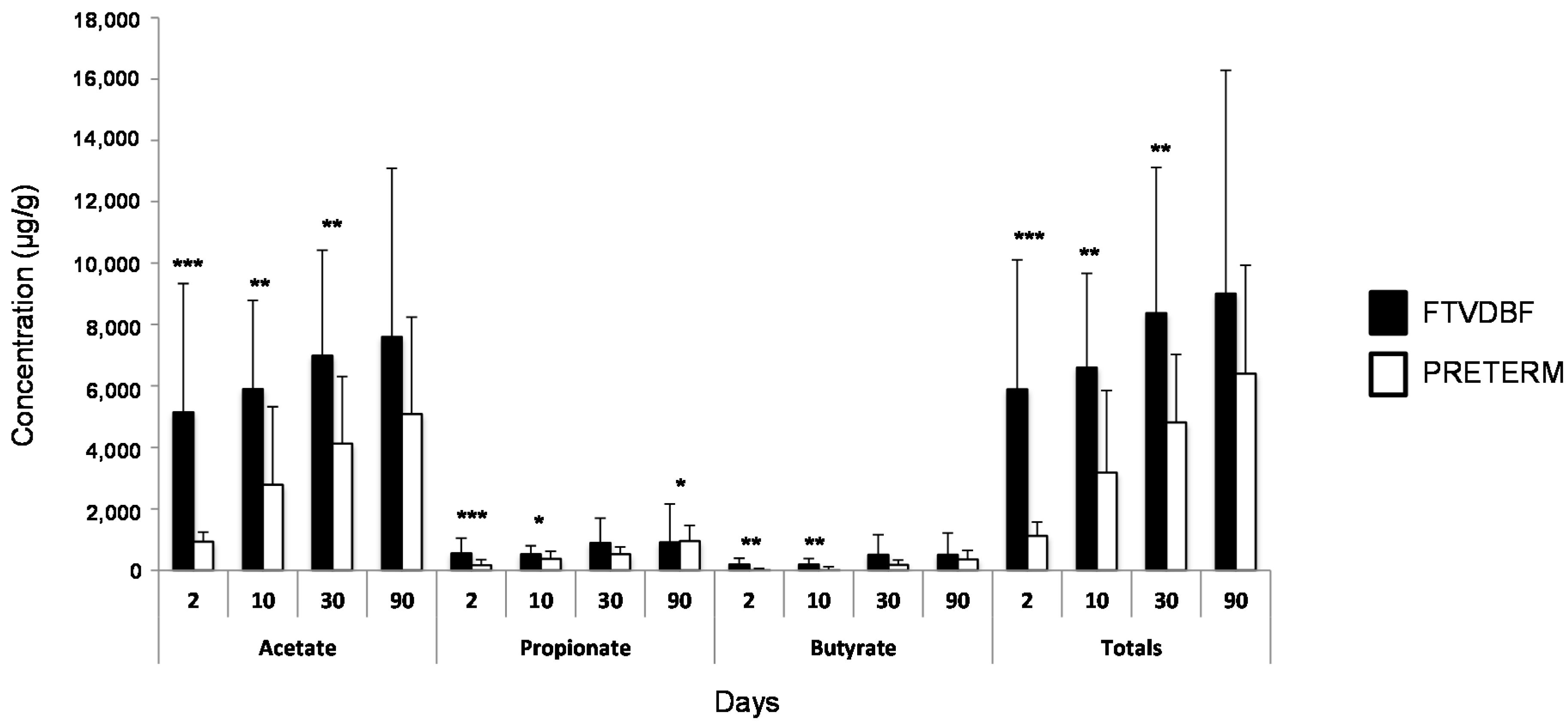
2.1. Establishment of the Intestinal Microbiota in Preterm and Full-Term Infants
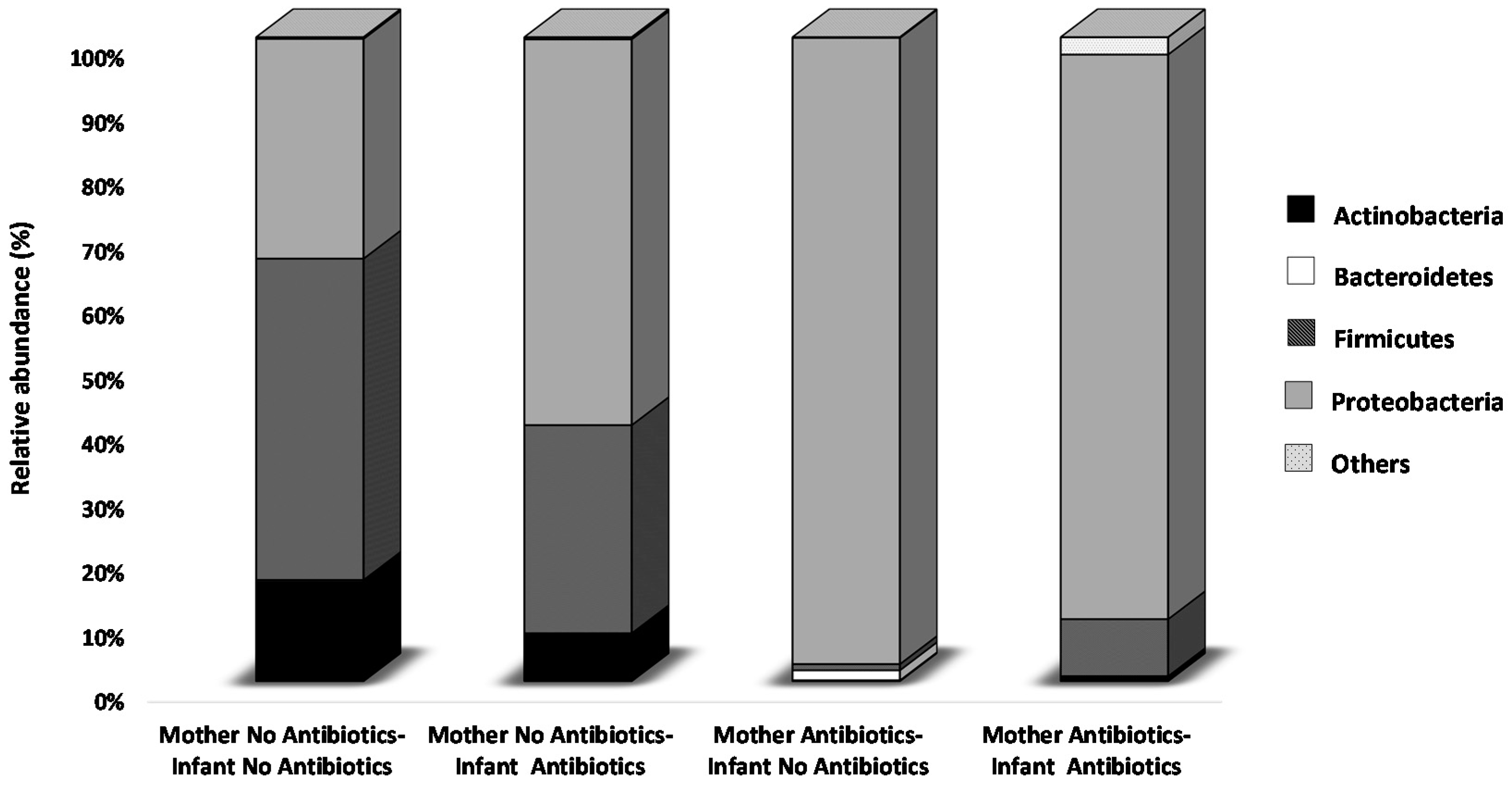
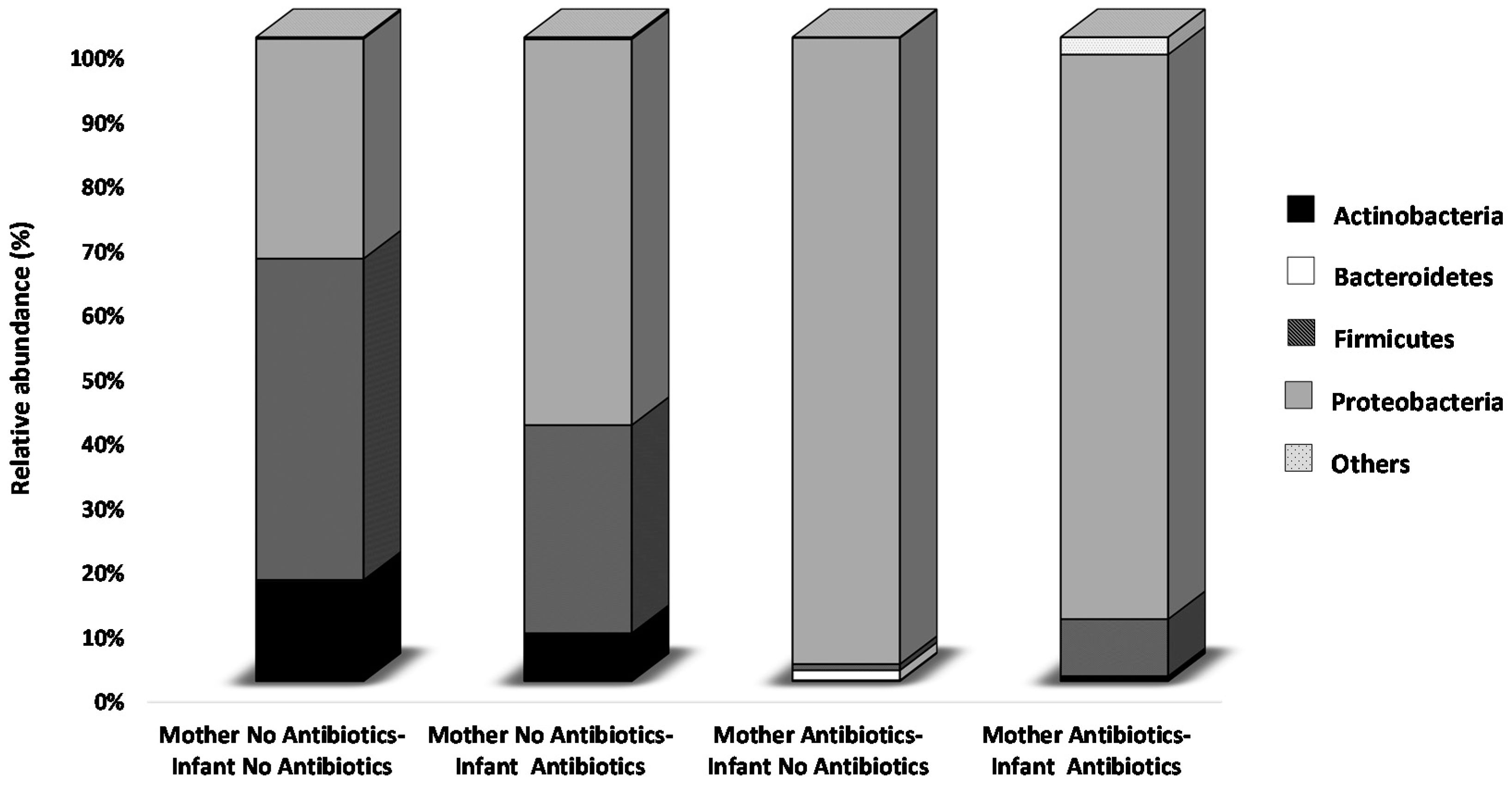
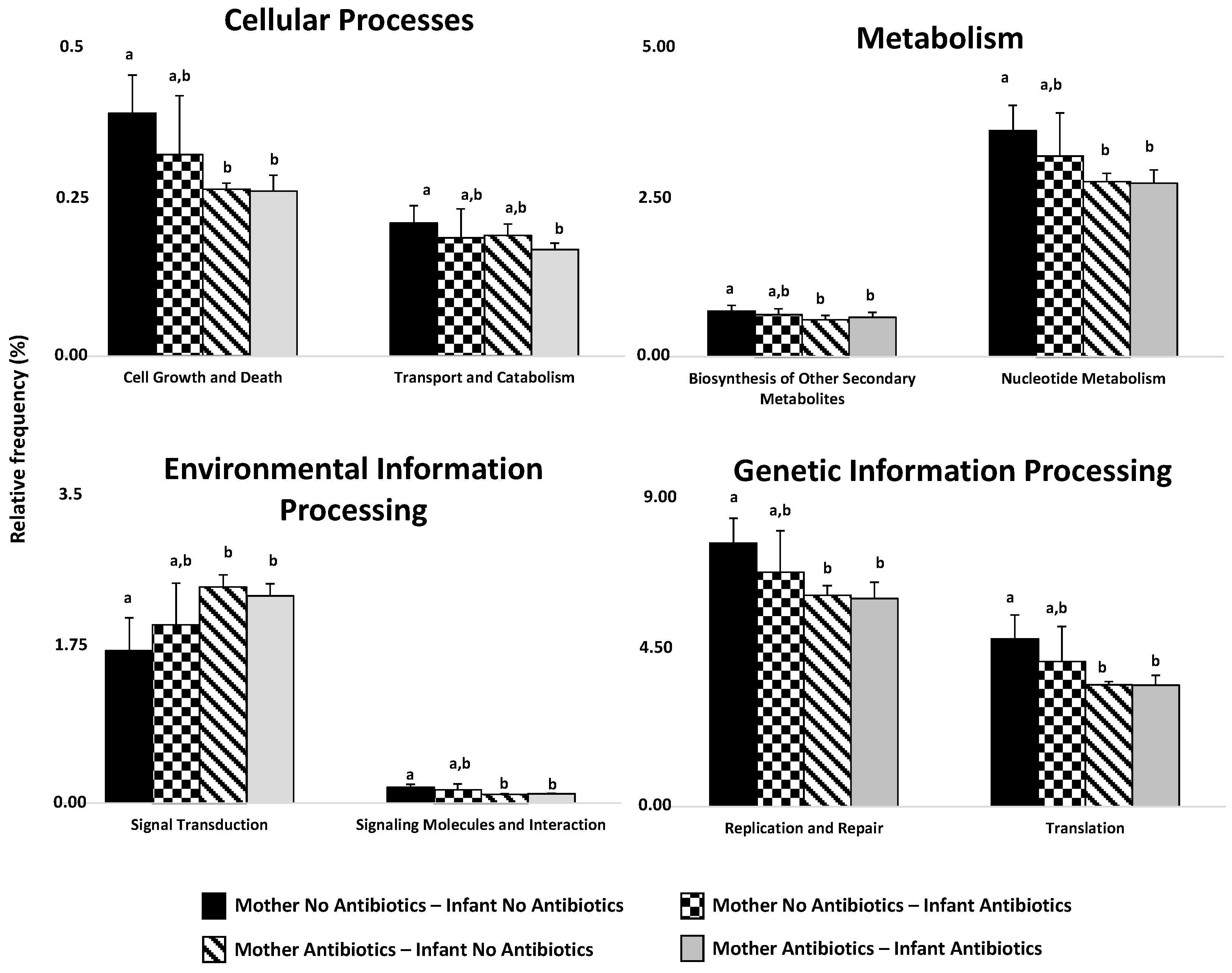
2.2. Effect of Perinatal Antibiotics on Microbiota Development in Preterm Infants
3. Materials and Methods
3.1. Volunteers and Samples
3.2. Intestinal Microbiota Analyses
3.3. Determination of SCFA in Feces
3.4. Functional Inference Analysis
3.5. Statistical Analyses
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Turnbaugh, P.J.; Hamady, M.; Yatsunenko, T.; Cantarel, B.L.; Duncan, A.; Ley, R.E.; Sogin, M.L.; Jones, W.J.; Roe, B.A.; Affourtit, J.P.; et al. A core gut microbiome in obese and lean twins. Nature 2009, 457, 480–484. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Arumugan, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Human Microbiome Project Consortium. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P.; et al. Human gut microbiome viewed across age and geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, F.H.; Tremaroli, V.; Nookaew, I.; Bergstrom, G.; Behre, C.J.; Fagerberg, B.; Nielsen, J.; Bäckhed, F. Gut metagenome in European women with normal, impaired and diabetic glucose control. Nature 2013, 498, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Sommer, F.; Bäckhed, F. The gut microbiota—Masters of host development and physiology. Nat. Rev. Microbiol. 2013, 11, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, G.P.; Lee, S.N.; Mazmanian, S.K. Gut biogeography of the bacterial microbiota. Nat. Rev. Microbiol. 2016, 14, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Matamoros, S.; Gras-Leguen, C.; Le Vacon, F.; Potel, G.; de La Cochetiere, M.F. Development of intestinal microbiota in infants and its impact on health. Trends Microbiol. 2013, 21, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Sim, K.; Powel, E.; Shaw, A.G.; McClure, Z.; Bangham, M.; Kroll, J.S. The neonatal gastrointestinal microbiota: The foundation of future health? Arch. Dis. Child. Fetal Neonatal Ed. 2013, 98, F362–F364. [Google Scholar] [CrossRef] [PubMed]
- Hansen, C.H.; Nielsen, D.S.; Kverka, M.; Zakostelska, Z.; Klimesova, K.; Hudcovic, T.; Tlaskalova-Hogenova, H.; Hansen, A.K. Patterns of early gut microbiota colonization shape future immune responses of the host. PLoS ONE 2012, 7, e34043. [Google Scholar] [CrossRef] [PubMed]
- Olszak, T.; An, D.; Zeissig, S.; Vera, M.P.; Richter, J.; Franke, A.; Glickman, J.N.; Siebert, R.; Baron, R.M.; Kasper, D.L.; et al. Microbial exposure during early life has persistent effects on natural killer T cell function. Science 2012, 336, 489–493. [Google Scholar] [CrossRef] [PubMed]
- Cox, L.M.; Yamanishi, S.; Sohn, J.; Alekseyenko, A.V.; Leung, J.M.; Cho, I.; Kim, S.G.; Li, H.; Gao, Z.; Mahana, D.; et al. Altering the intestinal microbiota during a critical developmental window has lasting metabolic consequences. Cell 2014, 158, 705–721. [Google Scholar] [CrossRef] [PubMed]
- Borre, Y.E.; O’Keeffe, G.W.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Microbiota and neurodevelopmental windows: Implications for brain disorders. Trends Mol. Med. 2014, 20, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Faa, G.; Gerosa, C.; Fanni, D.; Nemolato, S.; van Eyken, P.; Fanos, V. Factors influencing the development of a personal tailored microbiota in the neonate, with particular emphasis on antibiotic therapy. J. Matern. Fetal Neonatal Med. 2013, 26, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Bäckhed, F.; Roswall, J.; Peng, Y.; Feng, Q.; Jia, H.; Kovatcheva-Datchary, P.; Li, Y.; Xia, Y.; Xie, H.; Zhong, H.; et al. Dynamics and stabilization of the human gut microbiome during the first year of life. Cell Host Microbe 2015, 17, 690–703. [Google Scholar] [CrossRef] [PubMed]
- Del Chierico, F.; Vernocchi, P.; Petruca, A.; Paci, P.; Fuentes, S.; Pratico, G.; Capuani, G.; Masotti, A.; Reddel, S.; Russo, A.; et al. Phylogenetic and metabolic tracking of gut microbiota during perinatal development. PLoS ONE 2015, 10, e0137347. [Google Scholar]
- Jakobsson, H.E.; Abrahamsson, T.R.; Jenmalm, M.C.; Harris, K.; Quince, C.; Jernberg, C.; Björkstén, B.; Engstrand, L.; Andersson, A.F. Decreased gut microbiota diversity, delayed Bacteroidetes colonization and reduced Th1 responses in infants delivered by caesarean section. Gut 2014, 63, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Bello, M.G.; Costello, E.K.; Contreras, M.; Magris, M.; Hidalgo, G.; Fierer, N.; Knight, R. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc. Natl. Acad. Sci. USA 2010, 107, 11971–11975. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Bello, M.G.; de Jesus-Laboy, K.M.; Shen, N.; Cox, L.M.; Amir, A.; Gonzalez, A.; Bokullich, N.A.; Song, S.J.; Hoashi, M.; Rivera-Viñas, J.I.; et al. Partial restoration of the microbiota of caesarean-born infants via vaginal microbial transfer. Nat. Med. 2016, 22, 250–253. [Google Scholar] [CrossRef] [PubMed]
- Strunk, T.; Currie, A.; Richmond, P.; Simmer, K.; Burgner, D. Innate immunity in human newborn infants: Prematurity means more than immaturity. J. Matern. Fetal Neonatal Med. 2011, 24, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Arboleya, S.; Sánchez, B.; Milani, C.; Duranti, S.; Solís, G.; Fernández, N.; de los Reyes-Gavilán, C.G.; Ventura, M.; Margolles, A.; Gueimonde, M. Intestinal microbiota development in preterm neonates and effect of perinatal antibiotics. J. Pediatr. 2015, 166, 538–544. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hoenig, J.D.; Malin, K.J.; Qamar, S.; Petrof, E.O.; Sun, J.; Antonopoulos, D.A.; Chang, E.B.; Claud, E.C. 16S rRNA gene-based analysis of fecal microbiota from preterm infants with and without necrotizing enterocolitis. ISME J. 2009, 3, 944–954. [Google Scholar] [CrossRef] [PubMed]
- Arboleya, S.; Binetti, A.; Salazar, N.; Fernández, N.; Solís, G.; Hernández-Barranco, A.; Margolles, A.; de los Reyes-Gavilán, C.G.; Gueimonde, M. Establishment and development of intestinal microbiota in preterm neonates. FEMS Microbiol. Ecol. 2012, 79, 763–772. [Google Scholar] [CrossRef] [PubMed]
- Mshvildadze, M.; Neu, J.; Shuster, J.; Theriaque, D.; Li, N.; Mai, W. Intestinal microbial ecology in premature infants assessed by non-culture-based techniques. J. Pediatr. 2010, 156, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Arboleya, S.; Solís, G.; Fernández, N.; de los Reyes-Gavilán, C.G.; Gueimonde, M. Facultative to strict anaerobes ratio in the preterm infant microbiota: A target for intervention? Gut Microbes 2012, 3, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Barrett, E.; Kerr, C.; Murphy, K.; O’Sullivan, O.; Ryan, C.A.; Dempsey, E.M.; Murphy, B.P.; O’Toole, P.W.; Cotter, P.D.; Fitzgerald, G.F.; et al. The individual-specific and diverse nature of the preterm infant microbiota. Arch. Dis. Child. Fetal Neonatal Ed. 2013, 98, F334–F340. [Google Scholar] [CrossRef] [PubMed]
- Morrow, A.L.; Lagomarcino, A.J.; Schibler, K.R.; Taft, D.H.; Yo, Z.; Wang, B.; Altaye, M.; Wagner, M.; Gevers, D.; Ward, D.V.; et al. Early microbial and metabolomic signatures predict later onset of necrotizing enterocolitis in preterm infants. Microbiome 2013, 1. [Google Scholar] [CrossRef] [PubMed]
- Aloisio, I.; Quagliariello, A.; de Fanti, S.; Luiselli, D.; de Filippo, C.; Albanese, D.; Corvaglia, L.T.; Faldella, G.; di Gioia, D. Evaluation of the effects of intrapartum antibiotic prophylaxix on newborn intestinal microbiota using a sequencing approach targeted to multi hypervariate 16S rDNA regions. Appl. Microbiol. Biotechnol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Koenig, J.E.; Spor, A.; Scalfone, N.; Fricker, A.D.; Stombaugh, J.; Knight, R.; Angenent, L.T.; Ley, R.E. Succession of microbial consortia in the developing infant gut microbiome. Proc. Natl. Acad. Sci. USA 2011, 108, 4578–4585. [Google Scholar] [CrossRef] [PubMed]
- Arboleya, S.; Ang, L.; Margolles, A.; Yiyuan, L.; Dongya, Z.; Liang, X.; Solís, G.; Fernández, N.; de los Reyes-Gavilán, C.G.; Gueimonde, M. Deep 16S rRNA metagenomics and quantitative PCR analyses of the premature infant fecal microbiota. Anaerobe 2012, 18, 378–380. [Google Scholar] [CrossRef] [PubMed]
- Stewart, C.J.; Marrs, E.C.; Nelson, A.; Lanyon, C.; Perry, J.D.; Embleton, N.D.; Cummings, S.P.; Berrington, J.E. Development of the preterm gut microbiome in twins at risk of necrotising enterocolitis and sepsis. PLoS ONE 2013, 8, e73465. [Google Scholar] [CrossRef] [PubMed]
- Avershina, E.; Storro, O.; Oien, T.; Johnsen, R.; Pope, P.; Rudi, K. Major fecal microbiota shifts in composition and diversity with age in a geographically restricted cohort of mothers and their children. FEMS Microbiol. Ecol. 2014, 87, 280–290. [Google Scholar] [CrossRef] [PubMed]
- Morgan, X.C.; Tickle, T.L.; Sokol, H.; Gevers, D.; Devaney, K.L.; Ward, D.V.; Reyes, J.A.; Shah, S.A.; LeLeiko, N.; Snapper, S.B.; et al. Dysfunction of the intestinal microbiome in inflammatory bowel disease and treatment. Genome Biol. 2012, 13, R79. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Kobayashi, T.; Songjinda, P.; Tateyama, A.; Tsubouchi, M.; Kiyohara, C.; Shirakawa, T.; Sonomoto, K.; Nakayama, J. Influence of antibiotic exposure in the early postnatal period on the development of intestinal microbiota. FEMS Immunol. Med. Microbiol. 2009, 56, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Fallani, M.; Young, D.; Scott, J.; Norin, E.; Amarri, S.; Adam, R.; Aguilera, M.; Khanna, S.; Gil, A.; Edwards, C.A.; et al. Intestinal microbiota of 6-week-old infants across Europe: Geographic influence beyond delivery mode, breast-feeding and antibiotics. J. Pediatr. Gastroenterol. Nutr. 2010, 51, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Fouhy, F.; Guinane, C.M.; Hussey, S.; Wall, R.; Ryan, C.A.; Dempsey, E.M.; Murphy, B.; Ross, R.P.; Fitzgerald, G.F.; Stanton, C.; et al. High-Throughput sequencing reveals the incomplete, short-term recovery of infant gut microbiota following parenteral antibiotic treatment with ampicilin and gentamicin. Antimicrob. Agents Chemother. 2012, 56, 5811–5820. [Google Scholar] [CrossRef] [PubMed]
- Penders, J.; Thijs, C.; Vink, C.; Stelma, F.F.; Snijders, B.; Kummeling, I.; van den Brandt, P.A.; Stobberingh, E.E. Factors influencing the composition of the intestinal microbiota in early infancy. Pediatrics 2006, 118, 511–521. [Google Scholar] [CrossRef] [PubMed]
- Azad, M.B.; Konya, T.; Persaud, R.R.; Guttman, D.S.; Chari, R.S.; Field, C.J.; Sears, M.R.; Mandhane, P.J.; Turvey, S.E.; Subbarao, P.; et al. Impact of maternal intrapartum antibiotics, method of birth and breastfeeding on gut microbiota during the first year of life: A prospective cohort study. BJOG Int. J. Obstet. Gynaecol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Russell, S.L.; Gold, M.J.; Hartmann, M.; Willing, B.P.; Thorson, L.; Wlodarska, M.; Gill, N.; Blanchet, M.R.; Mohn, W.W.; McNagny, K.M.; et al. Early life antibiotic-driven changes in microbiota enhance susceptibility to allergic asthma. EMBO Rep. 2012, 13, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Chu, S.; Yu, H.; Chen, Y.; Chen, Q.; Wang, B.; Zhang, J. Periconceptional and gestational exposure to antibiotics and childhood asthma. PLoS ONE 2015, 10, e0140443. [Google Scholar] [CrossRef] [PubMed]
- Van Dyke, M.K.; Phares, C.R.; Lynfield, R.; Thomas, A.R.; Arnold, K.E.; Craig, A.S.; Mohle-Boetani, J.; Gershman, K.; Schaffner, W.; Petit, S.; et al. Evaluation of universal antenatal screening for group B streptococcus. N. Engl. J. Med. 2009, 360, 2626–2636. [Google Scholar] [CrossRef] [PubMed]
- Kenyon, S.; Boulvain, M.; Neilson, J.P. Antibiotics for preterm rupture of membranes. Cochrane Database Syst. Rev. 2010, 8, CD001058. [Google Scholar] [PubMed]
- Allen, V.M.; Yudin, M.H.; Bouchard, C.; Boucher, M.; Caddy, S.; Castillo, E.; Money, D.M.; Murphy, K.E.; Ogilvie, G.; Paquet, C.; et al. Management of group B streptococcal bacteriuria in pregnancy. J. Obstet. Gynaecol. Can. 2012, 34, 482–486. [Google Scholar] [CrossRef]
- King, J.F.; Flenady, V.; Murray, L. Prophylactic antibiotics for inhibiting preterm labour with intact membranes. Cochrane Database Syst. Rev. 2002, 4, CD000246. [Google Scholar] [PubMed]
- Russell, B.A.R.; Murch, S.H. Could peripartum antibiotics have delayed health consequences for the infant? BJOG Int. J. Obstet. Gynaecol. 2006, 113, 758–765. [Google Scholar] [CrossRef] [PubMed]
- Renz, H.; Brandtzaeg, P.; Hornef, M. The impact of perinatal immune development on mucosal homeostasis and chronic inflammation. Nat. Rev. Immunol. 2012, 12, 9–23. [Google Scholar] [CrossRef] [PubMed]
- Milani, C.; Hevia, A.; Foroni, E.; Duranti, S.; Turroni, F.; Lugli, G.A.; Sanchez, B.; Martín, R.; Gueimonde, M.; van Sinderen, D.; et al. Assessing the fecal microbiota: An optimized Ion Torrent 16S rRNA gene-based analysis protocol. PLoS ONE 2013, 8, e68739. [Google Scholar] [CrossRef] [PubMed]
- Langille, M.G.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- McDonald, D.; Clemente, J.C.; Kuczynski, J.; Rideout, J.R.; Stombaugh, J.; Wendel, D.; Wilke, A.; Huse, S.; Hufnagle, J.; Meyer, F.; et al. The Biological Observation Matrix (BIOM) format or: How I learned to stop worrying and love the ome-ome. Gigascience 2012, 1, 7. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Beiko, R.G. Identifying biologically relevant differences between metagenomic communities. Bioinformatics 2010, 26, 715–721. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| KEGG Pathways | Infants | Day 2 | Day 10 | Day 30 | Day 90 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean ± SD | p (t-Test) | q-Value | Mean ± SD | p (t-Test) | q-Value | Mean ± SD | p (t-Test) | q-Value | Mean ± SD | p (t-Test) | q-Value | ||
| Metabolism | |||||||||||||
| Amino Acid Metabolism | Preterm | 8.68 ± 0.58 | ns | 0.558 | 8.29 ± 0.64 | * | 0.031 | 8.35 ± 0.48 | * | 0.053 | 8.61 ± 0.66 | ns | 0.263 |
| Term | 8.43 ± 1.02 | 9.06 ± 0.78 | 8.86 ± 0.61 | 9.00 ± 0.67 | |||||||||
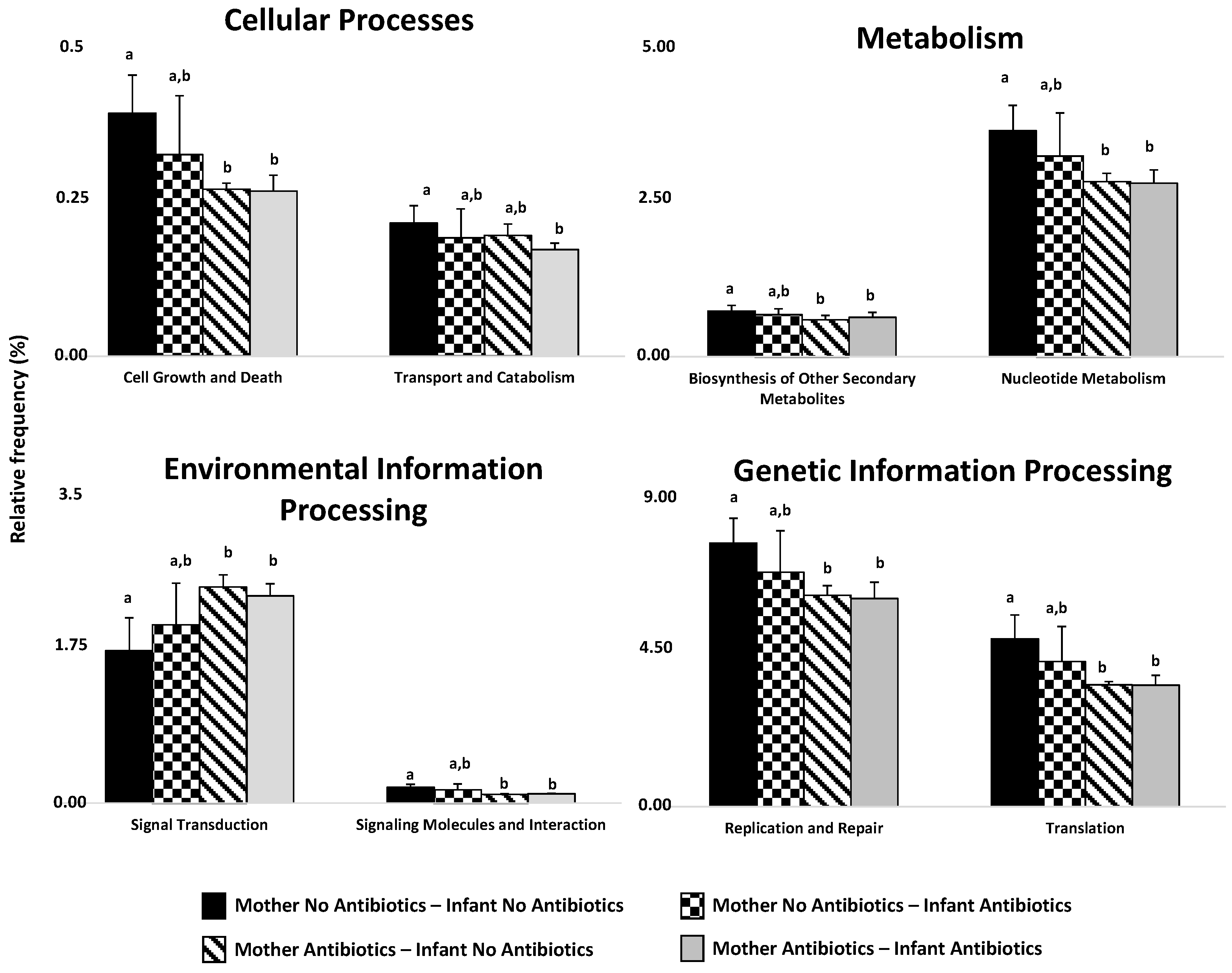
| Biosynthesis of Other Secondary Metabolites | Preterm | 0.77 ± 0.06 | ns | 0.776 | 0.66 ± 0.08 | * | 0.037 | 0.70 ± 0.10 | * | 0.056 | 0.75 ± 0.11 | ns | 0.391 |
| Term | 0.74 ± 0.24 | 0.86 ± 0.24 | 0.85 ± 0.19 | 0.82 ± 0.22 | |||||||||
| Carbohydrate Metabolism | Preterm | 11.53 ± 0.64 | ns | 0.383 | 10.99 ± 0.77 | ns | 0.902 | 10.86 ± 0.7 | * | 0.131 | 10.88 ± 0.60 | ns | 0.497 |
| Term | 11.16 ± 1.08 | 11.04 ± 0.80 | 11.31 ± 0.73 | 11.11 ± 0.76 | |||||||||
| Energy Metabolism | Preterm | 4.93 ± 0.19 | ns | 0.544 | 4.72 ± 0.12 | * | 0.033 | 4.75 ± 0.13 | * | 0.029 | 4.91 ± 0.31 | * | 0.550 |
| Term | 5.08 ± 0.63 | 5.2 ± 0.52 | 5.13 ± 0.42 | 5.26 ± 0.46 | |||||||||
| Enzyme Families | Preterm | 2.09 ± 0.07 | ns | 0.809 | 2.03 ± 0.05 | * | 0.039 | 2.03 ± 0.04 | * | 0.023 | 2.09 ± 0.07 | ns | 0.327 |
| Term | 2.08 ± 0.15 | 2.13 ± 0.10 | 2.13 ± 0.09 | 2.13 ± 0.09 | |||||||||
| Glycan Biosynthesis and Metabolism | Preterm | 1.89 ± 0.28 | * | 0.027 | 2.25 ± 0.38 | * | 0.112 | 2.35 ± 0.25 | ns | 0.307 | 2.22 ± 0.31 | ns | 0.343 |
| Term | 2.73 ± 0.82 | 2.71 ± 0.78 | 2.60 ± 0.67 | 2.50 ± 0.73 | |||||||||
| Lipid Metabolism | Preterm | 3.08 ± 0.24 | * | 0.006 | 2.89 ± 0.30 | ns | 0.493 | 2.78 ± 0.12 | ns | 0.561 | 2.85 ± 0.17 | ns | 0.640 |
| Term | 2.83 ± 0.16 | 2.81 ± 0.22 | 2.82 ± 0.19 | 2.90 ± 0.22 | |||||||||
| Metabolism of Cofactors and Vitamins | Preterm | 3.55 ± 0.55 | * | 0.044 | 3.80 ± 0.19 | * | 0.032 | 3.88 ± 0.26 | ns | 0.291 | 3.89 ± 0.13 | ns | 0.387 |
| Term | 4.07 ± 0.52 | 4.14 ± 0.39 | 4.02 ± 0.34 | 4.03 ± 0.18 | |||||||||
| Metabolism of Other Amino Acids | Preterm | 1.66 ± 0.06 | * | 0.230 | 1.68 ± 0.08 | ns | 0.892 | 1.70 ± 0.06 | ns | 0.367 | 1.65 ± 0.13 | ns | 0.510 |
| Term | 1.63 ± 0.07 | 1.69 ± 0.13 | 1.68 ± 0.07 | 1.61 ± 0.14 | |||||||||
| Metabolism of Terpenoids and Polyketides | Preterm | 1.74 ± 0.13 | * | 0.052 | 1.50 ± 0.19 | ns | 0.620 | 1.44 ± 0.08 | * | 0.034 | 1.46 ± 0.08 | ns | 0.295 |
| Term | 1.57 ± 0.20 | 1.53 ± 0.14 | 1.53 ± 0.09 | 1.52 ± 0.12 | |||||||||
| Nucleotide Metabolism | Preterm | 4.27 ± 0.52 | * | 0.125 | 3.23 ± 0.63 | * | 0.030 | 3.27 ± 0.57 | * | 0.064 | 3.40 ± 0.46 | ns | 0.279 |
| Term | 3.85 ± 0.63 | 3.78 ± 0.57 | 3.70 ± 0.51 | 3.67 ± 0.40 | |||||||||
| Xenobiotics Biodegradation and Metabolism | Preterm | 2.57 ± 0.35 | * | 0.000 | 2.23 ± 0.35 | * | 0.082 | 2.18 ± 0.28 | * | 0.036 | 2.08 ± 0.41 | ns | 0.269 |
| Term | 1.80 ± 0.15 | 1.84 ± 0.35 | 1.87 ± 0.31 | 1.80 ± 0.39 | |||||||||
| Cellular Processes | |||||||||||||
| Cell Growth and Death | Preterm | 0.50 ± 0.09 | * | 0.064 | 0.30 ± 0.09 | * | 0.034 | 0.30 ± 0.08 | * | 0.027 | 0.35 ± 0.10 | ns | 0.289 |
| Term | 0.40 ± 0.11 | 0.40 ± 0.10 | 0.40 ± 0.08 | 0.41 ± 0.08 | |||||||||
| Cell Motility | Preterm | 1.12 ± 0.47 | * | 0.151 | 2.06 ± 1.10 | ns | 0.496 | 1.98 ± 0.97 | * | 0.121 | 1.84 ± 0.81 | ns | 0.930 |
| Term | 1.85 ± 1.31 | 1.67 ± 1.30 | 1.51 ± 0.59 | 1.78 ± 0.91 | |||||||||
| Transport and Catabolism | Preterm | 0.20 ± 0.03 | * | 0.200 | 0.18 ± 0.03 | * | 0.049 | 0.19 ± 0.03 | * | 0.085 | 0.20 ± 0.06 | ns | 0.317 |
| Term | 0.29 ± 0.18 | 0.32 ± 0.19 | 0.29 ± 0.17 | 0.31 ± 0.18 | |||||||||
| Genetic Information Processing | |||||||||||||
| Folding, Sorting and Degradation | Preterm | 2.20 ± 0.05 | * | 0.100 | 2.18 ± 0.14 | * | 0.120 | 2.19 ± 0.10 | * | 0.112 | 2.20 ± 0.11 | ns | 0.749 |
| Term | 2.40 ± 0.28 | 2.34 ± 0.28 | 2.31 ± 0.20 | 2.32 ± 0.14 | |||||||||
| Replication and Repair | Preterm | 8.76 ± 1.06 | * | 0.126 | 6.52 ± 1.21 | * | 0.046 | 6.50 ± 0.98 | * | 0.035 | 7.00 ± 1.14 | ns | 0.360 |
| Term | 7.91 ± 1.29 | 7.88 ± 1.30 | 7.60 ± 1.07 | 7.76 ± 0.99 | |||||||||
| Transcription | Preterm | 2.70 ± 0.19 | ns | 0.839 | 3.02 ± 0.18 | * | 0.029 | 3.00 ± 0.19 | * | 0.191 | 3.05 ± 0.20 | ns | 0.261 |
| Term | 2.72 ± 0.29 | 2.73 ± 0.31 | 2.86 ± 0.27 | 2.90 ± 0.26 | |||||||||
| Translation | Preterm | 5.70 ± 0.82 | * | 0.053 | 3.93 ± 0.99 | * | 0.039 | 3.93 ± 0.82 | * | 0.045 | 4.19 ± 0.73 | ns | 0.266 |
| Term | 4.74 ± 1.08 | 4.81 ± 0.99 | 4.68 ± 0.80 | 4.57 ± 0.58 | |||||||||
| Environmental Information Processing | |||||||||||||
| Membrane Transport | Preterm | 15.15 ± 1.01 | ns | 0.345 | 16.86 ± 1.57 | * | 0.048 | 16.93 ± 1.31 | * | 0.075 | 16.54 ± 1.59 | ns | 0.292 |
| Term | 13.91 ± 3.38 | 14.05 ± 3.71 | 14.75 ± 3.11 | 14.53 ± 2.68 | |||||||||
| Signal Transduction | Preterm | 1.56 ± 0.43 | * | 0.105 | 2.50 ± 0.49 | * | 0.066 | 2.44 ± 0.47 | * | 0.009 | 2.23 ± 0.47 | ns | 0.283 |
| Term | 2.05 ± 0.71 | 1.89 ± 0.58 | 1.96 ± 0.31 | 1.93 ± 0.49 | |||||||||
| Signaling Molecules and Interaction | Preterm | 0.27 ± 0.07 | * | 0.147 | 0.16 ± 0.08 | * | 0.071 | 0.15 ± 0.06 | * | 0.039 | 0.17 ± 0.05 | ns | 0.353 |
| Term | 0.22 ± 0.08 | 0.22 ± 0.08 | 0.21 ± 0.06 | 0.20 ± 0.07 | |||||||||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arboleya, S.; Sánchez, B.; Solís, G.; Fernández, N.; Suárez, M.; Hernández-Barranco, A.M.; Milani, C.; Margolles, A.; De los Reyes-Gavilán, C.G.; Ventura, M.; et al. Impact of Prematurity and Perinatal Antibiotics on the Developing Intestinal Microbiota: A Functional Inference Study. Int. J. Mol. Sci. 2016, 17, 649. https://doi.org/10.3390/ijms17050649
Arboleya S, Sánchez B, Solís G, Fernández N, Suárez M, Hernández-Barranco AM, Milani C, Margolles A, De los Reyes-Gavilán CG, Ventura M, et al. Impact of Prematurity and Perinatal Antibiotics on the Developing Intestinal Microbiota: A Functional Inference Study. International Journal of Molecular Sciences. 2016; 17(5):649. https://doi.org/10.3390/ijms17050649
Chicago/Turabian StyleArboleya, Silvia, Borja Sánchez, Gonzalo Solís, Nuria Fernández, Marta Suárez, Ana M. Hernández-Barranco, Christian Milani, Abelardo Margolles, Clara G. De los Reyes-Gavilán, Marco Ventura, and et al. 2016. "Impact of Prematurity and Perinatal Antibiotics on the Developing Intestinal Microbiota: A Functional Inference Study" International Journal of Molecular Sciences 17, no. 5: 649. https://doi.org/10.3390/ijms17050649
APA StyleArboleya, S., Sánchez, B., Solís, G., Fernández, N., Suárez, M., Hernández-Barranco, A. M., Milani, C., Margolles, A., De los Reyes-Gavilán, C. G., Ventura, M., & Gueimonde, M. (2016). Impact of Prematurity and Perinatal Antibiotics on the Developing Intestinal Microbiota: A Functional Inference Study. International Journal of Molecular Sciences, 17(5), 649. https://doi.org/10.3390/ijms17050649