
Biochemical Characterization and Complete Conversion of Coenzyme Specificity of Isocitrate Dehydrogenase from Bifidobacterium longum

Abstract
:
1. Introduction
2. Results
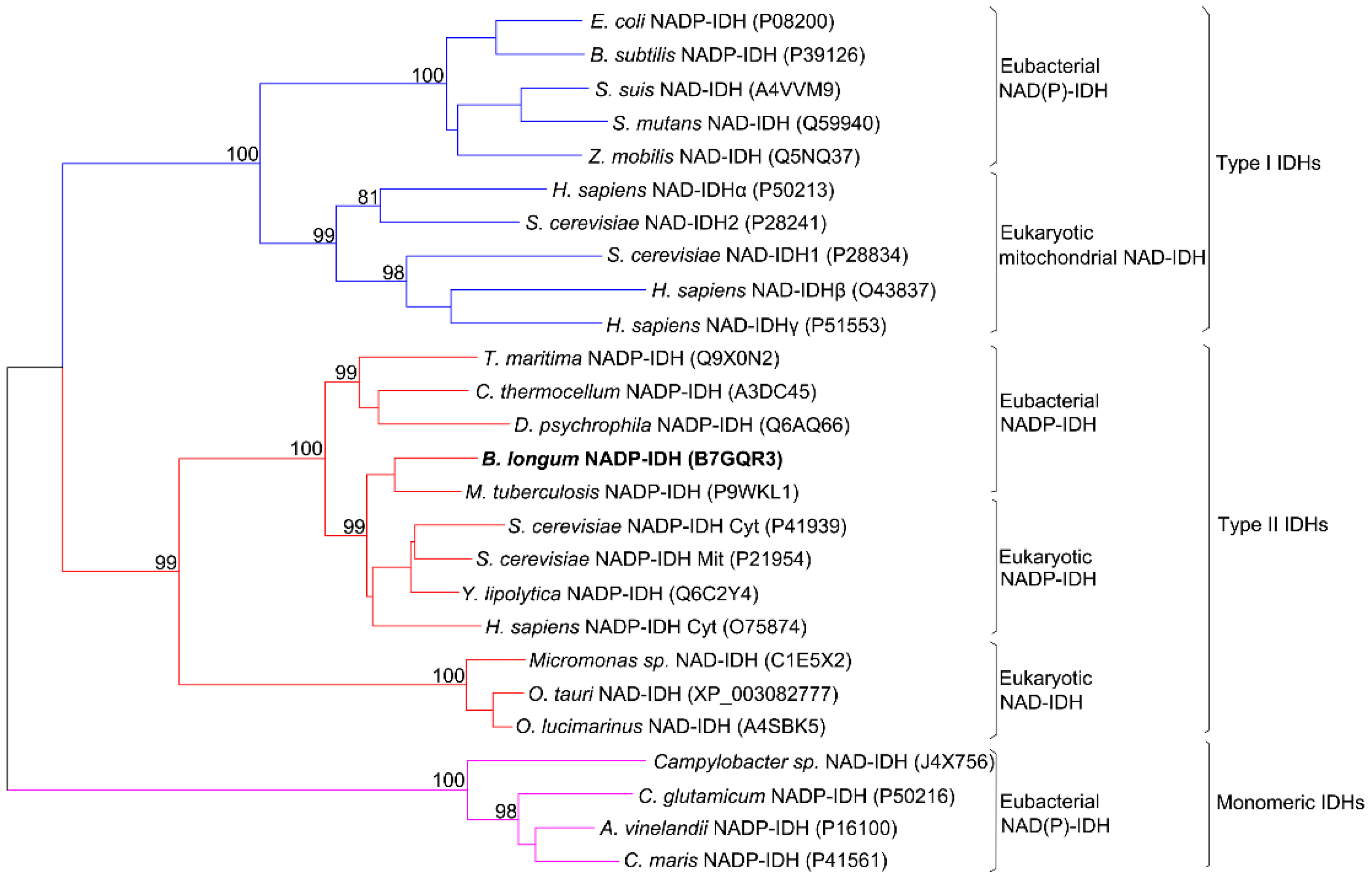
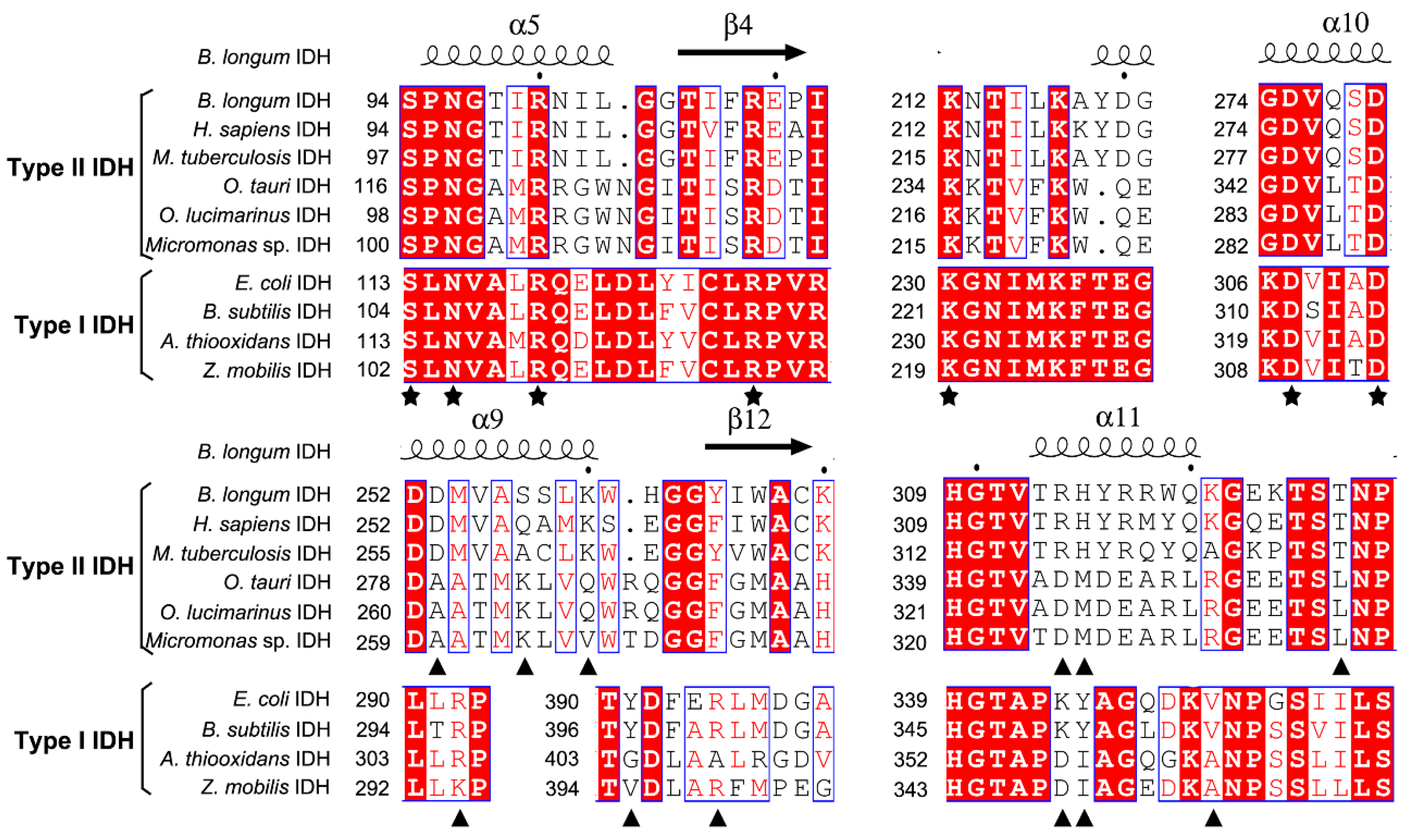
2.1. Sequence Alignment
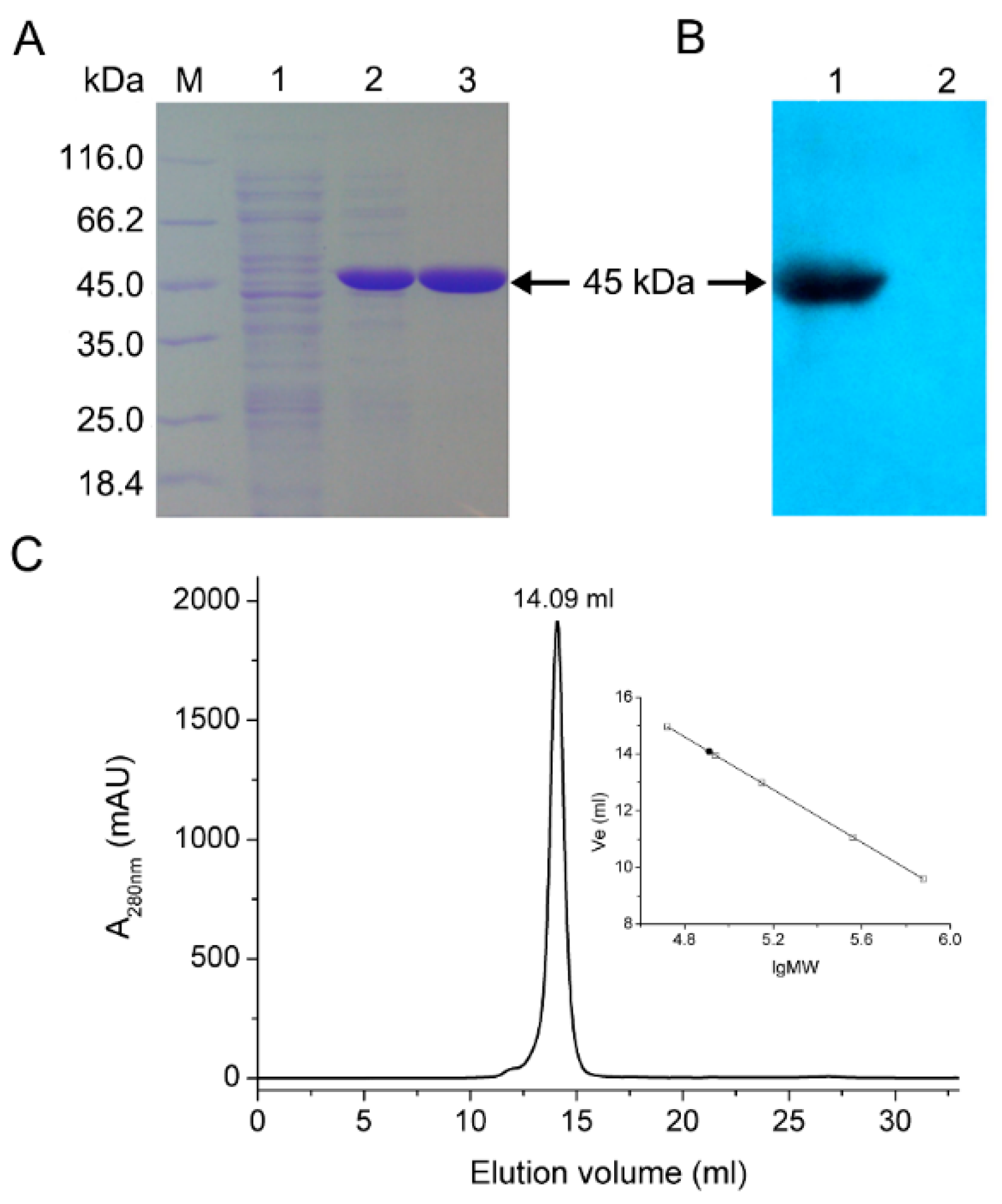
2.2. Expression and Purification
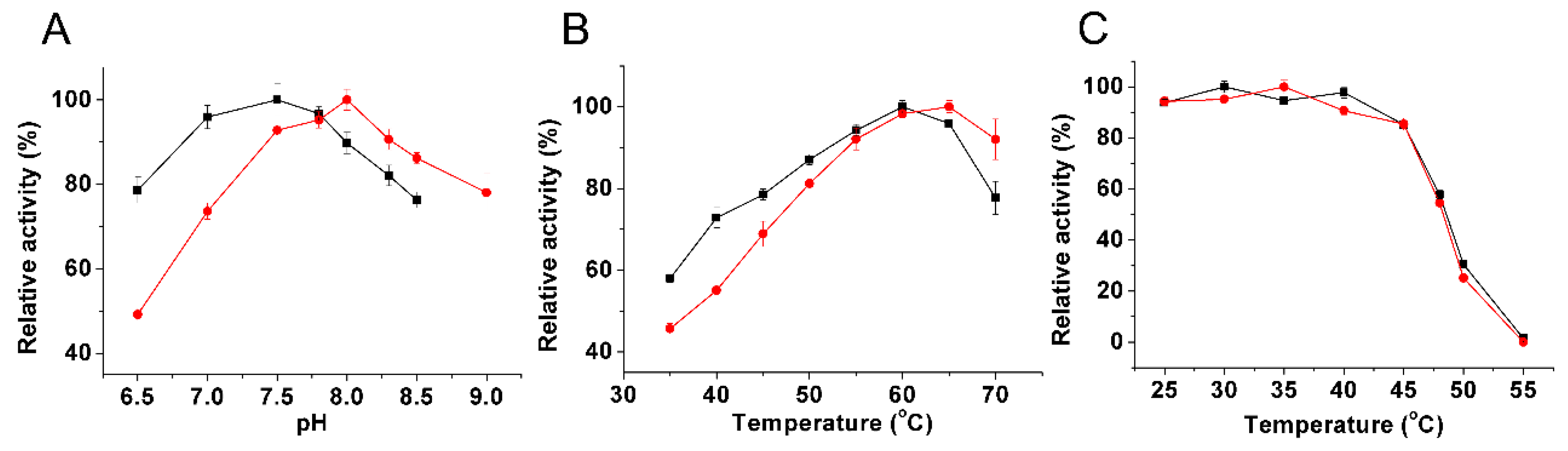
2.3. Kinetic Characterization
2.4. Switch of Coenzyme Specificity
3. Discussion
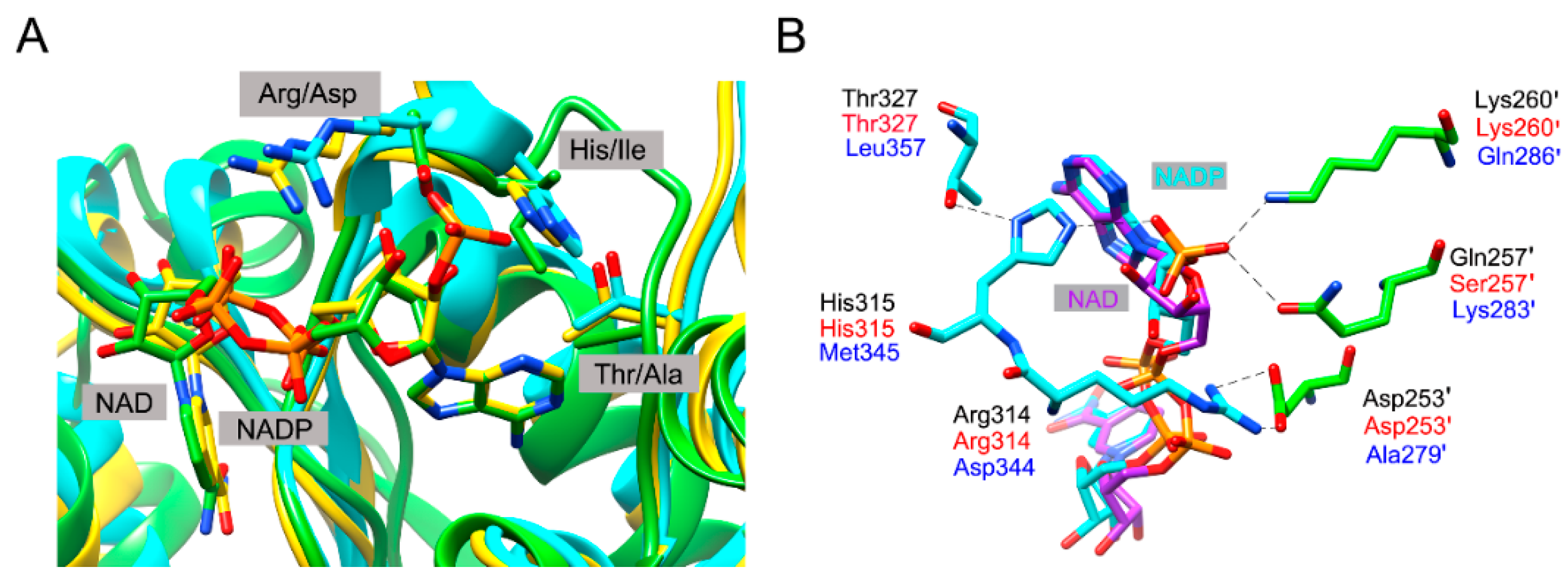
3.1. Coenzyme Specificity Determinants of BlIDH
3.2. Evolutionary Implications for NADP+ Use by Prokaryotic Type II IDHs
4. Experimental Section
4.1. Sequence Analysis
4.2. Bacterial Srains and Reagents
4.3. Recombinant Plasmid Construction
4.4. Site-Directed Mutagenesis
4.5. Overexpression and Purification
4.6. SDS-PAGE and Western Blotting
4.7. Gel Filtration Chromatography
4.8. Circular Dichroism Spectroscopy
4.9. Enzyme Assays and Kinetic Characterization
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| PCR | polymerase chain reaction |
| LB | Luria-Bertani |
| IPTG | isopropyl-1-thio-β-d-galactopyranoside |
| SDS-PAGE | sodium dodecyl sulfate polyacrylamide gel electrophoresis |
| CD | circular dichroism |
| SEC | size exclusion chromatography |
| IDH | isocitrate dehydrogenase |
| NADP+ | nicotinamide adenine dinucleotide phosphate |
| NAD+ | nicotinamide adenine dinucleotide |
| NADP-IDH | NADP+-dependent isocitrate dehydrogenase |
| NAD-IDH | NAD+-dependent isocitrate dehydrogenase |
| BlIDH | isocitrate dehydrogenase from Bifidobacterium longum |
| EcIDH | NADP-IDH from Escherichia coli |
| HcIDH | human cytosolic NADP-IDH |
| AtIDH | NAD-IDH from Acidithiobacillus thiooxidans |
| OtIDH | NAD-IDH from Ostreococcus tauri |
| Km | Michaelis constant |
| kcat | catalytic rate constant |
References
- Sivaraman, J.; Li, Y.; Banks, J.; Cane, D.E.; Matte, A.; Cygler, M. Crystal structure of Escherichia coli PdxA, an enzyme involved in the pyridoxal phosphate biosynthesis pathway. J. Biol. Chem. 2003, 278, 43682–43690. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, J.; Kobashi, N.; Nishiyama, M.; Yamane, H. Characterization of homoisocitrate dehydrogenase involved in lysine biosynthesis of an extremely thermophilic bacterium, Thermus thermophilus HB27, and evolutionary implication of β-decarboxylating dehydrogenase. J. Biol. Chem. 2003, 278, 1864–1871. [Google Scholar] [CrossRef] [PubMed]
- Tipton, P.A.; Beecher, B.S. Tartrate dehydrogenase, a new member of the family of metal-dependent decarboxylating R-hydroxyacid dehydrogenases. Arch. Biochem. Biophys. 1994, 313, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Dean, A.M.; Koshland, D.E., Jr. Kinetic mechanism of Escherichia coli isocitrate dehydrogenase. Biochemistry 1993, 32, 9302–9309. [Google Scholar] [CrossRef] [PubMed]
- Fernie, A.R.; Carrari, F.; Sweetlove, L.J. Respiratory metabolism: Glycolysis, the TCA cycle and mitochondrial electron transport. Curr. Opin. Plant Biol. 2004, 7, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Spaans, S.K.; Weusthuis, R.A.; van der Oost, J.; Kengen, S.W. NADPH-generating systems in bacteria and archaea. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Koh, H.J.; Park, D.C.; Song, B.J.; Huh, T.L.; Park, J.W. Cytosolic NADP+-dependent isocitrate dehydrogenase status modulates oxidative damage to cells. Free Radic. Biol. Med. 2002, 32, 1185–1196. [Google Scholar] [CrossRef]
- Jo, S.H.; Son, M.K.; Koh, H.J.; Lee, S.M.; Song, I.H.; Kim, Y.O.; Lee, Y.S.; Jeong, K.S.; Kim, W.B.; Park, J.W.; et al. Control of mitochondrial redox balance and cellular defense against oxidative damage by mitochondrial NADP+-dependent isocitrate dehydrogenase. J. Biol. Chem. 2001, 276, 16168–16176. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A.L.; Holmen, S.L.; Colman, H. IDH1 and IDH2 mutations in gliomas. Curr. Neurol. Neurosci. Rep. 2013, 13, 345. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Liu, Y.; Lu, C.; Cross, J.R.; Morris, J.P.; Shroff, A.S.; Ward, P.S.; Bradner, J.E.; Thompson, C.; Lowe, S.W. Cancer-associated IDH2 mutants drive an acute myeloid leukemia that is susceptible to Brd4 inhibition. Genes Dev. 2013, 27, 1974–1985. [Google Scholar] [CrossRef] [PubMed]
- Dang, L.; White, D.W.; Gross, S.; Bennett, B.D.; Bittinger, M.A.; Driggers, E.M.; Fantin, V.R.; Jang, H.G.; Jin, S.; Keenan, M.C.; et al. Cancer-associated IDH1 mutations produce 2-hydroxyglutarate. Nature 2009, 462, 739–744. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Lv, C.; Zhu, G. Novel type II and monomeric NAD+ specific isocitrate dehydrogenases: Phylogenetic affinity, enzymatic characterization, and evolutionary implication. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.G.; Song, P.; Cao, Z.Y.; Wang, P.; Zhu, G.P. A unique homodimeric NAD+-linked isocitrate dehydrogenase from the smallest autotrophic eukaryote Ostreococcus tauri. FASEB J. 2015, 29, 2462–2472. [Google Scholar] [CrossRef]
- Peng, Y.; Zhong, C.; Huang, W.; Ding, J. Structural studies of Saccharomyces cerevesiae mitochondrial NADP-dependent isocitrate dehydrogenase in different enzymatic states reveal substantial conformational changes during the catalytic reaction. Protein Sci. 2008, 17, 1542–1554. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zhao, J.; Xu, Z.; Peng, B.; Huang, Q.; Arnold, E.; Ding, J. Structures of human cytosolic NADP-dependent isocitrate dehydrogenase reveal a novel self-regulatory mechanism of activity. J. Biol. Chem. 2004, 279, 33946–33957. [Google Scholar] [CrossRef] [PubMed]
- Ceccarelli, C.; Grodsky, N.B.; Ariyaratne, N.; Colman, R.F.; Bahnson, B.J. Crystal structure of porcine mitochondrial NADP+-dependent isocitrate dehydrogenase complexed with Mn2+ and isocitrate. Insights into the enzyme mechanism. J. Biol. Chem. 2002, 277, 43454–43462. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Golding, G.B.; Dean, A.M. The selective cause of an ancient adaptation. Science 2005, 307, 1279–1282. [Google Scholar] [CrossRef] [PubMed]
- Srutkova, D.; Spanova, A.; Spano, M.; Drab, V.; Schwarzer, M.; Kozakova, H.; Rittich, B. Efficiency of PCR-based methods in discriminating Bifidobacterium longum ssp. longum and Bifidobacterium longum ssp. infantis strains of human origin. J. Microbiol. Methods 2011, 87, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Zhu, L.; Liu, X.; Li, T.; Zhang, Y.; Ying, T.; Wang, B.; Wang, J.; Dong, H.; Feng, E.; et al. A proteome reference map and proteomic analysis of Bifidobacterium longum NCC2705. Mol. Cell. Proteom. 2006, 5, 1105–1118. [Google Scholar] [CrossRef] [PubMed]
- Schell, M.A.; Karmirantzou, M.; Snel, B.; Vilanova, D.; Berger, B.; Pessi, G.; Zwahlen, M.C.; Desiere, F.; Bork, P.; Delley, M.; et al. The genome sequence of Bifidobacterium longum reflects its adaptation to the human gastrointestinal tract. Proc. Natl. Acad. Sci. USA 2002, 99, 14422–14427. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.Y.; Chang, F.J. Antioxidative effect of intestinal bacteria Bifidobacterium longum ATCC 15708 and Lactobacillus acidophilus ATCC 4356. Dig. Dis. Sci. 2000, 45, 1617–1622. [Google Scholar] [CrossRef] [PubMed]
- Iwabuchi, N.; Xiao, J.Z.; Yaeshima, T.; Iwatsuki, K. Oral administration of Bifidobacterium longum ameliorates influenza virus infection in mice. Biol. Pharm. Bull. 2011, 34, 1352–1355. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.Z.; Kondo, S.; Yanagisawa, N.; Miyaji, K.; Enomoto, K.; Sakoda, T.; Iwatsuki, K.; Enomoto, T. Clinical efficacy of probiotic Bifidobacterium longum for the treatment of symptoms of Japanese cedar pollen allergy in subjects evaluated in an environmental exposure unit. Allergol. Int. 2007, 56, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Sivan, A.; Corrales, L.; Hubert, N.; Williams, J.B.; Aquino-Michaels, K.; Earley, Z.M.; Benyamin, F.W.; Lei, Y.M.; Jabri, B.; Alegre, M.L.; et al. Commensal Bifidobacterium promotes antitumor immunity and facilitates anti-PD-L1 efficacy. Science 2015, 350, 1084–1089. [Google Scholar] [CrossRef] [PubMed]
- Yazawa, K.; Fujimori, M.; Amano, J.; Kano, Y.; Taniguchi, S. Bifidobacterium longum as a delivery system for cancer gene therapy: Selective localization and growth in hypoxic tumors. Cancer Gene Ther. 2000, 7, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Imada, K.; Tamura, T.; Takenaka, R.; Kobayashi, I.; Namba, K.; Inagaki, K. Structure and quantum chemical analysis of NAD+-dependent isocitrate dehydrogenase: Hydride transfer and co-factor specificity. Proteins 2008, 70, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wang, P.; Zhu, G.; Wang, B.; Zhu, G. Enzymatic characterization of a type II isocitrate dehydrogenase from pathogenic Leptospira interrogans serovar Lai strain 56601. Appl. Biochem. Biotechnol. 2014, 172, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Yang, H. A highly specific monomeric isocitrate dehydrogenase from Corynebacterium glutamicum. Arch. Biochem. Biophys. 2000, 383, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Kanao, T.; Kawamura, M.; Fukui, T.; Atomi, H.; Imanaka, T. Characterization of isocitrate dehydrogenase from the green sulfur bacterium Chlorobium limicola. Eur. J. Biochem. 2002, 269, 1926–1931. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.C.; Tian, C.Q.; Cheng, H.M.; Xu, L.; Wang, P.; Zhu, G.P. A novel type II NAD+-specific isocitrate dehydrogenase from the marine bacterium Congregibacter litoralis KT71. PLoS ONE 2015, 10, e0125229. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Jin, M.; Zhu, G. Biochemical and molecular characterization of NAD+-dependent isocitrate dehydrogenase from the ethanologenic bacterium Zymomonas mobilis. FEMS Microbiol. Lett. 2012, 327, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Dean, A.M.; Golding, G.B. Protein engineering reveals ancient adaptive replacements in isocitrate dehydrogenase. Proc. Natl. Acad. Sci. USA 1997, 94, 3104–3109. [Google Scholar] [CrossRef] [PubMed]
- Jennings, G.T.; Minard, K.I.; McAlister-Henn, L. Expression and mutagenesis of mammalian cytosolic NADP+-specific isocitrate dehydrogenase. Biochemistry 1997, 36, 13743–13747. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, P.; Ge, Y.; Wang, W.; Abbas, A.; Zhu, G. NADP+-specific isocitrate dehydrogenase from oleaginous yeast Yarrowia lipolytica CLIB122: Biochemical characterization and coenzyme sites evaluation. Appl. Biochem. Biotechnol. 2013, 171, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Liu, J.; Shen, G. Cloning, expression, and enzymatic characterization of isocitrate dehydrogenase from Helicobacter pylori. Protein J. 2009, 28, 443–447. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Yasutake, Y.; Tanaka, I.; Takada, Y. Elucidation of stability determinants of cold-adapted monomeric isocitrate dehydrogenase from a psychrophilic bacterium, Colwellia maris, by construction of chimeric enzymes. Microbiology 2005, 151, 1083–1094. [Google Scholar] [CrossRef] [PubMed]
- Hurley, J.H.; Chen, R.; Dean, A.M. Determinants of cofactor specificity in isocitrate dehydrogenase: Structure of an engineered NADP+→NAD+ specificity-reversal mutant. Biochemistry 1996, 35, 5670–5678. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Arnedo, A.; Camacho, M.; Llorca, F.; Bonete, M.J. Complete reversal of coenzyme specificity of isocitrate dehydrogenase from Haloferax volcanii. Protein J. 2005, 24, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Jin, M.; Su, R.; Song, P.; Wang, M.; Zhu, G. Enzymatic characterization of isocitrate dehydrogenase from an emerging zoonotic pathogen Streptococcus suis. Biochimie 2011, 93, 1470–1475. [Google Scholar] [CrossRef] [PubMed]
- Steen, I.H.; Lien, T.; Madsen, M.S.; Birkeland, N.K. Identification of cofactor discrimination sites in NAD-isocitrate dehydrogenase from Pyrococcus furiosus. Arch. Microbiol. 2002, 178, 297–300. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Bramucci, E.; Paiardini, A.; Bossa, F.; Pascarella, S. PyMod: Sequence similarity searches, multiple sequence-structure alignments, and homology modeling within PyMOL. BMC Bioinform. 2012, 13. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.N.; Hunt, H.D.; Horton, R.M.; Pullen, J.K.; Pease, L.R. Site-directed mutagenesis by overlap extension using the polymerase chain-reaction. Gene 1989, 77, 51–59. [Google Scholar] [CrossRef]
- Pace, C.N.; Vajdos, F.; Fee, L.; Grimsley, G.; Gray, T. How to measure and predict the molar absorption coefficient of a protein. Protein Sci. 1995, 4, 2411–2423. [Google Scholar] [CrossRef] [PubMed]
- Cvitkovitch, D.G.; Gutierrez, J.A.; Bleiweis, A.S. Role of the citrate pathway in glutamate biosynthesis by Streptococcus mutans. J. Bacteriol. 1997, 179, 650–655. [Google Scholar] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metal Ions | Relative Activity (%) | Metal Ions | Relative Activity 1 (%) | Metal Ions | Relative Activity 2 (%) |
|---|---|---|---|---|---|
| Mn2+ | 100.00 ± 1.07 | Mn2+ | 100.00 ± 1.07 | Mg2+ | 100.00 ± 0.82 |
| Mg2+ | 69.23 ± 0.82 | Mn2+ + Mg2+ | 69.17 ± 0.41 | Mg2+ + Mn2+ | 99.92 ± 0.41 |
| Ni2+ | 5.67 ± 0.54 | Mn2+ + Ni2+ | 47.42 ± 1.08 | Mg2+ + Ni2+ | 43.37 ± 1.46 |
| Co2+ | 14.30 ± 0.39 | Mn2+ + Co2+ | 30.80 ± 0.40 | Mg2+ + Co2+ | 39.58 ± 0.10 |
| Zn2+ | 0 | Mn2+ + Zn2+ | 5.19 ± 0.38 | Mg2+ + Zn2+ | 11.02 ± 0.13 |
| Cu2+ | 0 | Mn2+ + Cu2+ | 31.04 ± 0.37 | Mg2+ + Cu2+ | 61.38 ± 0.67 |
| Ca2+ | 0 | Mn2+ + Ca2+ | 62.33 ± 0.50 | Mg2+ + Ca2+ | 14.06 ± 0.50 |
| Li+ | 0 | Mn2+ + Li+ | 103.83 ± 0.65 | Mg2+ + Li+ | 100.52 ± 0.65 |
| Rb+ | 5.26 ± 0.39 | Mn2+ + Rb+ | 96.09 ± 0.57 | Mg2+ + Rb+ | 105.77 ± 0.35 |
| K+ | 4.89 ± 0.11 | Mn2+ + K+ | 101.98 ± 0.30 | Mg2+ + K+ | 103.75 ± 0.90 |
| Na+ | 6.05 ± 0.43 | Mn2+ + Na+ | 104.53 ± 0.15 | Mg2+ + Na+ | 109.27 ± 0.18 |
| Residues at 1 | NADP+ | NAD+ | Specificity (A/B) | Specificity (B/A) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 253 | 257 | 260 | 314 | 315 | 327 | Km (μM) | kcat (s−1) | kcat/Km (s−1·mM−1) (A) 2 | Km (μM) | kcat (s−1) | kcat/Km (s−1·mM−1) (B) 2 | ||
| D | S | K | R | H | T | 19.45 ± 2.6 | 36.38 ± 2.1 | 1870 ± 180 | 3584 ± 238.0 | 11.7 ± 5.2 | 3.3 ± 0.9 | 567 | 0.0017 |
| (wild-type) | |||||||||||||
| * | * | * | D | * | * | 1102 ± 92.6 | 2.15 ± 1.3 | 1.9 ± 0.7 | 3702 ± 117.1 | 0.88 ± 0.7 | 0.2 ± 0.1 | 9.5 | 0.105 |
| * | * | * | D | I | * | - | - | - | - | - | - | - | - |
| * | * | * | D | M | * | - | - | - | - | - | - | - | - |
| * | * | * | D | M | L | - | - | - | - | - | - | - | - |
| * | * | * | D | I | A | - | - | - | 4407 ± 196.7 | 0.41 ± 0.1 | 0.1 ± 0.01 | - | - |
| A | K | Q | D | I | A | 324.1 ± 27.9 | 0.307 ± 0.1 | 0.95 ± 0.05 | 130 ± 20.9 | 0.518 ± 0.03 | 4.0 ± 0.5 | 0.2375 | 4.21 |
| Enzymes & Subfamilies | NAD+ | NADP+ | Specificity (A/B) | References | ||||
|---|---|---|---|---|---|---|---|---|
| Km (μM) | kcat (s−1) | kcat/Km (s−1·μM−1) (A) c | Km (μM) | kcat (s−1) | kcat/Km (s−1·μM−1) (B) c | |||
| Type I NAD-IDH | ||||||||
| S. suis NAD-IDH (Mg2+) a | 233 | 41 | 0.176 | 9527 | 14 | 0.0015 | 117 | [39] |
| Z. mobilis NAD-IDH (Mg2+) | 312 | 88 | 0.282 | 8200 | 14 | 0.0017 | 165 | [31] |
| Engineered E. coli NAD-IDH (Mg2+) b | 99 | 16.2 | 0.164 | 5800 | 4.7 | 0.00081 | 202 | [37] |
| Type II IDH | ||||||||
| C. litoralis NAD-IDH (Mg2+) | 262.6 | 36.7 | 0.140 | - | - | - | - | [30] |
| Micromonas sp. NAD-IDH (Mg2+) | 126 | 22.5 | 0.179 | 1827 | 1.4 | 0.0008 | 224 | [12] |
| O. lucimarinus NAD-IDH (Mg2+) | 136.6 | 60.6 | 0.444 | 2211 | 10.0 | 0.0045 | 99 | [12] |
| O. tauri NAD-IDH (Mg2+) | 226 | 59 | 0.261 | 3354 | 17 | 0.0051 | 51 | [13] |
| B. longum NADP-IDH (Mg2+) | 3584 | 11.70 | 0.003 | 19.45 | 36.4 | 1.87 | 0.0016 | This study |
| Engineered B. longum NAD-IDH (Mg2+) b | 130 | 0.518 | 0.004 | 324.1 | 0.307 | 0.00095 | 4.21 | This study |
| Y. lipolytica NADP-IDH (Mg2+) | - | - | - | 59 | 72 | 1.22 | - | [34] |
| Engineered Y. lipolytica NAD-IDH (Mg2+) b | 47,000 | 0.38 | 8.1 × 10−6 | 2410 | 4.24 | 1750 | 4.6×10-9 | [34] |
| Monomeric NAD-IDH | ||||||||
| Campylobacter sp. NAD-IDH (Mg2+) | 28.9 | 7.0 | 0.242 | 513.2 | 1.9 | 0.004 | 61 | [12] |
| C. curvus NAD-IDH (Mg2+) a | 74.2 | 10.8 | 0.146 | 475.9 | 2.0 | 0.004 | 37 | [12] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, S.-P.; Cheng, H.-M.; Wang, P.; Zhu, G.-P. Biochemical Characterization and Complete Conversion of Coenzyme Specificity of Isocitrate Dehydrogenase from Bifidobacterium longum. Int. J. Mol. Sci. 2016, 17, 296. https://doi.org/10.3390/ijms17030296
Huang S-P, Cheng H-M, Wang P, Zhu G-P. Biochemical Characterization and Complete Conversion of Coenzyme Specificity of Isocitrate Dehydrogenase from Bifidobacterium longum. International Journal of Molecular Sciences. 2016; 17(3):296. https://doi.org/10.3390/ijms17030296
Chicago/Turabian StyleHuang, Shi-Ping, Hong-Mei Cheng, Peng Wang, and Guo-Ping Zhu. 2016. "Biochemical Characterization and Complete Conversion of Coenzyme Specificity of Isocitrate Dehydrogenase from Bifidobacterium longum" International Journal of Molecular Sciences 17, no. 3: 296. https://doi.org/10.3390/ijms17030296
APA StyleHuang, S.-P., Cheng, H.-M., Wang, P., & Zhu, G.-P. (2016). Biochemical Characterization and Complete Conversion of Coenzyme Specificity of Isocitrate Dehydrogenase from Bifidobacterium longum. International Journal of Molecular Sciences, 17(3), 296. https://doi.org/10.3390/ijms17030296