
Investigation of the Antiproliferative Properties of Natural Sesquiterpenes from Artemisia asiatica and Onopordum acanthium on HL-60 Cells in Vitro


 ,
, 
Abstract
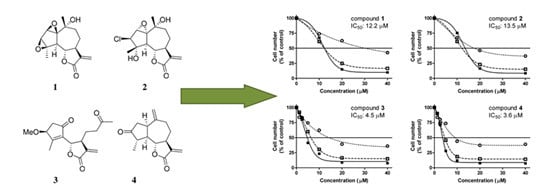
:
1. Introduction
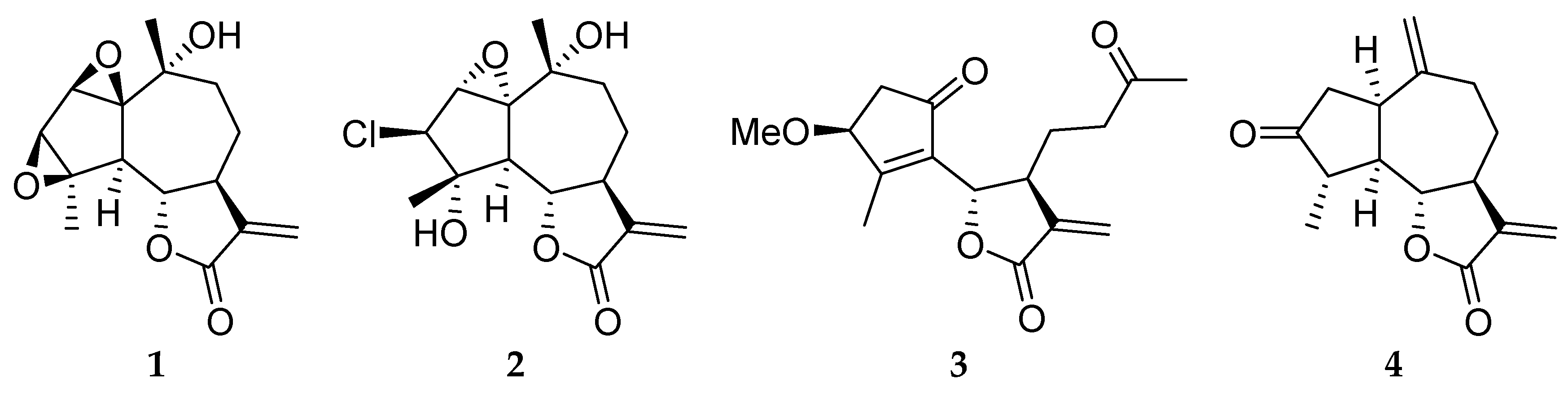
2. Results and Discussion
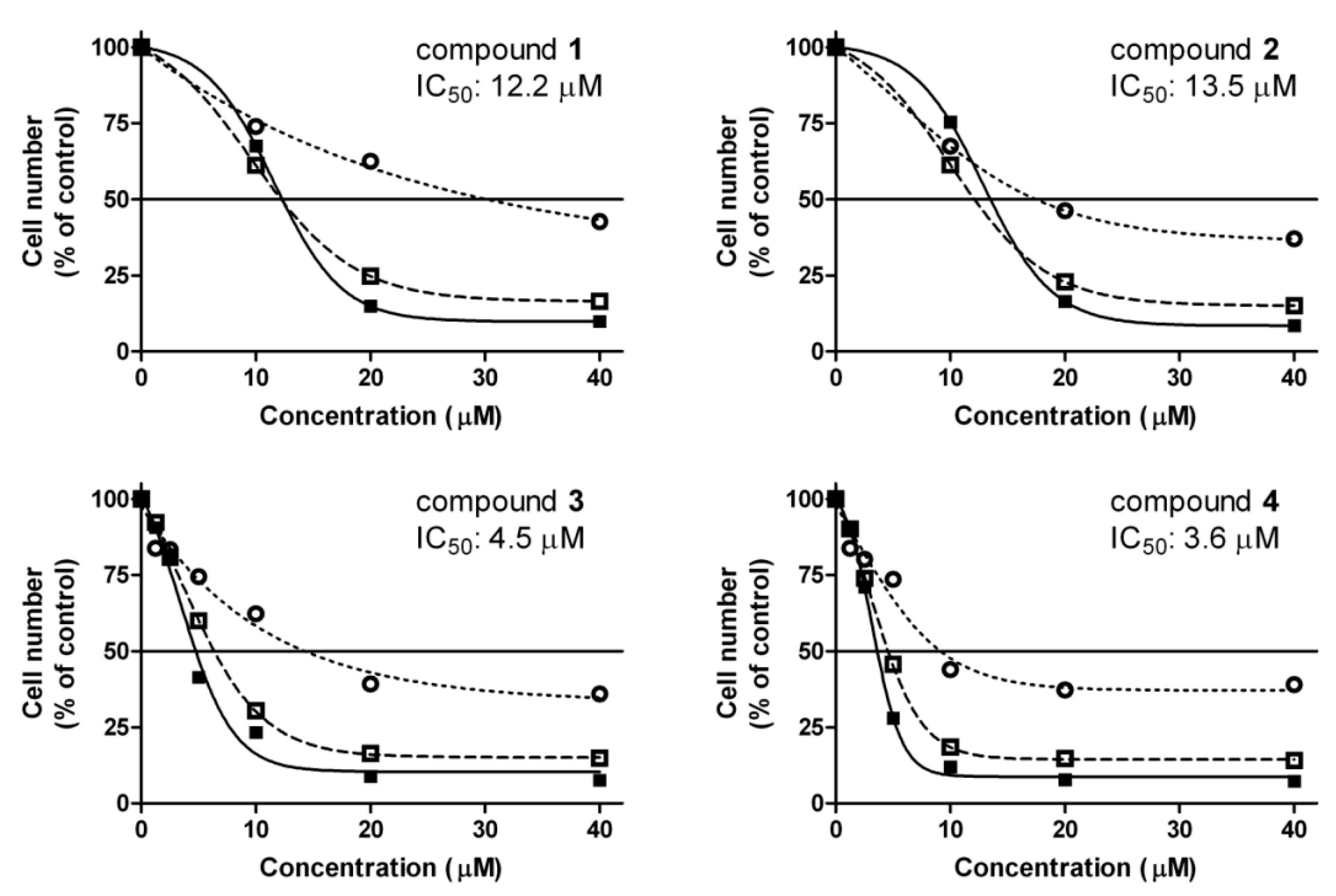
2.1. Antiproliferative Assay
 ), 48 (
), 48 (  ) and 72 h (■) and their IC50 values calculated from 72-h data. Data are means ± SEM from three experiments.
), 48 ( ) and 72 h (■) and their IC50 values calculated from 72-h data. Data are means ± SEM from three experiments.
) and 72 h (■) and their IC50 values calculated from 72-h data. Data are means ± SEM from three experiments.
), 48 ( ) and 72 h (■) and their IC50 values calculated from 72-h data. Data are means ± SEM from three experiments.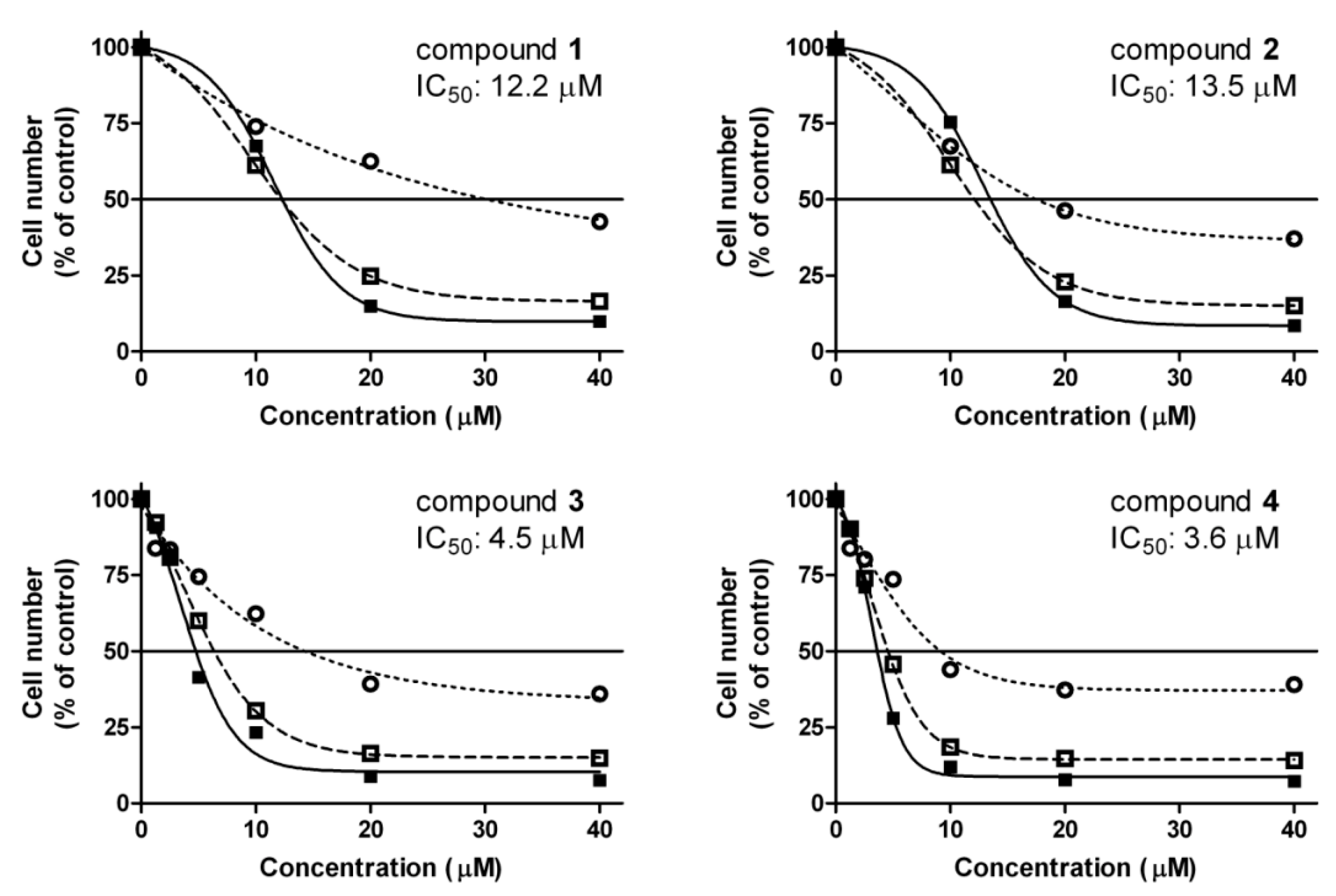
2.2. Cell Viability Measurements
and ■ indicate fibroblast and HL-60 cells, respectively. Data are means ± SEM from three experiments.
and ■ indicate fibroblast and HL-60 cells, respectively. Data are means ± SEM from three experiments.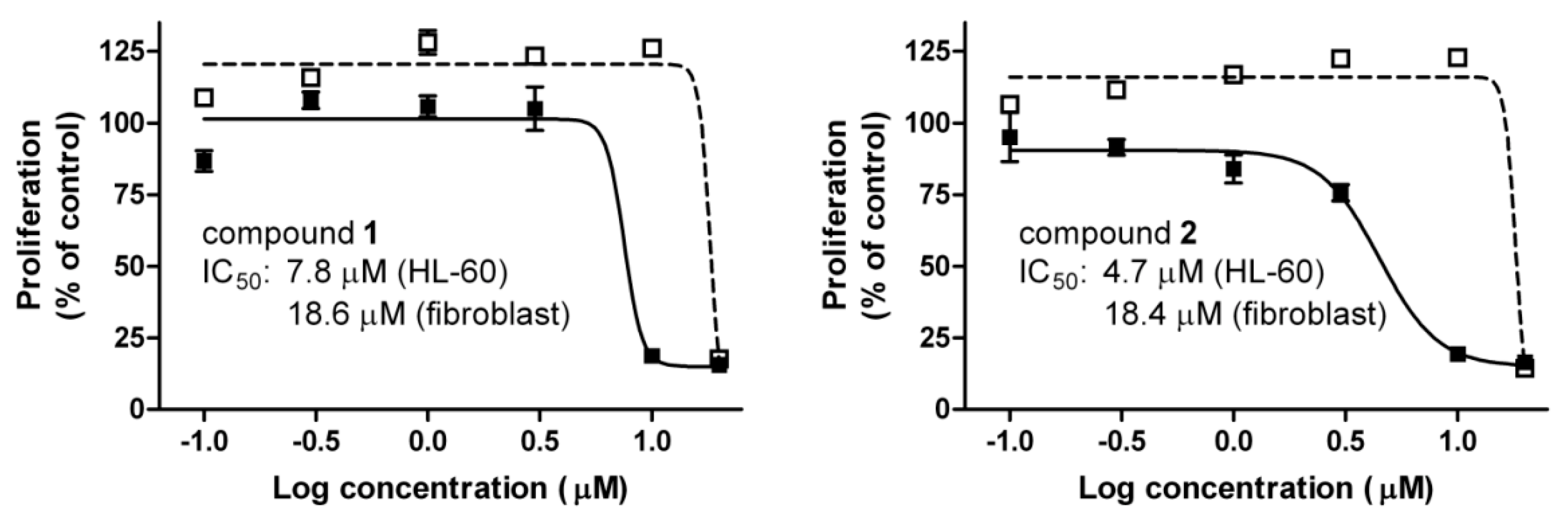
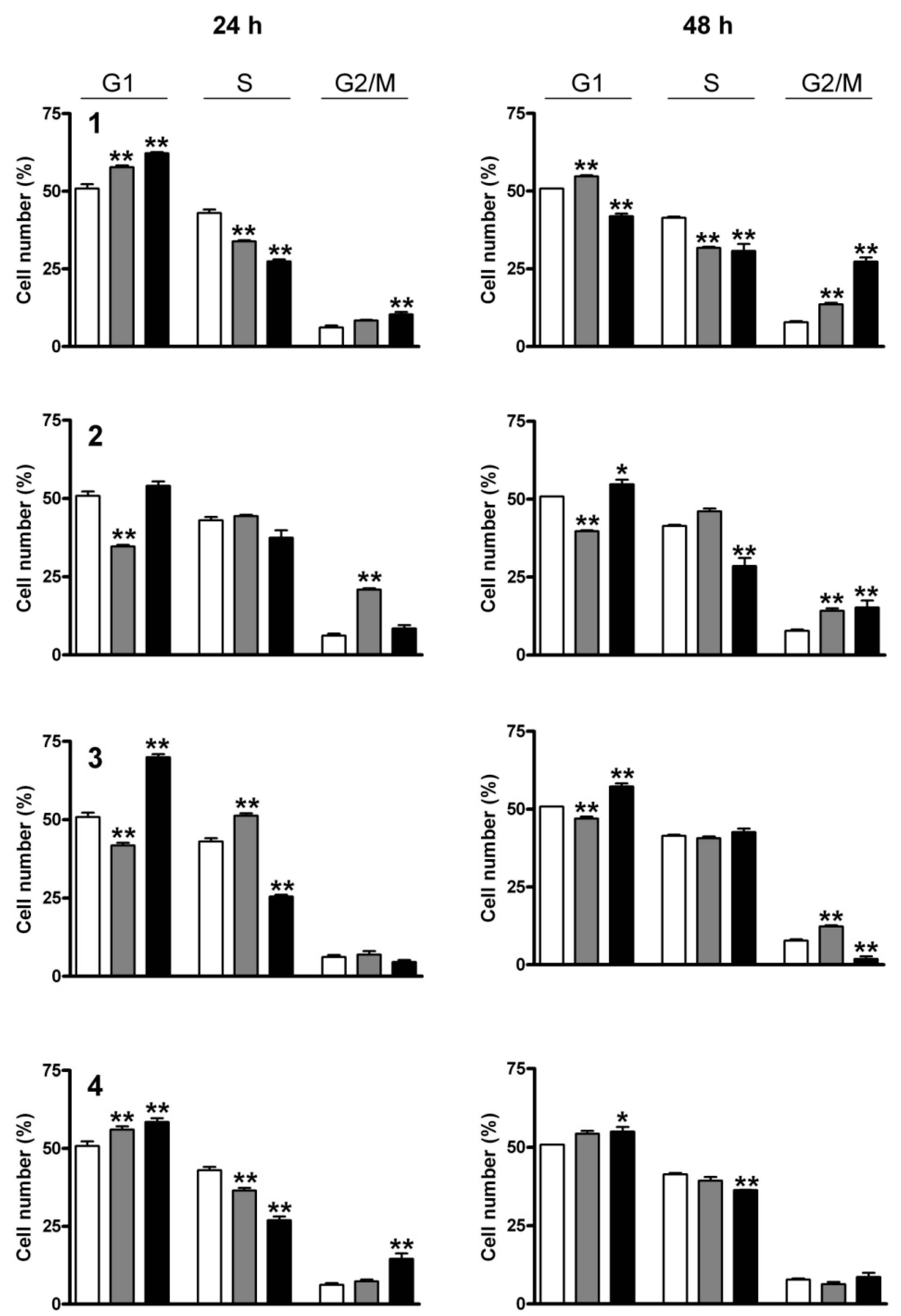
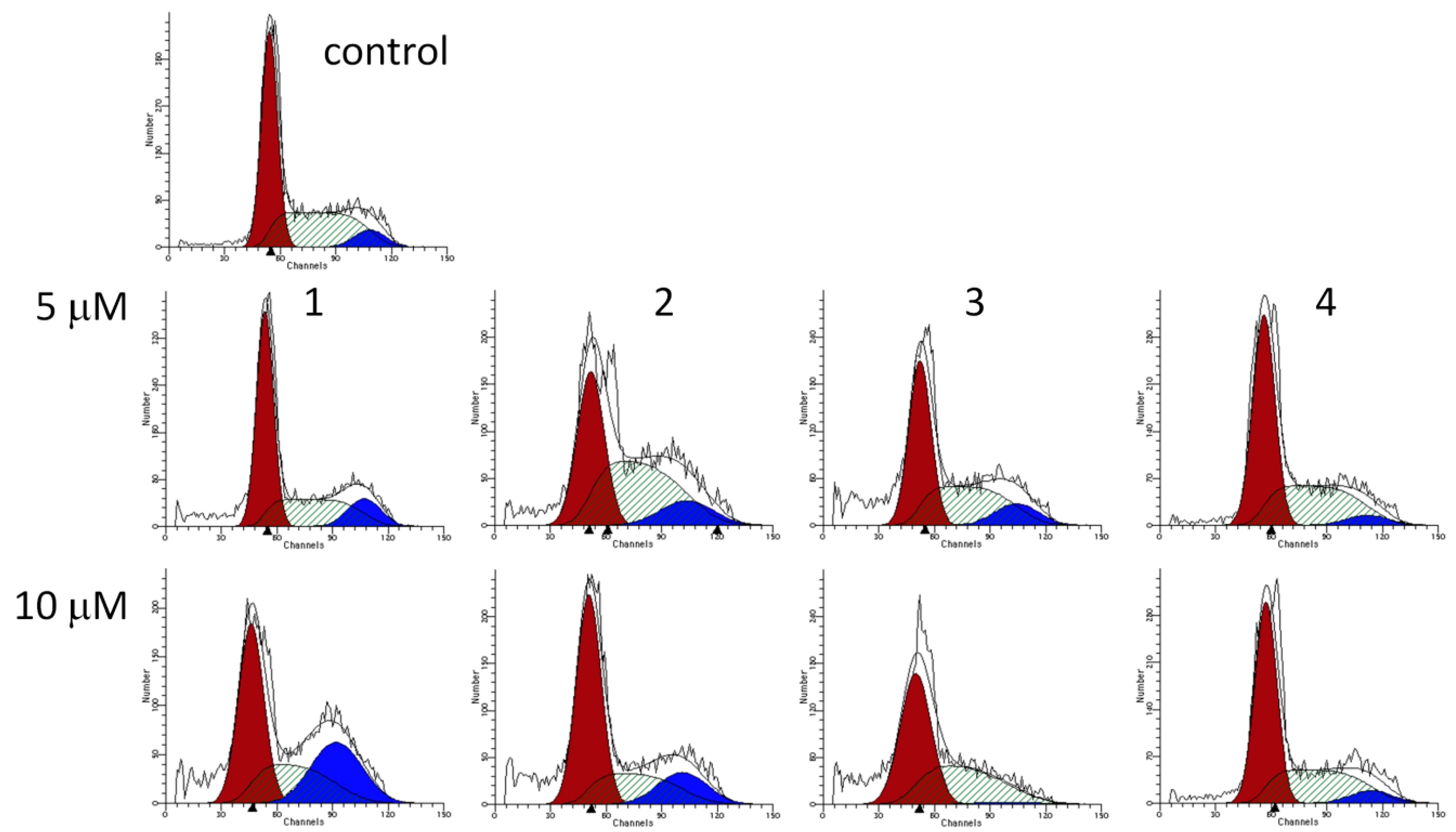
2.3. Cell Cycle Analysis
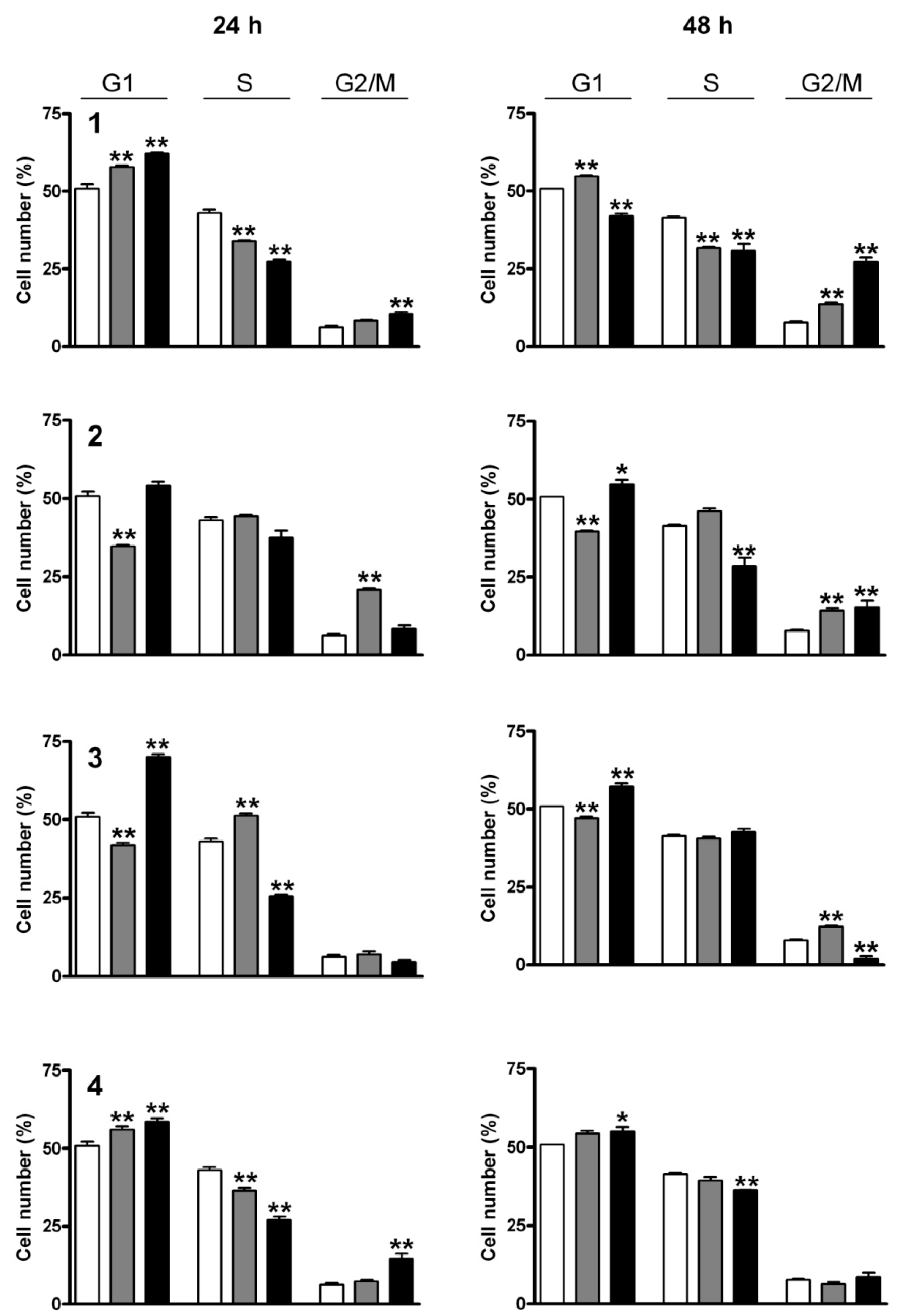
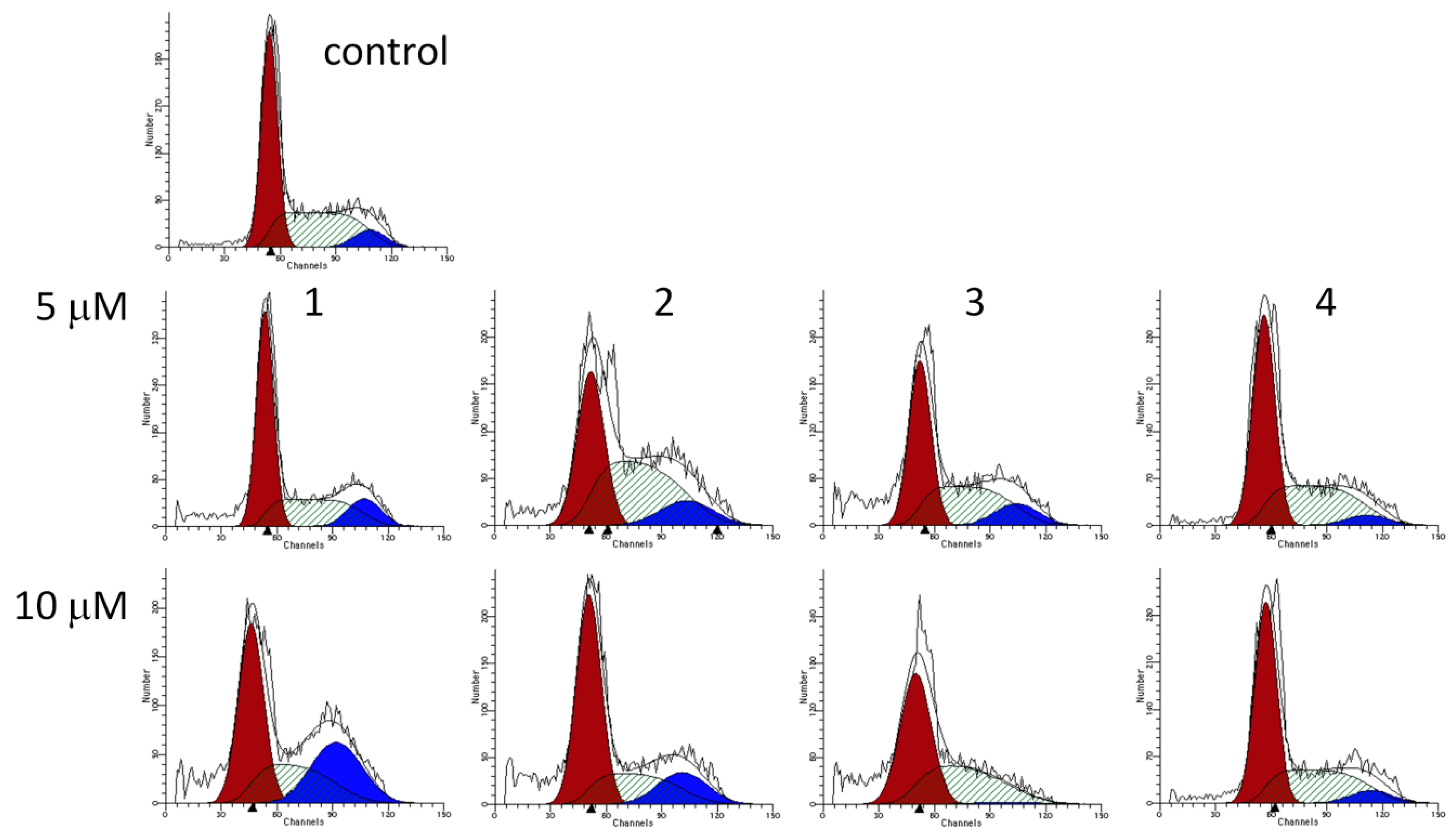
2.4. Fluorescent Staining

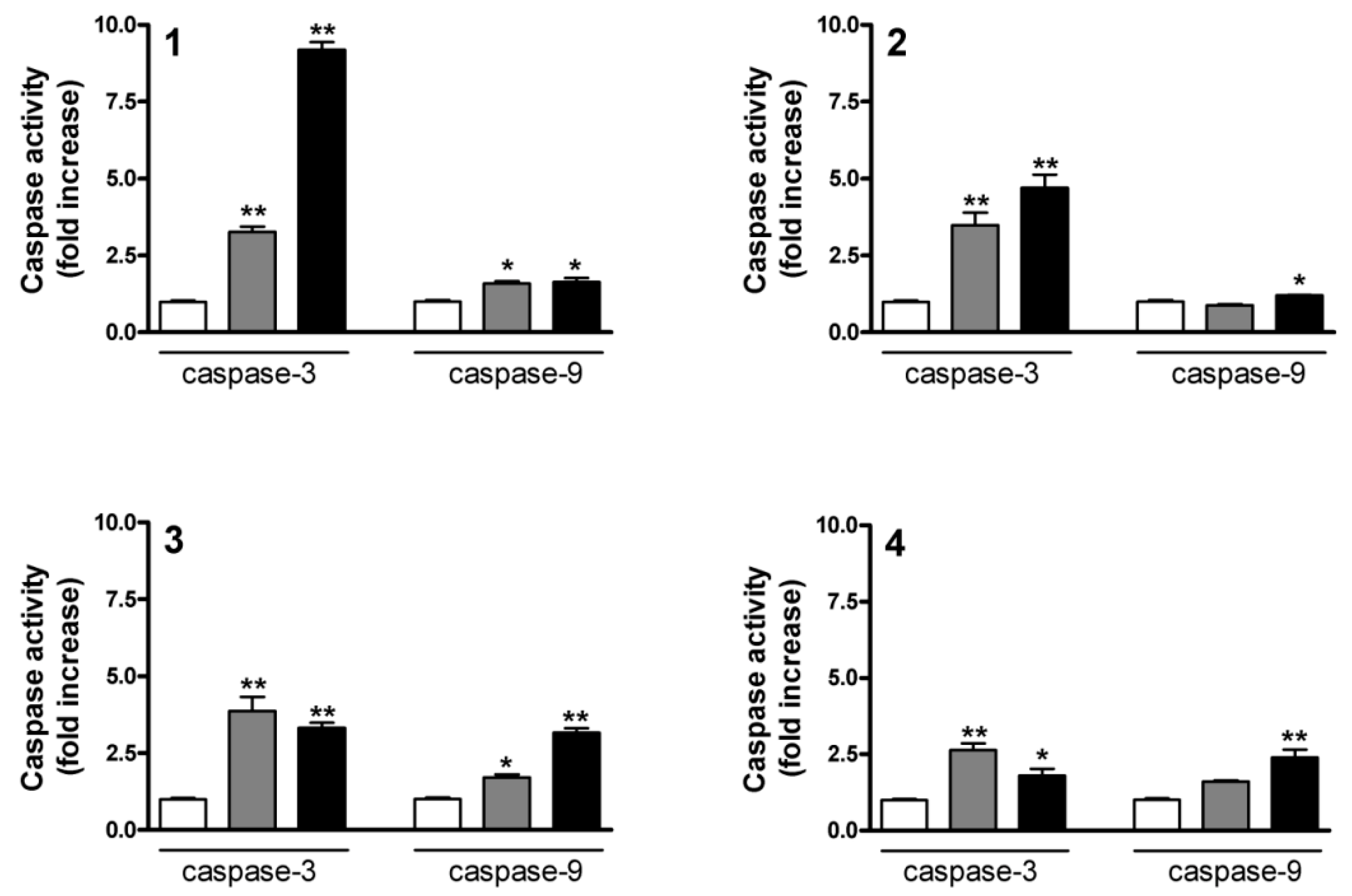
2.5. Determination of Caspase-3 and Caspase-9 Activities
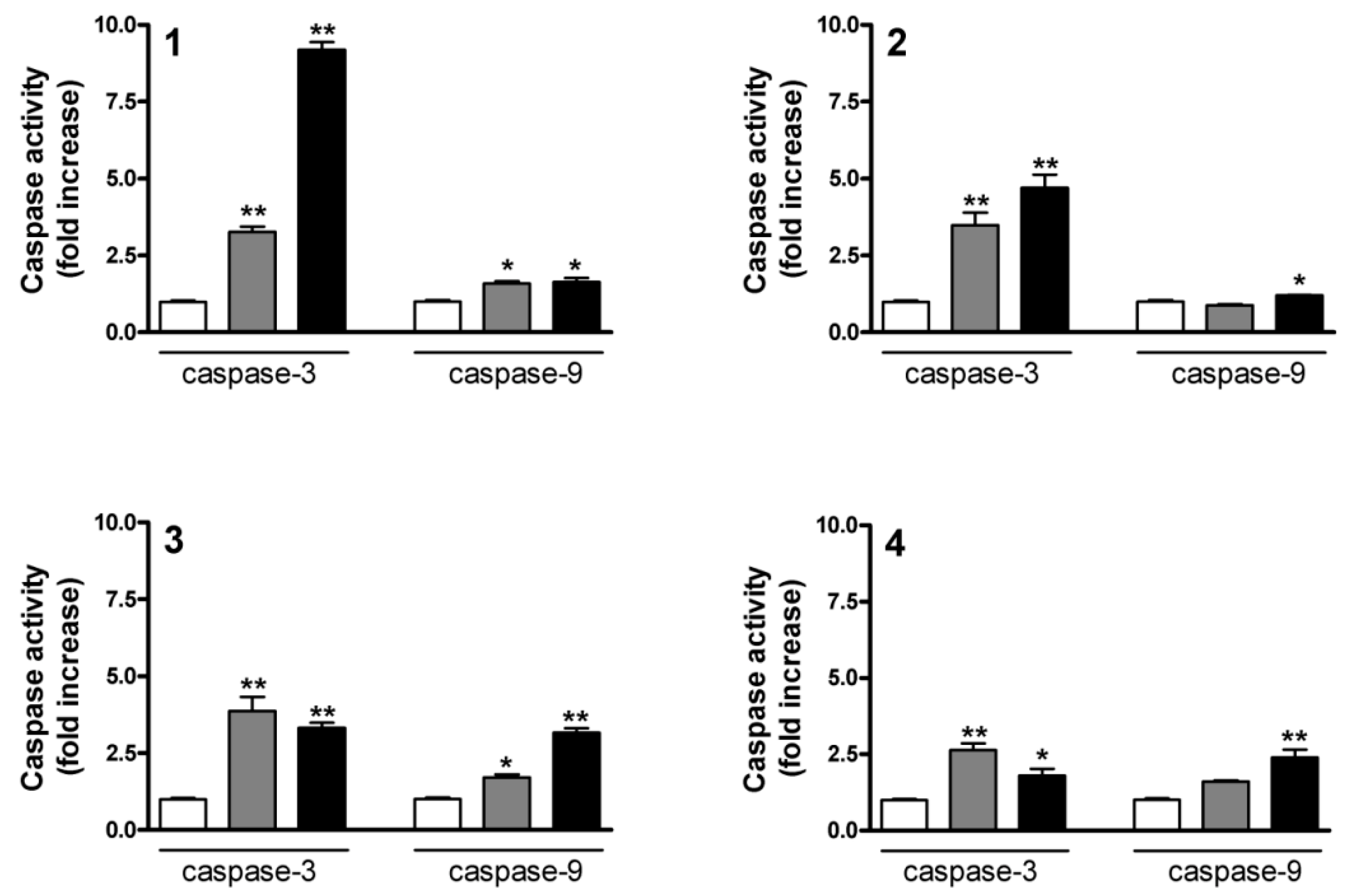
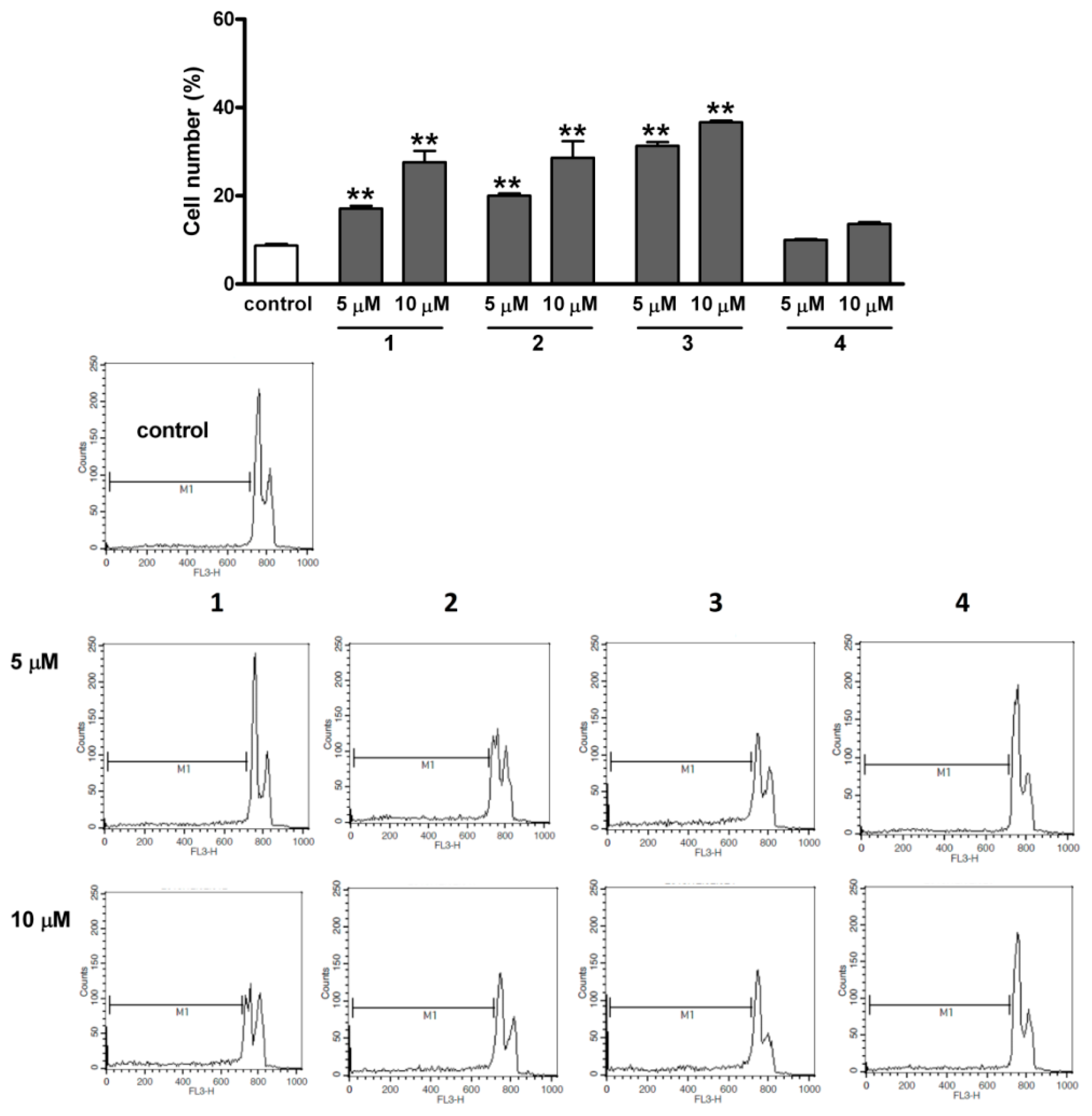
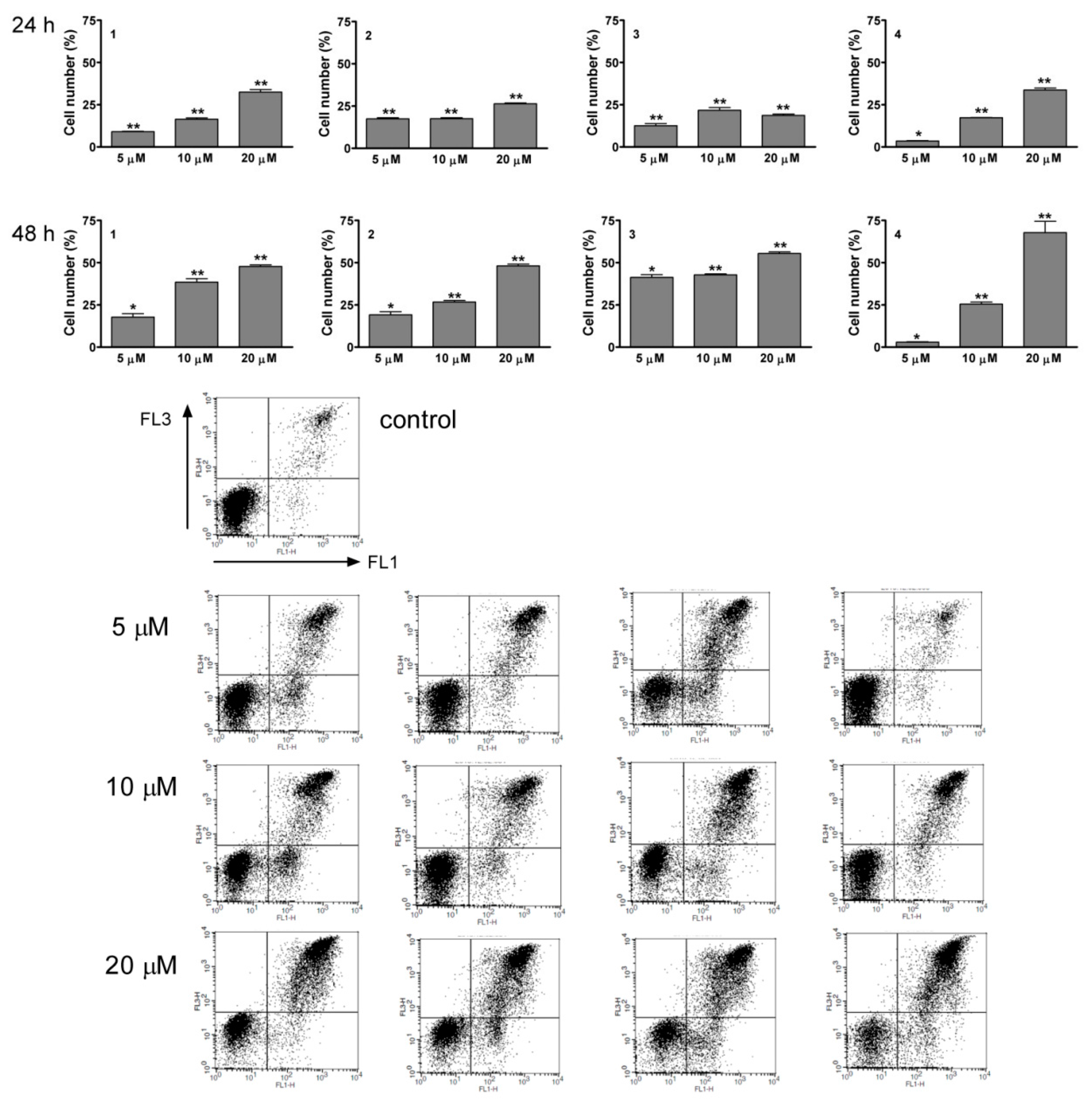
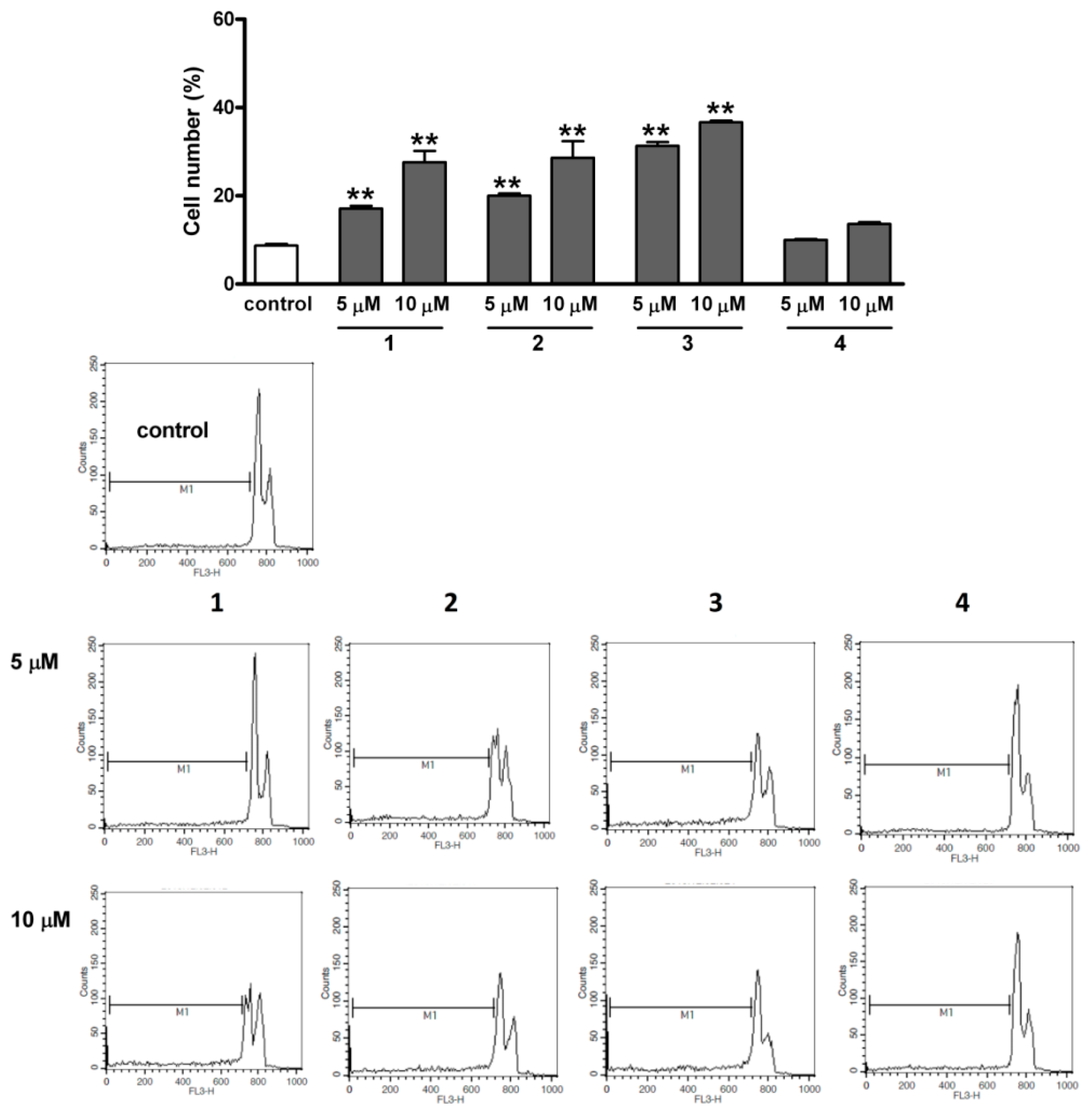
2.6. Flow Cytometric Apoptosis Assay
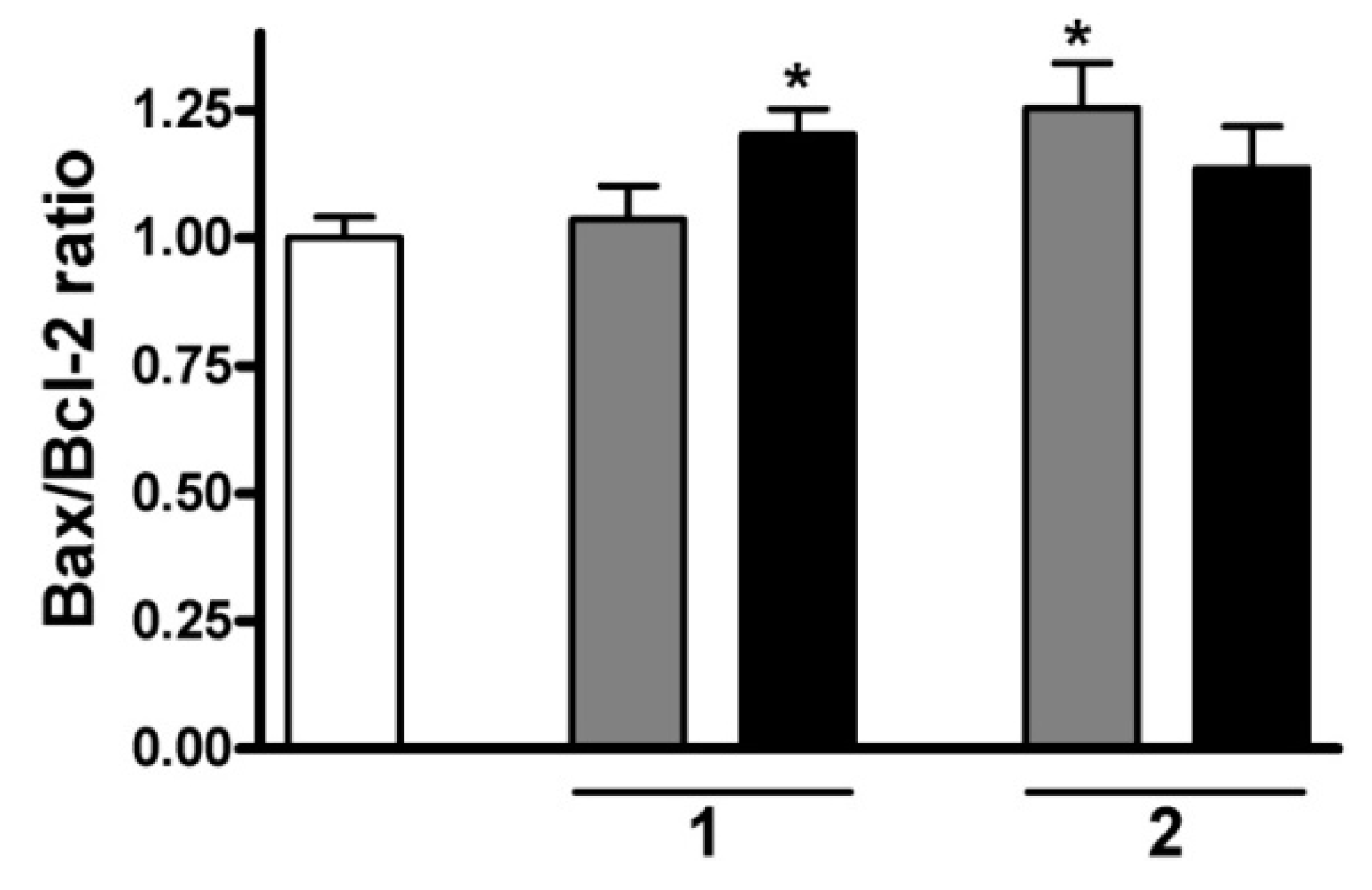
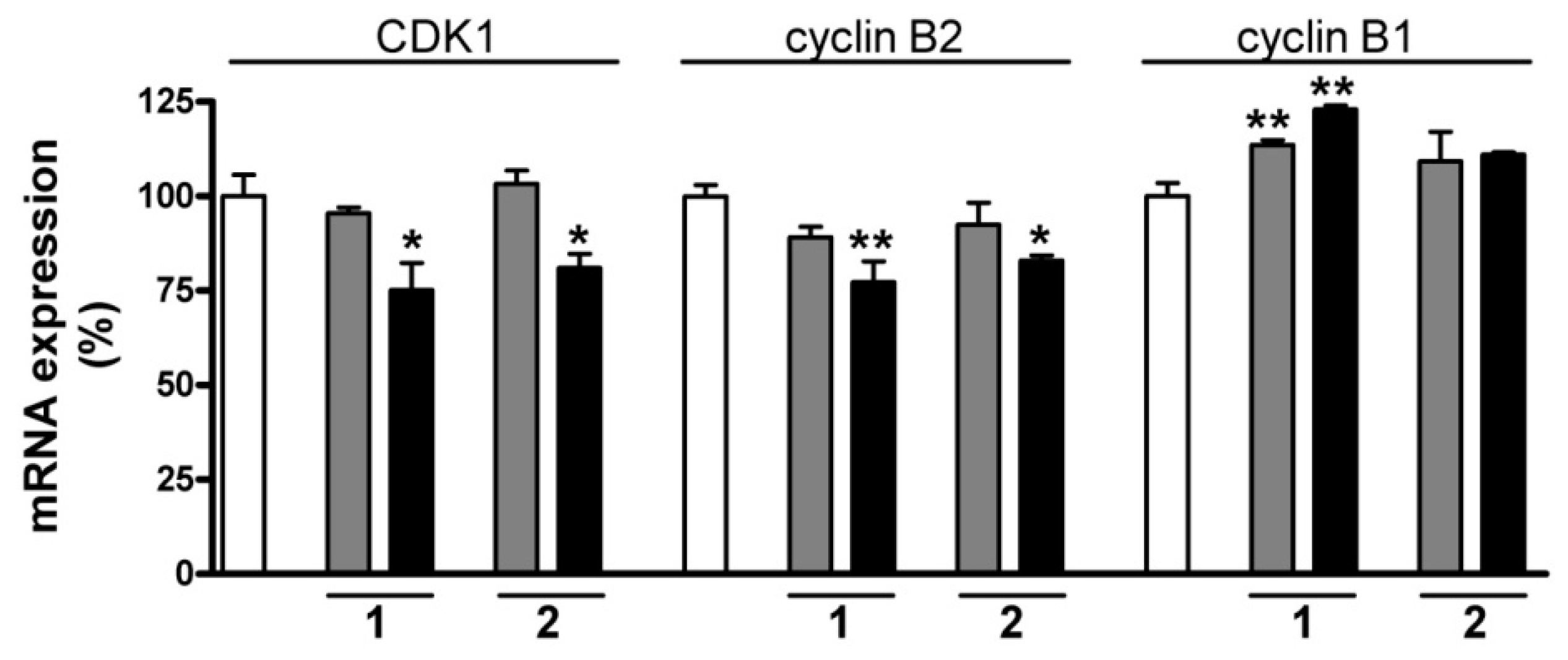
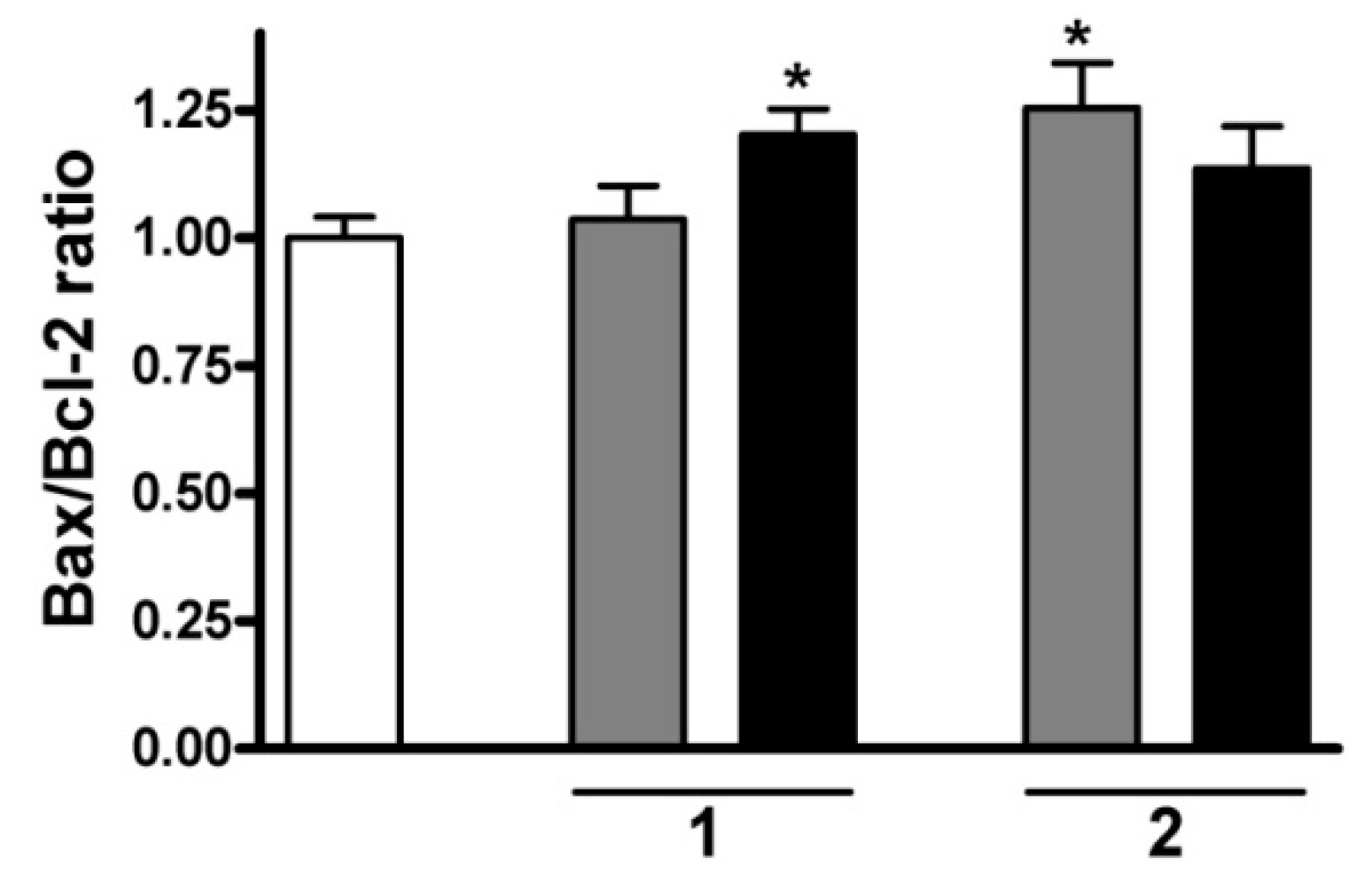
2.7. RT-PCR Studies
3. Experimental Section
3.1. Chemicals
3.2. Cell Culturing and Antiproliferative Assay
3.3. Cell Viability Measurements
3.4. Cell Cycle Analysis by Flow Cytometry
3.5. Double Staining with Hoechst 33258 and PI
3.6. Caspase-3 Assay
3.7. Caspase-9 Assay
3.8. Reverse Transcription-Polymerase Chain Reaction (RT-PCR) Studies

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Primer Sequence | Gene ID | Product Size (bp) | Coupling Temp. (°C) |
|---|---|---|---|---|
| CDK1 | F: ACTGGCTGATTTTGGCCTTGCC | 983 | 118 | 62 |
| R: TGAGTAACGAGCTGACCCCAGCAA | ||||
| cyclin B1 | F: AATAAGGAGGGAGCAGTGCG | 891 | 51 | 60 |
| R: GAAGAGCCAGCCTAGCCTCAG | ||||
| cyclin B2 | F: GCGTTGGCATTATGGATCG | 9133 | 51 | 60 |
| R: TCTTCCGGGAAACTGGCTG | ||||
| Bax | F: TGGCAGCTGACATGTTTTCTGAC | 581 | 195 | 53 |
| R: CGTCCCAACCACCCTGGTCT | ||||
| Bcl-2 | F: GACTTCGCCGAGATGTCCAG | 596 | 225 | 51 |
| R: CAGGTGCCGGTTCAGGTACT | ||||
| hGAPDH | F: ACCCAGAAGACTGTGGATGG | 2597 | 415 | 55 |
| R: TGCTGTAGCCAAATTCGTTG |
3.9. Flow Cytometric Apoptosis Assay
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Are, C.; Rajaram, S.; Are, M.; Raj, H.; Anderson, B.O.; Chaluvarya Swamy, R.; Vijayakumar, M.; Song, T.; Pandey, M.; Edney, J.A.; et al. A review of global cancer burden: Trends, challenges, strategies, and a role for surgeons. J. Surg. Oncol. 2013, 107, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Cragg, G.M.; Grothaus, P.G.; Newman, D.J. Impact of natural products on developing new anti-cancer agents. Chem. Rev. 2009, 109, 3012–3043. [Google Scholar] [CrossRef] [PubMed]
- Cragg, G.; Newman, D. Nature: A vital source of leads for anticancer drug development. Phytochem. Rev. 2009, 8, 313–331. [Google Scholar] [CrossRef]
- Heywood, V.H.; Harborne, J.B.; Turner, B.L. The Biology and Chemistry of the Compositae; Academic Press: London, UK, 1977. [Google Scholar]
- Hegnauer, R. Chemotaxonomie der Pflanzen; Birkhauser Verlag: Basel, Switzerland, 1989. [Google Scholar]
- Kuete, V.; Sandjo, L.P.; Wiench, B.; Efferth, T. Cytotoxicity and modes of action of four Cameroonian dietary spices ethno-medically used to treat cancers: Echinops giganteus, Xylopia aethiopica, Imperata cylindrica and Piper capense. J. Ethnopharmacol. 2013, 149, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Csupor-Löffler, B.; Hajdú, Z.; Réthy, B.; Zupkó, I.; Máthé, I.; Rédei, T.; Falkay, G.; Hohmann, J. Antiproliferative activity of Hungarian Asteraceae species against human cancer cell lines. Part II. Phytother. Res. 2009, 23, 1109–1115. [Google Scholar] [CrossRef] [PubMed]
- Réthy, B.; Csupor-Löffler, B.; Zupkó, I.; Hajdú, Z.; Máthé, I.; Hohmann, J.; Rédei, T.; Falkay, G. Antiproliferative activity of Hungarian Asteraceae species against human cancer cell lines. Part I. Phytother. Res. 2007, 21, 1200–1208. [Google Scholar] [CrossRef] [PubMed]
- Csupor-Löffler, B.; Hajdú, Z.; Zupkó, I.; Molnár, J.; Forgo, P.; Vasas, A.; Kele, Z.; Hohmann, J. Antiproliferative constituents of the roots of Conyza canadensis. Planta Med. 2011, 77, 1183–1188. [Google Scholar] [CrossRef] [PubMed]
- Hajdú, Z.; Zupkó, I.; Réthy, B.; Forgo, P.; Hohmann, J. Bioactivity-guided isolation of cytotoxic sesquiterpenes and flavonoids from Anthemis ruthenica. Planta Med. 2010, 76, 94–96. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Kundu, J.K.; Lee, J.S.; Oh, T.Y.; Na, H.K.; Surh, Y.J. Chemopreventive effects of the standardized extract (DA-9601) of Artemisia asiatica on azoxymethane-initiated and dextran sulfate sodium-promoted mouse colon carcinogenesis. Nutr. Cancer 2008, 60 (Suppl. 1), 90–97. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Oh, T.Y.; Ahn, B.O.; Cho, H.; Kim, W.B.; Kim, Y.B.; Surh, Y.J.; Kim, H.J.; Hahm, K.B. Involvement of oxidative stress in experimentally induced reflux esophagitis and Barrett’s esophagus: Clue for the chemoprevention of esophageal carcinoma by antioxidants. Mutat. Res. 2001, 480–481, 189–200. [Google Scholar] [CrossRef]
- Hajdú, Z.; Hohmann, J.; Forgo, P.; Máthé, I.; Molnár, J.; Zupkó, I. Antiproliferative activity of Artemisia asiatica extract and its constituents on human tumor cell lines. Planta Med. 2014, 80, 1692–1697. [Google Scholar] [CrossRef] [PubMed]
- Csupor-Löffler, B.; Zupkó, I.; Molnár, J.; Forgo, P.; Hohmann, J. Bioactivity-guided isolation of antiproliferative compounds from the roots of Onopordum acanthium. Nat. Prod. Commun. 2014, 9, 337–340. [Google Scholar] [PubMed]
- Muhammad, I.; Takamatsu, S.; Mossa, J.S.; El-Feraly, F.S.; Walker, L.A.; Clark, A.M. Cytotoxic sesquiterpene lactones from Centaurothamnus maximus and Vicoa pentanema. Phytother. Res. 2003, 17, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Tolomeo, M.; Simoni, D. Drug resistance and apoptosis in cancer treatment: Development of new apoptosis-inducing agents active in drug resistant malignancies. Curr. Med. Chem. Anticancer Agents 2002, 2, 387–401. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S. Caspase function in programmed cell death. Cell Death Differ. 2007, 14, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Kuypers, F.A.; Lewis, R.A.; Hua, M.; Schott, M.A.; Discher, D.; Ernst, J.D.; Lubin, B.H. Detection of altered membrane phospholipid asymmetry in subpopulations of human red blood cells using fluorescently labeled annexin V. Blood 1996, 87, 1179–1187. [Google Scholar] [PubMed]
- Brinkmann, K.; Kashkar, H. Targeting the mitochondrial apoptotic pathway: A preferred approach in hematologic malignancies? Cell Death Dis. 2014, 5, e1098. [Google Scholar] [CrossRef] [PubMed]
- Lindqvist, A.; Kallstrom, H.; Lundgren, A.; Barsoum, E.; Rosenthal, C.K. Cdc25B cooperates with Cdc25A to induce mitosis but has a unique role in activating cyclin B1-Cdk1 at the centrosome. J. Cell Biol. 2005, 171, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Millimouno, F.M.; Dong, J.; Yang, L.; Li, J.; Li, X. Targeting apoptosis pathways in cancer and perspectives with natural compounds from Mother Nature. Cancer Prev. Res. 2014, 7, 1081–1107. [Google Scholar] [CrossRef] [PubMed]
- Kreuger, M.R.; Grootjans, S.; Biavatti, M.W.; Vandenabeele, P.; D’Herde, K. Sesquiterpene lactones as drugs with multiple targets in cancer treatment: Focus on parthenolide. Anticancer Drugs 2012, 23, 883–896. [Google Scholar] [PubMed]
- Fernandes, M.B.; Scotti, M.T.; Ferreira, M.J.; Emerenciano, V.P. Use of self-organizing maps and molecular descriptors to predict the cytotoxic activity of sesquiterpene lactones. Eur. J. Med. Chem. 2008, 43, 2197–2205. [Google Scholar] [CrossRef] [PubMed]
- Gach, K.; Dlugosz, A.; Janecka, A. The role of oxidative stress in anticancer activity of sesquiterpene lactones. Naunyn Schmiedebergs Arch. Pharmacol. 2015, 388, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Won, Y.K.; Ong, C.N.; Shen, H.M. Anti-cancer potential of sesquiterpene lactones: Bioactivity and molecular mechanisms. Curr. Med. Chem. Anticancer Agents 2005, 5, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Gach, K.; Janecka, A. α-Methylene-γ-lactones as a novel class of anti-leukemic agents. Anticancer Agents Med. Chem. 2014, 14, 688–694. [Google Scholar] [CrossRef] [PubMed]
- Chadwick, M.; Trewin, H.; Gawthrop, F.; Wagstaff, C. Sesquiterpenoids lactones: Benefits to plants and people. Int. J. Mol. Sci. 2013, 14, 12780–12805. [Google Scholar] [CrossRef] [PubMed]
- Ghantous, A.; Sinjab, A.; Herceg, Z.; Darwiche, N. Parthenolide: From plant shoots to cancer roots. Drug Discov. Today 2013, 18, 894–905. [Google Scholar] [CrossRef] [PubMed]
- Doan, H.Q.; Gulati, N.; Levis, W.R. Ingenol mebutate: Potential for further development of cancer immunotherapy. J. Drugs Dermatol. 2012, 11, 1156–1157. [Google Scholar] [PubMed]
- Shaikenov, T.E.; Adekenov, S.M.; Williams, R.M.; Prashad, N.; Baker, F.L.; Madden, T.L.; Newman, R. Arglabin-DMA, a plant derived sesquiterpene, inhibits farnesyltransferase. Oncol. Rep. 2001, 8, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Puskás, L.G.; Fehér, L.Z.; Vizler, C.; Ayaydin, F.; Rásó, E.; Molnár, E.; Magyary, I.; Kanizsai, I.; Gyuris, M.; Madácsi, R.; et al. Polyunsaturated fatty acids synergize with lipid droplet binding thalidomide analogs to induce oxidative stress in cancer cells. Lipids Health Dis. 2010, 9, 56. [Google Scholar] [CrossRef] [PubMed]
- Vermes, I.; Haanen, C.; Reutelingsperger, C. Flow cytometry of apoptotic cell death. J. Immunol. Methods 2000, 243, 167–190. [Google Scholar] [CrossRef]
- Minorics, R.; Szekeres, T.; Krupitza, G.; Saiko, P.; Giessrigl, B.; Wölfling, J.; Frank, E.; Zupkó, I. Antiproliferative effects of some novel synthetic solanidine analogs on HL-60 human leukemia cells in vitro. Steroids 2011, 76, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Ribble, D.; Goldstein, N.B.; Norris, D.A.; Shellman, Y.G. A simple technique for quantifying apoptosis in 96-well plates. BMC Biotechnol. 2005, 5, 12. [Google Scholar] [CrossRef] [PubMed]
- Molnár, J.; Ocsovszki, I.; Puskás, L.; Ghane, T.; Hohmann, J.; Zupkó, I. Investigation of the antiproliferative action of the quinoline alkaloids kokusaginine and skimmianine on human cell lines. Curr. Signal Transduct. Ther. 2013, 8, 148–155. [Google Scholar] [CrossRef]
- Molnár, J.; Frank, É.; Minorics, R.; Kádár, Z.; Ocsovszki, I.; Schönecker, B.; Wölfling, J.; Zupkó, I. A click approach to novel D-ring-substituted 16a-triazolylestrone derivatives and characterization of their antiproliferative properties. PLoS ONE 2015, 10, e0118104. [Google Scholar] [CrossRef] [PubMed]
- Nagy, L.I.; Fehér, L.Z.; Szebeni, G.J.; Gyuris, M.; Sipos, P.; Alföldi, R.; Ózsvári, B.; Hackler, L.; Balázs, A.; Batár, P.; et al. Curcumin and its analogue induce apoptosis in leukemia cells and have additive effects with bortezomib in cellular and xenograft models. Biomed. Res. Int. 2015, 2015, 968981. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molnár, J.; Szebeni, G.J.; Csupor-Löffler, B.; Hajdú, Z.; Szekeres, T.; Saiko, P.; Ocsovszki, I.; Puskás, L.G.; Hohmann, J.; Zupkó, I. Investigation of the Antiproliferative Properties of Natural Sesquiterpenes from Artemisia asiatica and Onopordum acanthium on HL-60 Cells in Vitro. Int. J. Mol. Sci. 2016, 17, 83. https://doi.org/10.3390/ijms17020083
Molnár J, Szebeni GJ, Csupor-Löffler B, Hajdú Z, Szekeres T, Saiko P, Ocsovszki I, Puskás LG, Hohmann J, Zupkó I. Investigation of the Antiproliferative Properties of Natural Sesquiterpenes from Artemisia asiatica and Onopordum acanthium on HL-60 Cells in Vitro. International Journal of Molecular Sciences. 2016; 17(2):83. https://doi.org/10.3390/ijms17020083
Chicago/Turabian StyleMolnár, Judit, Gábor J. Szebeni, Boglárka Csupor-Löffler, Zsuzsanna Hajdú, Thomas Szekeres, Philipp Saiko, Imre Ocsovszki, László G. Puskás, Judit Hohmann, and István Zupkó. 2016. "Investigation of the Antiproliferative Properties of Natural Sesquiterpenes from Artemisia asiatica and Onopordum acanthium on HL-60 Cells in Vitro" International Journal of Molecular Sciences 17, no. 2: 83. https://doi.org/10.3390/ijms17020083
APA StyleMolnár, J., Szebeni, G. J., Csupor-Löffler, B., Hajdú, Z., Szekeres, T., Saiko, P., Ocsovszki, I., Puskás, L. G., Hohmann, J., & Zupkó, I. (2016). Investigation of the Antiproliferative Properties of Natural Sesquiterpenes from Artemisia asiatica and Onopordum acanthium on HL-60 Cells in Vitro. International Journal of Molecular Sciences, 17(2), 83. https://doi.org/10.3390/ijms17020083